Étude du rôle des récepteurs NMDA du mésencéphale ventral dans la récompense induite par la stimulation électrique du mésencéphale postérieur chez le rongeur
120
0
0
Texte intégral
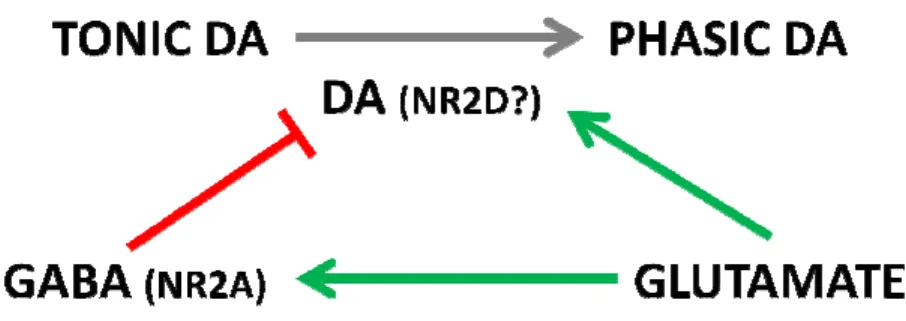
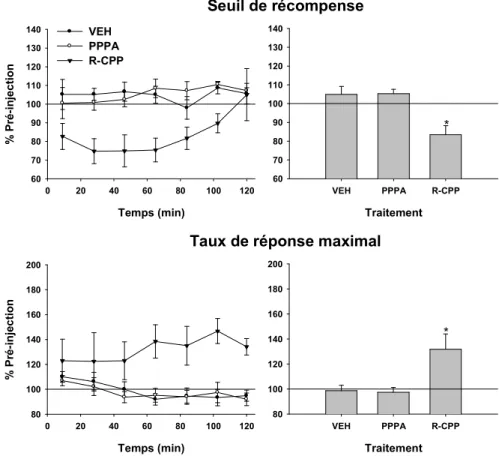
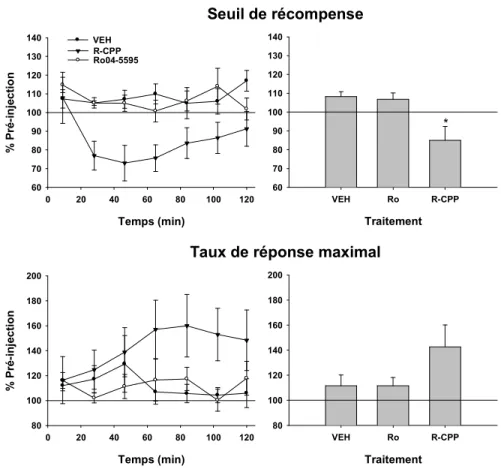
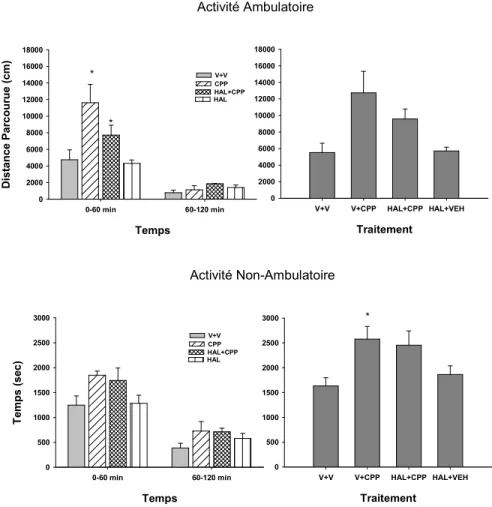
Figure
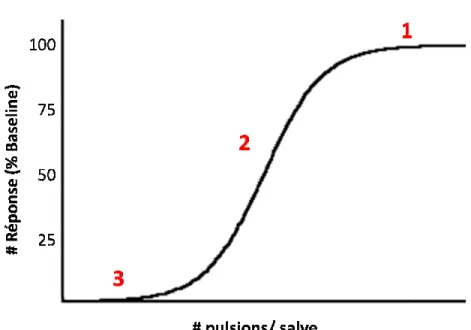
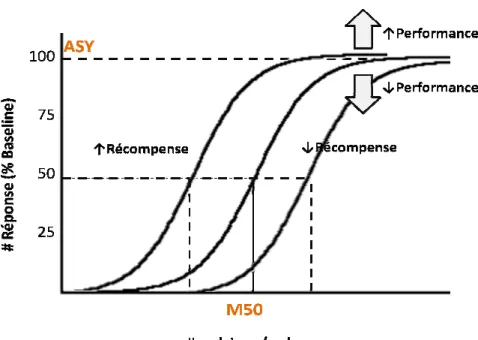
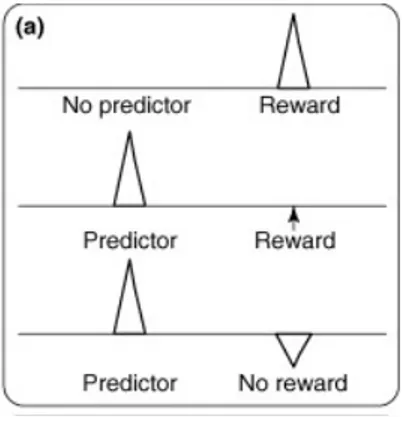
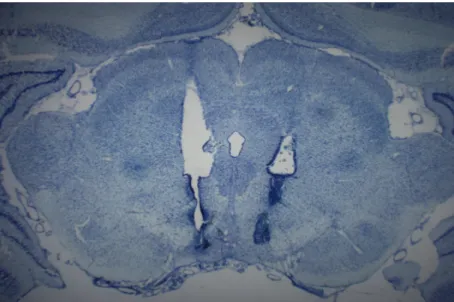
+6

Documents relatifs
