Approches anatomique et par imagerie des nerfs du plexus brachial : application pour le bloc échoguidé lors de chirurgie de l'épaule
80
0
0
Texte intégral
Figure
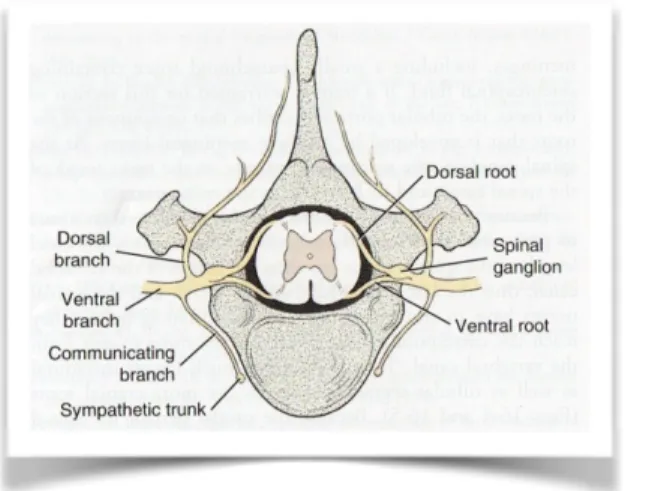
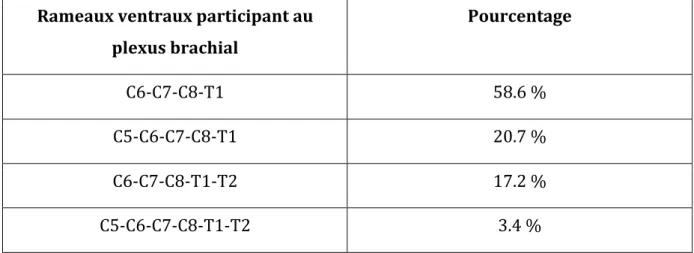
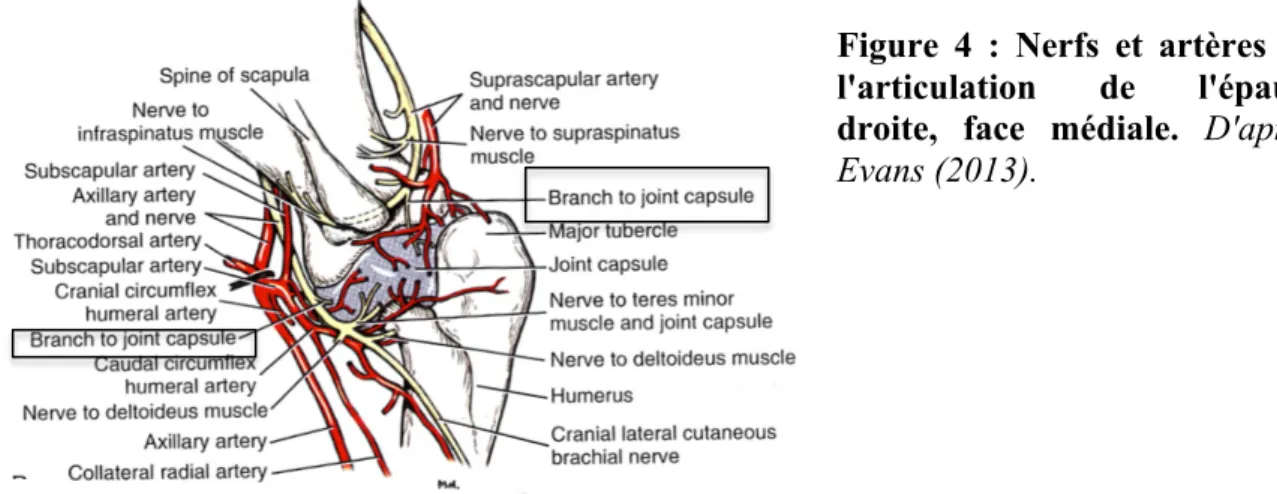
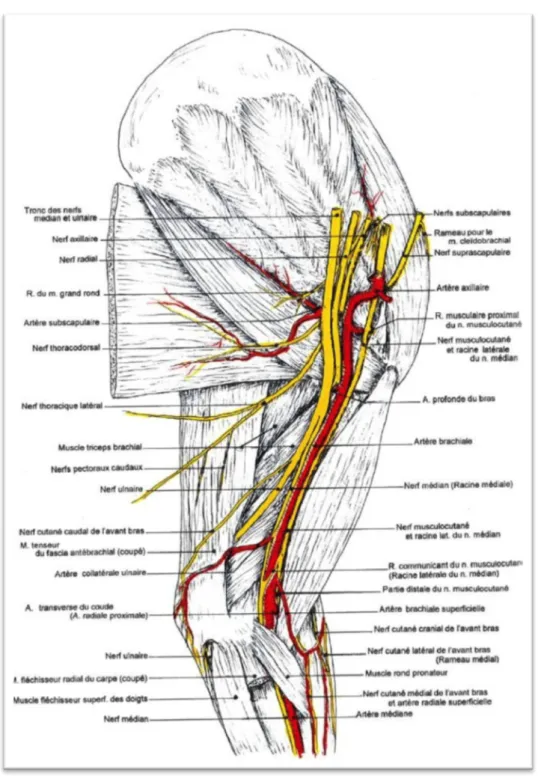
+7
Outline
Documents relatifs