REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHESCIENTIFIQUE UNIVERSITE BATNA 1
INSTITUT DES SCIENCES VETERINAIRES ET DES SCIENCES AGRONOMIQUES
MEMOIRE
Pour l’obtention du diplôme de
Filièr
e
Sciences vétérinaires
Option
Histologie et histopathologie des animaux domestiques
Présenté par :
BOUSSOUAR Hadjira
THEME
JURY
Grade et université
Président : HELEILI Nouzha M.C.A - Université Batna 1 Examinateur : BENHAMZA Louiza Prof - Université Constantine 1 Examinateur : KHENENOU Tarek M.C.A- Université Souk Ahras Rapporteur : BENNOUNE Omar Professeur - Université Batna 1
MAGISTER
EFFET DE L’EXPOSITION A L’ETHANOL SUR
L’OSTEOGENESE ET LA RETINE DES
REMERCIEMENTS
Tout d’abord je remercie le DIEU ‹‹ ALLAH ›› tout puissant pour la volonté, la santé et la patience qu'il nous a donné pour mener à bien ce mémoire.
Je tiens à exprimer mes plus vifs remerciements à mon directeur de mémoire, le Professeur BENNOUNE Omar pour son encadrement, ses encouragements et de nous avoir soigneusement guidé et dirigé tout au long de la réalisation de ce modeste
travail.
Un remerciement particulier aux membres de jury, le Dr HELEILI Nouzha pour avoir accepté de présider mon jury de mémoire ainsi que le Professeur BENHAMZA
Louiza, et le Dr KHENENOU Tarek.
Je tiens à remercier vivement le Professeur AYACHI Ammar, le professeur ALLOUI
Nadir, le Dr ADILI Nezar et le Dr ZGHINA Djamel. Merci pour votre
encouragement et votre aide.
Je tiens à remercier les docteurs YOUSFI Mohamed et MAHDAOUI
Abderrahmane pour l’aide qu’ils m’ont procuré.
Je tiens à remercier aussi tous mes collègues et confrères au sein de notre département des sciences vétérinaires, université Batna1, qui m’ont aidé dans
l’élaboration de ce travail.
Je tiens enfin, à remercier tous ceux et celles qui se reconnaîtront, pour leur contribution à la réalisation de ce mémoire.
DEDICACES
Je dédie ce modeste travail :
A l’âme de ma mère ) qui m’a donné durant toute sa vie, l’amour, le soutien, et la joie que dieu lui accorde sa grâce et sa miséricorde.
A mon père. A qui ce travail vous est particulièrement dédié. Je ne saurais vous exprimer toute ma reconnaissance pour vos sacrifices, votre grand amour, votre soutien moral que vous n’avez cessé de me prodiguer. Que dieu vous protège et vous
offre une longue vie.
A mon cher fiancé le docteur vétérinaire MOKRANI Anis,
A mes frères et mes sœurs,
A mon cousin SAADI Messaoud,
A toute ma famille : BOUSSOUAR,
A tous mes chers ami(e)s, et mes collègues,
A tous ceux que j’aime,
SOMMAIRE
Table des illustrations
Liste des abréviations
Introduction
01Chapitre I
Etude bibliographique
1. Incubation et développement embryonnaire 03
1.1. Le développement embryonnaire précoce 03
1.1.1. La fécondation 03
1.1.2. La segmentation et la blastulation 03
1.1.3. La gastrulation 05
1.1.3.1. Développement de l’hypoblaste 07
1.1.3.2. Formation de la ligne primitive 08
1.1.3.3. Mise en place de l’endoderme et du mésoderme 08
1.1.4. L’organogenèse 09
1.1.4.1. La neurulation 09
1.1.4.1.1. Mouvements généraux des feuillets 10
1.1.4.2. La somitogénèse 10
1.1.5. Développement des membranes extra-embryonnaires 11
1.2. L’incubation 14
1.2 .1. Paramètres techniques de l’incubation des œufs de poules 14
1.2 .1.1. La température 14
1.2 .1.2. La ventilation 14
1.2.1.3. L’humidité 15
1.2 .2.1. Retournement des œufs 15
1.2 .2.2. Mirages 15
1.2 .3. L’éclosion 16
2. Histologie du tissu osseux et de l’œil 17
2.1. Histologie du tissu osseux 17
2.1.1. Les cellules osseuses 17
2.1.1.1. Les cellules ostéoprogénitrices 17
2.1.1.2. Les ostéoblastes 18
2.1.1.3. Les ostéocytes 19
2.1.1.4. Les ostéoclastes 19
2.1.2. Classification du tissu osseux 21
2.1.2.1. Le tissu osseux non lamellaire (le tissu osseux primaire) 21
2.1.2.2. Le tissu osseux lamellaire (le tissu osseux secondaire) 21
2.1.2.2.1. Le tissu osseux lamellaire haversien (l’os compact) 22
2.1.2.2.1.1. Systèmes fondamentaux 23
2.1.2.2.1.2. Systèmes de Havers 23
2.1.2.2.2. Le tissu osseux lamellaire non haversien (l’os trabéculaire) 24
2.1.3. Le périoste et l’endoste 25
2.1.4. L’ostéogénèse 27
2.1.4.1. L'ossification intramembraneuse 27
2.1.4.2. L’ossification endochondrale 29
2.2. Histologie de l’œil 32
2.2.1. Les tuniques de l’œil 33
2.2.1.1.2. La sclère (sclérotique) 35
2.2.1.2. La tunique vasculaire de l’œil 36
2.2.1.2.1. La choroïde 37
2.2.1.2.2. L’iris 37
2.2.1.2.3. Le corps ciliaire 37
2.2.1.3. La tunique nerveuse de l’œil (la rétine) 38
2.2.1.3.1. Structure générale 38
2.2.1.3.2. Couches de la rétine 38
Chapitre II
Matériel et méthodes
1. Etude de l’effet de l’éthanol sur l’ostéogénèse 44
1.1. Matériels 44
1.2. Méthodes d’analyse 45
1.2.1. Préparation et stérilisation des solutions pour l'injection 45
1.2.2. Nettoyage et désinfection de l’incubateur 45
1.2.3. Les œufs et la répartition des groupes 47
1.2.4. Injection des substances 47
1.2.5. Visualisation du squelette 49
1.2.5. 1. Procédure de la coloration 49
1.2.6. Traitement histologique et observation 50
1.2.6.1. Réalisation des coupes histologiques 50
1.2.6.1. 1. Fixation 50
1.2.6.1. 2. Lavage des échantillons 50
1.2.6.1. 3. Inclusion à la paraffine 50
1.2.6.1. 3.1. La déshydratation 51
1.2.6.1. 3.2. L’éclaircissement 51
1.2.6.1. 3.3. L’imprégnation en paraffine 51
1.2.6.1. 4. Coupes au microtome et étalement 53
1.2.6.1. 5. Coloration des lames 55
1.2.6.1. 5.1. Principe de la coloration H&E 55
1.2.6.1. 5.2. Méthode de la coloration des lames 55
1.2.6.1. 6. Le montage des lames 56
1.2.6.2. L’observation microscopique des lames 56
2. Etude de l’effet de l’éthanol sur la rétine 57
2. 1. Matériels 57
2. 2. Préparation et stérilisation des solutions pour l'injection 57
2. 3. Groupes exposés à l’éthanol 57
2. 4. Mésure de la longueur totale et du poids corporel des poussins 58
2. 5. Examen histologique 59
3. Analyses statistiques des données expérimentales 60
Chapitre III
Résultats et discussion
1. Effet de l’éthanol sur l’ostéogénèse 61
1.1. Effets de l'éthanol sur la viabilité des embryons 61
1.2. La coloration macroscopique intégrale avec le rouge d’alizarine 63
1.3. Effet de l’éthanol sur la longueur de la zone proliférative et d’hypertrophie cartilagineuse de l’humérus
64
2. Effet de l’éthanol sur la rétine 69
2.1. Effets de l’éthanol sur la viabilité des poussins 69
2.2. Effets de l’éthanol sur le poids corporel total et la longueur totale du poussin 70
2.3. Effet de l’éthanol sur la structure microscopique de la rétine 74
Conclusion
81Références bibliographiques
Résumé
TABLE DES ILLUSTRATIONS
1. Liste des figures :
Étude bibliographique
Figure 1 : Clivage méroblastique discoïde chez un embryon de poulet 04
Figure 2 : Gastrulation des oiseaux. 06
Figure 3 : Neurulation chez l’embryon d’oiseau. 09
Figure 4 : Une paire de somites. 10
Figure 5 : Représentation schématique des membranes et du fluide extra-embryonnaires.
12
Figure 6 : A : L’ossification intra membranaire (× 540).
B : Prévalence de la lacune des ostéocytes en forme de fuseau dans la section histologique transversale du plateau du sternum de poulet (H&E).
18
Figure 7 : Os compact (Humérus décalcifié du poulet). 22
Figure 8 : Os compact (section longitudinale). 23
Figure 9 : Structure de l’os compact (x 400). 24
Figure 10 : Os trabéculaire (H&E, x 50). 24
Figure 11 : Quelques caractéristiques de la structure osseuse dans une coupe longitudinale à travers une extrémité d'un os long.
26
Figure 12 : Dynamique des sutures crâniennes. 28
Figure 13 : Ostéogénèse membranaire-os pariétal (H&E, x 90). 29
Figure 14 : Développement de l'os endochondral. 30
Figure 16 : Coupe méridienne semi-schématique de l’œil. 32
Figure 17 : La cornée. 34
Figure 18 : Sclère de poulet (x 62,5). 35
Figure 19 : Schéma simplifié de l’architecture de la rétine en trois couches cellulaires.
39
Figure 20 : La rétine avec ses dix couches décrites (× 270). 41
Matériel et méthodes
Figure 21 : Incubateur automatique (vue externe). 46
Figure 22 : Incubateur automatique (vue interne). 46
Figure 23 : Les différents stades de l’injection in ovo. 48
Figure 24 : Automate à inclusion. 51
Figure 25 : Etapes de la mise en blocs. 52
Figure 26 : Cassettes contenant du tissu paraffiné. 53
Figure 27 : Confection et étalement des coupes. 54
Figure 28 : La coloration classique des lames (H&E). 56 Figure 29 : Evaluation du développement par la mesure de la longueur des
poussins.
58
Figure 30 : Evaluation du développement par la mesure du poids corporel des poussins.
59
Résultats et discussion
Figure 31 : Effet de l'éthanol sur le taux de survie des embryons de poulet. 63 Figure 32 : Embryon de poulet à 10 jours coloré avec le rouge d’alizarine. 64 Figure 33 : Coupe transversale au niveau de l’humérus. 65 Figure 34 : Effet de l’éthanol sur la longueur de la zone proliférative. 66 Figure 35 : Effet de l’éthanol sur la longueur de la zone hypertrophique. 67
Figure 36 : Effet de l'éthanol sur la viabilité des poussins. 69 Figure 37 : Effet de l’éthanol sur le poids corporel des poussins. 73 Figure 38 : Effet de l'éthanol sur la longueur des poussins. 74 Figure 39 : Coupe transversale au niveau de la rétine du groupe témoin négatif
(H&E, x100).
75
Figure 40 : Coupe transversale au niveau de la rétine du groupe témoin positif (H&E,x 100).
75
Figure 41 : Coupe transversale au niveau de la rétine du groupe exposé à l’éthanol 10% (H&E, x 100).
76
Figure 42 : Effet de l’exposition à l’éthanol 10% sur la rétine. 77 Figure 43 : Coupe transversale au niveau de la rétine du groupe exposé à
l’éthanol 30% (H&E, x 100).
77
Figure 44 : Effet de l’exposition à l’éthanol 30% sur la rétine. 78 Figure 45 : Coupe transversale au niveau de la rétine du groupe exposé à
l’éthanol 50% (H&E, x 100).
79
2. Liste des tableaux :
Étude bibliographique
Tableau 1 : Caractéristiques des ostéoblastes, des ostéocytes et des ostéoclastes. 20 Tableau 2 : Classification osseuse basée sur l'observation microscopique. 21 Tableau 3 : Classification osseuse basée sur l'apparence brute. 25
Tableau 4 : Les couches de la cornée. 34
Tableau 5 : Les couches de la sclérotique. 36
Tableau 6 : Comparaison entre les bâtonnets et les cônes. 40
Tableau 7 : Les couches de la rétine. 42
Matériel et méthodes
Tableau 8 : Répartition des groupes et traitements 47
Tableau 9 : Les étapes de l’inclusion à la paraffine. 52 Tableau 10 : Le protocole de la coloration H&E. 55
Tableau 11 : Traitement des groupes. 57
Résultats et discussion
Tableau 12 : La viabilité embryonnaire. 61
Tableau 13 : Les moyennes du taux de survie des embryons. 62
Tableau 14 : La viabilité des poussins. 69
Tableau 15: Le poids corporel et la longueur des poussins du groupe témoin (A). 70 Tableau 16 : Le poids corporel et la longueur des poussins du groupe exposé à 70
Tableau 17 : Le poids corporel et la longueur des poussins du groupe exposé à 10% EtOH (C).
71
Tableau 18 : Le poids corporel et la longueur des poussins du groupe exposé à 30% EtOH (D).
71
Tableau 19 : Le poids corporel et la longueur des poussins du groupe exposé à 50% EtOH (E).
71
Tableau 20 : Les moyennes du poids corporel et de la longueur des poussins. 72
LISTE DES SYMBOLES & ABREVIATIONS
% : pour cent. l : microlitre. m : micromètre. °C : degré Celsius.
CG : couche de cellules ganglionnaires. cm : centimètre.
CO : dioxide de carbone. EtOH : éthanol.
g : gramme. h : heure.
H&E : coloration hématoxyline éosine. HR : humidité relative.
Kg : kilogramme. Kcal : kilocalorie.
KOH : hydroxyde de potassium. MEC : matrice extracellulaire. mg : milligramme.
min : minute.
O : oxygène.
PI : couche plexiforme interne. sec : seconde.
INTRODUCTION
L'alcool est le tératogène le plus commun dans le monde occidental, et sa consommation pendant la grossesse peut donner lieu à une affection appelée syndrome d’alcoolisme fœtal ou, sous sa forme plus douce, les effets de l'alcoolisme fœtal. Cela est particulièrement inquiétant, compte tenu de l'augmentation mondiale de l'alcoolisme chez les femmes (Chaudhuri, 2004). Bien que le taux de consommation d'alcool soit demeuré relativement constant au cours des 30 dernières années, le pourcentage de femmes enceintes qui ont consommé de l'alcool a augmenté de 12,4% en 1991 à 16,3% en 1995. Entre-temps, le nombre de nouveau-nés souffrant du syndrome d'alcoolisme fœtal (FAS) a été multiplié par quatre entre 1979 et 1993 avec un taux d'incidence de 6,7 pour 10 000 naissances en 1993 (Miller et al. ,2001).
Les conséquences cliniques négatives de la consommation d'alcool pendant la grossesse peuvent inclure une mortalité néonatale accrue, des anomalies de la tête et du visage, une microcéphalie, des troubles des organes corporels (Bupp Becker et Shibley, 1998). Y compris le retard de croissance, les anomalies musculaires et squelettiques et les déficiences intellectuelles et comportementales (McGill et al., 2009).
Les anomalies les plus fréquentes qui sont observées chez les enfants atteints du syndrome d'alcoolisme fœtal sont la dégénérescence de la rétine et l'hypoplasie du nerf optique. L'alcool altère le développement de la rétine provoquant une migration cellulaire retardée et défectueuse, une perte de neurones dans les couches internes de la rétine, en particulier dans la couche des cellules ganglionnaires. De plus, la réduction du nombre de myéline-axones de la couche de fibres nerveuses optiques est très sensible(Aguileraa et al., 2005).
Divers modèles des animaux ont été utilisés par les chercheurs pour étudier les facteurs et les effets de diverses maladies liées à l'alcool. De tels modèles ont aidé les chercheurs à explorer les mécanismes par lesquels la consommation à court terme et à long terme peut interférer avec les processus normaux de développement de l'embryon(Kamran et al., 2011).
L'embryon aviaire a prouvé son utilité pour étudier les effets néfastes de l'éthanol sur l'embryon. Le poulet et la caille sont des modèles établis de longue date pour la recherche en biologie du développement (Smith, 2008).
L'embryon de poulet (Gallus gallus) est un modèle extrêmement utile pour la biologie du développement, l'embryologie expérimentale et la tératologie. Des descriptions minutieuses et complètes du développement des poussins sont largement disponibles(Drake et al., 2006).
Les modèles aviaires offrent plusieurs avantages pour la recherche développementale. Les œufs fertiles sont peu coûteux, disponibles dans la commerce et ne nécessitent qu'un incubateur pour se développer. La coquille est facilement fenêtrée pour visualiser directement ou manipuler l'embryon et facilement oblitérée pour continuer le développement (Smith, 2008). Les chercheurs peuvent hisser l'œuf, examiner l'embryon et cibler précisément les expositions à des stades de développement spécifiques. Les agents tels que les facteurs de croissance et les tératogènes peuvent être ciblés sur des tissus spécifiques à l'aide d'implants ou de greffons. Les contributions du métabolisme maternel sont absentes d'un système in ovo, qui permet d'isoler directement les effets tératogènes. En bref, le modèle d'embryon de poulet fournit un système pratique et peu coûteux dans lequel appliquer des outils expérimentaux modernes pour déterminer comment un tératogène interfère avec des mécanismes spécifiques qui sous-tendent l'organogenèse et la morphogenèse(Drake et al., 2006).
L'œil aviaire, largement utilisé pour l'étude des fonctions parasympathiques dans le développement et le vieillissement, est devenu un modèle animal privilégié pour l'ophtalmologie expérimentale en raison de sa croissance rapide, de ses qualités visuelles élevées et de sa facilité de manipulation générale(Egle De Stefano et Mugnaini, 1997).
Ce travail a été conçu pour étudier l'effet tératogène de l'exposition in ovo à l'éthanol sur l’ostéogénèse et le développement de la rétine dans un système de modèle d'embryon de poulet; compte tenu de la dose de l’éthanol injecté et le stade de développement embryonnaire.
Chapitre I
Etude bibliographique
1. Incubation et développement embryonnaire :
1.1. Le développement embryonnaire précoce : 1.1.1. La fécondation :
La fécondation se produit lorsque les membranes cellulaires des spermatozoïdes et des ovocytes fusionnent. Après le coït, l'exposition des spermatozoïdes à l'environnement de l'appareil reproducteur féminin entraîne la capacitation, l'élimination des glycoprotéines de surface et du cholestérol de la membrane du sperme, ce qui permet la fécondation (Klein et Enders, 2007).
Plusieurs spermatozoïdes entrent dans l'ovule, mais la fertilisation est assurée par un seul.
Dans le cas du poulet, le premier clivage se produit environ cinq heures après la fécondation (Houillon, 1972).
1.1.2. La segmentation et la blastulation :
La segmentation correspond fondamentalement à une succession de divisions cellulaires donnant des cellules filles ou blastomères, dont le contenu cytoplasmique provient pour une grande part de la ségrégation du cytoplasme ovocytaire(Franquinet et Foucrier, 1998).
Le clivage commence par un zygote, progresse à travers un stade morula et se termine au stade blastocyste (blastula)(McGeady, et al., 2006).
Le blastodisque en segmentation ou blastoderme contient huit blastomères centraux à limites nettes et huit blastomères périphériques dont les limites avec le vitellus sont peu distinctes. Aux stades 32 et 64, les blastomères centraux acquièrent une limite inférieure. Plusieurs assises cellulaires se mettent en place par multiplication des cellules du disque central. Une cavité se creuse entre celles-ci et le vitellus sous-jacent formant le blastocœle primaire(Le Moigne et Foucrier, 1992).
Figure 1: Clivage méroblastique discoïde chez un embryon de poulet(Hyttel et al., 2010). Quatre stades de clivage (A-D) vues du pôle animal, E: un embryon de début de clivage vu de côté.
La zone centrale translucide du blastoderme s'appelle la zone pellucide, tandis que l’anneau périphérique des cellules, qui est toujours en contact avec le jaune d'œuf, s'appelle la zone opaque (figure 1). C'est l'anneau des cellules de la zone opaque qui constitue la structure de l'embryon(Fasenko, 2007).
La zone pellucide se compose d’un ou deux couches cellulaires et la zone opaque se compose de quatre à six couches cellulaires. Lorsque les deux régions distinctes sont formées, le blastodisque devient un blastoderme(Reijrink et al., 2008 ).
1.1.3. La gastrulation :
La division cellulaire se ralentit et finit par s'arrêter, et la gastrulation commence. La gastrulation implique le mouvement d'un grand nombre de cellules pour invaginer la blastula autour du pôle végétatif(Drasdo et Forgacs, 2000).
Dans les embryons aviaires et mammifères, les principaux événements de la gastrulation comprennent la formation et la progression de la ligne primitive, et l'entrée de cellules à travers elle pour former les trois couches(Lawson et al., 2001) (figure 2).
La gastrulation est l'événement morphogénétique à travers lequel les trois couches germinales définitives, l'ectoderme, le mésoderme et l'endoderme sont établis (Wei et Mikawa, 2000).
Figure 2: Gastrulation des oiseaux(Kardong, 2011).
(a) : vue dorsale de la ligne primitive, (b) : une section transversale à travers l'embryon illustre le flux
interne des cellules, (c) : une section médiale longitudinale à travers l'embryon montre la migration
vers l'avant d'un flux distinct de cellules qui produisent la notochorde, (d) : vue tridimensionnelle de
Chez le poulet, le processus de gastrulation implique la formation de la ligne primitive, qui est d'abord visible comme un épaississement de la région épiblastique postérieure due à la migration des cellules épiblastiques de la région latérale vers le centre de l'embryon. Au stade 3, la ligne primitive s’étend de la région postérieure à l’antérieure. À l'extrémité antérieure de la ligne primitive se trouve le nœud de Hensen (Faure et al. ,2002).Ce nœud est responsable d'au moins trois fonctions principales: induction du neuroectoderme (induction neurale), dorsalisation du mésoderme et générer les mouvements d'extension convergents qui conduisent l'allongement du neuraxis(Darnell et al., 1999).
1.1.3.1. Développement de l'hypoblaste:
Les oiseaux et les mammifères gastro-intestinaux forment une ligne primitive au lieu d'un blastopore circulaire, et une couche de tissu sous-jacente évidente, l'hypoblaste (Arendt et Nu¨bler-Jung, 1999).
Chez le poussin, une couche endodermique précoce est présente avant la gastrulation, qui a un destin extra-embryonnaire, mais elle est composée d'au moins deux types cellulaires: l'hypoblaste proprement dit «endophylle ; hypoblaste primaire » et l'endoblaste « endoblaste falciforme ; hypoblaste secondaire »(Foley et al., 2000).
Au moment de la ponte, le blastoderme contient environ 20 000 cellules. La plupart des cellules pellucides de la zone restent à la surface et forment l'épiblaste. Un certain nombre de cellules pellucides se délaminent et migrent dans la cavité subgerminale où elles constituent l'hypoblaste primaire, une collection lâche de groupes de cellules contenant chacune 5-20 cellules. Ensuite, une feuille de cellules provenant d'un épaississement local au bord postérieur du blastoderme (faucille de Koller) migre en avant et pousse les cellules hypoblastiques primaires en face d'eux, formant ainsi l'hypoblaste secondaire, ou endoblaste (Hyttel et al., 2010).
L'hypoblaste contient deux populations cellulaires distinctes: l'une comprend les cellules polyinvaginées qui descendent verticalement de l'épiblaste, l'autre comprend des cellules contribuant à la zone marginale (MZ)(Hefzibah et Oded, 1989).L’épiblaste donne naissance à des structures extra-embryonnaires et à toutes les structures de l'embryon proprement dit, tandis que l'hypoblaste ne donne naissance qu'à des structures extra-embryonnaires(Yang et al., 2002).
1.1.3.2. Formation de la ligne primitive :
La ligne primitive a été définie comme un passage transitoire pour les cellules entre les couches épiblastiques et les couches profondes, en tant que centre blastémique pour la prolifération, ainsi que par les types de cellules prospectives qu'il contient et par l'expression de marqueurs moléculaires(Psychoyos et Stern, 1996).
Au début de l'embryogenèse, la ligne primitive apparaît d'abord comme un épaississement de l'épiblaste dans la zone marginale postérieure, la future extrémité caudale de l’embryon(Sho et al., 2007).
La ligne primitive atteint sa pleine longueur après l'incubation d'un œuf pendant environ dix-neuf heures. À l'extrémité de la ligne prolongée, forme une structure épaissie, où les trois couches de germes sont étroitement associées. Depuis sa découverte par Hensen (1876) dans une étude sur les embryons de lapin, le terme « nœud de Hensen», en abrégé nœud, est largement utilisé(Boettger et al., 2001).
Au cours de la régression de la ligne primitive, une série de cellules filles provenant du nœud sont disposées le long de la ligne médiane, formant des chordamesoderme qui se différencient finalement en notochorde (Mikawa et al., 2004).
La notochorde joue un rôle essentiel dans la structuration des tissus ectodermiques et mésodermiques, tels que le tube neural et les dérivés somitiques(Cleaver et Krieg, 2001). La ligne primitive peut également jouer un rôle dans la spécification du devenir cellulaire, en fonction de la gamme de gènes qu'elle exprime au cours de son développement (Lawson et Schoenwolf, 2003).
1.1.3.3. Mise en place de l’endoderme et du mésoderme :
L’embryon se développe uniquement à partir de l’épiblaste de la blastula. Les tissus de l’aire opaque constituent l’ectoderme extra-embryonnaire dont les cellules du front migrent activement à la surface du jaune, prolifèrent et tendent à l’envelopper par épibolie. Elles sont à l’origine de l’ectoderme des annexes. Ce feuillet est doublé intérieurement par l’expansion du mésoderme extra-embryonnaire(Le Moigne et Foucrier, 1992).
1.1.4. L’organogenèse :
L’organogénèse débute par la neurulation, peu après le stade de la ligne primitive maximale. La plaque neurale se forme d’abord dans la région antérieure, et en avant du nœud de Hensen. C’est l’ébauche de la vésicule cérébrale dont l’apex se fermera tardivement (neuropore antérieur)(De Vos et Van Gansen, 1980).
1.1.4.1. La neurulation :
Le processus embryonnaire, suivant ou se chevauchant la gastrulation, en commençant par l'apparition de la plaque neurale et se terminant par l'achèvement du tube neural(Dye, 2012).
1.1.4.1.1. Mouvements généraux des feuillets :
Les plis neuraux apparaissent après 20-21 heures d’incubation, de part et d’autre du neurectoderme, délimitant la plaque neurale et se rencontrent dans l’axe médian au niveau du cerveau moyen après 26 heures. La fermeture progresse vers l’avant, isolant le cerveau antérieur vers 30-33 heures. La fermeture du tube neural s’achève au niveau caudal après 44 heures d’incubation(Le Moigne et Foucrier, 1992) (figure 3).
A 48 h, alors que la partie antérieure se régionalise avec notamment la différenciation de la zone pharyngienne, la partie troncale reste encore inachevée avec un intestin moyen ne présentant pas de paroi ventrale et donc restant ouvert sur la masse vitelline sous-jacente. Dans la partie caudale se manifeste un début de repli de l’endoderme suite un décollement de l’embryon par rapport au jaune. Ainsi se met en place, à l’instar du processus observé dans la région antérieure, l’intestin postérieur à partir duquel apparait, aux 60 heures, le diverticule allantoïdien(Franquinet et Foucrier, 1998).
1.1.4.2. La somitogénèse :
Dans l'embryon de vertébré, la symétrie bilatérale est particulièrement évidente au niveau de l'arrangement des somites, qui forment deux colonnes symétriques de segments mésodermiques des deux côtés de l'axe embryonnaire(Vermot et Pourquie, 2005).
Dans les embryons aviaires, les somites constituent l'unité morphologique du motif métamérique. Les somites sont des épithéliums formés à partir d'un mésenchyme, la plaque segmentaire, et sont ensuite réorganisés en dermatome, myotome et sclérotome. Quelques heures après leur apparition, les somites subissent d'autres transformations qui conduisent à la production d'un épithélium à deux couches (figure 4), le dermamyotome, et d'un mésenchyme, le sclérotome; le premier se différencie en derme et en muscles squelettiques, tandis que le second donne naissance au cartilage des vertèbres (Duband, et al., 1987).
1.1.5. Développement des membranes extra-embryonnaires :
Chez les oiseaux et généralement chez les reptiles, les membranes extra-embryonnaires se forment peu après l'établissement des couches germinales de base(Kardong, 2011).
Le sac vitellin, composé d'une couche endodermique et d'une couche mésodermique, est une extension de l'intestin de l'embryon, auquel il est annexé(Simonsen, 1996).
Le sac vitellin est important dans le développement précoce : - Source des cellules germinales primordiales.
- Rôle dans les fonctions hématopoïétiques.
- Impliqué dans la formation du tube digestif (Papka, 1995).
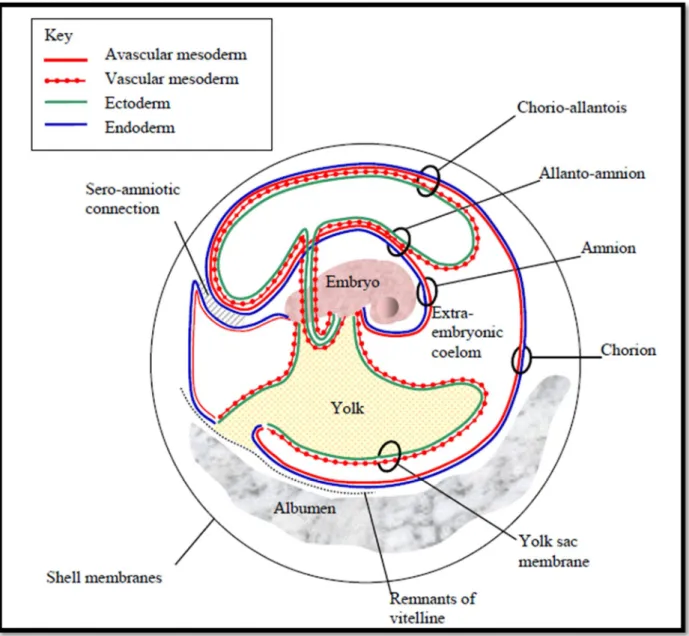
La feuille somatopleurale se développe vers le haut au-dessus de l'embryon en tant que plis amniotique, qui se rencontrent finalement et fusionnent à la ligne médiane. Deux membranes sont produites à partir des plis amniotiques. L'un est l'amnios qui entoure immédiatement l’embryon. L'autre est le chorion le plus périphérique(Kardong, 2011) (figure 5). Le tissu à l'extrémité postérieure de la strie primitive se renfle dans la cavité pro-amniotique et forme le pli amniotique postérieur. Il est continu avec les plus petits plis amniotiques latéraux qui s'unissent pour former le pli amniotique antérieur(Theiler et al., 1972).
Figure 5: Représentation schématique des membranes et du fluide extra-embryonnaires (Baggott, 2009).
L'intérêt particulier pour l'amnios dans l'embryogenèse est dû à son rôle spécifique d'entourer directement l'embryon formant une cavité amniotique remplie de fluide. La membrane amniotique se compose de deux couches cellulaires: la monocouche épithéliale interne et la couche mésenchymateuse externe avec des cellules musculaires lisses (Nechaeva et al., 2004). Le chorion apparaît pendant la gastrulation par déplacement de l'ectoderme extra-embryonnaire sur le toit de la cavité exocoélomique nouvellement formée. C'est une tissue bicouche de double origine, composée de cellules extraembryonnaires. L'ectoderme dérivé du trophectoderme et du mésoderme extra-embryonnaire dérivé de l'épiblaste proximal (Zeigler et al., 2006).
Lorsque l'intestin s'est fermé, le bourgeon allantoïdien s'est formé comme une petite vésicule tapissée par le mésoderme à l'extérieur et l'endoderme à l’intérieur (Caprioli et al., 2001). En grandissant, elle envahit le cœlome extraembryonnaire coiffe l’amnios et s’étale sous la séreuse jusqu’à atteindre le blanc qu’elle va digérer (De Vos et Van Gansen, 1980). Finalement, l'allantoïde externe et le chorion fusionnent pour former une seule membrane composite, la membrane chorio-allantoïque(Kardong, 2011).
L'allantoïde est une structure mésodermique extra-embryonnaire qui forme une connexion entre l'embryon postérieur et le chorion au début du développement (Naiche et Papaioannou, 2003).
1.2. L’incubation :
1.2 .1. Paramètres techniques de l’incubation des œufs de poules :
Pour se développer et éclore avec succès, les embryons aviaires doivent être exposés aux conditions physiques correctes pendant l’incubation (Reid et al., 2000). Deux des conditions environnementales les plus importantes pendant l'incubation, connues pour influer sur le développement embryonnaire, sont la température et l'oxygène( Molenaar et al., 2010).
1.2 .1.1. La température :
Les embryons d'oiseaux sont effectivement ectothermes et dépendent de la chaleur fournie par les parents assis sur les œufs pour atteindre des températures appropriées pour le développement (Martin et al., 2007). Cependant, la température a été suggérée comme le facteur le plus important pour contrôler la croissance et le développement des embryons(Leksrisompong et al., 2007; Lourens et al. , 2005), car une température élevée constante accélère initialement la croissance embryonnaire et l'utilisation des nutriments et de l'énergie du jaune et de l'albumen, mais diminue la croissance embryonnaire plus tard(Lourens et al. , 2007 ).
La température d'incubation optimale pour les espèces de volaille est entre 37 à 38° C, bien que l'éclosabilité soit possible entre 35 et 40,5° C (French, 1997). À partir du 10 ème jour, tout dérèglement de la température, quel qu’en soit le sens, réduit les performances d’éclosion (Hulet et al., 2007). Lorsque les œufs sont transférés en éclosoir, ils subissent un refroidissement qu’il faut d’abord compenser ; ensuite, compte-tenu du fort dégagement calorique (11 Kcal/œuf du 19ème au 21ème jour inclus) et des dangers que présentent des températures trop élevées, l’éclosoir doit être refroidi en permanence pour y conserver une température de 37,5°C (Sauveur, 1988).
1.2 .1.2. La ventilation :
La ventilation correcte de l'incubateur est aussi importante que sa température et son humidité. En ce qui concerne la teneur en oxygène de l'air, il peut y avoir une concentration optimale pour une capacité d'écoulement maximale à 21%. En ce qui concerne le dioxyde de carbone, la figure n'est toujours pas claire. On sait cependant qu'un niveau de dioxyde de carbone supérieur à 1% entraîne une diminution de l'éclosion et du taux de croissance, et aussi que la sensibilité de l'embryon au dioxyde de carbone diminue avec l’âge (Freeman et Vince, 1974).
1.2.1.3. L’humidité :
La porosité de la coquille d'œuf en vapeur d'eau et en gaz vitaux (C0 et 0 ) pendant l'incubation influence l'éclosion des œufs, la croissance embryonnaire et le poids des poussins à l’éclosion(Peebles et al., 1987).L’optimum est une perte d'humidité de l'œuf égale à environ 12 à 14% du poids initial de l'œuf. Comme la perte de poids de l'œuf est entièrement due à la perte d'humidité, cette perte de poids peut être utilisée pour évaluer la perte d’humidité (Brugére, 2015).La gamme optimale d'humidité relative a été rapportée comme étant assez large, entre 40 et 70% HR, avec une éclosabilité maximale obtenue autour de 50% HR(Bruzual et al., 2000).
1.2 .2. Autres paramètres de l’incubation : 1.2 .2.1. Retournement des œufs :
Au cours de l'incubation artificielle, on a signalé que le retournement des œufs réduisait la malposition de l’embryon pour empêcher l'adhérence anormale de l'embryon ou des membranes embryonnaires à la membrane de la coquille et favoriser la fermeture complète et rapide des chorioallantoide à la petite extrémité de l'œuf. Le tournage est nécessaire pour assurer une utilisation correcte de l'albumen par l'embryon en développement dans la période d'incubation normale (Tona et al., 2003). Ces avantages ont été liés à des influences sur la physiologie de l'embryon telles que l'accumulation de protéines dans le liquide amniotique, le taux de croissance de la zone vasculeuse, l'échange gazeux(El-Sagheer, 2012).
Cependant, les œufs qui ne sont pas retournés pendant l'incubation présentent une plus grande fréquence de caries de développement embryonnaire et une trappe réduite par rapport aux œufs tournés(Cutchin et al., 2009).
1.2 .2.2. Mirages :
Compte tenu de la main-d’œuvre nécessaire au mirage et des risques que présente cette manipulation pour le développement embryonnaire (1à 3% de mortalité), les mirages ne sont pratiquement plus jamais effectués au 5ème jour d’incubation comme il était jadis d’usage pour détecter les œufs clairs et les embryons morts précocement (germes morts). Le mirage au 18ème jour a pour but essentiel d’éviter un encombrement excessif des éclosoirs et de contrôler la bonne marche de l’incubation ; à côté de ces avantages, il est lui aussi couteux en main-d’œuvre et est donc quelquefois supprimé, peut-être à tort, dans certains grands couvoirs.
Lorsqu’il est pratiqué, il doit l’être sans heurts et rapidement pour éviter le refroidissement des œufs(Sauveur, 1988).
1.2 .3. L’éclosion :
Les facteurs d'amélioration qui affectent l’éclosion comprennent la souche, la santé, la nutrition et l'âge du troupeau, la taille et le poids des œufs, le poids et la qualité, la durée et les conditions de stockage des œufs, l'hygiène des œufs et la saison de l'année. L'âge des obtenteurs affecte l'éclosion, parce qu’elle est liée à la qualité de l'œuf à couver telle que la composition ou le rapport interne de l'œuf, le poids de l'œuf et la qualité de la coquille, ce qui influence l'état d'incubation et le développement de l'embryon de poulet. L'éclosion est influencée par le stockage des œufs, car la qualité de l'œuf se déprécie, ce qui affecte l'activité métabolique de l'embryon de poulet, ce qui influe sur le développement embryonnaire du poussin (Yassin et al., 2008).
2. Histologie du tissu osseux et de l’œil :
2.1. Histologie du tissu osseux:
L'os forme une proportion majeure des tissus conjonctifs du corps, et fournit un exemple de la combinaison de deux composants pour donner un tissu avec des propriétés mécaniques particulières(Holmes, 1965).
Le composant structurel de la phase organique est le collagène de type I (fibreux), qui comprend environ 90% de la protéine osseuse; la phase inorganique est principalement de minuscules cristaux de l'hydroxyapatite minérale alcaline, Ca (Po ) (OH) (Tim, 2003).
Il est le support mécanique essentiel du squelette, permet la locomotion, transmet les forces issues de la contraction musculaire d’une partie du corps à une autre pendant le mouvement et assure la protection des organes internes. Enfin, il joue un rôle extrêmement important dans le maintien de l’homéostasie car il est un réservoir métabolique de sels minéraux, en particulier de calcium(Toppets et al., 2004) .
2.1.1. Les cellules osseuses:
L'os est composé de quatre types de cellules. Les ostéoblastes, les ostéoclastes et les cellules de la muqueuse osseuse sont présentes sur les surfaces osseuses, tandis que les ostéocytes pénètrent dans l'intérieur minéralisé(Bilezikian et al., 2002) (tableau 1).
Les cellules osseuses sont particulièrement reconnaissables dans le tissu osseux fibreux réticulé, pendant la croissance intensive propre à la période embryonnaire les cellules ostéoprogénitrices, les ostéoblastes et les ostéocytes représentent différents stades fonctionnels d’un même type cellulaire(Dhem, 2004).
2.1.1.1. Les cellules ostéoprogénitrices :
Les cellules ostéoprogénitrices sont de petites cellules fusiformes résidant sur toutes les surfaces osseuses non résorptives. Ils constituent la couche profonde du périoste qui investit chaque os et aussi l'endoste qui tapisse la cavité médullaire, les canaux haversiens et d'autres espaces de tissus mous (Cormack, 2001). Les cellules ostéoprogénitrices sont des cellules semblables à des fibroblastes dans la couche interne du périoste adjacente à la surface de l'os: elles fonctionnent comme des cellules souches. Dans les os en croissance active ou après une fracture, elles subissent une mitose. Certaines cellules de la progéniture se différencient en ostéoblastes tandis que d'autres restent en tant que cellules ostéoprogénitrices (Mitchell et Peel, 2009).
2.1.1.2. Les ostéoblastes :
Les ostéoblastes sont la forme immature des cellules osseuses ; ils sont responsables de la synthèse et de la sécrétion de l’ostéoïde, composant organique de la matrice extracellulaire de l’os (figure 6-A). Les ostéoblastes prisonniers dans la substance osseuse, deviennent des ostéocytes(Wheater et al., 1979).
La fonction principale de l’ostéoblaste est de synthétiser et de minéraliser la matrice osseuse au cours de la croissance du squelette, du renouvellement de la matrice osseuse chez l’adulte et de la réparation osseuse tout au long de la vie(Pierre, 2001).
A la fin de la phase de formation osseuse, les ostéoblastes peuvent être insérés dans l'os sous forme d'ostéocytes, devenir des ostéocytes inactifs, ou subir une mort cellulaire programmée (apoptose). Sous certaines conditions, ils peuvent se différencier en cellules productrices d'os chondroïde(Dallas et Bonewald, 2010 ; Franz-Odendaal et al., 2006).
Figure 6 : A : L’ossification intra membranaire (× 540)(Gartner et Hiatt, 2007). B : Prévalence de la lacune des ostéocytes en forme de fuseau dans la section histologique transversale du plateau du sternum de poulet (H&E)(Domenis
et al., 2009).
Ob : ostéoblastes, Oc : ostéoclastes.
2.1.1.3. Les ostéocytes :
Les ostéocytes sont les cellules osseuses prédominantes et il est généralement admis que les ostéocytes sont les mécanosensors professionnels de l’os (Djien et al. ,2007). L’ostéocyte c’est une cellule fusiforme contenue dans une logette appelée ostéoplaste. Il présente de nombreux prolongements cellulaires contenus dans des canalicules interostéoplastiques (figure 6-B). Les prolongements de deux ostéocytes sont réunis par des jonctions de type Gap. Contrairement aux ostéoblastes qui se divisent activement, les ostéocytes n’ont plus la capacité de se diviser, d’autant que la rigidité de la MEC osseuse s’y oppose(Prugnolle et Thoreau, 1996).
Les ostéocytes, cellules matures de l'os, possèdent un rapport noyau-cytoplasme élevé et relativement peu d'organites cytoplasmiques (Ovalle et Nahirney, 2013). Après leur formation, les ostéocytes deviennent de taille réduite. L'espace dans la matrice occupée par un ostéocyte est appelé la lacune ostéocytaire(Antonio, 2012).
2.1.1.4. Les ostéoclastes :
Ce sont des cellules d’origine hématopoïétique provenant de cellules souches communes à la lignée monocytes-macrophages(Sybille, 2012).
Après avoir atteint la surface osseuse, les précurseurs ostéoclastiques y adhèrent et fusionnent de manière asynchrone, entraînant la formation d’une cellule multinucléée, l’ostéoclaste mûr (figure 6-A). Lorsqu’il résorbe activement l’os, l’ostéoclaste est caractérisé par une bipolarité morphologique et fonctionnelle(Baron, 2001).
Les ostéoclastes s’attachent à la surface osseuse au niveau du site de résorption osseuse active, souvent au sein de dépressions où ils ont érodé l’os. Ces dépressions sont dites niches de résorption ou lacunes de Howship (Stevens et Lowe, 1993).
Tableau 1: Caractéristiques des ostéoblastes, des ostéocytes et des ostéoclastes (Michael et Wojciech, 2015).
Caractéristiques Osteoblaste Osteocyte Osteoclaste Emplacement Surface osseuse ; cône
de fermeture des canaux de résorption
Lacunes et canalicules de la matrice osseuse
Surface osseuse ; cône de coupe des canaux de résorption
Pourcentage de toutes les cellules de l'os
>5% 95% >1%
Fonction Dépôts de la matrice osseuse; initie la minéralisation en libérant des vésicules matricielles Maintient la matrice osseuse ressent le stress mécanique; régule l'hémostase du calcium et du phosphate
Résorbe l'os par hydrolyse enzymatique de la matrice osseuse minéralisée Morphologie cellulaire Cuboïde ou polygonale, cellule mononucléaire, cytoplasme basophile; Golgi négatif Petite cellule mononucléaire ovale; cytoplasme pâle; processus de cellule longue Grande cellule multinucléaire; cytoplasme acidophile; bordure ébouriffée; la lacune sous-jacente de Howship
Cellules précurseurs Cellule
ostéoprogénitrice Osteoblast Cellules hématopoïétiques (GMP, CFU-GM) Processus de différenciation / facteurs de transcription CBFA1 (RUNX2); IGF-1 Le processus de sélection des
ostéoblastes n'est pas connu c-fos; NF-B; RANK-RANKL signalisation Principaux récepteurs hormonaux régulateurs RANKL, récepteurs de la PTH RANKL, récepteurs de la PTH
RANG, les récepteurs de la calcitonine; récepteurs de la phosphatase acide tartrate-résistante (TRAP)
Durée de vie Semaines (12 jours) Années (10-20 ans) Jours (3 jours) Marqueurs biochimiques Ostéocalcine, sialoprotéine osseuse (BSP-2) Dentine-matrice protéine-1 (DMP-1); la podoplanine (protéine E11); la sclérostine; facteur de croissance fibroblastique-23 (FGF-23) La phosphatase acide résistante aux tartrates (TRAP); la cathepsine K; métalloprotéinase matricielle-9(MMP-9)
2.1.2. Classification du tissu osseux :
Au cours du développement et tout au long de la vie se forment plusieurs types de tissu osseux. Il est ainsi possible de faire la distinction entre le tissu osseux non lamellaire et le tissu osseux lamellaire(Bertrand, 2008) (Tableau 2).
2.1.2.1. Le tissu osseux non lamellaire (le tissu osseux primaire):
L’os primaire, immature ou tissé est le premier type d'os formé au cours du développement du fœtus, de la réparation osseuse et du renouvellement tissulaire. Les caractéristiques de l'os primaire sont des ostéocytes abondants, une faible teneur en minéraux et une organisation irrégulière des fibres de collagène. L'os tissé est mécaniquement faible. Il est temporaire et remplacé par un tissu osseux secondaire(Bobrysheva et Kachshenko).
2.1.2.2. Le tissu osseux lamellaire (le tissu osseux secondaire) :
L'os secondaire est un os mûr composé de lamelles osseuses parallèles ou concentriques de 3 à 7 m d'épaisseur (Gartner et Hiatt, 2007). Les ostéocytes dans l'os lamellaire ont tendance à être aplatis avec moins de canalicules, et ils ont une disposition plus organisée(Anusuya, 2011).
Les os lamellaires sont des os à dépôt lent avec des arrangements bien organisés de collagène comme les os de mammifères, les os longs aviaires sont constitués d'os cortical compact, qui est la coquille extérieure entourant l'os spongieux ou trabéculaire et l'espace de la moelle(Rath et al., 2000)(tableau 3).
Tableau 2: Classification osseuse basée sur l'observation microscopique (Cui et al., 2011).
Os primaire (os immature)
Arrangement irrégulier
Lamelles sans motif organisé; pas très minéralisé Développer le fœtus Développement osseux Os secondaire (os mature) Arrangement régulier Motif lamellaire bien organisé; fortement minéralisé Adultes Protection et support
2.1.2.2.1. Le tissu osseux lamellaire haversien (l’os compact) :
L'os compact est organisé en unités, appelées ostéons, entourant un vaisseau sanguin central. Le cylindre de la matrice osseuse autour d'un vaisseau sanguin peut également être appelé système haversien (Rogers, 1983).
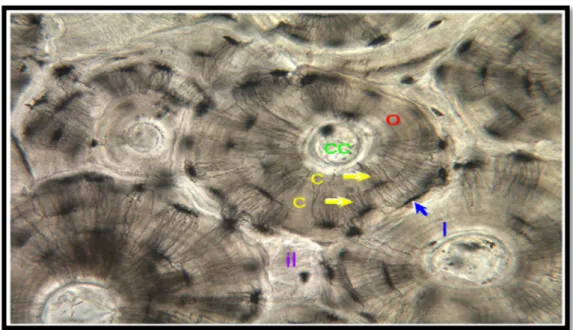
Chaque ostéon est formé par des lamelles concentriques ou des anneaux de matrices qui entourent un canal central. Le système canaliculaire s'étend à travers les lamelles. Les vaisseaux sanguins et les nerfs sont situés dans le canal central. Les canaux centraux d'ostéons osseux compacts se connectent à la surface de l'os par des canaux perforants (canal de Volkmann) qui se déroulent perpendiculairement aux canaux centraux (figure 7). Les lamelles interstitielles complètent l'espace entre les ostéons tandis que les lamelles circonférentielles recouvrent la surface externe et interne de l'os compact(Eurell, 2004).
Figure 7 : Os compact (Humérus décalcifié du poulet)(Bacha et Bacha, 2014). 5:canal Haversien (canal central), 12: ostéocyte, 14 : canal de Volkmann (canal perforant).
2.1.2.2.1.1. Systèmes fondamentaux :
Comporte les lamelles circonférentielles extérieures ; des piles de lamelles s'étendent au moins partiellement autour de la circonférence extérieure d'un os long. Le dépôt de ces lamelles par le périoste entraîne une augmentation de l'épaisseur dans la paroi de la diaphyse et les lamelles circonférentielles intérieures ; des piles de lamelles s'étendent au moins partiellement autour de la circonférence intérieure d'un os long faisant face à la cavité de la moelle (figure 8). Le dépôt de ces lamelles par l'endoste entraîne une augmentation de l'épaisseur de la paroi de la diaphyse(Pakurar et Bigbee, 2004).
Figure 8 : Os compact (section longitudinale)(Eroschenko, 1996).
2.1.2.2.1.2. Systèmes de Havers :
Un ostéon (ou système haversien) se réfère au complexe de lamelles concentriques, typiquement de 100-250 m de diamètre, entourant un petit canal central qui contient des vaisseaux sanguins, des nerfs, du tissu conjonctif lâche et de l'endoste(Mescher, 2016) (Figure 9). Les ostéons sont séparés les uns des autres, et notamment des lamelles interstitielles, par une ligne de ciment particulièrement distincte(Krstic, 1985).
Figure 9 : Structure de l’os compact (x 400)(Eurell et Frappier, 2006). O : osteon, CC : canal central, L : lacune, C : canaliculi, il : lamelles interstitielles.
2.1.2.2.2. Le tissu osseux lamellaire non haversien (l’os trabéculaire) :
L'os spongieux forme la partie la plus interne des axes des os longs, la couche délimitant immédiatement la cavité médullaire; les extrémités des os longs; tout sauf la coquille des os courts(Clarkson).
Figure 10 : Os trabéculaire (H&E, x 50)(Heath et al., 2006 ). MH : espaces médullaires hématopoiétique, T : trabécules.
L'os spongieux est constitué de lamelles trabéculaires. Les lamelles trabéculaires forment les espaces dans l'os spongieux qui sont remplis de la moelle osseuse. Les ostéocytes sont également logés dans les trabécules (figure 10) et sont reliés par de minuscules passages tubulaires ramifiés appelés canalicules. Les canalicules fournissent un canal pour l'échange de métabolites entre les ostéocytes et l'espace extracellulaire (Zoetis et al., 2003).
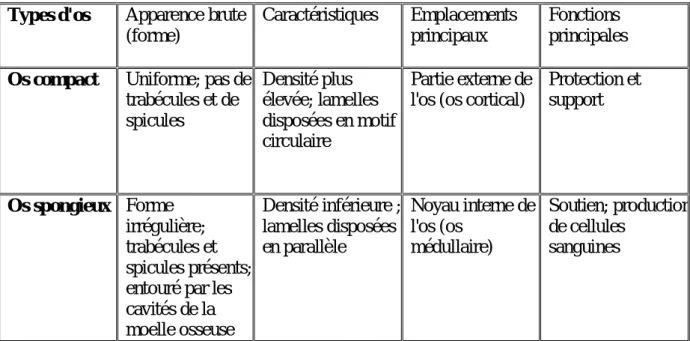
Tableau 3: Classification osseuse basée sur l'apparence brute(Cui et al. 2011). Types d'os Apparence brute
(forme)
Caractéristiques Emplacements principaux
Fonctions principales
Os compact Uniforme; pas de trabécules et de spicules Densité plus élevée; lamelles disposées en motif circulaire Partie externe de l'os (os cortical)
Protection et support Os spongieux Forme irrégulière; trabécules et spicules présents; entouré par les cavités de la moelle osseuse Densité inférieure ; lamelles disposées en parallèle Noyau interne de l'os (os médullaire) Soutien; production de cellules sanguines 2.1.3. Le périoste et l’endoste :
Les os sont entourés, extérieurement, par un tissu conjonctif particulier, le périoste, qui a des potentialités ostéogènes et qui est donc également concerné par le dépôt de tissu osseux nouveau ; son rôle est particulièrement important en cas de fracture(Dhem, 2004).
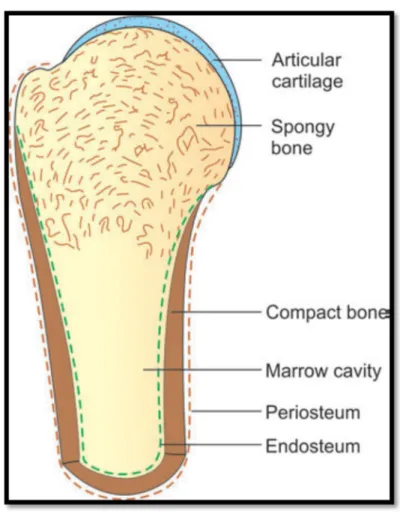
Le périoste est la couverture externe d'un os entier à l'exception des surfaces articulaires (figure 11) (Banks, 1993), des insertions tendineuses et ligamentaires et certains sites comme la zone sous-capsulaire du col du fémur. Il est très vascularisé. Il est constitué de deux couches : le périoste fibreux, externe et le périoste cellulaire, interne (Toppets et al., 2004).
Figure 11: Quelques caractéristiques de la structure osseuse dans une coupe longitudinale à travers une extrémité d'un os long(Neelam et Sabita, 2014).
L'endoste est un tissu conjonctif spécialisé qui unit les cavités de la moelle et fournit des cellules ostéoprogénitrices et des ostéoblastes pour la croissance et la réparation des os (Gartner et Hiatt, 2015). L'endoste est une mince couche d'ostéoblastes actifs et inactifs, qui recouvre toutes les surfaces internes de l'os; les ostéoblastes sont également nécessaires pour la croissance osseuse(Mescher, 2013).
2.1.4. L’ostéogénèse:
La formation du tissu osseux peut se produire par deux processus distincts: l'ossification endochondrale (croissance du cartilage) et l'ossification intramembranaire (croissance de la membrane)(Fratini et al., 2013).
L'ossification osseuse intramembraneuse et endochondrale se produit à proximité de la croissance vasculaire (Kanczler et Oreffo, 2008). La formation osseuse endochondrale se produit après la différenciation des cellules dans les condensations mésenchymateuses en chondrocytes. Ces cellules sécrètent une matrice extracellulaire cartilagineuse riche en protéoglycanes, en glycosaminoglycanes (GAG) et en collagène (types II et X). Les chondrocytes subissent un processus de différenciation stratifiée et d'apoptose, permettant le remplacement du cartilage par l'os. En revanche, au cours de l'ossification intramembranaire, les cellules mésenchymateuses se différencient directement en ostéoblastes osseux (Rigueur et Lyons, 2014).
2.1.4.1. L'ossification intramembraneuse :
Au cours du développement des os intramembranaires, aucun précurseur cartilagineux n'est nécessaire pour la formation osseuse primaire (Crowder et Stout, 2011). La formation osseuse intramembraneuse est obtenue par transformation directe des cellules mésenchymateuses en ostéoblastes, les cellules squelettiques impliquées dans la formation osseuse. C'est le processus responsable du développement des os plats de la voûte crânienne, y compris les lignes de suture crânienne, certains os du visage et des parties de la mandibule et de la clavicule(Bilezikian et al, 2008)(figure 12).
Dans l'ossification intramembraneuse, l'os se développe à partir de mésenchyme du tissu conjonctif. Certaines cellules se différencient directement dont les ostéoblastes et produisent la matrice osseuse (Eroschenko, 2008). Les cellules ostéoprogénitrices se différencient en ostéoblastes. Ces cellules commencent à synthétiser et à sécréter l'ostéoïde. Le premier composant sécrété de l'ostéoïde est le collagène, et les constituants restants de la substance fondamentale sont produits un peu plus tard(Eurell et Frappier, 2006).
Figure 12 : Dynamique des sutures crâniennes(Ducy ,2001).
Les ostéoblastes sécrètent la matrice osseuse qui est ensuite calcifiée, avec comme résultat l’encapsulation des ostéoblastes qui deviennent alors des ostéocytes (Junqueira et al., 2001), et leurs fines expansions cytoplasmiques se rétractent pour former de fins prolongements à l’intérieur des canalicules. Les cellules ostéoprogénitrices situées à la surface des centres d’ossification se divisent par mitose pour donner naissance à des ostéoblastes qui fabriquent davantage d’os. La constitution de l’os se fait progressivement par fusion de centres d’ossification adjacents, aboutissant à la formation d’un os d’aspect grossièrement spongieux (Heath et al., 2006 ) (figure 13).
Figure 13 : Ostéogénèse membranaire-os pariétal (H&E, x 90)(Kuhnel, 2003). 1 : lamelles ostéoïdes avec ostéocytes, 2 : tissu conjonctif embryonnaire (mésenchyme), 3: ostéoclaste.
2.1.4.2. L’ossification endochondrale :
Au cours de la formation de l'os endochondral, les cellules formant le cartilage et l'os expriment des composants matriciels extracellulaires différents et uniques à différents stades. Formation d'une matrice de collagène, dépôt d'ions calcium et apparition séquentielle de protéines osseuses telles que l'ostéocalcine, la phosphatase alcaline, l'ostéopontine et la sialoprotéine osseuse dans la cascade d'ossification endochondrale ont été caractérisées (Chang et al., 2007).
Au cours de la formation osseuse endochondrale normale, la dégradation de la matrice cartilagineuse minéralisée par les ostéoclastes / chondroclastes et le dépôt d'une matrice osseuse caractéristique par les ostéoblastes sont étroitement coordonnés (Nakashima et al., 2002).
La croissance et le développement des os longs se produise à travers le processus d'ossification endochondrale dans lequel un modèle de cartilage est formé et progressivement remplacé par un tissu osseux(Ahmed et al., 2013) (figure 15).
Figure 14 : Développement de l'os endochondral(Long et Ornitz, 2013).
Au début de la formation osseuse endochondrale, les cellules progénitrices mésenchymateuses se condensent et se différencient en chondrocytes pour former des rudiments cartilagineux(Daisuke et al., 2012) (Figure 14 ; A et B).
Les chondrocytes jouent un rôle central dans ce processus, contribuant à la croissance longitudinale grâce à une combinaison de prolifération, de sécrétion de la matrice extracellulaire (MEC) et d’hypertrophie(Mackie et al., 2011) (figure 14 ; C).
La zone des chondrocytes hypertrophiques est envahie par les vaisseaux sanguins, les ostéoblastes et les ostéoclastes pour initier l'ossification de la matrice cartilagineuse (Estrada et al., 2011)(Figure 14 ; D).
Figure15: La formation de l'os endochondral (× 14) (Gartner et Hiatt, 2000; Gartner et Hiatt, 2007). C : chondrocytes, flèches : complexe osseux, b : os, P : périoste.
2.2. Histologie de l’œil :
Le premier signe du développement des yeux est un gonflement sur les côtés latéraux du prosencéphale à peu près au stade neuf de développement. Ce sont les rudiments des vésicules optiques qui se trouvent sous l'ectoderme de la tête. Au stade 10, chaque vésicule a commencé à se contracter à sa base jusqu'à ce que sa connexion au cerveau se réduise à une tige étroite, mais la lumière de la vésicule optique est toujours en communication avec le prosencéphale via la tige(Bellairs et Osmond, 2005).
L'oeil mesure environ 25 mm de diamètre. Il est suspendu dans la cavité orbitaire osseuse par six muscles extrinsèques qui contrôlent son mouvement. Une épaisse couche de tissu adipeux entoure et amortit partiellement l'œil lorsqu'il se déplace dans l'orbite. Les muscles extraoculaires sont coordonnés de sorte que les yeux se déplacent symétriquement autour de leurs propres axes centraux(Michael et Wojciech, 2010).
Les yeux aviaires prennent un volume considérable de la tête de l'oiseau et sont très grands par rapport à la taille du cerveau. En termes généraux, la structure de leurs yeux n'est pas très différente de celle des autres vertébrés(Whittow, 1999).
L'œil a trois couches ou tuniques principales:
- La tunique fibreuse, comprend la sclère et la cornée.
- La tunique vasculaire, une membrane mince, fortement vascularisée, comprend les corps ciliaires et l’iris.
- La tunique nerveuse, est formée par la rétine (Yash, 2013). 2.2.1. Les tuniques de l’œil :
2.2.1.1. La tunique fibreuse de l’œil :
La tunique fibreuse recouvre la plus grande partie du globe oculaire sous la forme d'une couche fibreuse résistante, appelée sclère. La sclère protège les parties les plus faibles de l'œil et offre des points d'insertion aux muscles oculaires, constitués de tissu musculaire squelettique, qui permettent à la boule oculaire de se déplacer à l'intérieur de son orbite. En face, la tunique fibreuse est mince et transparente et prend une courbure beaucoup plus forte. Cette partie est la cornée. Sa surface est tapissée d'un épithélium d'origine ectodermique (Lommel van, 2003).
2.2.1.1.1. La cornée:
Une partie convexe et transparente de la couche externe des yeux appartenant à la média dioptrique de l'œil, avec une épaisseur de 0,8-0,9 mm au centre et de 1,1 mm en périphérie (Krstic, 1984).
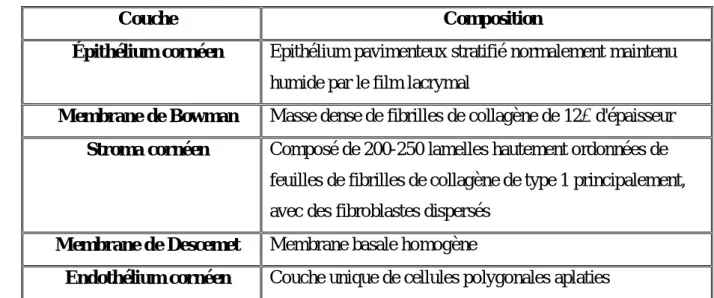
Elle se compose de trois couches cellulaires distinctes à la fois en apparence et en origine. Ces couches sont séparées par deux membranes importantes qui apparaissent homogènes lorsqu'elles sont vues au microscope optique (figure 17). Ainsi, les cinq couches de la cornée vues dans une coupe transversale sont : l’épithélium cornéen, la membmne de Bowman, stroma cornéen, membrane de Descemet (membrane basale postérieure) et l’endothélium cornéen(Michael, 2002) (tableau 4).
Figure 17: La cornée(Mescher, 2011).
1 : épithélium cornéen, 2 : stroma cornéen, 3 : endothélium cornéen.
La cornée aviaire est histologiquement similaire à celle des mammifères, sauf que la membrane de Bowman n'est pas toujours différenciée(Bolton, 1976).
La membrane de Bowman, cependant est plus épaisse et donc plus apparente dans les préparations histologiques. La membrane de dissipation est relativement mince et moins distincte(Bacha et Bacha, 2014).
Tableau 4 : Les couches de la cornée(Vaughan, 2002).
Couche Composition
Épithélium cornéen Epithélium pavimenteux stratifié normalement maintenu humide par le film lacrymal
Membrane de Bowman Masse dense de fibrilles de collagène de 12 d'épaisseur Stroma cornéen Composé de 200-250 lamelles hautement ordonnées de
feuilles de fibrilles de collagène de type 1 principalement, avec des fibroblastes dispersés
Membrane de Descemet Membrane basale homogène
2.2.1.1.2. La sclère (sclérotique) :
La sclère, qui représente environ les cinq sixièmes de la surface de l'œil, commence à la périphérie de la cornée et s'étend vers l'arrière jusqu'au nerf optique(Stacey, 2007).
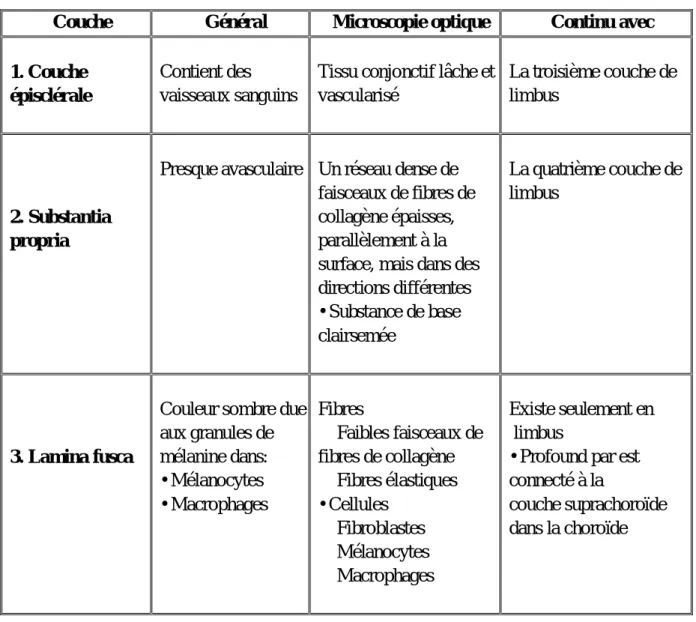
La sclère des poussins se compose à la fois d'une couche cartilagineuse interne et d'une couche fibreuse externe plus mince qui ressemble à la sclère des mammifères (Phillips et al., 2000) (tableau 5).
Figure 18 : Sclère de poulet (x 62,5)(Bacha et Bacha, 2014). 11 : cartilage scléral, 12 : osselet scléral, 13 : artefact spatial.
La sclérotique aviaire a un anneau d'osselets scléraux se chevauchant dans un tissu conjonctif dense(Aughey et Frye, 2001) (figure 18).
Tableau 5: Les couches de la sclérotique(Karnov et al., 2017).
Couche Général Microscopie optique Continu avec
1. Couche épisclérale
Contient des vaisseaux sanguins
Tissu conjonctif lâche et vascularisé
La troisième couche de limbus
2. Substantia propria
Presque avasculaire Un réseau dense de faisceaux de fibres de collagène épaisses, parallèlement à la surface, mais dans des directions différentes • Substance de base clairsemée La quatrième couche de limbus 3. Lamina fusca
Couleur sombre due aux granules de mélanine dans: • Mélanocytes • Macrophages Fibres Faibles faisceaux de fibres de collagène Fibres élastiques • Cellules Fibroblastes Mélanocytes Macrophages Existe seulement en limbus
• Profound par est connecté à la
couche suprachoroïde dans la choroïde
2.2.1.2. La tunique vasculaire de l’œil :
La couche uvéale est une couche hautement vascularisée qui forme de trois composants: la choroïde, le corps ciliaire et l’iris. La choroïde est un tissu conjonctif lâche caractérisé par sa richesse en vaisseaux sanguins et la pigmentation qui absorbe la lumière atteignant la choroïde. Le corps ciliaire contient des fibres musculaires lisses entourant la lentille et contrôle la mise au point de la lentille. L'iris est situé entre les chambres antérieure et postérieure. La pupille centrale de l'iris régule la quantité de lumière entrant dans le globe oculaire (Shu-xin, 1999).
2.2.1.2.1. La choroïde :
La choroïde est une couche relativement mince qui fournit l'apport sanguin à la rétine et elle est située entre la lamina fusca de la sclère et l'épithélium pigmentaire rétinien (Scudamore, 2014).
La couche vasculaire de la choroïde contient des vaisseaux sanguins et de grands espaces incorporés dans un tissu conjonctif lâche. La couche choriocapillaire est séparée de l'épithélium pigmentaire de la rétine par une membrane de Bruch indistincte. Aucun tapetum lucidum n'est présent chez le poulet (Bacha et Bacha, 2014).
La structure lâche de la choroïde joue un rôle majeur dans le maintien de la pression intraoculaire (PIO). Les fluides tissulaires peuvent être filtrés à partir de l'endothélium capillaire ou réabsorbés dans les capillaires eux-mêmes, en fonction des changements du gradient de pression hydrostatique(Egle De Stefano et Mugnaini, 1997).
2.2.1.2.2. L’iris :
L'iris est l'extension la plus antérieure de l'uvée (couche intermédiaire) qui couvre partiellement la lentille, laissant une ouverture ronde au centre appelée pupille. Il est composé de tissu conjonctif lâche contenant un grand nombre de cellules pigmentées et de nombreux vaisseaux sanguins. Sa surface antérieure est rainurée et striée d'une couche de recouvrement discontinue de cellules de Chapeau ressemblant à un épithélium pavimenteux, mais il s'agit probablement de fibroblastes(Bacon et Niles, 1983).
La face antérieure de l'iris, exposée à la chambre antérieure, n'est pas recouverte d'épithélium, mais consiste en une couche irrégulière et discontinue de fibroblastes et de mélanocytes, densément tassée et avec des processus interdigités. Plus profond dans l'iris, le stroma est un tissu conjonctif lâche plus typique avec microvasculature. La surface postérieure de l'iris est lisse, avec un épithélium à deux couches continues avec celui couvrant le corps ciliaire et ses processus(Mescher, 2010).
2.2.1.2.3. Le corps ciliaire :
Le corps ciliaire, situé entre la cornée et la sclérotique, contient une région de muscle lisse (muscle ciliaire), qui change la forme de la lentille (accommodation) de sorte qu'il concentre la lumière sur la rétine (Peckham, 2011). La structure histologique du corps ciliaire est essentiellement un tissu conjonctif lâche (riche en fibres élastiques, en vaisseaux et en mélanocytes) entourant le muscle ciliaire. Les surfaces du corps ciliaire qui font face au corps vitré, à la chambre postérieure et à la lentille sont couvertes par l'extension antérieure de la rétine(Junqueira et Carneiro, 2005).
2.2.1.3. La tunique nerveuse de l’œil (la rétine) : 2.2.1.3.1. Structure générale:
La rétine est une couche mince et délicate composée de deux composants:
• La rétine neurale est une couche interne qui contient des récepteurs sensibles à la lumière et des réseaux neuronaux complexes.
• L'épithélium pigmentaire rétinien est une couche externe composée de cellules cubiques contenant de la mélanine(Michael, 2010).
2.2.1.3.2. Couches de la rétine :
La rétine photosensible du poulet est composée de dix couches, comme chez les mammifères, mais contrairement à celle des mammifères est avasculaire (Aughey et Frye, 2001). Les cellules de l'épithélium pigmenté sont considérablement différentes dans le poulet. Elles sont grandes et étroites plutôt que cuboïdes. Le noyau occupe la plus petite région basale de chaque cellule, qui contient peu ou pas de granules de pigments. La partie apicale est remplie de granules de pigments en forme de bâtonnets orientés parallèlement à l'axe long de la cellule. Le cytoplasme apical semble souvent être séparé en des touffes ou des brins de granules de pigments (Bacha et Bacha, 2014).
La rétine, la troisième et la plus intérieure tunique de l'œil, est sa partie neurale, qui contient les cellules photoréceptrices, connues sous le nom de bâtonnets et de cônes (Gartner et Hiatt, 2007) (figure 19). Chaque bâtonnet ou cône est constitué d'un segment externe qui est relié au segment interne par une tige étroite. Le segment externe contient une pile de disque membraneux.
Figure 19 : Schéma simplifié de l’architecture de la rétine en trois couches cellulaires (Guillaume, 2008).
Les segments extérieurs des bâtonnets sont de forme cylindrique tandis que les segments externes du cône sont coniques (tableau 6). Les segments externes sont des cils hautement modifiés et les cellules en bâtonnets et en cônes sont des cellules neurales modifiées qui convertissent l'énergie photonique en un potentiel générateur qui dépolarise les fibres des cellules nerveuses en contact avec le nerf optique(Feeback, 1987).