présentée en vue de l’obtention du titre de Docteur de l’Université de Toulouse délivré par
l’Université Toulouse III – Paul Sabatier
Spécialité : Neurosciences
Bases neurales des comportements orientés vers un but :
Étude des corrélats de l’activité unitaire préfrontale et
hippocampique dans une tâche de navigation
par
Vincent
H
OK
Soutenue le 19/12/2007 devant la commission d’examen composée de :
MM. M. Giurfa Université de Toulouse Président
R. Jaffard Université de Bordeaux Rapporteur
J.-M. Lassalle Université de Toulouse Co-directeur de thèse B. Poucet Aix-Marseille Université Directeur de thèse
présentée en vue de l’obtention du titre de Docteur de l’Université de Toulouse délivré par
l’Université Toulouse III – Paul Sabatier
Spécialité : Neurosciences
Bases neurales des comportements orientés vers un but :
Étude des corrélats de l’activité unitaire préfrontale et
hippocampique dans une tâche de navigation
par
Vincent
H
OK
Soutenue le 19/12/2007 devant la commission d’examen composée de :
MM. M. Giurfa Université de Toulouse Président
R. Jaffard Université de Bordeaux Rapporteur
J.-M. Lassalle Université de Toulouse Co-directeur de thèse B. Poucet Aix-Marseille Université Directeur de thèse
Le travail réalisé au cours de cette thèse, a été effectué sous la direction de Bruno Poucet, au sein du Laboratoire de Neurobiologie de la Cognition (UMR 6155, Aix-Marseille Université – CNRS). Cette thèse a été financée par le Minis-tère de l’Enseignement Supérieur et de la Recherche et par la Fondation pour la Recherche Médicale (FDT-2006-1208711).
Mea culpa, je n’ai pas toujours fait preuve d’un sens aigu de la diplomatie, ni de la demi-mesure (“Une intro générale ? Ben si, elle est là. Oui, elle fait une page recto-verso, mais l’essentiel est dedans. . . Non ?”). Ce côté “brut de décoffrage”, agrémenté peut-être d’un poil de paresse, se ressentira probablement à la lecture de ce manuscrit : trente pages d’anatomie pouvant laisser une impression d’indigestion. Je tiens donc à remercier les membres du jury qui ont bien voulu lire et juger ce travail de thèse.
Je revois encore Bruno me demandant quand je comptais venir à Marseille pour débuter les expériences. J’étais alors en DEA, nous étions au mois de novembre, pas tout à fait à la fin des cours, mais déjà avec les examens à préparer. Tout naturellement, je lui ai répondu : janvier, avec une petite intonation interrogative pleine d’espoir, précédée d’un “euh” trahissant trop vivement mon angoisse . . . Il a eu, sans aucun doute, raison d’insister pour que je commence les expériences dès décembre. Mais si je ne devais te remercier que d’une seule chose, Bruno, ce serait de ton enthousiasme sans faille. Et. . . ta disponibilité aussi. Ta franchise. Et ta patience (mise à rude épreuve, face à mon scepticisme que même certains referees de mauvaise foi n’oseraient pas afficher).
Étienne a sa part de responsabilité dans cette entreprise, il faut qu’il en assume ici même les conséquences : merci de ta disponibilité (ce n’est pas un lieu commun, mais juste un point commun, et j’ai même cru comprendre que ce n’était pas si commun que ça d’ailleurs), ton humour (Marie appelle ça un “humour de papa”, je n’ai jamais vraiment compris le sens exact de cette expression, mais je crois que c’est positif), ta capacité à identifier clairement les failles d’un protocole, d’un raisonnement, d’un manuscrit (en clair, si Étienne vous dit : “je ne suis pas convaincu que ce soit une très bonne idée. . .”, passez à autre chose, c’est voué à l’échec).
Je pense avoir eu beaucoup de chance de travailler dans ce laboratoire. Bon, d’accord, pour l’environnement scientifique, c’est vrai, mais surtout, j’étais content de venir travailler le matin (je ne tolérerai aucun commentaire sur ce sujet), peut-être tout simplement parce qu’il y règne une excellente ambiance, et que chacun y contribue à hauteur de son degré d’ébriété chronique (pas uniquement, mais ça compte quand même un peu).
Un grand merci à ceux qui permettent que tout (ou presque) soit possible : Dany, Rémy, Luciana (qu’est ce qu’on ferait sans toi ?) et Riri. Merci de votre disponibilité et de votre gentillesse.
J’ai une pensée toute particulière pour celles et ceux qui nous ont quittés en cours de route pour des cieux plus cléments : Louis et Audrey (Mont-réal), Sébastien (Freiburg), Cécile (Washington), Vietminh (Bordeaux), Carole (Marseille ; oui, il faut avouer que c’est une ville qu’on ne quitte pas facile-ment), Bénédicte et Julien (Bourg-en-Bresse). J’aimerais aussi remercier tous les membres de la dream team de m’avoir accepté dans leur bureau de mai à septembre 2005, alors que je ne voulais pas quitter la SOGERES (j’avais peur de ne pas avoir assez au restaurant universitaire, finalement, j’en ai vraiment eu assez).
Un grand merci également à Francesca, Sébastien et Marie de votre lecture attentive des premières versions de ce manuscrit, et puis, tout simplement, de m’avoir consacré un peu de votre temps, qui, comme chacun sait, est élastique (sous certaines conditions néanmoins). Merci encore une fois à Sébastien, Sylvie et Nathalie de leurs précieux conseils sur les techniques d’injection ; les résultats auraient été encore plus étonnants sans leurs suggestions. Merci à Boris de ses conseils éclairés en matière d’analyse et de traitement des signaux continus (même si j’ai encore un peu de mal à comprendre comment une moyenne de sinusoïdes décalées dans le temps. . .). Merci à Brian de ta rigueur et de ton esprit critique, tous deux d’une extrême préciosité. Merci à Alexandrine d’avoir accepté (mais le savais-tu seulement au début ?) de passer le plus clair de ton temps enfermée, seule, dans la pénombre, avec pour seuls compagnons tes adorables petits rats. Ton calvaire ne sera pas vain, je te le promets. Merci également à Paul, Alice, Tiffany et Corentin, Sophie et Xavier, Stéphane et Lætitia, Alexandre et Jean-Baptiste, Bjørg, Anna et Andrea, Elisa, Frédéric et Virginie, Sylvie, Sébastien et Amélie, Francesca, Barbara, Christelle et Paolo, Marianne, Sabrina, Tiphaine pour les bons moments passés en votre compagnie. Vous avez très largement contribué à ce que j’aie envie de rester un peu plus. . . et bien plus encore.
Marie a sacrifié ses trois semaines de vacances pour me voir travailler sur les étapes finales de la rédaction de ce mémoire. Quelques années auparavant, Marie a accepté sans sourciller que je parte de Tours pour poursuivre mes études à Toulouse. Bien des années après, Marie s’est résignée à vivre à plus de sept cents kilomètres de moi, lorsque je suis parti m’installer à Marseille, alors qu’elle habitait Paris (Montréal, Québec, Ottawa, Vienne, rayer les mentions inutiles, cela fera toujours trop loin). Aujourd’hui, Marie a, sans hésitation, décidé de me rejoindre à Dublin. Je ne sais pas trop comment te remercier de tout cela, tout ce que je peux faire, c’est te dire combien je t’aime, et combien tu comptes pour moi.
Remerciements iii
Liste des tableaux ix
Table des figures xi
Résumé xiii
Abstract xv
Introduction xvii
I
Mécanismes Neuronaux de la Cognition Spatiale
1
1 Neuroanatomie 3
1.1 Définition de la région hippocampique . . . 3
1.2 Localisation de la formation hippocampique et de la région parahippocampique . . . 3
1.3 Connexions du gyrus dentelé . . . 4
1.3.1 Connexions au sein du gyrus dentelé . . . 4
1.3.2 Entrées et sorties du gyrus dentelé . . . 6
1.4 Connexions de l’hippocampe . . . 6
1.4.1 Connexions au niveau CA3 . . . 8
1.4.2 Connexions au niveau CA1 . . . 8
1.5 Connexions du subiculum . . . 9
1.5.1 Connexions hippocampique et parahippocampique . . . 10
1.5.2 Connexions afférentes . . . 10
1.5.3 Connexions efférentes . . . 10
1.6 Cortex entorhinal . . . 12
1.6.1 Connexions avec la formation hippocampique . . . 12
1.6.2 Connexions avec la région parahippocampique . . . 15
1.6.3 Connexions extrinsèques, entrées et sorties . . . 15
1.7 Cortex périrhinal et postrhinal . . . 18
1.7.1 Connexions hippocampiques et parahippocampiques . . 18
1.7.2 Connexions extrinsèques . . . 19
1.8 Cortex préfrontal . . . 19
1.8.1 Connexions du cortex préfrontal . . . 20
1.8.2 Cortex préfrontal médian . . . 21
2 Activité électrophysiologique spatio-sélective 29 2.1 Cellules de lieu . . . 29
2.1.1 Propriétés des cellules de lieu . . . 29
2.1.2 Caractéristiques et distribution des champs d’activité . . 30
2.1.4 Remapping . . . 34
2.2 Cellules de direction de la tête . . . 36
2.2.1 Propriétés des cellules de direction de la tête . . . 37
2.2.2 Circuit des cellules de direction de la tête . . . 38
2.2.3 Influence des informations sensorielles sur l’activité des cellules de direction de la tête . . . 40
2.2.4 Relations entre les cellules de direction de la tête et les cellules de lieu hippocampiques . . . 41
2.3 Grid cells du cortex entorhinal . . . 41
2.3.1 Propriétés des grid cells . . . 43
2.3.2 Influence des informations sensorielles sur l’activité des grid cells . . . 44
2.3.3 Grid cells et remapping hippocampique . . . 44
3 Navigation : physiologie et modélisation 51 3.1 Corrélats comportementaux des cellules de lieu . . . 51
3.1.1 Tâches multiples ou séquentielles . . . 51
3.1.2 Effet attentionnel sur l’activité des cellules de lieu . . . . 55
3.1.3 Influence des stratégies comportementales . . . 57
3.1.4 Cellules de lieu et codage du but . . . 58
3.2 Cortex préfrontal et corrélats comportementaux . . . 64
3.2.1 Tâches multiples ou séquentielles . . . 64
3.2.2 Cellules du cortex préfrontal et comportement orienté vers un but . . . 65
3.3 Carte cognitive et navigation topologique . . . 67
3.3.1 Concepts . . . 67
3.3.2 Planification de trajet et modèle distribué . . . 70
II
Contribution Expérimentale
77
4 Activité liée au but des neurones du cortex préfrontal 79 4.1 Introduction . . . 794.2 Matériels et méthodes . . . 81
4.2.1 Sujets . . . 81
4.2.2 Fabrication des plots d’électrodes . . . 82
4.2.3 Comportement . . . 82
4.2.4 Chirurgie . . . 83
4.2.5 Technique d’enregistrement . . . 84
4.2.6 Analyses des données . . . 85
4.2.7 Caractéristiques numériques . . . 85
4.3 Résultats . . . 86
4.4 Discussion . . . 87
Publication # 1 89
5 Activité liée au but des cellules de lieu 113 5.1 Introduction . . . 113 5.2 Résultats . . . 114 5.3 Discussion . . . 116 Publication # 3 119 Publication # 4 133 6 Interactions fronto-hippocampiques 147 6.1 Introduction . . . 147 6.2 Matériels et méthodes . . . 148 6.3 Résultats . . . 150 6.3.1 Observations générales . . . 150 6.3.2 Résultats comportementaux . . . 150
6.3.3 Analyse des caractéristiques électrophysiologiques . . . 150
6.3.4 Analyse des caractéristiques spatiales . . . 151
6.3.5 Analyse de la décharge liée au but . . . 154
6.4 Discussion . . . 159
III
Discussion Générale
161
Bibliographie 169
4.1 Caractéristiques principales des neurones enregistrés dans le cortex cingulaire antérieur dorsal et les aires prélimbique et infralimbique . . . 86 6.1 Analyse des variables comportementales dans la tâche de
navi-gation continue . . . 150 6.2 Caractéristiques électrophysiologiques principales des cellules
pyramidales et des interneurones dans la tâche de navigation continue . . . 151 6.3 Caractéristiques spatiales de la décharge des cellules de lieu
1.1 Organisation tridimensionnelle de la région hippocampique . . 5
1.2 Couches de la formation hippocampique . . . 7
1.3 Projections du subiculum . . . 11
1.4 Organisation topologique de la voie perforante . . . 13
1.5 Projections vers LEA et MEA . . . 14
1.6 Connexions de la région parahippocampique . . . 15
1.7 Distribution des entrées vers le LEA et MEA . . . 17
1.8 Cortex préfrontal chez le rat . . . 21
1.9 Projections cortico-sous-corticales du cortex préfrontal . . . 22
1.10 Projections principales des aires IL et PL . . . 24
1.11 Principales voies excitatrices de la région hippocampique . . . 27
2.1 Cartes d’activité spatiale de cellules pyramidales et thêta . . . 31
2.2 Remapping des cellules de lieu . . . 35
2.3 Rate remapping et global remapping . . . 37
2.4 Exemple de cellule de direction de la tête. . . 38
2.5 Anatomie du système des cellules de direction de la tête . . . . 39
2.6 Intégration de trajet angulaire . . . 41
2.7 Cellules de direction et cellules de lieu . . . 42
2.8 Caractéristiques de l’activité spatiale des grid cells . . . 45
2.9 Patterns d’activité des neurones du MEA . . . 46
2.10 Résumé des connexions des structures spatio-sélectives . . . 50
3.1 Dispositif utilisé dans l’expérience de Wood et al. (2000) . . . . 53
3.2 Cellules de lieu et tâches séquentielles . . . 54
3.3 Champs d’activité dans le labyrinthe aquatique circulaire . . . 59
3.4 Indépendance de l’activité vis-à-vis du comportement instantané 60 3.5 Dynamique des champs d’activité et apprentissage d’un but . . 62
3.6 Schéma du dispositif de l’expérience de Batuev et al. (1990) . . 65
3.7 Activité des neurones préfrontaux et mémoire de travail . . . . 68
3.8 Concept de représentation topologique . . . 69
3.9 Fusion amer–azimut (PrPh) . . . 71
3.10 Construction d’une carte cognitive . . . 72
3.11 Planification et états stables . . . 73
3.12 Transitions et planification de trajet . . . 73
3.13 Schéma général de l’architecture du modèle de planification . . 74
3.14 Schéma général de l’architecture du modèle de Gaussier et al. (2002) . . . 75
4.1 Activité des neurones préfrontaux dans une tâche d’exploration 80 4.2 Représentation schématique du protocole expérimental de Gra-non & Poucet (1995) . . . 81 4.3 Représentation schématique de la tâche de navigation continue 83
4.4 Cartes d’activité des cellules du cortex préfrontal dans la tâche de navigation continue . . . 87 5.1 Analyse temporelle de l’activité des cellules de lieu dans la zone
du but . . . 115 5.2 Analyse temporelle de l’activité extra-champ pour les tâches
indicée et non-indicée . . . 117 5.3 Analyse temporelle de l’activité de l’activité locomotrice . . . . 117 6.1 Protocole utilisé pour les enregistrements hippocampiques et
inactivation préfrontale . . . 149 6.2 Distribution spatiale des centroïdes des champs d’activité des
cellules de lieu . . . 152 6.3 Fréquence de décharge et sélectivité spatiale . . . 154 6.4 Modulation des fréquences de décharge selon le type de session 155 6.5 Enregistrement simultané d’interneurone et de cellule de lieu . 156 6.6 Enregistrement simultané d’interneurone et de cellule de lieu . 157 6.7 Effet de l’inactivation du cortex préfrontal sur la décharge
b a s e s n e u r a l e s d e s c o m p o r t e m e n t s o r i e n t é s v e r s u n b u t : é t u d e d e s c o r r é l at s d e l’activité unitaire préfrontale et h i p p o c a m p i q u e d a n s u n e t â c h e d e nav i g at i o n
L’hippocampe est aujourd’hui reconnu pour jouer un rôle majeur dans le traitement des informations spatiales. Bien que son rôle ne se limite pas simplement à ce type de traitement, la présence de cellules de lieu, neurones déchargeant lorsque l’animal occupe certaines localisations spatiales, a suscité nombre d’études, visant à déterminer la nature de la représentation spatiale au sein de cette structure. Néanmoins, il n’existe pas de consensus sur l’implication de l’hippocampe dans la genèse des trajets de navigation ou dans le codage des buts spatiaux. D’autre part, du fait de son rôle probable dans les comportements de planification, le cortex préfrontal apparaît comme une structure clé dans la réalisation d’un comportement orienté vers un but. L’objectif des recherches réalisées au cours de cette thèse est donc de tenter de comprendre comment ces deux structures (hippocampe et cortex préfrontal) participent à la genèse des comportements orientés vers un but. Nous montrons, dans une première expérience, que les neurones du cortex préfrontal médian (aires prélimbique et infralimbique) enregistrés chez des animaux réalisant une tâche de navigation, présentent une activité spatio-sélective. Les lieux à forte valeur motivationnelle semblent être particulièrement représentés par cette activité préfrontale.
Dans une seconde expérience, nous montrons que les cellules de lieu enregistrées chez des animaux réalisant la tâche de navigation présentent une activité liée au but, en plus de leur activité spatio-sélective classique. Cette décharge est spécifiquement liée à certaines phases temporelles de la tâche. Ce type de signal, en conjonction avec l’activité des neurones préfrontaux, pourrait servir de base à un système de planification de trajet.
Dans la dernière partie de cette thèse, nous présentons des données récentes concernant les interactions entre l’hippocampe et le cortex préfrontal, et plus particulièrement les conséquences de l’inactivation du cortex préfrontal sur l’activité hippocampique. Les résultats montrent que l’activité des cellules de lieu à l’endroit du but n’est pas d’origine préfrontale. De plus, le cortex préfron-tal n’est pas impliqué dans le contrôle en ligne de ce type de comportement orienté vers un but. Ces derniers résultats confortent l’hypothèse que cette activité préfrontale serait vraisemblablement le résultat d’une trace mnésique formée au cours de l’acquisition de cette tâche comportementale.
Mots clés : hippocampe, cortex préfrontal, cellules de lieu, aires prélimbique et infralim-bique, enregistrements unitaires, rat, comportement orienté vers un but.
n e u r a l b a s e s o f g oa l-directed behaviour: prefrontal and h i p p o c a m pa l n e u r o na l a c t i v i t y i n a p l a c e nav i g at i o n ta s k
The hippocampus is well known to play a key role in spatial information processing as shown by the existence of place cells. Such cells are active when the animal occupies particular locations in particular behavioural contexts. However, little evidence has been found in support of the hypothesis that the hippocampus could be involved in the generation of navigation paths and coding of spatial goals. On the other hand, the prefrontal cortex, in line with its role in planning, could be a key structure in the mechanisms involved in goal-directed behaviour. The objective of the research carried out during this thesis is thus to attempt to understand how these two structures (hippocampus and prefrontal cortex) take part in the emergence of goal-directed behaviour. In a first study, we show that prefrontal cortex neurons recorded while the animal performed a goal-oriented navigation task, display spatio-selective activity, especially at locations with a high motivational value.
In a second study, we show that hippocampal place cells recorded in animals trained in the same goal navigation task do have a goal firing pattern, i.e. out-of-field goal related activity, that is time-locked to particular phases of the task. This kind of signal could serve for path planning in combination with prefrontal neurons activity.
In the last study, we present recent data that deal with the interactions be-tween these two structures in the same task, looking for particular modulations of hippocampal place cells activity after prefrontal inactivation. Our results show that the prefrontal cortex does not trigger hippocampal goal-related ac-tivity and suggest a preferential role of the prefrontal cortex in the acquisition of the task rather than in the on-line monitoring.
keywords: hippocampus, prefrontal cortex, place cells, prelimbic and infralimbic areas, unit recording, rat, goal-directed behaviour.
Suite à la découverte en 1971 des cellules de lieu (O’Keefe & Dostrovsky, 1971), on a longtemps attribué une fonction essentiellement spatiale à l’hippo-campe. Les cellules de lieu sont des neurones pyramidaux sélectivement activés lorsque l’animal se trouve dans certaines régions de l’environnement (champs d’activité). L’organisation spatiale de ces champs d’activité à l’échelle d’une population de cellules de lieu est caractéristique d’un environnement donné (Muller & Kubie, 1987 ; Wilson & McNaughton, 1993). Ces cellules découvertes originellement chez le rat, sont aussi présentes chez l’homme (Ekstrom et al., 2003), chez lequel des lésions du lobe temporal entraînent des amnésies rétrogrades et antérogrades (Scoville & Milner, 2000). On considère aujourd’hui que l’activité de ces neurones hippocampiques est intrinsèquement liée à la mémoire épisodique, puisqu’elle offre un cadre spatio-temporel indispensable à la formation d’une telle mémoire.
Le succès des cellules de lieu comme modèle neuronal de support de la mémoire, est lié à plusieurs facteurs : d’une part, la relative facilité d’accès de cette structure chez le rongeur à diverses techniques d’investigation (lésion, inactivation, électrophysiologie), et, d’autre part, l’extraordinaire activité pa-tente des cellules de lieu. Très récemment (Hafting et al., 2005), une activité spatio-sélective complémentaire a été découverte dans le cortex entorhinal médian, principale structure d’entrée et de sortie de l’hippocampe. Combi-nées à d’autres recherches mettant en évidence une activité spatio-sélective dans d’autres structures sous-corticales (Sharp & Green, 1994), ces données indiquent clairement que l’hippocampe fait partie d’un système beaucoup plus global dédié à la navigation, et plus généralement à la mémoire. De plus, des résultats récents (Jones & Wilson, 2005a,b) suggèrent que l’hippocampe et le cortex préfrontal possèdent une relation privilégiée impliquée dans les processus de mémorisation et de décision. L’objectif des recherches que j’ai réalisées au cours de cette thèse a été de tenter de comprendre comment ces structures participent à la genèse des comportements orientés vers un but.
D’un point de vue strictement neuroanatomique, le cortex préfrontal est particulièrement associé à l’hippocampe ventral, dans lequel il existe également des cellules de lieu, mais dont les champs d’activité sont beaucoup plus larges, plus bruités que dans la partie dorsale (Poucet et al., 1994 ; Jung et al., 1994). Néanmoins, les neurones préfrontaux ne montrent pas de sélectivité spatiale lorsque l’animal explore aléatoirement l’environnement (Poucet, 1997 ; Jung et al., 1998 ; Gemmell et al., 2002). Comme nous le verrons par la suite, mon travail a permis de montrer que ces mêmes neurones présentent une décharge spatialement sélective en différents endroits correspondant aux buts spatiaux de l’animal, lorsque celui-ci doit effectuer une tâche orientée vers un but spatial dans le même environnement (Hok et al., 2005).
Dans le même type de tâche, je montre également que les cellules hippo-campiques présentent une brève activité lorsque l’animal se trouve au but. Je montre qu’une telle décharge ne peut pas être interprétée par le comportement du rat, ni sur la base d’un mécanisme d’estimation du décours temporel (Hok et al., 2007b), mais repose vraisemblablement sur une plus grande excitabilité
de neurones déchargeant de manière synchrone (Hok et al., 2007a). En com-binaison avec les neurones préfrontaux, les cellules de lieu hippocampiques pourraient participer à un réseau neuronal permettant au rat de planifier des trajectoires précises dans l’espace.
Une question naturelle, découlant de ces deux expériences, porte sur la nature du signal hippocampique observé à l’endroit du but. Ainsi, il pourrait s’agir d’une forme modulée d’un signal préfrontal en direction de l’hippocampe transitant par les cortex périrhinal et entorhinal latéral. Pour étudier cette question, j’ai mis en place une expérience permettant d’enregistrer l’activité hippocampique tout en inactivant de manière réversible le cortex préfrontal. Les résultats montrent qu’une telle inactivation provoque une augmentation globale de l’activité des neurones hippocampiques (cellules de lieu et interneurones inclus). Néanmoins, ces résultats suggèrent également que l’activité extra-champ des cellules de lieu à l’endroit du but n’est pas affectée par l’inactivation du cortex préfrontal.
Cette thèse se compose de deux parties. La première partie, allant des chapitres 1 à 3, passe en revue les données expérimentales de la littérature scientifique récente, tandis que la seconde partie, chapitres 4 à 6, couvre l’ensemble de la contribution expérimentale effectuée au cours de cette thèse. Le chapitre 1 présente les données récentes de la neuroanatomie concernant les régions hippocampique et préfrontale, comprenant une description de la connectivité interne de ces structures ainsi qu’un résumé des afférences et efférences de ces différentes régions. Le chapitre 2 se limite à une description des propriétés spatio-sélectives des différents types neuronaux impliqués dans un système global de traitement des informations spatiales. Le chapitre 3 constitue à proprement parler une revue de littérature sur les corrélats spatiaux et comportementaux des cellules de lieu, ainsi que ceux rapportés pour les neurones du cortex préfrontal. Ce chapitre comprend également une brève description d’un modèle biologiquement réaliste permettant d’offrir un cadre cohérent de compréhension et d’investigation des relations entre les différentes structures impliquées dans la navigation.
Le chapitre 4 présente les résultats expérimentaux obtenus sur la base d’enregistrements électrophysiologiques de l’activité du cortex préfrontal dans une tâche de navigation spatiale continue. Le résultat principal de cette étude fait état d’une activité spatio-sélective des neurones préfrontaux qui semble liée à des endroits particuliers de l’environnement (Poucet et al., 2004 ; Hok et al., 2005). Le chapitre 5 traite des corrélats spatiaux et comportementaux des cellules de lieu enregistrées dans la même tâche que précédemment, mettant en évidence une décharge extra-champ à l’endroit du but (Hok et al., 2007a). Cette décharge extra-champ ne peut pas être expliquée sur la base de comportements locomoteurs particuliers, et n’est pas reliée à un mécanisme d’estimation du décours temporel (Hok et al., 2007b). Le dernier chapitre de la contribution expérimentale (chapitre 6) présente mes données les plus récentes sur les interactions fronto-hippocampiques à l’aide d’inactivations du cortex préfrontal, couplées à des enregistrements hippocampiques dans le même type de tâche, et met en avant le rôle putatif du cortex préfrontal dans l’acquisition de ce type de comportement orienté vers un but.
Ce manuscrit se termine par une discussion générale de l’ensemble des résultats, en insistant particulièrement sur les questions expérimentales et théoriques découlant de mes travaux expérimentaux.
M É C A N I S M E S N E U R O N A U X D E L A C O G N I T I O N
S PAT I A L E
1
N E U R O A N AT O M I E
1.1 d é f i n i t i o n d e l a r é g i o n h i p p o c a m p i q u e
La convention utilisée pour décrire les différentes régions hippocampiques est celle de Witter & Amaral (2004). Le terme de région hippocampique com-prend deux ensembles de structures corticales, à savoir la formation hippo-campique (archicortex) et la région parahippohippo-campique. Ces deux ensembles se distinguent avant tout sur la base du nombre de couches cellulaires et sur leur connectivité générale. La formation hippocampique est constituée de trois régions cytoarchitectoniquement distinctes : le gyrus dentelé (DG), l’hippocampe, lui-même divisé en trois sous-régions (CA1, CA2 et CA3) et le subiculum. Ces trois régions de la formation hippocampique possèdent trois couches cellulaires propres à l’allocortex. La connectivité entre ces trois couches est largement unidirectionnelle. Les cellules granulaires du gyrus dentelé pro-jettent via les fibres moussues vers le champ CA3 de l’hippocampe, et bien que les cellules de CA3 envoient des collatérales vers les couches profondes ou polymorphiques du gyrus dentelé, ces axones n’innervent pas les cellules granulaires. Cette unidirectionnalité se retrouve également pour les autres connexions intrinsèques principales (CA3 → CA1 ; CA1 → subiculum) de la formation hippocampique.
La région parahippocampique comprend le cortex entorhinal (EC)1 , les cortex périrhinal (PER) et postrhinal (POR) ainsi que le présubiculum et le parasubiculum2
.
1.2 l o c a l i s at i o n d e l a f o r m at i o n h i p p o c a m p i q u e e t d e l a r é g i o n pa r a h i p p o c a m p i q u e
La localisation tridimensionnelle de la formation hippocampique dans le cerveau du rat est assez complexe (Fig. 1.1). La formation hippocampique ap-paraît comme une structure en forme de C allongé, partant rostro-dorsalement des noyaux septaux du prosencéphale, au-dessus et derrière le diencéphale, et finit caudo-ventralement à la naissance du lobe temporal. Le plus long axe de la formation hippocampique est appelé axe septotemporal (le pôle septal étant localisé dorsalement et rostralement) et l’axe orthogonal est appelé axe
1
Certains auteurs incluent le cortex entorhinal dans la formation hippocampique du fait que la projection des cellules entorhinales vers les cellules du gyrus dentelé via la voie perforante est unidirectionnelle, mais le cortex entorhinal compte plus de trois couches cellulaires et d’autres projections vers le champ CA1 et le subiculum sont réciproques.
2
Le subiculum, le présubiculum (PrS) et parasubiculum (PaS) sont parfois regroupés sous le terme de complexe subiculaire. Une des raisons pour lesquelles Witter et Amaral n’utilisent pas cette dénomination réside dans le fait que le subiculum possède trois couches cellulaires alors que le PrS et le PaS possèdent plus de trois couches ; une autre raison consiste à dire que le subiculum est une structure de sortie de la formation hippocampique, tandis que les deux autres structures constitueraient une voie d’entrée du thalamus antérieur vers la formation hippocampique. Pour une discussion complète de ce point particulier, voir la section Should the subiculum, presubiculum and parasubiculum be grouped together as the “subicular complex ?” de Witter & Amaral (2004).
transverse. Afin de décrire la localisation des différentes régions le long de cet axe transverse, le gyrus dentelé servira de point de référence, définissant ainsi l’extrémité proximale de la formation hippocampique. Par exemple, la portion de CA1 proche de CA2 sera appelée portion proximale, tandis que la portion plus proche du subiculum sera appelée portion distale. Il est moins évident de décrire la position des champs hippocampiques à partir de la vue extérieure de la formation hippocampique, car ceux-ci composent la structure à différents niveaux septotemporaux (Fig. 1.1A et 1.1D). Aux niveaux septaux extrêmes, par exemple, seuls le gyrus dentelé et les champs CA1-3 de l’hippocampe sont présents. La région parahippocampique devrait être envisagée comme la partie la plus caudale et la plus ventrale du manteau cortical, enroulée autour des parties caudales et ventrales de la formation hippocampique (Fig. 1.1B et 1.1C). Ainsi, les champs qui composent la région parahippocampique se trouvent situés caudalement et ventralement par rapport aux autres champs de la for-mation hippocampique. Le présubiculum et le parasubiculum forment le bord le plus médian de ce manteau cortical, dont la partie centrale est constituée par le cortex entorhinal. Les cortex périrhinal et postrhinal forment le bord latéral et caudodorsal externe. La frontière du cortex entorhinal avec les cortex périrhinal et postrhinal est approximativement située à hauteur du sulcus rinal (Fig. 1.1B et 1.1C).
1.3 c o n n e x i o n s d u g y r u s d e n t e l é
Le gyrus dentelé possède trois couches cellulaires, les cellules principales3 étant situées dans la couche granulaire. La couche moléculaire est comparati-vement pauvre en corps cellulaires, tandis que la troisième couche, la couche polymorphique, contient des neurones aux corps cellulaires de grande taille, qui peuvent être confondus avec les neurones de CA3 sur la base de colorations de Nissl classiques4
.
1.3.1 Connexions au sein du gyrus dentelé
Les cellules de la couche granulaire donnent naissance aux fibres moussues qui innervent principalement les cellules de CA3, mais arrosent également les cellules de la couche polymorphique du gyrus dentelé via des collatérales. Au sein de la couche polymorphique, les collatérales innervent plusieurs types d’interneurones ainsi que les cellules moussues. Au niveau de la couche granulaire, les cellules granulaires sont soumises à un certain nombre de contrôles inhibiteurs en provenance des autres couches du gyrus dentelé. Pour un détail de ces boucles inhibitrices au sein du gyrus dentelé, voir la revue de Freund & Buzsáki (1996), et pour une description avancée des feedback de la couche polymorphique vers la couche moléculaire, voir Witter & Amaral (2004) ; Deller (1998).
3
On retrouvera dans la littérature le terme de cellules principales pour désigner l’ensemble des cellules pyramidales de CA1/CA3 ainsi que les cellules granulaires du gyrus dentelé.
4
Cette couche polymorphique est aussi appelée hile ou hilus, et fut également décrite par Lorente de Nó sous le terme CA4, laissant entendre qu’il s’agissait d’une région similaire aux couches des cellules pyramidales de l’hippocampe.
Fig. 1.1 –Représentation schématique de l’organisation tridimensionnelle de la région hippocampique dans le cerveau de rat. A La forme en C de l’hippocampe est représentée en transparence dans le cerveau de rat. L’axe septo-temporal est représenté le long de la structure par une flèche. B Reconstruction en vue latérale de la localisation des cortex périrhinal (PER), postrhinal (POR), et entorhinal (EC). C Reconstruction de la partie caudale de l’hémisphère gauche, montrant la subdivision du cortex entorhinal en aires caudale (CE), médiane (ME), dorsolatérale (DLE), dorsal-intermédiaire (DIE), ventral-intermédiaire (VIE) et amygdaloentorhinale (AE). D Vue tridimensionnelle de la formation hippocampique seule, montrant trois sections horizontales (numérotées de 1à 3) à différents niveaux dorsoventraux (noter particulièrement l’absence du cortex entorhinal à des niveaux de coupe les plus dorsaux, alors qu’il est apparent à des niveaux intermédiaire et plus ventraux), et trois sections frontales (numérotées de 1’ à 3’) à différents niveaux rostrocaudaux. Abréviations : CA1, CA2 et CA3, champs hippocampiques ; DG, gyrus dentelé ; f, fornix ; S, subiculum ; rs, sulcus rhinal. D’après Witter & Amaral (2004).
1.3.2 Entrées et sorties du gyrus dentelé
Le gyrus dentelé ne donne pas d’autres projections extrinsèques que les fibres moussues à destination du champ CA3 hippocampique. L’entrée prin-cipale de cette structure provient du cortex entorhinal via la voie perforante, et le gyrus dentelé ne reçoit aucune autre entrée en provenance de structures corticales. En revanche, le gyrus dentelé reçoit un certain nombre d’entrées sous-corticales, en particulier en provenance des noyaux du septum, la région supramamillaire de l’hypothalamus postérieur, et également plusieurs régions du tronc cérébral telles que le locus coeruleus et les noyaux du raphé. De plus, les cellules pyramidales des champs CA3, et particulièrement celles situées au niveau proximal, ainsi que les cellules pyramidales de CA2, donnent quelques collatérales en direction de la couche polymorphique du gyrus dentelé. 1.4 c o n n e x i o n s d e l’hippocampe
L’hippocampe lui-même peut être subdivisé en deux régions principales, une région proximale (voir Sec. 1.2) possédant des cellules à large corps neuronal, une région distale possédant des corps neuronaux plus petits. Ramón y Cajal désigna ces deux régions sous les termes de région inférieure et région supérieure respectivement. La terminologie employée par Lorente de Nó est néanmoins quelque peu différente, mais est entrée dans l’usage courant. Cet auteur a divisé l’hippocampe en différents champs ammoniques, à savoir CA1, CA2 et CA3. Les champs CA3 et CA2 sont équivalents à la région inférieure de Ramón y Cajal et le champ CA1 est équivalent à la région supérieure. En plus de ces différences cytoarchitectoniques concernant la taille des cellules, les champs CA3 et CA1 se distinguent également sur la base de leur connectivité. Par exemple, les cellules de CA3 reçoivent une entrée importante d’informations en provenance du gyrus dentelé via les fibres moussues, alors que les cellules de CA1 ne sont pas concernées par cette projection.
L’existence du champ CA2 a été l’objet de controverses. Tel que défini à l’origine par Lorente de Nó, CA2 est constitué d’une fine zone de cellules interposées entre CA1 et CA3, possédant des corps cellulaires de grande taille comme CA3, mais non innervées par les fibres moussues à l’instar de CA1. D’autres études montrent que la connectivité de cette région CA2 présente un certain nombre de différences vis-à-vis des autres champs hippocampiques, et bien que certains auteurs pensent le champ CA2 comme une extension de CA3, il apparaît que CA2 se distingue de CA35
.
L’organisation lamellaire des différentes couches cellulaires est globalement la même pour tous les champs hippocampiques (voir Fig. 1.2). La couche principale de cellules est appelée couche des cellules pyramidales (stratum pyramidale). La fine couche située en profondeur de cette couche pyramidale est désignée sous le nom de stratum oriens (cette couche ne contient que très peu de corps cellulaires), et, situé encore en dessous, se trouve l’alveus (cette couche ne contient que des fibres de passage). Au niveau du champ CA3,
5
Pour une discussion complète sur la validité de l’individualisation de CA2, voir la section Connections of CA2 dans Witter & Amaral (2004). Par la suite, je ne parlerai plus de ce champ particulier, la grande majorité des études, tant électrophysiologiques que lésionnelles ou encore théoriques, ne font pas mention particulière de CA2, mais se focalisent sur les champs CA3 et CA1.
Fig. 1.2 –Cette figure présente les différentes régions et couches de la formation hippocampique. Abréviations : ab, faisceau angulaire ; fi, fimbria ; PaS, parasubiculum ; PrS, présubiculum ; ML, GL, et PoDG, respectivement couches moléculaire, granulaire et polymorphique du gyrus dentelé ; so, stratum oriens ; pcl, couche des cellules pyrami-dales ; sl, stratum lucidum ; sr, stratum radiatum ; s l-m. stratum lacunosum-moleculare. D’après Witter & Amaral (2004).
mais pas au niveau CA1 ni CA2, se trouve une couche située au-dessus de la couche pyramidale, appelée stratum lucidum. Cette couche contient les axones de la projection des fibres moussues en provenance du gyrus dentelé et à destination du champ CA3. Dans la terminaison distale de cette couche, on peut observer une densification de la projection des fibres moussues qui marque la délimitation entre les champs CA3 et CA2. Superficiellement au stratum lucidum, au niveau CA3, et immédiatement au-dessus de la couche pyramidale, aux niveaux CA1 et CA2, se trouve le stratum radiatum. Cette couche peut être définie comme la région supra-pyramidale où se trouvent les connexions récurrentes CA3/CA3 et les connexions collatérales de Schaffer de CA3 vers CA1. La partie la plus superficielle de l’hippocampe est appelée stratum lacunosum-moleculare. C’est dans cette couche que se terminent les fibres de la voie perforante en provenance du cortex entorhinal ; on y trouve également d’autres afférences telles que celles en provenance du nucleus reuniens.
Le principal type cellulaire des neurones hippocampiques est représenté par les cellules pyramidales qui se trouvent dans la couche pyramidale de l’hippocampe. Ces cellules pyramidales possèdent un arbre dendritique basal qui s’étend jusque dans le stratum oriens et un arbre dendritique apical qui s’étend jusqu’au niveau de la fissure hippocampique.
1.4.1 Connexions au niveau CA3
Les cellules pyramidales de CA3 donnent naissance à de nombreuses col-latérales qui innervent aussi bien les champs ammoniques de l’hippocampe ipsilatéral (CA3, CA2 et CA1) que ceux de l’hippocampe contralatéral (pro-jections commissurales innervant les mêmes champs que précédemment). Les cellules pyramidales du champ CA3, et plus particulièrement celles situées au niveau proximal, envoient également des projections vers les neurones de la couche polymorphique du gyrus dentelé. En revanche, ces neurones CA3 ne semblent pas envoyer de projection vers le subiculum, présubiculum, parasubiculum, ou encore le cortex entorhinal.
Les projections de CA3 à destination des cellules CA1 (collatérales de Schaffer) sont organisées topologiquement selon l’axe septo-temporal, c.-à-d. que les cellules CA3 proches du gyrus dentelé (proximales) ont tendance à innerver les cellules CA1 situées au niveau septal, alors que les cellules CA3 proches de CA1 (distales) ont tendance à innerver les cellules CA1 situées au niveau temporal. De même, les terminaisons de ces projections sont plus ou moins superficielles au sein du stratum radiatum suivant la localisation proximale/distale des cellules CA1 cibles.
Les projections récurrentes de CA3 (collatérales axoniques reliant les cellules CA3 à d’autres cellules CA3) sont organisées également selon une distribution septo-temporale, c.-à-d. que les cellules CA3 proximales ne communiquent uniquement qu’avec d’autres cellules CA3 proximales situées à un même niveau septo-temporal, tandis que les cellules CA3 plus distales ont tendance à innerver d’autres cellules CA3 suivant un axe transverse et septo-temporal beaucoup plus large.
1.4.2 Connexions au niveau CA1
Quelques interneurones au niveau CA1 projettent de manière importante vers CA3 et le hile du gyrus dentelé. Ces cellules sont situées à la bordure du stratum oriens et de l’alveus, et possèdent des arbres dendritiques horizontaux principalement localisés dans le stratum oriens. Les axones de ces cellules forment des synapses symétriques sur les dendrites des cellules principales et fournissent une information inhibitrice en retour. Néanmoins, la projection principale du champ CA1 est topologiquement organisée en direction du subiculum, c.-à-d. que les cellules CA1 proximales projettent vers la partie distale du subiculum, tandis que les cellules CA1 distales projettent sur la partie proximale. Notons également que les cellules CA1 reçoivent une faible entrée en provenance du subiculum (Commins et al., 2002).
Au contraire de CA3, CA1 ne possède pas de collatérales récurrentes, il existe néanmoins quelques collatérales d’axones de CA1 à destination des cellules du subiculum qui projettent à leur tour sur les dendrites basales d’autres cellules CA1. Parmi les cellules CA1 recevant ces projections, beaucoup d’entre elles sont des interneurones, telles les cellules en panier situées dans le stratum oriens, qui reçoivent des entrées directes des cellules pyramidales CA1, inhibant en retour les cellules pyramidales. Un autre aspect de la connectivité de CA1 contraste avec celle de CA3 : la faible projection contralatérale à travers les fibres commissurales. L’ensemble de ces différences de connectivité entre
CA3 et CA1 laisse penser que ces deux types de champ ammonique présentent des caractéristiques fonctionnelles complètement différentes, un point sur lequel je reviendrai en détail dans la section 2.1.4.
Le champ CA1 reçoit des afférences de structures tant sous-corticales que corticales. Les entrées principales des structures corticales proviennent de la région parahippocampique, tandis que les entrées sous-corticales proviennent du septum ; néanmoins, ces dernières entrées sont moins importantes qu’au niveau du champ CA3. Les deux tiers de CA1 au niveau temporal, et plus particulièrement dans la partie distale, proche du subiculum, reçoivent des entrées en provenance du complexe amygdalaire. En ce qui concerne les entrées thalamiques, notons essentiellement les projections en provenance du nucleus reuniens à destination du stratum lacunosum-moleculare du champ CA1. Ces projections ne passent pas par le trajet classique de la fimbria/fornix mais passent à travers la capsule interne et le faisceau cingulaire. Ces projections se distribuent tout le long de l’axe septo-temporal, mais avec une préférence pour les niveaux intermédiaires. Cette projection est également organisée de manière topologique, la partie dorsale du nucleus reuniens ayant tendance à se projeter vers les parties septales de CA1, tandis que la partie ventrale se projette sur les parties plus temporales. Les fibres de la projection du reuniens forment des synapses excitatrices (asymétriques) tant au niveau des cellules principales que des interneurones.
La région CA1 possède bien plus de projections extrinsèques que la région CA3. De fait, la région CA1 ne projette pas seulement sur l’aire septale latérale, qui possède également des entrées en provenance de CA3, mais aussi sur de nombreuses structures corticales et sous-corticales. Ces projections suivent une organisation générale selon un axe septo-temporal (cortex rétrosplénial, portions dorsales du septum latéral, portions rostrolatérales du nucleus ac-cumbens, noyau de la bande diagonale de Broca pour la partie septale de CA1 ; cortex pédonculé dorsal, aires pré- et infralimbique du cortex préfrontal médian pour la partie plus temporale de CA1).
Les projections en provenance de CA1 à destination du cortex préfrontal médian sont importantes mais moins que celles prenant naissance au niveau du subiculum. Ces projections CA1 innervent de manière diffuse toutes les couches au niveau préfrontal. Ces entrées en direction des aires prélimbique et infralimbique innervent aussi bien les cellules pyramidales que les interneu-rones.
1.5 c o n n e x i o n s d u s u b i c u l u m
La couche de cellules principales du subiculum est constituée de cellules pyramidales dont les dendrites apicales s’étendent jusque dans la couche mo-léculaire et les dendrites basales jusque dans les portions les plus profondes de la couche pyramidale. On y trouve également des neurones à petit corps cellulaire, probablement de type interneurone. D’un point de vue physio-logique, on distingue deux classes de neurones dont le profil de décharge est de type regular firing ou bursting (O’Mara et al., 2001). La localisation de ces deux catégories cellulaires au sein des différentes couches du subiculum n’est pas clairement établie, de plus, une distinction proximale/distale vient complexifier cette tentative de classification. Néanmoins, il semblerait que les
neurones de type bursting soient les seuls à projeter vers le cortex entorhinal. De manière générale, les neurones du subiculum projettent principalement vers le présubiculum, le cortex entorhinal et, dans une moindre mesure, vers la région CA1.
1.5.1 Connexions hippocampique et parahippocampique
Le subiculum reçoit ses principales entrées hippocampiques en provenance de la région CA1. Le subiculum est également fortement innervé par les fibres des cortex entorhinal, périrhinal et postrhinal. Les entrées sont plus faibles en provenance du pré- et parasubiculum. Il n’existe aucune interaction entre le subiculum et le gyrus dentelé ou encore entre le subiculum et les régions CA3/CA2 de l’hippocampe. Au contraire, comme nous l’avons vu plus haut (Sec. 1.4.2), le subiculum envoie de faibles projections vers le champ CA1 innervant aussi bien les cellules pyramidales que les interneurones.
Les projections à destination des pré- et parasubiculum sont très largement réciproques et topologiquement organisées. Le subiculum envoie également des projections à destination des cortex entorhinal, périrhinal et postrhinal. 1.5.2 Connexions afférentes
La nature des afférences corticales à destination du subiculum est controver-sée, notamment en ce qui concerne les afférences du cortex cingulaire antérieur (aire 24) et celles du cortex rétrosplénial. En outre, il apparaît que les structures corticales qui projettent massivement sur le cortex entorhinal ne se retrouvent pas au niveau du subiculum. De fait, aucune preuve d’éventuelles afférences directes en provenance des aires pré- et infralimbique du cortex préfrontal médian à destination du subiculum n’a pu être démontrée.
En revanche, les afférences en provenance des structures sous-corticales sont parfaitement documentées, le subiculum recevant des projections en provenance du complexe amygdalaire, du complexe septal et de structures thalamiques (comme p.ex. le nucleus reuniens).
1.5.3 Connexions efférentes
Le subiculum est une des principales sorties de la formation hippocampique, et innerve un certain nombre de structures tant corticales que sous-corticales (Fig. 1.3). Citons en particulier la projection à destination du cortex préfrontal médian comprenant les parties médiane et ventrale du cortex orbitofrontal ainsi que les aires pré- et infralimbique. Au niveau de l’aire prélimbique et du cortex orbitofrontal, le subiculum innerve principalement les couches profondes de ces structures, tandis que les couches superficielles sont également innervées au niveau de l’aire infralimbique. Il semblerait que le subiculum proximal dans sa partie septale (dorsale) projette préférentiellement vers l’aire prélimbique du cortex préfrontal médian, tandis que la partie temporale (ventrale) projette essentiellement vers l’aire infralimbique. Notons également une projection relativement faible vers le cortex cingulaire antérieur, plus importante pour le cortex rétrosplénial, et modérée vers les cortex périrhinal et postrhinal.
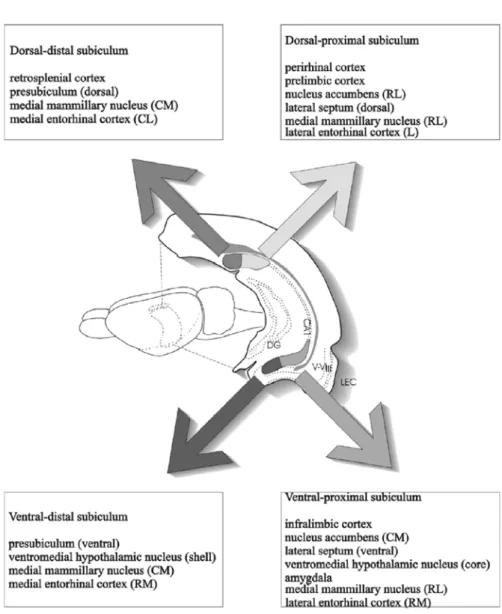
Fig. 1.3 – Schéma représentant l’origine des différentes projections du subiculum en fonction des axes septo-temporal (dorso-ventral) et transverse. Le subiculum peut être subdivisé en quatre régions principales, à savoir : dorsal-distal, dorsal-proximal, ventral-distal et ventral-proximal. DG, gyrus dentelé ; LEC, cortex latéral entorhinal ; V–VI, couches corticales profondes V et VI ; III, couche corticale superficielle III ; CM, caudomédian ; CL, caudolatéral ; RL, rostrolatéral ; RM, rostromédian ; L, latéral. D’après Naber & Witter (1998).
nucleus accumbens, les tubercules mamillaires ainsi que les régions hypothala-miques adjacentes. Les structures thalahypothala-miques, telles que le nucleus reuniens, reçoivent également des projections subiculaires.
1.6 c o r t e x e n t o r h i na l
1.6.1 Connexions avec la formation hippocampique
Le cortex entorhinal est subdivisé en deux sous-régions, à savoir une ré-gion latérale (cortex latéral entorhinal ou LEA) et une réré-gion médiane (cortex médian entorhinal ou MEA). Comme nous le verrons plus loin, les afférences de ces deux sous-structures se distribuent de manière différente sur chacune d’entre elles. Le cortex entorhinal est avant tout la source principale d’infor-mations (via la voie perforante) en direction du gyrus dentelé et l’une des plus importantes en direction de l’hippocampe et du subiculum. Les fibres de cette voie perforante prennent naissance à partir des neurones des couches cellulaires II et III, mais les couches profondes, dans une moindre mesure il est vrai, semblent également y contribuer (Wouterlood et al., 2004). Cette projec-tion est principalement glutamatergique, et se projette de manière strictement unidirectionnelle sur toutes les subdivisions de la formation hippocampique (gyrus dentelé, hippocampe et subiculum, Fig. 1.4).
Il existe néanmoins des différences subtiles dans l’innervation des diffé-rentes composantes de la formation hippocampique. En effet, tandis que les projections à destination du gyrus dentelé et des régions CA3/CA2 naissent des neurones de la couche II du cortex entorhinal, celles à destination de la région CA1 et, dans une moindre mesure, à destination du subiculum, naissent des neurones de la couche III du cortex entorhinal. Une autre différence fon-damentale entre ces deux groupes (gyrus dentelé, CA3/CA2 d’un côté, et CA1/subiculum de l’autre), réside dans le fait que les informations en prove-nance du LEA et du MEA sont mélangées au niveau DG/CA36
mais restent ségrégées selon un axe transverse au niveau de CA1/subiculum (voir Fig. 1.4 et 1.5 pour une vision schématique de l’ensemble).
La région CA1 est également la première région de la formation hippocam-pique à envoyer une projection vers le cortex entorhinal, ce qui la différencie encore de la région CA3 et du gyrus dentelé. Ces projections naissent tout le long des axes septotemporal et transverse, et terminent principalement dans le MEA plutôt que dans le LEA, préférentiellement dans la couche V du cortex en-torhinal. Le subiculum est la deuxième région de la formation hippocampique à renvoyer une information vers le cortex entorhinal, avec une connectivité, là encore, proche de celle de CA1, les projections du subiculum se terminant majoritairement dans la couche V du cortex entorhinal.
6
Bien que cette projection à destination de DG/CA3 ne soit pas différenciée selon l’axe trans-verse, il semble y avoir malgré tout une organisation radiale. En effet, tandis que les projections en provenance du LEA se terminent dans la partie superficielle du stratum lacunosum-moleculare, celles en provenance du MEA se situent plus en profondeur de cette même couche.
Fig. 1.4 –Schéma illustrant l’organisation topologique et laminaire de la projection de la voie perforante vers le gyrus dentelé, l’hippocampe et le subiculum. Le dessin de gauche représente la surface du cortex entorhinal, le sulcus rhinal (non représenté) se trouverait sur la gauche. Le cortex entorhinal a été divisé en aires médiane (MEA) et latérale (LEA). Cette figure montre l’organisation laminaire de la projection de la voie perforante vers le gyrus dentelé, en distinguant le LEA qui se projette superficiellement dans la couche moléculaire et le stratum lacunosum-moleculare, et le MEA qui se projette plus en profondeur. En revanche, en ce qui concerne CA1 et le subiculum, le LEA se projette sur la bordure CA1/subiculum, alors que le MEA se projette à un niveau plus proximal sur CA1 et plus distal sur le subiculum. Les portions situées latéralement au niveau du cortex entorhinal se projettent sur les régions septales de la formation hippocampique, alors que les portions plus médianes se projettent sur des régions temporales. Les projections du présubiculum se terminent exclusivement sur le MEA et les projections du cortex rétrosplénial (RSP Ctx) se terminent sur la partie caudale du MEA. Les entrées principales du cortex entorhinal en provenance du cortex périrhinal se terminent sur les bordures latérales du MEA et du LEA. D’après Witter & Amaral (2004).
Fig. 1.5 – Projections vers les aires médiane et latérale du cortex entorhinal. Les afférences en provenance de ces deux structures sont mélangées au niveau de DG et CA3, mais restent ségrégées au niveau de CA1 et du subiculum. D’après Burwell (2000).
Fig. 1.6 – Représentations aplaties de la région parahippocampique décrivant les connexions intrinsèques (gauche) et interconnexions (droite). Le cortex postrhinal (POR) est représenté en gris foncé, et le cortex périrhinal (aires 35 et 36) est représenté en gris moyen. Le cortex entorhinal (LEA et MEA) est représenté suivant un dégradé de trois gris clairs symbolisant les bandes de projection latérales vers médianes. D’après Burwell (2000).
1.6.2 Connexions avec la région parahippocampique
Le cortex entorhinal envoie des projections vers les cortex périrhinal et postrhinal (Fig. 1.6). Ces projections prennent principalement naissance dans les couches profondes du cortex entorhinal, bien que celles envoyées vers l’aire 35du cortex périrhinal prennent naissance à partir des couches superficielles du LEA. Les connexions les plus fortes semblent avoir pour origine les bords latéral et caudal du cortex entorhinal, c’est-à-dire les parties du LEA et du MEA adjacentes aux cortex périrhinal et postrhinal et qui sont réciproquement connectées à la partie septale de l’hippocampe. Les entrées principales du LEA proviennent majoritairement de l’aire périrhinale 35, alors que celles du MEA proviennent principalement du cortex postrhinal (Burwell & Amaral, 1998). Les projections en provenance des cortex périrhinal et postrhinal se terminent principalement dans la couche III du cortex entorhinal.
Bien qu’il existe des projections en provenance des couches profondes du cortex entorhinal en direction du présubiculum et du parasubiculum, celles-ci sont d’une importance relativement faible comparativement aux projections réciproques, ce qui amènent Amaral et Witter à considérer ces structures comme structures d’entrée au niveau du cortex entorhinal et non l’inverse. 1.6.3 Connexions extrinsèques, entrées et sorties
Connexions corticales afférentes
Le cortex entorhinal reçoit des entrées en provenance de nombreuses struc-tures corticales. Ces entrées peuvent être séparées en deux groupes : celles qui se projettent sur les couches superficielles (I–III), et celles qui se terminent préfé-rentiellement dans les couches profondes (IV–VI). La première catégorie délivre des informations aux neurones entorhinaux ; celles-ci sont ensuite envoyées vers le gyrus dentelé, l’hippocampe et le subiculum. Le deuxième groupe
d’en-trées se projette sur les neurones des couches profondes du cortex entorhinal qui reçoivent des informations en provenance des champs hippocampiques et projettent à leur tour vers des structures corticales7
.
Les principales voies d’entrées des couches superficielles proviennent essen-tiellement des structures olfactives du télencéphale, et plus particulièrement du bulbe olfactif, du noyau antérieur olfactif et du cortex piriforme.
Les afférences corticales des couches profondes du cortex entorhinal pro-viennent d’aires corticales appartenant aux cortex limbique et paralimbique. On compte parmi ces aires corticales : le cortex insulaire agranulaire, qui se distribue préférentiellement sur la partie ventrale du sulcus rhinal (cortex péri-rhinal), le cortex rétrosplénial, qui se distribue dans la portion la plus caudale du MEA, et enfin le cortex préfrontal, tant dans sa composante médiane que orbito-frontale (Fig. 1.7).
Les entrées en provenance du cortex préfrontal à destination du LEA et du MEA composent environ 10% des entrées corticales du système. Néanmoins, la composition de ces entrées est assez hétérogène. Les entrées à destination du LEA incluent les régions médiane et orbitale alors que celles à destination du MEA proviennent majoritairement de la région orbito-médiane. La majorité de ces entrées proviennent de la couche II, bien que les couches plus pro-fondes, telles que les couches V et VI de la région orbito-médiane, contribuent également (Burwell & Amaral, 1998).
Les afférences principales du cortex entorhinal proviennent des deux struc-tures voisines que sont les cortex périrhinal et postrhinal, chacune recevant des afférences de structures corticales extrêmement diverses. Le cortex entorhinal reçoit également des entrées d’autres structures corticales (plus faibles que celles sus-mentionnées) incluant le bulbe olfactif, le noyau olfactif antérieur, le cortex piriforme, le cortex insulaire agranulaire, le cortex préfrontal médian (en particulier les aires prélimbique et infralimbique), les cortex cingulaire anté-rieur et postéanté-rieur (rétrosplénial), le cortex insulaire, le cortex ventro-temporal postérieur et le cortex pariétal postérieur.
En plus de ces connexions afférentes, le cortex entorhinal reçoit des pro-jections de nombreuses structures sous-corticales, incluant le thalamus (en particulier du nucleus reuniens et du nucleus centralis medialis), le complexe septal médian, l’amygdale et l’hypothalamus (en particulier les noyaux su-pramamillaires et l’aire hypothalamique latérale). On trouve également des entrées monoaminergiques en provenance de l’aire tegmentale ventrale, du noyau raphé dorsal et du locus coeruleus.
Connexions sous-corticales afférentes
Le cortex entorhinal reçoit des afférences en provenance de structures sous-corticales également impliquées dans l’innervation des structures hippo-campique et parahippohippo-campique. Une des connexions les plus importantes provient du complexe septal médian, là encore avec une distribution légère-ment différente entre le MEA et le LEA. Le cortex entorhinal reçoit égalelégère-ment
7
Cette vision schématique de l’organisation des couches au sein du cortex entorhinal doit être encore une fois nuancée. Il existe également des connexions allant des couches profondes vers les couches superficielles, si bien qu’une entrée corticale se terminant dans la couche V, p.ex. les projections en provenance du cortex préfrontal, peut également influencer les informations à destination de l’hippocampe (Kloosterman et al., 2003a).
Fig. 1.7 –Distribution des entrées vers les cortex entorhinal latéral (LEA) et entorhinal médian (MEA) en provenance des cortex piriforme, frontal, insulaire, temporal, cingu-laire, pariétal et occipital. Les bandes latérale, intermédiaire et médiane représentent les aires du LEA et MEA projetant sur les parties dorsale, médiane et ventrale de la formation hippocampique. D’après Burwell (2000).
une projection du complexe amygdalaire, en particulier des noyaux latéral, basal, basal accessoire, médian et cortical postérieur. En ce qui concerne les en-trées thalamiques, elles proviennent essentiellement des noyaux central médian et reuniens (RE). Il est intéressant de noter que les neurones du RE projettent non seulement vers le cortex entorhinal, mais aussi vers le champ CA1 hippo-campique et le subiculum. Pour finir, le cortex entorhinal reçoit des projections de nombreuses structures appartenant à l’hypothalamus (noyaux suprama-millaires et tubéromasuprama-millaires, aire hypothalamique latérale, aire tegmentale ventrale et noyau dorsal du raphé) et au tronc cérébral (région pontine). Connexions corticales efférentes
Comme on l’a vu précédemment (Sec. 1.6.1), les principales projections efférentes du cortex entorhinal en direction de l’ensemble des régions de la formation hippocampique proviennent des couches II et III, formant ainsi la voie perforante. Les projections en retour de l’hippocampe se distribuent largement dans la couche V du cortex entorhinal. Les cellules principales de la couche II, les cellules stellaires, sont organisées en amas, tandis que les cellules des autres couches sont organisées de manière plus diffuse.
De manière schématique, les projections corticales efférentes du cortex en-torhinal atteignent les régions corticales limbiques, paralimbiques et olfactives. Ces projections sont principalement dédiées aux cortex périrhinal et postrhinal, en provenance des couches profondes du cortex entorhinal. Les cortex périrhi-nal et postrhipérirhi-nal envoient à leur tour des projections vers les aires sensorielles unimodalitaires et les aires associatives multimodalitaires. De plus, des projec-tions directes vers les aires du cortex olfactif, prélimbique (cortex préfrontal médian), orbitofrontal agranulaire insulaire et rétrosplenial ont également été décrites. Les projections sous-corticales sont destinées majoritairement vers le septum, surtout latéral, l’amygdale et le nucleus accumbens (striatum ventral). Connexions sous-corticales efférentes
A l’image de la formation hippocampique, le cortex entorhinal projette vers la région septale. Ces fibres en provenance du cortex entorhinal se terminent
préférentiellement dans la partie latérale du complexe septal, bien que la partie médiane soit également innervée dans une moindre mesure. On trouve égale-ment des projections très importantes en direction du complexe amygdalaire, notamment vers le noyau basal, les parties médianes du noyau latéral, le noyau basal accessoire et le noyau cortical postérieur. Il est à noter qu’aucune de ces projections ne semble émerger du MEA, toutes prenant naissance dans la couche V du LEA.
Le cortex entorhinal projette également bilatéralement vers le striatum, en particulier dans sa partie ventrale, c.-à-d. vers le nucleus accumbens et les par-ties adjacentes aux tubercules olfactifs. Les projections entre le cortex entorhinal et le nucleus accumbens sont organisées topologiquement, de sorte que les parties médianes du MEA et du LEA projettent sur les parties caudomédianes du nucleus accumbens, tandis que les parties plus latérales du cortex entorhinal projettent sur des parties plus latérales du nucleus accumbens. Finalement, aucune projection du cortex entorhinal vers le thalamus ou le tronc cérébral n’a été décrite dans la littérature.
1.7 c o r t e x p é r i r h i na l e t p o s t r h i na l
La différence entre les cortex périrhinal et postrhinal n’a été établie que très récemment (Burwell et al., 1995). A l’origine, ces deux aires corticales étaient vues comme appartenant à la même structure, à savoir le cortex périrhinal ; cette structure étant composée des aires 35 et 36 (ou encore aire 35 et cortex ectorhinal8
). Les cortex périrhinal et postrhinal, globalement agranulaires, sont, comme leurs noms l’indiquent, situés à proximité de la fissure rhinale. La frontière entre ces deux structures, longtemps sujette à controverse, ne fut clairement définie que très récemment (Burwell, 2000 ; Burwell & Amaral, 1998 ; Burwell et al., 1995).
1.7.1 Connexions hippocampiques et parahippocampiques
Il est clairement établi aujourd’hui que ni le cortex périrhinal ni le cortex postrhinal ne projettent vers CA3 ou le gyrus dentelé (Canning & Leung, 1997 ; Naber et al., 1999). En revanche, ces deux structures donnent naissance à des projections à destination de la région CA1 et du subiculum (Naber et al., 2001a,b), qui possèdent également des connexions réciproques avec le cortex périrhinal et le cortex postrhinal (Kloosterman et al., 2003b). Il est particulière-ment intéressant de noter, là encore, une certaine organisation topologique des projections du PER et du POR, proche de celle observée pour les projections entorhino-hippocampiques. Ces projections terminent préférentiellement dans les deux-tiers dorsaux de l’hippocampe (région septale), les projections en pro-venance du cortex périrhinal innervant le subiculum proximal et la région CA1 distale selon l’axe transverse, tandis que celles du cortex postrhinal innervent le subiculum distal et la région CA1 proximale. L’organisation transverse de ces connexions est donc à rapprocher directement de celles observées entre le cortex périrhinal et le cortex entorhinal latéral d’une part, et le cortex postrhinal et le cortex entorhinal médian d’autre part (Sec. 1.6.1).
Il existe également des connexions directes en provenance de CA1 et du 8
subiculum à destination des cortex périrhinal et postrhinal. Aucune connexion entre les cortex périrhinal et postrhinal d’une part, et le pré- et para-subiculum d’autre part n’a été mise en évidence.
1.7.2 Connexions extrinsèques
Les entrées corticales des cortex périrhinal et postrhinal n’ont été étudiées de manière systématique que très récemment (Burwell & Amaral, 1998). En ce qui concerne le cortex périrhinal, les aires 35 et 36 reçoivent des informations corticales de nature complètement différente, c.-à-d. que l’aire 36 reçoit surtout des informations en provenance du cortex ventro-temporal, alors que l’aire 35reçoit surtout des connexions en provenance des cortex piriforme, latéral entorhinal et insulaire. Pour le cortex postrhinal, les principales entrées, par ordre décroissant d’importance, proviennent des cortex visuel associatif, parié-tal, rétrosplénial et ventro-temporal. De même que pour le cortex périrhinal, les connexions du cortex postrhinal sont très largement réciproques.
Les connexions sous-corticales des cortex périrhinal et postrhinal sont moins bien documentées que les connexions corticales, néanmoins, on sait que ces deux structures possèdent des connexions réciproques avec l’amygdale ainsi que les structures thalamiques.
1.8 c o r t e x p r é f r o n ta l
L’existence d’un cortex préfrontal chez les mammifères non primates, et plus particulièrement chez le rat, fut l’objet de controverses, du fait essentiellement du manque de données anatomiques et fonctionnelles. Une première tentative de définition fut proposée par Rose & Woolsey (1948), mais uniquement basée sur des critères anatomiques, c.-à-d. sur les aires de projections corticales du noyau dorsomédian du thalamus. Il apparaît de manière claire aujourd’hui que ce seul critère anatomique ne suffit pas pour déterminer d’une part, d’éventuelles homologies, et, d’autre part, d’éventuelles analogies9
, entre les structures du cortex préfrontal chez le rat et celles chez le primate.
Les premières études visant à définir structurellement le cortex préfrontal, s’appuyaient sur l’existence d’une couche agranulaire (couche IV) située en avant des aires prémotrices agranulaires. Pour cette raison, le cortex préfrontal était aussi connu sous le nom de “cortex frontal agranulaire”. Cette définition est inadaptée dès lors que l’on s’intéresse au rongeur, p.ex. les aires motrices primaires sont granulaires chez cette espèce alors qu’elles sont agranulaires chez le primate. Les seuls arguments cytoarchitectoniques ne peuvent donc être utilisés pour caractériser cette structure. Le pattern de connectivité, les propriétés fonctionnelles (tant électrophysiologiques que comportementales),
9
On trouve dans la littérature concernant les lobes frontaux les termes ‘équivalentes’, ‘compara-bles’ ou ‘analogues’, mais rarement ‘homologues’ pour désigner les caractéristiques communes des cortex préfrontaux chez différents groupes phylogénétiques. Dans le champ de la biologie évo-lutionniste, deux structures sont dites homologues si elles dérivent de l’évolution d’une structure ancestrale commune. Le terme d’analogie est utilisé lorsque deux structures remplissent la même fonction mais que leur développement provient d’une évolution parallèle et non commune. De fait, deux structures peuvent être homologues sans avoir nécessairement la même fonction. Le terme d’homologie fonctionnelle prend alors tout son sens. Pour ce point particulier concernant le cortex préfrontal, voir Preuss (1995), pour une discussion plus générale, voir Striedter (2002).