A Monsieur le Professeur Pascal CHAVANET et Monsieur le Professeur William COUET qui ont accepté de consacrer de leur temps à la lecture et à l’appréciation de ce manuscrit, qu’ils trouvent ici le témoignage de notre respectueuse gratitude.
A Monsieur le Professeur Henri DABERNAT et Monsieur le Docteur Pascal SANDERS qui nous ont fait l’honneur d’accepter de faire partie de notre jury de thèse, qu’ils soient assurés de nos plus sincères remerciements.
A Messieurs les Professeurs Pierre-Louis TOUTAIN et Alain BOUSQUET-MELOU qui m’ont accueillie comme doctorante et qui m’ont guidée tout au long de cette thèse, qu’ils soient assurés de ma plus sincère reconnaissance pour m’avoir permis de réaliser ce travail dans les meilleures conditions possibles.
J’adresse mes remerciements à tous les membres de l’équipe « Antibiothérapie et Antibiorésistance » de l’UMR181 qui ont contribué à ce travail. A Madame le Docteur Véronique DUPOUY pour son aide précieuse en microbiologie et en biologie moléculaire. A Madame Nathalie ARPAILLANGE pour son soutien technique en microbiologie. A Madame Sylvie PUEL et Mademoiselle le Docteur Marlène LACROIX pour la réalisation des dosages de marbofloxacine.
Je souhaite également remercier toute l’équipe zootechnique de l’UMR181 pour avoir pris soin des souris pendant ce travail de thèse : Mademoiselle Mihaëla ASTRUC, Monsieur Sylvain BRUYAS, Monsieur Jean-Pierre GAU, Monsieur Joseph MALIGOY, Monsieur Valentin RODRIGUEZ, Monsieur Patrice ROUBY et Monsieur René SCHAN.
Je tiens à remercier les doctorants anciens, actuels et futurs, avec qui j’ai partagé l’expérience de la thèse, pour les bons moments que nous avons passé ensemble. A Delphine BIBBAL pour tout le soutien et les conseils qu’elle m’a apportés pendant ces trois ans, Anne-Sylvie KESTEMAN ma binôme de thèse, Julie ANTIC, Séverine COLLET, Elisabeth JEUNESSE, Julien LEGHAIT et Béatrice ROQUES.
A tous les membres de l’UMR181, pour toute l’aide indispensable, qu’elle soit technique, administrative ou scientifique, qu’ils m’ont apportée.
A ma famille qui m’a permis, par son soutien et sa confiance, de trouver ma voie dans la recherche.
Avant-propos
Les travaux de recherche de cette thèse ont, jusqu’à présent, fait l’objet des publications et communications suivantes :
Articles
1. Aude A. Ferran, Anne-Sylvie Kesteman, Pierre-Louis Toutain, and Alain Bousquet-Mélou
Pharmacokinetic/pharmacodynamic analysis of the Influence of Inoculum Size on the Selection of Resistance in Escherichia coli by a Quinolone in a Mouse Thigh Bacterial Infection Model
Antimicrob. Agents Chemother. 2009 53 : 3384-3390
2. Aude Ferran, Véronique Dupouy, Pierre-Louis Toutain, and Alain Bousquet-Mélou
Influence of Inoculum Size on the Selection of Resistant Mutants of Escherichia coli in Relation to Mutant Prevention Concentrations of Marbofloxacin
Antimicrob. Agents Chemother. 2007 51: 4163-4166.
Communications orales
1. Aude Ferran, Anne-Sylvie Kesteman, Pierre-Louis Toutain and Alain Bousquet-Mélou A PK/PD Analysis of the selection of resistance to marbofloxacin in Escherichia coli in a mouse-thigh infection model
Eleventh International Congress of the European Association for Veterinary Pharmacology and Toxicology. July 14, 2009. Leipzig, Germany.
2. Aude Ferran, Anne-Sylvie Kesteman, Pierre-Louis Toutain and Alain Bousquet-Mélou
Effects of inoculum size on the selection of resistance in Escherichia coli by marbofloxacin in an in vitro pharmacodynamic model and in a mouse-thigh infection model.
Fourth International Conference on Antimicrobial Agents in Veterinary Medicine. August 24, 2008. Prague, Czech Republic
Posters
1. Aude Ferran, Anne-Sylvie Kesteman, Pierre-Louis Toutain and Alain Bousquet-Mélou
A PK/PD Analysis of the Selection of Resistance to Marbofloxacin in Escherichia coli in a Mouse-thigh Infection Model
Third symposium on Antimicrobial Resistance in Animals ad Environment. June 1-3, 2009. Tours, France
2. Aude Ferran, Véronique Dupouy, Sylvie Puel, Charles-Adrien Richard, Pierre-Louis Toutain and Alain Bousquet-Mélou
Effect of inoculum size on the selection resistant Escherichia coli mutants by marbofloxacin
TABLE DES ILLUSTRATIONS ... 9
GLOSSAIRE ... 11
INTRODUCTION ... 13
1 ETUDE BIBLIOGRAPHIQUE... 17
1.1 METHODES D’ETUDE DE L’ACTIVITE DES ANTIBIOTIQUES... 19
1.1.1 Détermination de la Concentration Minimale Inhibitrice ... 19
1.1.2 Courbes de bactéricidie avec des concentrations en antibiotique constantes... 20
1.1.3 Courbes de bactéricidie avec des concentrations en antibiotique variables... 22
1.1.4 Modèles d’études des antibiotiques sur animaux... 25
1.2 LA MARBOFLOXACINE... 27
1.2.1 Spectre d’activité ... 27
1.2.2 Propriétés pharmacocinétiques et pharmacodynamiques ... 27
1.3 MECANISME D’ACTION DES FLUOROQUINOLONES... 29
1.3.1 Cibles intrabactériennes ... 29
1.3.2 Mécanisme de fixation des fluoroquinolones aux complexes enzymes/ADN ... 30
1.3.3 Paramètres prédictifs de l’efficacité des fluoroquinolones ... 31
1.4 RESISTANCES AUX FLUOROQUINOLONES... 37
1.4.1 Mécanismes de tolérance... 37
1.4.2 Mécanismes de résistance ... 38
1.4.3 Paramètres prédictifs de l’émergence de résistance corrélés à la CMI ... 44
1.4.4 Concept de Concentration Prévenant les Mutants ... 46
1.4.5 Paramètres prédictifs de l’émergence de résistance corrélés à la CPM... 47
1.5 INFLUENCE DE LA TAILLE DE L’INOCULUM... 53
1.5.1 Différentes modalités d’antibiothérapie en médecine vétérinaire... 53
1.5.2 Influence de la taille de l’inoculum sur l’activité bactéricide des antibiotiques ... 55
1.5.3 Influence de la taille de l’inoculum sur la sélection de bactéries résistantes ... 56
1.6 CONCLUSION ET PROJET DE THESE... 59
2 ETUDE EXPERIMENTALE ... 61
2.1 INFLUENCE DE LA TAILLE DE L’INOCULUM SUR L’ACTIVITE BACTERICIDE IN VITRO... 63
2.1.1 Problématique... 63
2.1.2 Objectif... 63
2.1.3 Matériels et méthodes ... 63
2.1.4 Résultats ... 66
2.1.5 Discussion... 72
2.2 INFLUENCE DE LA TAILLE DE L’INOCULUM SUR LA SELECTION DE RESISTANCE IN VITRO... 75
2.2.1 Problématique... 75 2.2.2 Objectifs ... 75 2.2.3 Matériel et méthodes ... 76 2.2.4 Résultats ... 79 2.2.5 Discussion... 85 2.2.6 Article 1 ... 89
2.3 MODELE MURIN D’INFECTION DE LA CUISSE AVEC ESCHERICHIA COLI... 95
2.3.1 Problématique... 95 2.3.2 Objectifs ... 95 2.3.3 Matériels et méthodes ... 95 2.3.4 Résultats ... 99 2.3.5 Discussion... 107 2.3.6 Article 2 ... 111
2.4 MODELE MURIN D’INFECTION PULMONAIRE A PASTEURELLA MULTOCIDA... 119
2.4.1 Problématique... 119 2.4.2 Objectifs ... 119 2.4.3 Matériel et méthodes ... 120 2.4.4 Résultats ... 122 2.4.5 Discussion... 129 DISCUSSION GENERALE ... 133 REFERENCES ... 141
Tables des illustrations
Figure 1 : Méthode de détermination de la CMI. ... 19
Figure 2 : Représentation schématique de courbes de bactéricidie. ... 20
Figure 3 : Schéma d’une relation concentration-effet de type Emax... 21
Figure 4 : Système in vitro dynamique ... 22
Figure 5 : Schéma des entrées et des sorties du compartiment central... 23
Figure 6 : Représentation d’une molécule de marbofloxacine. ... 27
Figure 7 : Schéma du mécanisme d’action de la gyrase. ... 29
Figure 8 : Schéma de la fixation de la sous-unité gyrA à l’ADN... 30
Figure 9 : Représentation schématique des concentrations plasmatiques en antibiotique ... 32
Figure 10 : Nombre de bactéries versus AUC/CMI, Cmax/CMI et T>CMI... 33
Figure 11 : Impact de l’AUIC sur l’éradication bactérienne ... 34
Figure 12 : Description des mécanismes de résistances aux fluoroquinolones ... 38
Figure 13 : Méthode de détermination de la CPM. ... 46
Figure 14 : Schéma d’une fenêtre de sélection... 47
Figure 15 : Profils de concentrations différents avec un même Cmax/CMI. ... 48
Figure 16 : Représentation schématique d’administration simultanée de 2 antibiotiques (1)... 51
Figure 17 : Représentation schématique d’administration simultanée de 2 antibiotiques (2)... 51
Figure 18 : Représentation des aires utilisées pour le calcul de l'effet bactéricide... 65
Figure 19 : Détermination de la CMI de la marbofloxacine pour Escherichia coli ATCC 25922... 66
Figure 20 : Croissance d’Escherichia coli ATCC 25922 à partir d’un inoculum de 105 UFC/mL ... 66
Figure 21 : Croissance d'Escherichia coli ATCC 25922 à partir d'un inoculum de 108 UFC/mL ... 67
Figure 22 : Croissance des bactéries en incubation à 37°C avant l'ajout de marbofloxacine. ... 67
Figure 23 : Courbes de bactéricidie obtenues à partir d'un inoculum de 105 UFC/mL. ... 68
Figure 24 : Courbes de bactéricidie obtenues à partir d'un inoculum de 107 UFC/mL. ... 68
Figure 25 : Courbes de bactéricidie obtenues à partir d'un inoculum de 109 UFC/mL. ... 68
Figure 26 : Vitesses de bactéricidie de la marbofloxacine en fonction de l’inoculum. ... 69
Figure 27 : Effets bactéricides de la marbofloxacine en fonction de l’inoculum... 69
Figure 28 : Courbes de bactéricidie obtenues à partir d'un inoculum de 107 UFC/mL (1)... 70
Figure 29 : Courbes de bactéricidie obtenues à partir d'un inoculum de 107 UFC/mL (2)... 71
Figure 30 : Vitesses de bactéricidie en fonction du stade de croissance. ... 71
Figure 31 : Effets bactéricides en fonction du stade de croissance. ... 72
Figure 32 : Schéma de la structure du système in vitro dynamique. ... 77
Figure 33 : Représentation de l’aire utilisée pour le calcul de l'effet bactéricide ... 78
Figure 34 : Détermination de la CPM de la marbofloxacine pour Escherichia coli ATCC 25922 ... 79
Figure 35 : Schéma de la fenêtre de sélection des premiers mutants ... 80
Figure 36 : Croissance des bactéries dans le système in vitro dynamique ... 80
Figure 37 : Concentrations de marbofloxacine mesurées dans le compartiment central. ... 81
Figure 38 : Nombres de bactéries vivantes observées ... 82
Figure 39 : Fréquence de bactéries résistantes ... 84
Figure 40 : Concentrations plasmatiques de marbofloxacine en fonction du temps... 100
Figure 41 : Populations bactériennes totales en fonction de la dose de marbofloxacine ... 102
Figure 42 : Proportions de bactéries résistantes en fonction de la dose de marbofloxacine (1) ... 104
Figure 43 : Proportions de bactéries résistantes en fonction de la dose de marbofloxacine (2) ... 105
Figure 44 : Proportions de bactéries résistantes versus AUC24/CMI, Cmax/CMI et TFS... 106
Figure 45 : Survie des souris après infection ... 122
Figure 46 : Croissance de Pasteurella multocida dans le poumon de souris ... 123
Figure 47 : Concentrations plasmatiques de marbofloxacine ... 124
Tableau 1 : Valeurs minimales de l’indice AUC/CMI prévenant la sélection de résistances. ... 44
Tableau 2 : Valeurs minimales de l’indice Cmax/CMI prévenant la sélection de résistances... 45
Tableau 3 : Valeurs des paramètres du modèle Emax pour chaque inoculum... 70
Tableau 4 : Paramètres pharmacocinétiques pour chaque profil de concentrations ... 81
Tableau 5 : Activité bactéricide de la marbofloxacine et sélection de mutants résistants (1)... 83
Tableau 6 : Activité bactéricide de la marbofloxacine et sélection de mutants résistants (2)... 83
Tableau 7 : Activité bactéricide de la marbofloxacine et sélection de mutants résistants (3)... 83
Tableau 8 : Paramètres pharmacocinétiques de la marbofloxacine ... 100
Tableau 9 : Valeurs estimées des indices TFS, AUC24/CMI et Cmax/CMI (1) ... 101
Tableau 10 : Valeurs estimées des indices TFS, AUC24/CMI et Cmax/CMI (2) ... 101
Tableau 11 : Populations totales de bactéries dans les cuisses (1) ... 102
Tableau 12 : Valeurs des paramètres du modèle Emax inhibition pour chaque inoculum ... 103
Tableau 13 : Populations totales de bactéries dans les cuisses (2) ... 103
Tableau 14 : Bilan cliniques et microbiologiques ... 125
Tableau 15 : Populations totales de bactéries dans les poumons ... 125
Tableau 16 : Pourcentages de poumons contenant des bactéries résistantes (1) ... 126
Tableau 17 : Pourcentages de poumons contenant des bactéries résistantes (2) ... 127
Tableau 18 : Valeurs des indices PK/PD ... 127
Équation 1 ... 20 Équation 2 ... 24 Équation 3 ... 24 Équation 4 ... 24 Équation 5 ... 65 Équation 6 ... 69 Équation 7 ... 73 Équation 8 ... 99 Équation 9 ... 103
Glossaire
ATCC American Type Cell Culture
AUC Aire sous la courbe des concentrations (Area Under the Curve)
AUIC Aire sous les concentrations inhibitrices (Area Under the Inhibitory Curve) Cmax Concentration maximale en antibiotique
Cl Clairance
CLSI Clinical and Laboratory Standard Institute CMB Concentration Minimale Bactéricide CMI Concentration Minimale Inhibitrice CPM Concentration Prévenant les Mutants. ENVT Ecole Nationale Vétérinaire de Toulouse
EUCAST European Commitee on Aantimicrobial Susceptibility Testing FS Fenêtre de Sélection
HPLC High Performance Liquid Chromatography INRA Institut National de Recherche Agronomique
MH Mueller-Hinton
PD Pharmacodynamie
PK Pharmacocinétique
PMQR Plasmid-mediated Quinolone Resistance QRDR Quinolone-Resistance Determining Region
T>CMI Pourcentage du temps pendant lequel les concentrations sont au-dessus de la
CMI
T>CPM Pourcentage du temps pendant lequel les concentrations sont au-dessus de la
CPM
TFS Pourcentage du temps pendant lequel les concentrations sont dans la Fenêtre
de Sélection TA Toxine-Antitoxine
UFC Unité Formant une Colonie UMR Unité Mixte de Recherche
Les travaux de recherche présentés dans ce document ont été menés dans l’équipe « Antibiothérapie et Antibiorésistance » de l’UMR181 Physiopathologie et Toxicologie Expérimentales, INRA, ENVT. L’équipe « Antibiothérapie et Antibiorésistance » s’intéresse aux effets de l’antibiothérapie sur les bactéries pathogènes ainsi que sur les bactéries de la flore commensale en se focalisant principalement sur l’identification de stratégies thérapeutiques (choix de l’antibiotique, doses, voies d’administration,…) capables d’éradiquer l’agent pathogène tout en limitant la sélection de bactéries résistantes au niveau du site infectieux et de la flore commensale. Mon travail de thèse, sous la direction du Professeur Pierre-Louis Toutain et du Professeur Alain Bousquet-Mélou, s’est intégré dans la thématique de recherche relative à la détermination de schémas posologiques capables de prévenir l’émergence de résistances bactériennes au niveau du site infectieux.
Les fluoroquinolones constituent une classe d’antibiotiques, développée à partir des années 70 ; elles sont très efficaces sur un large spectre de bactéries et de nombreuses infections telles que les infections cutanées, respiratoires ou urinaires sont traitées avec ces antibiotiques. Cependant, l’usage croissant des fluoroquinolones aussi bien en médecine humaine qu’en médecine vétérinaire engendre progressivement l’émergence de résistances préoccupantes qu’il est nécessaire de limiter ou de prévenir en proposant de meilleures pratiques d’antibiothérapie.
En médecine vétérinaire, les antibiothérapies préventives telles que la prophylaxie et la métaphylaxie sont très fréquentes notamment en élevage. La métaphylaxie consiste à traiter tous les animaux d’un troupeau lorsque seulement quelques individus présentent des symptômes cliniques. Les animaux, qui sont alors traités de manière préventive avec des doses d’antibiotique curatives, ne sont a priori porteurs que d’une très petite quantité de bactéries, une caractéristique qui pourrait être favorable à un succès de l’antibiothérapie dans l’éradication des bactéries pathogènes et à la prévention de l’émergence de résistances bactériennes.
C’est dans ce contexte que nous avons décidé d’évaluer l’interaction entre la charge
bactérienne et l’exposition à une fluoroquinolone commercialisée pour un usage
vétérinaire, la marbofloxacine, sur l’éradication bactérienne et sur la sélection de bactéries résistantes au niveau du site infectieux.
Une première partie bibliographique présentera les différentes méthodes d’études de l’activité des antibiotiques utilisées dans notre travail de thèse (chapitre 1.1). Les principales propriétés de la marbofloxacine (chapitre 1.2), le mécanisme d’action des fluoroquinolones (chapitre 1.3) ainsi que les mécanismes de résistances des bactéries à ces antibiotiques (chapitre 1.4) seront également décrits. En parallèle, les indices pharmacocinétiques/ pharmacodynamiques capables de prédire l’activité des fluoroquinolones et l’émergence de résistance seront présentés ainsi que les valeurs seuils de ces indices proposées dans la littérature. Enfin, les différentes modalités d’utilisation des antibiotiques en médecine vétérinaire ainsi que l’influence de la taille de l’inoculum bactérien sur l’activité des antibiotiques et sur l’émergence de résistance seront discutées (chapitre 1.5).
Les travaux de recherche pour cette thèse ont débuté par deux études menées in vitro destinées à évaluer l’interaction entre la taille d’un inoculum bactérien composé d’Escherichia coli et l’exposition des bactéries à la marbofloxacine sur la bactéricidie (chapitre 2.1) et sur la sélection de bactéries résistantes (chapitre 2.2). Les hypothèses émises in vitro ont ensuite été vérifiées lors d’une étude réalisée in vivo (chapitre 2.3) sur un modèle d’infection de cuisses de souris avec la même bactérie, Escherichia coli. Enfin, l’impact d’un traitement précoce a été évalué sur un modèle murin d’infection expérimentale pulmonaire à Pasteurella multocida plus représentatif des infections rencontrées en élevages bovins et porcins (chapitre 2.4).
1.1 Méthodes d’étude de l’activité des
antibiotiques
Il existe actuellement deux méthodes principales pour étudier l’activité des antibiotiques in
vitro.
La détermination de la Concentration Minimale Inhibitrice (CMI) est la plus simple. Elle
donne des informations sur la sensibilité in vitro des bactéries à un antibiotique après exposition à une concentration constante pendant une durée fixée.
Les courbes de bactéricidie permettent de connaître les effets in vitro des antibiotiques au
cours du temps sur des bactéries exposées : à des concentrations constantes ou
à des concentrations variables en antibiotique par le développement de systèmes
in vitro dynamiques.
Enfin, les modèles animaux permettent de prendre en compte l’accessibilité de l’antibiotique au site infectieux in vivo et l’influence du système immunitaire.
1.1.1 Détermination de la Concentration
Minimale Inhibitrice
La CMI permet d’évaluer la sensibilité des bactéries aux antibiotiques. Cette concentration est déterminée par une méthode standardisée par le CLSI (Clinical and Laboratory Standarts Institute) [CLSI, 2003 #113]. Un inoculum de 5.105 bactéries/mL est exposé à une gamme de concentrations d’antibiotique obtenue par une série de dilutions au demi. La croissance des bactéries est observée à l’œil nu après 15 à 18 heures d’exposition à l’antibiotique à 35°C. La CMI est alors la concentration la plus faible pour laquelle aucune croissance n’est visible à l’œil nu (figure 1). 16 8 4 2 1 CMI dilution par 2 Concentrations en antibiotique (µg/mL) 16 8 4 2 1 CMI dilution par 2 16 8 4 2 1 CMI dilution par 2 Concentrations en antibiotique (µg/mL)
Figure 1 : Méthode de détermination de la CMI.
Les zones grisées correspondent à une croissance bactérienne visible à l’œil nu. La CMI est la concentration en antibiotique pour laquelle aucune croissance n’est visible après 18 heures d’incubation à 35°C.
Cette méthode est simple, rapide et standardisée ce qui permet les comparaisons entre laboratoires. Cependant, il est important de remarquer que la méthode ne nécessitant que 5.105 bactéries/mL permet uniquement d’obtenir la sensibilité de la souche majoritaire de la population bactérienne échantillonnée. En effet, une seule colonie est prélevée au hasard à partir d’un ensemencement sur gélose et, par conséquent, cette colonie est le plus souvent représentative de la souche majoritaire dans le prélèvement. De plus, la dilution de l’inoculum pour obtenir une suspension de 5.105 bactéries/mL réduit fortement la probabilité de conserver dans l’échantillon des mutants spontanés dont la probabilité d’apparition est de 10-8 (13). Par ailleurs, la détermination de la CMI ne permet d’évaluer l’effet de l’antibiotique
qu’après un temps d’incubation fixé. Pour connaître l’activité de l’antibiotique au cours du
temps, il faut dénombrer les bactéries à intervalles réguliers afin d’obtenir des courbes de bactéricidie.
1.1.2 Courbes de bactéricidie avec des
concentrations en antibiotique constantes
Les courbes de bactéricidie sont le plus souvent obtenues par des dénombrements de bactéries exposées à des concentrations constantes en antibiotiques. Ces courbes réalisées pour plusieurs classes d’antibiotiques ont permis de mettre en évidence deux modes d’action (figure 2) : un mode concentration-dépendant et un mode temps-dépendant.Temps UF C/ mL UFC / mL Temps Antibiotiques concentration-dépendants Antibiotiques temps-dépendants 0 Concentration 1 Concentration 2 Concentration 3 avec C3>C2>C1 Temps UF C/ mL UFC / mL Temps Antibiotiques concentration-dépendants Antibiotiques temps-dépendants 0 Concentration 1 Concentration 2 Concentration 3 avec C3>C2>C1 0 Concentration 1 Concentration 2 Concentration 3 avec C3>C2>C1
Figure 2 : Représentation schématique de courbes de bactéricidie.
La figure de gauche représente le cas d’un antibiotique concentration-dépendant ; l’activité bactéricide augmente avec la concentration. La figure de droite représente le cas d’un antibiotique temps-dépendant ; l’activité
bactéricide n’augmente pas avec la concentration mais avec le temps.
Les antibiotiques agissent sur des cibles bactériennes et leur activité sur ces récepteurs peut être facilement décrite par un modèle de type Emax (équation 1) qui décrit la relation
concentration – effet lors d’une interaction entre une molécule et un récepteur (74).
50 max
EC
C
C
E
E
Équation 1où E est l’effet observé pour la concentration C, Emax l’effet maximal, EC50 la concentration
nécessaire pour obtenir un effet égal à 50 % de l’effet maximal et γ le coefficient de sigmoïdicité.
La relation concentration-effet décrite par ce modèle est typiquement une relation sigmoïde (figure 3). C oncen tr ati o n s attei ntes pou r le s tem p s-dé p e n d a n ts Concentrations en antibiotique Effets Emax EC50 Conc en tr ation s attei n tes pou r le s con cen tr atio n-dé p e n d a n ts C oncen tr ati o n s attei ntes pou r le s tem p s-dé p e n d a n ts Concentrations en antibiotique Effets Emax EC50 Conc en tr ation s attei n tes pou r le s con cen tr atio n-dé p e n d a n ts
Figure 3 : Schéma d’une relation concentration-effet de type Emax.
L’effet augmente avec la concentration en antibiotique .
Pour des concentrations en antibiotique faibles, l’effet est quasiment nul puis quand les concentrations augmentent, l’effet augmente proportionnellement aux concentrations (plus ou moins vite selon la pente de la courbe) et enfin, pour de fortes concentrations, un effet maximal est atteint, on parle alors de « plateau ».
Selon la toxicité des molécules, les concentrations plasmatiques permettent ou non d’atteindre un effet maximal. Pour des antibiotiques tels que les bêta-lactamines pour lesquels les doses administrées peuvent être élevées, les concentrations permettent d’obtenir un effet maximal situé au niveau du plateau et un changement de concentrations n’a alors que peu d’influence sur l’intensité de l’effet qui reste maximal. L’activité de l’antibiotique dépend donc principalement du temps passé au niveau du plateau. Ces antibiotiques sont dits temps-dépendants. A l’opposé, pour certains antibiotiques aux marges thérapeutiques étroites tels que les aminoglycosides ou les fluoroquinolones, les concentrations pour des raisons de toxicité ne permettent pas d’atteindre l’effet maximal et la décroissance des concentrations est alors associée à une diminution de l’effet. Ces antibiotiques dont l’effet dépend des concentrations sont dits concentration-dépendants.
La classification des antibiotiques en concentration ou temps-dépendants a conduit à une meilleure efficacité des antibiothérapies par une adaptation des schémas posologiques. Ainsi, des administrations fréquentes ont été préconisées pour les antibiotiques
temps-dépendants afin de maintenir longtemps les concentrations au-dessus de la CMI alors que des administrations uniques sont recommandées pour les antibiotiques concentration-dépendants afin d’atteindre des concentrations élevées sans pour autant couvrir tout l’intervalle d’administration.
Les courbes de bactéricidie, obtenues en exposant les bactéries à des concentrations en antibiotique constantes, sont faciles à réaliser et permettent de connaître rapidement la modalité d’action d’un antibiotique sur une souche bactérienne. Cependant, cette méthode ne prend pas en compte la dynamique temporelle des concentrations en antibiotiques in
vivo.
1.1.3 Courbes de bactéricidie avec des
concentrations en antibiotique variables
Des systèmes appelés in vitro dynamiques ont été développés pour permettre d’exposer les bactéries à des concentrations en antibiotique variables au cours du temps. L’intérêt de ces systèmes est de mimer l’élimination des antibiotiques par l’organisme in vivo.1.1.3.1 Structures de systèmes in vitro dynamiques
La structure générale des systèmes dynamiques consiste en plusieurs compartiments matérialisés par des béchers immergés dans un bain-marie à 37°C ou placés dans une chambre thermostatée à 37 °C (figure 4).
Bain-marie Agitateur magnétique Déchets Bouillon + bactéries + antibiotiques Bouillon Pompe 37°C Prélèvements Thermostat Bain-marie
Agitateur magnétique Bain-marie
Agitateur magnétique Déchets Bouillon + bactéries + antibiotiques Bouillon Pompe 37°C Prélèvements Thermostat
Figure 4 : Système in vitro dynamique
Les bactéries sont mises en contact avec l’antibiotique dans le compartiment central immergé dans un bain-marie thermostaté à 37°C. Une pompe assure la dilution des antibiotiques et l’autre maintient le volume constant. Un
agitateur magnétique homogénéise la suspension bactérienne.
Le compartiment central contenant du bouillon de culture représente le système vasculaire et l’objectif est d’y retrouver l’équivalent des concentrations plasmatiques d’antibiotique
injectés directement dans le compartiment central. Pour simuler une administration extravasculaire, une pompe prélève les antibiotiques dans un compartiment périphérique et les amène progressivement au compartiment central. Suite à l’arrivée des antibiotiques dans ce compartiment, leur élimination débute. Une pompe apporte du bouillon de culture sans antibiotique au compartiment central afin de diluer les antibiotiques selon le débit choisi (voir paragraphe 1.1.3.2). Une autre pompe permet de maintenir le volume du compartiment central constant par une élimination de bouillon vers un compartiment déchet. Dans la plupart des systèmes, l’homogénéisation des bactéries est assurée par une agitation magnétique. Cependant, dans certains systèmes avec des structures en dérivation, des pompes assurent l’homogénéisation à l’intérieur des structures contenant les bactéries (12).
La culture bactérienne est réalisée dans différents compartiments selon le degré de complexité du système. Dans les systèmes les plus simples, les bactéries sont en suspension dans le compartiment central et sont donc éliminées vers le compartiment déchet en même temps que les antibiotiques (figure 4) (2, 39, 92). Dans les systèmes plus sophistiqués, les bactéries sont isolées du compartiment central pour prévenir leur élimination au cours de l’expérience. La séparation entre la suspension bactérienne et le compartiment central peut être matérialisée par un filtre dont les pores, qui ont un diamètre de 0.4 µm, permettent le passage des antibiotiques mais pas celui des bactéries (43, 67).
Enfin, dans les systèmes les plus complexes (12, 46), les bactéries sont placées dans des structures isolées en dérivation. Le contenu du compartiment central contenant les antibiotiques circule à travers ces structures grâce à une pompe, les bactéries étant maintenues par des filtres de pores de 0.4 µm.
1.1.3.2 Débit d’élimination des antibiotiques
Dans les systèmes in vitro dynamiques, les antibiotiques sont éliminés progressivement du compartiment central selon un débit d’élimination choisi. Afin de garder un volume constant, le débit d’apport en bouillon sans antibiotique est identique au débit d’élimination des antibiotiques (figure 5).
Compartiment central Avec une quantité x d’antibiotique
répartie dans un volume V Débit d’apport en bouillon
sans antibiotique
Débit d’élimination des antibiotiques (Clairance)
Figure 5 : Schéma des entrées et des sorties du compartiment central.
Le débit d’élimination choisi pour faire décroître les concentrations en antibiotique à la même vitesse qu’in vivo est aussi appliquée à l’entrée du système.
On peut affecter aux systèmes in vitro dynamiques la notion de clairance (Cl) qui est le coefficient de proportionnalité entre la vitesse d’élimination de l’antibiotique et la concentration d’antibiotique au temps t dans le système (équation 2).
)
/
(
)
/
(
lim
'
) ( ) (mL
µg
ion
Concentrat
h
µg
ination
é
d
vitesse
Cl
t t
Équation 2La clairance du système s’exprime en mL/h et correspond au volume du système complètement épuré de l’antibiotique par unité de temps. Par définition, le rapport entre la clairance et le volume total du système correspond à la fraction du volume complètement épurée par unité de temps (k10) (équation 3).
V
Cl
K
10
Équation 3où Cl est la clairance et V le volume du système
Pour reproduire une élimination similaire à celle observée in vivo, il faut connaître le temps de demi-vie (T1/2) de l’antibiotique. Il est alors possible de calculer la fraction à épurer par
unité de temps (k10) pour obtenir ce temps de demi-vie d’après l’équation 4.
2 / 1 10
2
ln
T
K
Équation 4Connaissant la fraction à épurer par unité de temps et le volume du système in vitro, il est alors possible de calculer la clairance qui est le débit à appliquer aux pompes situées en amont et en aval du compartiment central afin d’obtenir une décroissance des concentrations en antibiotique similaire à celle observée in vivo et de maintenir le volume du compartiment central constant.
Cependant, même les systèmes d’études in vitro les plus perfectionnés capables de mimer l’élimination progressive des antibiotiques au contact des bactéries ne permettent pas de prendre en compte toutes les paramètres qui peuvent influer sur l’efficacité d’un traitement antibiotique in vivo tels que l’accessibilité au site infectieux.
1.1.4 Modèles d’études des antibiotiques sur
animaux
Les études de l’activité des antibiotiques sur modèles animaux sont très utilisées dans le développement d’antibiotiques en parallèle des études in vitro pour évaluer rapidement la capacité des antibiotiques à atteindre et à tuer les bactéries in vivo. Le modèle de septicémie de la souris est, par exemple, un modèle de choix dans le screening des molécules antibiotiques qui a été utilisé pour étudier les effets du premier sulfamide en 1935 (24).
Dans les premières étapes du développement d’un antibiotique, les études sont souvent limitées à des évaluations de la survie des animaux alors que dans les étapes ultérieures, le nombre de bactéries après traitement est directement évalué dans le tissu infecté.
Les modèles animaux sont actuellement le meilleur moyen de tester l’activité et la
sécurité de nouveaux antibiotiques avant l’administration à des patients et sont aussi un
excellent moyen de trouver de nouvelles approches thérapeutiques (dose, intervalle d’administration,…). Il a par exemple été montré que les données de valeurs seuils d’indice PK/PD pour prédire une guérison microbiologique ou clinique obtenues dans des modèles animaux depuis 15 ans avaient une bonne concordance avec les valeurs retrouvées dans des études cliniques (6).
Le site d’infection choisi dans le modèle animal est, dans la plupart des cas, le site de
l’infection naturelle. Cependant, il existe un modèle qui a été très utilisé pour décrire
l’activité des antibiotiques in vivo et qui ne correspond pas à une infection naturelle ; il s’agit du modèle d’infection de la cuisse de souris immunodéprimées. Ce modèle a été utilisé la première fois en 1950 par Eagle et al. (30) pour tester les effets de la pénicilline G sur différents streptocoques. Le modèle a ensuite été amélioré par Craig et al. (44) qui ont rendu les souris neutropéniques par l’injection de cyclophosphamide. De nombreuses combinaisons antibiotique-bactérie ont été étudiées dans ce modèle du fait de sa facilité de mise en place qui consiste en une injection intramusculaire d’une suspension de la souche bactérienne choisie. Enfin, bien que l’infection du muscle ne soit pas une infection d’intérêt en médecine, les résultats obtenus dans ce modèle semblent être représentatifs des traitements des septicémies puisque le plus souvent les concentrations libres des antibiotiques au niveau musculaire sont très proches des concentrations plasmatiques libres.
En conclusion, les modèles d’études in vitro et in vivo qui sont complémentaires ont permis de mieux connaître l’activité antibactérienne de nombreux antibiotiques. Ces modèles sont maintenant de plus en plus utilisés pour l’étude de l’émergence de
résistances bactériennes et les résultats expérimentaux de cette thèse ont pour la plupart
1.2 La marbofloxacine
La marbofloxacine est un agent antibactérien de synthèse, à usage vétérinaire, appartenant à la famille des fluoroquinolones. Cet antibiotique ayant des indications thérapeutiques aussi bien chez les animaux de compagnie que chez les animaux de rente a été utilisé pour l’ensemble des études expérimentales de ce travail de thèse.
1.2.1 Spectre
d’activité
La marbofloxacine a un spectre d’activité large et est indiquée dans le traitement de diverses infections à bactéries à Gram négatif ou à Gram positif.
Elle est indiquée dans le traitement d’infections respiratoires à Pasteurella multocida,
Mannheimia haemolytica et Mycoplasma bovis chez les bovins et à Actinobacillus pleuropneumoniae, Mycoplasma hyopneumoniae et Pasteurella multocida chez le porc. La
marbofloxacine est également utilisable pour le traitement des gastro-entérites dues à
Escherichia coli K99 chez le veau et pour le traitement du syndrome
mammite-métrite-agalactie chez la truie.
Chez le chien, la marbofloxacine est indiquée dans le traitement des pyodermites, des infections respiratoire et des infections du tractus urinaire dues à Escherichia coli ou à
Proteus mirabilis. Chez le chat, la marbofloxacine est indiquée dans le traitement des
infections cutanées. Pour les carnivores domestiques, la formulation injectable de la marbofloxacine peut aussi administrée en prophylaxie chirurgicale.
1.2.2 Propriétés
pharmacocinétiques
et
pharmacodynamiques
La figure 6 présente la structure de la molécule de marbofloxacine.
CH3 N N N N F O O CH3 O OH
Figure 6 : Représentation d’une molécule de marbofloxacine. Les groupements d’intérêts pour l’activité antibactérienne sont entourés.
Les 2 atomes d’oxygène entourés sur la figure 6 permettent la liaison de la molécule à ses cibles bactériennes. Deux autres groupements chimiques de la molécule, le fluor et le groupement méthyle, sont particulièrement intéressants. La présence d’un fluor facilite le passage de la membrane des bactéries à Gram positif et le groupement méthyle confère à la marbofloxacine une activité bactéricide sur les Pseudomonas.
La biodisponibilité de la marbofloxacine est élevée. Le pourcentage de la dose qui parvient au système vasculaire après une administration extravasculaire varie entre 62 à 100 % selon la voie d’administration. Le temps nécessaire pour atteindre le pic de concentration plasmatique est court (inférieur à 2 heures).
La diffusion dans l’organisme de la marbofloxacine est large ; le volume de distribution à l’équilibre est d’environ 1.3 L/kg. La liaison aux protéines plasmatiques est faible (inférieure à 10% pour les carnivores domestiques et les porcs, inférieure à 30% pour les bovins). L’élimination est lente. La marbofloxacine est essentiellement éliminée sous forme active, pour 2/3 par voie urinaire et pour 1/3 par voie fécale dans la plupart des espèces animales. Le temps de demi-vie est de 4 à 14 heures selon l’espèce.
La marbofloxacine pénètre dans le cytoplasme des bactéries à Gram positif par diffusion. Le passage de la membrane externe des bactéries à Gram négatif se fait par la complexation des ions magnésium qui provoque une désorganisation des lipopolysaccharides ou par un passage actif à travers des protéines transmembranaires appelées porines. Le mécanisme d’action de la marbofloxacine, similaire à celui des autres fluoroquinolones, est présenté dans la section suivante (section 1.3).
1.3 Mécanisme d’action des
fluoroquinolones
Les connaissances actuelles sur le mécanisme d’action des fluoroquinolones ont fait l’objet d’une revue en 2008 par Drlica et al. (26).
1.3.1 Cibles
intrabactériennes
Les cibles bactériennes des fluoroquinolones sont deux enzymes : la gyrase et la
topoisomérase IV. La gyrase a une structure tétramérique composée de 2 sous-unités A
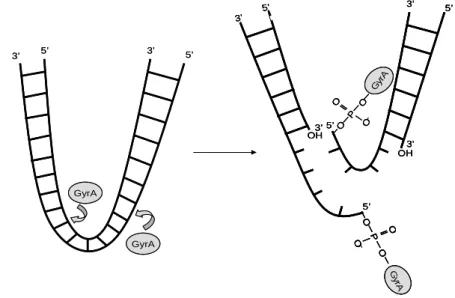
codées par le gène gyrA et de 2 sous-unités B codées par le gène gyrB. Cette enzyme défait les superenroulements de l’ADN en amont de la fourchette de réplication par des cassures d’ADN suivies de religation (Figure 7).
b
ADN
double
brin
Gyrase
a
a
b
b
b
b
ADN
double
brin
Gyrase
a
a
b
Figure 7 : Schéma du mécanisme d’action de la gyrase.
L’ADN double brin est clivé par la gyrase puis passe à travers la cassure avant que la boucle soit refermée. La topoisomérase IV a aussi une structure tétramérique dont les 2 sous-unités A sont codées par le gène parC et les 2 sous-unités B par le gène parE. Elle intervient dans la décaténation de l’ADN, c'est-à-dire dans la séparation des brins d’ADN en fin de réplication (51, 56, 96).
1.3.2 Mécanisme de fixation des
fluoroquinolones aux complexes
enzymes/ADN
La gyrase et la topoisomérase IV sont des enzymes intervenant dans la réplication de l’ADN qui créent au cours de leur activité des paires de fragments simples brins à plusieurs endroits sur l’ADN. Elles se lient alors de manière covalente à l’extrémité 5’ de l’ADN clivé (Figure 8). 3’ 5’ 3’ 5’ GyrA GyrA 5’ 3’ 5’ 3’ G yrA O P O O -O Gyr A O P O O -O 5’ 5’ 3’ 3’ OH OH 3’ 5’ 3’ 5’ GyrA GyrA 5’ 3’ 5’ 3’ G yrA O P O O -O Gyr A O P O O -O 5’ 5’ 3’ 3’ OH OH 5’ 3’ 5’ 3’ G yrA O P O O -O G yrA O P O O -O O P O O -O Gyr A O P O O -O 5’ 5’ 3’ 3’ OH OH
Figure 8 : Schéma de la fixation de la sous-unité gyrA à l’ADN.
Les brins simples après la cassure de l’ADN sont liés de manière covalente à GyrA par l’extrémité 5’.
Les quinolones qui se lient au complexe enzyme/ADN se fixent rapidement à la gyrase ou à la topoisomérase IV probablement avant que l’ADN ne soit clivé ; il se forme alors un
complexe ternaire. L’affinité des quinolones aux simples brins d’ADN est corrélée à la
puissance des quinolones in vitro. La liaison aux sous-unités GyrA ou ParC se fait au niveau de leur hélice 4 qui est située près la cassure de l’ADN. Les mutations, des substitutions d’acides aminés, qui sont à l’origine de la diminution de la sensibilité des bactéries aux fluoroquinolones se situent sur cette hélice ; elles diminuent l’affinité des fluoroquinolones pour ces cibles.
La fixation des fluoroquinolones aux complexes enzyme-ADN inhibe la religation des fragments simples brins de l’ADN en maintenant les enzymes fixées à l’ADN. Ces structures, appelées « complexes clivés », se situent au niveau de nombreux sites préférentiels sur le chromosome et inhibent la synthèse d’ADN en bloquant la fourche de réplication. Pour les bactéries à Gram négatif où la gyrase est la cible préférentielle pour la majorité des fluoroquinolones, la réplication est bloquée en quelques minutes. Pour les bactéries à Gram
l’inhibition de la synthèse d’ADN est plus lente probablement parce que la topoisomérase IV est située en aval des fourches de réplication alors que la gyrase est en amont. La collision entre la fourche et le complexe clivé n’entraîne pas la libération des fragments d’ADN, les brèches de l’ADN sont refermées par la libération des topoisomérases ou lors du retrait de l’antibiotique. Le fait que ces complexes clivés soient réversibles suggère qu’ils ne sont pas létaux. En effet, en présence de chloramphénicol qui est un inhibiteur de la synthèse protéique, l’acide oxolinique inhibe toujours la synthèse d’ADN mais n’est plus bactéricide. Il semble donc exister un facteur protéique qui est responsable de la mort des bactéries.
1.3.3 Paramètres prédictifs de l’efficacité des
fluoroquinolones
L’efficacité d’un antibiotique sur une bactérie dépend de la sensibilité et de l’exposition de la souche à cet antibiotique.
Des paramètres pharmacodynamiques indiquent la sensibilité de la souche et des
paramètres pharmacocinétiques décrivant les concentrations d’antibiotique au contact des
bactéries permettent d’évaluer l’exposition des bactéries.
1.3.3.1 Sensibilité d’une souche à l’antibiotique
En pratique, l’évaluation de la sensibilité d’une souche pour le choix de l’antibiothérapie repose sur un antibiogramme. La technique consiste à imbiber plusieurs antibiotiques sur des disques de papier et à les déposer sur une gélose ensemencée avec la souche à tester en culture pure. Après 24 à 48 heures d’incubation, le diamètre de la zone autour du disque où la croissance des bactéries a été inhibée (diamètre d’inhibition) est mesuré. Il permet alors de classer les bactéries comme sensibles, intermédiaires ou résistantes à l’antibiotique. Les valeurs de diamètres qui définissent les limites entre les trois classes sont déterminées par des organismes normatifs tels que le CLSI (Clinical and Laboratory Standards Institute) ou l’EUCAST (European Committee on Antimicrobial Susceptibility Testing). Il existe une relation entre ce diamètre d’inhibition et la CMI décrite par des courbes de concordance et à ce titre, la CMI est indirectement utilisée pour évaluer la sensibilité d’une souche bactérienne.
Il faut cependant remarquer que la CMI est toujours déterminée après une exposition des bactéries à des concentrations constantes en antibiotiques ce qui diffère des expositions in
vivo. D’autres paramètres ont donc été choisis pour prendre en compte l’évolution des
Trois paramètres pharmacocinétiques (PK) sont généralement utilisés pour décrire l’exposition d’un organisme à un antibiotique (figure 9):
T>Concentration : le pourcentage de temps au-dessus d’une concentration,
AUC : l’aire sous la courbe des concentrations plasmatiques,
Cmax : la concentration plasmatique maximale.
AUC
Temps Concentrations plasmatiquesC
maxT
>ConcentrationAUC
Temps Concentrations plasmatiquesC
maxT
>ConcentrationFigure 9 : Représentation schématique des concentrations plasmatiques en antibiotique
au cours du temps après une administration extravasculaire. Les paramètres pharmacocinétiques qui décrivent l’exposition sont notés en gras.
Les indices pharmacocinétique/pharmacodynamique (PK/PD) sont hybrides ; ils tiennent compte à la fois de la sensibilité et de l’exposition de la souche à l’antibiotique. Les valeurs des paramètres PK utilisées dans le calcul de ces indices sont les valeurs correspondant aux concentrations plasmatiques libres qui sont les seules à pouvoir atteindre les bactéries. Par ailleurs, les concentrations plasmatiques sont très souvent représentatives des concentrations tissulaires au contact des bactéries. En effet, les bactéries pathogènes en médecine vétérinaire ont pour la plupart une localisation extracellulaire et l’équilibre qui s’établit entre les concentrations libres dans le plasma et les tissus indique que les concentrations plasmatiques libres sont les mêmes que les concentrations libres tissulaires.
Les indices PK/PD sont obtenus en couplant les trois paramètres PK à la CMI de l’antibiotique :
T>CMI : le pourcentage de temps entre 0 et 24 heures à l’équilibre pendant lequel les
concentrations plasmatiques sont au-dessus de la CMI. Cet indice est exprimé en pourcents, il ne tient pas compte de l’intensité de l’exposition (concentrations) mais seulement de la durée de l’exposition à une concentration critique qui est la CMI.
Cmax/CMI : la concentration plasmatique maximale divisée par la CMI. Cet indice
est un rapport entre 2 concentrations et n’a pas d’unité. A l’opposé du T>CMI, il tient
AUC/CMI : l’aire sous la courbe des concentrations plamsatiques entre 0 et 24
heures à l’équilibre divisée par la CMI ce qui correspond à l’AUC de 0 à l’infini divisée par la CMI après l’adminsitration d’une dose unique. Cet indice est exprimé en heures et il tient compte de la durée et de l’intensité de l’exposition.
Il est intéressant de remarquer qu’il a aussi été décrit un indice AUIC (area under the inhibitory curve) qui a pris différentes définitions depuis les années 90 (78). Le plus souvent, cet indice est, de manière similaire à l’AUC/CMI, un rapport entre une aire et la CMI. Cependant, l’AUC calculée pour cet indice n’est pas l’aire totale de 0 à 24 heures mais seulement l’aire entre les temps où les concentrations sont supérieures à la CMI à l’équilibre. Dans certaines conditions, l’AUIC peut être identique à l’AUC/CMI. Par exemple, si l’absorption est rapide ou si l’administration est intravasculaire, les concentrations sont supérieures à la CMI dès le temps 0 et si, en plus, l’élimination est lente, les concentrations restent au-dessus de la CMI pendant toute la durée de l’intervalle d’administration. Compte-tenu de la ressemblance de l’AUIC avec l’AUC/CMI et de l’absence de définition précise, nous ne nous intéresserons plus à cet indice par la suite.
La pertinence des trois indices PK/PD les plus communément décrits (AUC/CMI, Cmax/CMI et
T>CMI) pour prédire l’efficacité des fluoroquinolones a été testée in vitro, sur des modèles
animaux et lors d’études cliniques.
Lors d’infections expérimentales de souris traitées avec une fluoroquinolone, la gatifloxacine, les indices AUC/CMI et Cmax/CMI ont montré une forte corrélation avec la guérison
microbiologique (figure 10) (8).
Figure 10 : Nombre de bactéries versus AUC/CMI, Cmax/CMI et T>CMI
dans les cuisses de souris traitées après 24 heures d’exposition à la gatifloxacine. L’AUC/CMI et le Cmax/CMI sont
corrélés avec la réduction du nombre de bactéries. D’après (8).
D’autres études ont été menées en parallèle pour déterminer des valeurs de ces indices capables de garantir une bonne efficacité des fluoroquinolones.
1.3.3.2 Pertinence et valeurs seuils de l’indice AUC/CMI
1.3.3.2.1 Etudes sur modèles animaux
Une valeur d’AUC/CMI de 100 heures, c’est-à-dire des concentrations en moyenne 4 fois supérieures à la CMI pendant 24 heures, a été nécessaire pour obtenir 100% de survie chez des souris infectées par Klebsiella pneumoniae et traitées avec de la gatifloxacine (59). L’AUC/CMI a aussi été corrélée à la réduction du nombre de bactéries lors de traitements de pneumonies chez la souris avec des fluoroquinolones (93, 94). Lors d’infections de cuisses ou de poumons de souris immunodéprimées, le nombre de bactéries (Staphylocoques, Streptocoques et Entérobactéries) a été maintenu constant avec des valeurs d’AUC/CMI comprises entre 16 et 70 heures. La valeur d’AUC/CMI associée à une réduction du nombre de bactéries d’un facteur 10 chez des souris infectées par Mycobacterium tuberculosis et traitées avec diverses fluoroquinolones a été de 100 à 150 heures (94). Il est aussi intéressant de remarquer que le système immunitaire a un rôle important puisque l’AUC/CMI nécessaire pour obtenir un même effet était 2 fois plus faible pour des souris immunocompétentes que pour des souris immunodéprimées (8).
1.3.3.2.2 Etudes cliniques
Chez l’homme, lors du traitement d’infections bactériennes par des fluoroquinolones, il a été montré que l’indice AUC/CMI était corrélé positivement à la guérison microbiologique (7, 8, 29, 42). La guérison clinique des patients, quant à elle, semble dépendre de nombreux autres facteurs dont l’âge et le site d’infection. Une valeur seuil d’AUC/CMI de 125 heures a été déterminée pour obtenir la guérison microbiologique de patients gravement infectés (figure 11) (42).
Figure 11 : Impact de l’AUIC sur l’éradication bactérienne
en fonction de la durée de traitement. Le nombre de patients par groupe (en fonction de l’AUIC) avec une éradication bactérienne est représenté. Pour des valeurs d’AUIC supérieures à 125, la guérison microbiologique
est supérieure à 80%. D’après (42).
Une autre valeur d’AUC/CMI de 87 heures a permis la guérison microbiologique de patients atteins d’infections nosocomiales à Streptococcus pneumoniae (29). De manière comparable aux données obtenues sur modèles animaux, il semble que les valeurs seuils d’AUC/CMI
soient inférieures lorsque le système immunitaire est fonctionnel. En effet, la guérison de patients immunocompétents infectés par Streptococcus pneumoniae a été obtenue avec des AUC/CMI de seulement 34 heures (7).
1.3.3.2.3 Expériences in vitro
In vitro, l’indice AUC/CMI est également corrélé aux effets des fluoroquinolones sur les
inoculums bactériens (41, 69). Lors de l’exposition in vitro de Streptococcus pneumoniae à des fluoroquinolones autres que la ciprofloxacine, qui est moins efficace, les valeurs d’AUC/CMI nécessaires pour éradiquer des inoculums bactériens de 106 à 108 UFC/mL étaient comprises entre 30 et 60 heures (58, 63, 65, 106). Bien que ces valeurs d’AUC/CMI soient inférieures à celles recommandées pour la guérison microbiologique de patients, les modèles in vitro restent pertinents pour étudier les relations entre les AUC/CMI et l’efficacité des fluoroquinolones. De plus, il a été montré que ces relations entre l’AUC/CMI et l’efficacité étaient identiques pour différentes bactéries exposées à un même antibiotique mais variaient en fonction de l’antibiotique. Par conséquent, l’AUC/CMI est aussi utilisée pour comparer différentes fluoroquinolones in vitro (40).
D’après les résultats obtenus in vitro et in vivo, la corrélation entre les AUC/CMI et les effets des fluoroquinolones semble être bien établie. Cependant, il a été décrit que l’indice Cmax/CMI pouvait être dans certains cas plus prédictif de l’activité des fluoroquinolones (27).
1.3.3.3 Pertinence et valeurs seuils de l’indice C
max/CMI
1.3.3.3.1 Etude sur modèles animaux
Une étude sur des rats septicémiques a montré que l’indice Cmax/CMI était plus prédictif de la
survie que l’indice AUC/CMI lorsque la valeur de Cmax/CMI était supérieure à 10 (27). Une
valeur de Cmax/CMI égale à 10.6 a également permis d’obtenir 100% de survie sur des souris
infectées par Streptococcus pneumoniae et traitées avec de la ciprofloxacine (97).
1.3.3.3.2 Etudes
cliniques
Une étude prospective sur les effets de différentes doses de lévofloxacine a montré qu’une valeur de Cmax/CMI supérieure à 12.2 permettait de prédire une guérison clinique et
microbiologique (89). Dans cette étude, 82 % des patients étaient exposés à des Cmax/CMI
En résumé, les indices AUC/CMI et Cmax/CMI sont capables de prédire l’activité des
fluoroquinolones. Ces indices représentent l’intensité de l’exposition à l’antibiotique ce qui est en accord avec le mode d’action « concentration-dépendant » des fluoroquinolones. Par ailleurs, l’AUC et le Cmax sont proportionnels à la dose pour les substances qui ont des
cinétiques linéaires et, pour des administrations uniques journalières, l’augmentation de la dose conduit à une augmentation proportionnelle de l’AUC/CMI et du Cmax/CMI. Dans la
majorité des études, il est donc assez difficile de discriminer quel indice est le plus représentatif de l’activité. Seules des études plus nombreuses menées avec des doses fractionnées associées à la même AUC sur 24 heures mais à un Cmax divisé par le nombre
d’administrations journalières permettraient d’identifier quel est, de l’AUC/CMI ou du Cmax/CMI, l’indice le plus pertinent.
1.4 Résistances aux fluoroquinolones
1.4.1 Mécanismes de tolérance
La tolérance est un mécanisme particulier de survie en présence de l’antibiotique qui se distingue de la résistance par le fait que la sensibilité des bactéries aux fluoroquinolones, en termes de valeur de CMI, n’est pas altérée.
La présence de bactéries tolérantes ou persistantes a été pour la première fois décrite par J. Bigger en 1944 quand il a remarqué que la pénicilline ne pouvait pas complètement stériliser une culture de Staphylococcus aureus in vitro. Les bactéries persistantes qui ne sont pas tuées par l’antibiotique ne portent pas de résistantes génétiques et restent sensibles à cet antibiotique si elles sont remises en culture dans du milieu frais (9, 60 ). En effet, leur CMI est inchangée et lorsque ces bactéries sont mises en croissance et exposées de nouveau à un antibiotique, la vitesse de bactéricidie est identique à celle observée sur la population originale. En phase de croissance exponentielle, les bactéries persistantes sont rares alors que leur nombre augmente dès qu’elles entrent en phase stationnaire (55). Le phénomène de persistance est probablement un mécanisme de protection pour la bactérie dans des conditions de stress, de privation ou d’exposition aux antibiotiques. L’impact in vivo de ces bactéries persistantes est très important au niveau des biofilms ou lorsque le système immunitaire est incompétent et ne peut pas éliminer ces quelques bactéries qui persistent après l’action de l’antibiotique (61).
Bien que le phénomène de persistance ait été découvert dans les années 40, le mécanisme expliquant la présence de ces bactéries a été peu étudié. Les hypothèses émises pour expliquer ce phénomène sont que ces bactéries sont à un stade de division cellulaire sur lequel l’antibiotique ne peut pas agir ou qu’elles sont dans un état dormant. La première étude moléculaire a été menée par Moyed et al. (79) qui ont mis en évidence un gène porté par Escherichia coli appelé hipA (high persistence) dont la mutation induit une augmentation des bactéries persistantes de 10 à 1000 fois. La protéine HipA forme avec HipB un complexe toxine-antitoxine (TA) où la toxine HipA est régulée par le répresseur HipB. Balaban et al. (9) ont mis en évidence par l’étude de la croissance des mutants hipA7 dans des microfluides que les bactéries persistantes constituaient une population préexistante générée principalement en phase stationnaire. Il a été montré récemment que l’activité sérine kinase de la protéine HipA était à l’origine de la formation des bactéries persistantes (19). Cependant, le gène hipA n’étant pas présent sur toutes les bactéries présentant des phénomènes de persistance, il ne peut pas expliquer à lui seul la présence de bactéries persistantes. Une implication de complexes TA autres que hipBA tels que relBE et mazEF
inhibant la synthèse de peptidoglycanes, d’ADN ou d’ARN a par ailleurs été démontrée. D’autres mutants, les hipQ, persistent en présence de norfloxacine. Ces bactéries persistantes, dites de type II, ont une croissance faible mais non nulle et leur nombre dépend du nombre total de bactéries (9).
D’autres protéines toxiques produites par un plasmide telles que des protéines du système chaperon peuvent aussi être responsables de la formation de bactéries persistantes ce qui remet en cause la théorie des modules TA (100). Récemment, il a aussi été découvert que le gène phoU était impliqué dans la formation de bactéries persistantes et que son inactivation était responsable d’une augmentation de la sensibilité à de nombreux antibiotiques et au stress (62). Cette protéine présente dans de très nombreuses espèces bactériennes pourrait donc expliquer le phénomène de persistance bien que le mécanisme de suppression de l’activité métabolique cellulaire ne soit pas encore élucidé.
1.4.2 Mécanismes de résistance
La perte d’efficacité d’un antibiotique, qui se traduit par une résistance de la bactérie à cet antibiotique et donc à une diminution de sa sensibilité à l’antibiotique (c’est-à-dire à une augmentation de la CMI), peut être due soit à une mutation de ses cibles soit à une diminution de sa concentration intrabactérienne (figure 12). Dans la suite de ce paragraphe, le terme « résistance faible » fait référence à une modification de la CMI d’un facteur 2 à 4 par rapport à la souche parente et le terme « résistance élevée » fait référence à une multiplication de la CMI par un facteur supérieur à 4.
AcrB gyrB gyrA ParE ParC Porine Pompe à efflux Topoisomérase IV Gyrase AcrA TolC
Figure 12 : Description des mécanismes de résistances aux fluoroquinolones (cibles, pompes à efflux, porines) d’une bactérie à Gram négatif (soulignés sur le schéma).
1.4.2.1 Mutations sur les gènes codant pour les cibles
Pour Escherichia coli et les autres bactéries à Gram négatif, la gyrase est la cible préférentielle et la topoisomérase IV est la cible secondaire pour la plupart des fluoroquinolones. Des mutations sur les gènes gyrA ou gyrB codant pour la gyrase entraînent le franchissement d’un premier palier de résistance chez les bactéries à Gram négatif (80). Les mutations sur les gènes parC et parE codant pour la topoisomérase IV ne provoquent une diminution de la sensibilité que lorsque la gyrase a été préalablement modifiée (51). L’ordre des mutations conférant des résistances est inversé pour les bactéries à Gram positif pour lesquelles la première mutation est le plus souvent sur la topoisomérase IV et la deuxième sur la gyrase.1.4.2.1.1 Mutations sur gyrA
Les points de mutations conférant des résistances aux fluoroquinolones se regroupent dans des régions appelées QRDR (Quinolone-Resistance Determining Region). Pour la sous-unité GyrA, cette région chez Escherichia coli est située entre les acides aminés 67 et 106 qui sont très proches de l’acide aminé Tyr122 responsable de la liaison covalente temporaire aux groupements phosphates de l’ADN au cours de l’activité gyrase. Il semblerait que ces modifications d’acides aminés altèrent le site de fixation des fluoroquinolones au complexe ADN-gyrase et que la résistance soit due à cette diminution d’affinité pour ce complexe. La substitution d’un nucléotide sur le codon 83 est la mutation la plus couramment observée sur
gyrA (34, 105). Les bactéries les plus résistantes aux fluoroquinolones présentent souvent
une substitution d’une sérine par une leucine. Les acides aminés Trp, Ala et Val ont aussi été retrouvés sur cette position suite à la substitution d’un nucléotide (51, 91). Des mutations sur le codon 87 sont aussi plus rarement observées sur gyrA (51, 84, 91). Les souches présentant une de ces substitutions sur le codon 87 ont une résistance moins élevée que celles avec une seule mutation sur le codon 83 (84). L’association de 2 substitutions de codons sur gyrA en 83 et 87 conduit à des résistances élevées (34). D’autres mutations de
gyrA sur des positions différentes dans la QRDR ont aussi été observées beaucoup plus
rarement. Pasquali et al. (85) ont obtenus des mutants Gly81Cys et Ser83Leu en exposant
Escherichia coli ATCC25922 à des concentrations suboptimales, c’est-à-dire dans la fenêtre
de sélection (voir paragraphe 1.4.4.2), d’enrofloxacine et de ciprofloxacine. Une seule mutation, une substitution Ala196Glu a été observé hors de la QRDR (105). Ces mutations confèrent des résistances moins élevées que celles sur les codons 83 et 87 (105). La place du codon modifiée a une importance dans le niveau de résistance conféré par la mutation. Cependant, l’acide aminé qui intervient après la mutation a aussi une influence notoire car ses groupements et ses charges électriques conditionnent l’interaction des quinolones avec le complexe ADN-gyrase par la formation de liaisons hydrogènes et de liaison ioniques (91).
1.4.2.1.2 Mutations sur parC
Pour les bactéries à Gram négatif, les mutations sur les gènes codant pour la topoisomérase IV n’entraînent une diminution de la sensibilité que si elles sont associées à une première mutation sur le gène gyrA. Ces mutations supplémentaires entraînent un haut niveau de résistance aux fluoroquinolones (51). Du fait des homologies entre les sous-unités de la topoisomérase IV et de la gyrase, les codons 83, 81 et 87 de gyrA d’Escherichia coli correspondent aux codons 78, 80 et 84 de parC. La substitution la plus courante dans la sous-unité ParC est Ser80Ile (34, 91, 101) suivie par la substitution Ser80Arg. Il a aussi été observé des substitutions de l’acide aminé Glu84 par Lys, Val et Gly (34, 91). Les changements d’acide aminé en position 80 réduisent les possibilités de liaisons hydrogènes alors que les substitutions en position 84 entraînent la perte de charges négatives (91, 101). D’autres mutations sur parC beaucoup plus rares ont été observées dans et hors de la QRDR (91).
1.4.2.1.3 Autres
mutations
Les mutations sur gyrB et parE sont rarement observées et confèrent des résistances bien moins élevées que celles dues à des mutations sur gyrA et parC.
1.4.2.2 Surexpression de pompes à efflux
chromosomiques
Les résistances dues à une baisse des concentrations intrabactériennes proviennent de variations d’expression des protéines de transports membranaires telles que les porines et les pompes à efflux. Les porines permettent aux fluoroquinolones de franchir la membrane externe des bactéries à Gram négatif pour entrer dans la cellule. Les pompes à efflux, à l’opposé, font sortir l’antibiotique de la cellule par un transport actif. Les variations de l’expression de ces deux transporteurs peuvent être dues à des modifications de transcription de l’opéron mar (multiple antibiotic resistance) codant pour MarA (activateur), MarB, MarC et MarR (répresseur) qui induisent un phénotype mar. Des composés contenant des noyaux phénol tels que les fluoroquinolones, des mutations sur le gène marR ou des régulateurs de transcription peuvent diminuer l’activité répressive de MarR. MarA est alors surexprimé et entraîne la transcription du gène micF qui produit un ARN anti-sens de l’ARN qui code pour OmpF, une des 2 porines d’Escherichia coli. Cependant, la diminution de la sensibilité aux fluoroquinolones associée à une mutation sur marA n’est que partiellement due à une diminution des porines à la surface des bactéries (18). En effet, MarA induit aussi la transcription des gènes acrA et acrB codant pour les sous-unités d’une pompe à efflux (4, 82). La diminution des concentrations intrabactériennes en antibiotique sans mutation