ETUDE DE L'EFFET DE LA CONTAMINATION EN METAUX LOURDS SUR FRANKIA SPP. ET SA SYMBIOSE AVEC L'AULNE NOIR
(ALNUS GLUTINOSA (L.) GAERTN).
par
Pier-Anne Belanger
Memoire presente au Departement de biologie en vue de l'obtention du grade de maitre es sciences (M.Sc.)
FACULTE DES SCIENCES UNIVERSITE DE SHERBROOKE
1*1
Library and Archives Canada Published Heritage Branch 395 Wellington Street Ottawa ON K1A 0N4 Canada Bibliotheque et Archives Canada Direction du Patrimoine de I'edition 395, rue Wellington Ottawa ON K1A 0N4 CanadaYour file Votre reference ISBN: 978-0-494-65614-3 Our file Notre reference ISBN: 978-0-494-65614-3
NOTICE: AVIS:
The author has granted a
non-exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
telecommunication or on the Internet, loan, distribute and sell theses
worldwide, for commercial or non-commercial purposes, in microform, paper, electronic and/or any other formats.
L'auteur a accorde une licence non exclusive permettant a la Bibliotheque et Archives Canada de reproduce, publier, archiver, sauvegarder, conserver, transmettre au public par telecommunication ou par Plnternet, preter, distribuer et vendre des theses partout dans le monde, a des fins commerciales ou autres, sur support microforme, papier, electronique et/ou autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur conserve la propriete du droit d'auteur et des droits moraux qui protege cette these. Ni la these ni des extraits substantiels de celle-ci ne doivent etre imprimes ou autrement
reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
Conformement a la loi canadienne sur la protection de la vie privee, quelques
formulaires secondaires ont ete enleves de cette these.
While these forms may be included in the document page count, their removal does not represent any loss of content from the thesis.
Bien que ces formulaires aient inclus dans la pagination, il n'y aura aucun contenu manquant.
1*1
Le 19 mai 2010
lejury a accepte le memoire de Madame Pier-Anne Belanger dans sa version finale.
Membres du jury
Professeur Sebastien Roy Directeur de recherche Departement de biologie
Professeur Francois Malouin Membre
Departement de biologie
Monsieur Ryszard Brzezinski President rapporteur Departement de biologie
SOMMAIRE
Les principaux responsables de la contamination des-sol's par les metaux sont les exploitations minieres et les activites industrielles. Evidemment, de tels ajouts de contaminants au niveau du sol affecteront directement sa microflore qui comprend plusieurs microorganismes d'importance environnementale comme les bacteries fixatrices d'azote. Les methodes actuelles de decontamination des sols s'averent couteuses et frequemment dommageables pour l'environnement en plus d'etre limitees au niveau du volume de sol pouvant etre traite. C'est pourquoi dans les dernieres annees les efforts de recherche se concentraient a deVelopper de nouvelles biotechnologies de decontamination utilisant les capacites naturelles de certains microorganismes (bioremediation) et de certaines plantes (phytoremediation). C'est dans cette optique que pourraient etre utilises les bacteries du genre Frankia spp. et les aulnes.
Frankia spp. se retrouve dans le sol partout a l'echelle mondiale. Elle fait partie de la classe des actinomycetes et possede la capacite de fixer l'azote en mode saprophyte et symbiotique (symbiose actinorhizienne). La symbiose actinorhizienne a ete observee chez plus de deux cent especes vegetales sur tous les continents sauf en Antarctique et s'avere aussi efficace dans la fixation d'azote atmospherique que la symbiose entre les Rhizobium et les legumineuses.
Les aulnes possedent dgalement une distribution geographique tres vaste. Ce sont des plantes pionnieres (arbres ou arbustes) qui enrichissent graduellement le sol par l'apport de matiere organique et de matiere azotee. Leurs utilisations actuelles sont tres variees (foresterie, agroforesterie, revegetalisation de sites contamines, etc.). Parmi les especes d'aulnes, Paulne noir (Alnus glutinosa), est utilise sur plusieurs sites d'Europe contamines en metaux.
A la vue des informations disponibles sur Frankia spp. et A. glutinosa, l'hypothese que Frankia spp. et sa symbiose avecA glutinosa tolereraient les metaux dans leur environnement et pourraient servir en revegetalisation et/ou phytoremediation a ete posee. De cette hypothese decoulent deux objectifs specifiques et originaux : 1- Evaluer la resistance de Frankia spp. face aux metaux et 2- Evaluer 1'impact des metaux sur la symbiose et sur la plante hote.
Dans le cadre de ce projet, une methode de criblage en milieu de culture liquide a ete developpee afin de determiner la tolerance de cinq souches de Frankia sp. envers douze metaux. La methode de revelation choisie impliquait la reduction d'un sei de tetrazolium par le metabolisme respiratoire des souches. Ces travaux ont permis de mettre en evidence une diversite au niveau de la tolerance des souches de Frankia sp. face aux metaux.
La tolerance de la symbiose actinorhizienne face aux metaux a ete etudiee grace a la souche Frankia ACN14a et A. glutinosa. La culture hydroponique de ces organismes dans des conditions gnotobiotiques et dans un environnement controle (cycle de luminosite, temperature et humidite) a permis d'observer plusieurs phenomenes interessants. Les resultats globaux demontrent que la symbiose serait davantage affectee par les metaux lors de son etablissement (denombrement des nodules) que lors de son developpement (poids frais des nodules). De plus, la symbiose avec Frankia ACN 14a favoriserait un developpement superieur des parties aeriennes de l'aulne. Aussi, aucun metal teste (aux concentrations permettant la survie de la plante) n'a aboli completement la symbiose et la capacite fixatrice d'azote de Frankia ACN14a dans les nodules. Le dosage deTactivite nitrogenase a ete mesure via la reduction de 1'acetylene en ethylene par chromatographic en phase gazeuse. Finalement, la quantite de metal se retrouvant dans les parties aeriennes a ete quantifiee par ICP-MS et les resultats ont demontre la possibilite que l'aulne noir accumule des metaux au niveau des tiges et feuilles. A long terme, ces resultats pourraient etre transposes a grande echelle et servir a i'elaboration de strategies de decontamination employant les aulnes noirs et Frankia spp..
REMERCIEMENTS
J'aimerais remercier mon directeur de recherche le professeur Sebastien Roy pour l'opportunite qu'il m'a offerte, mais surtout pour son encadrement et son soutien tout au long de ces deux dernieres annees. Je dois egalement exprimer des remerciements sinceres a mes conseillers de maitrise les docteurs Ryszard Bfzezinski et Francis Malouin pour leurs idees ainsi que leurs judicieux conseils.
Aussi, un grand merci a tous les membres de mon laboratoire de recherche ainsi qu'aux membres des laboratoires des docteurs Carole Beaulieu et Ryszard Brzezinski pour leur esprit critique et les discussions scientiflques toujours interessantes.
Je ne pourrais passer sous silence l'aide precieuse du docteur Jean-Philippe Bellenger de PUniversite de Princeton pour les analyses ICP-MS ainsi que 1'interpretation des resultats obtenus.
Je me dois egalement de mentionner le Fonds quebecois de la recherche sur la nature et les technologies pour son soutien financier par le biais de leur bourse de maitrise en recherche.
Enfin, je voudrais aussi remercier ma famille pour leurs encouragements et Simon Despins pour sa patience et son opinion concernant certains travaux et presentations.
TABLE DES MATIERES
SGMMAIRE .- I REMERCIEMENTS ....Ill TABLE DES MATIERES IV LISTE DES ABREVIATIONS VII LISTE DES TABLEAUX IX LISTE DES FIGURES X INTRODUCTION I
1. Problematique etudiee : la contamination anthropogenique des sols par les metaux.. 1
1.1 Metaux frequemment retrouves dans les sols contamines... 2
1.2 Methode d'assainissement des sites contamines ..3
1.2.1 Les methodes ex situ 3 1.2.2 Les methodes in situ ;.... 4
2. Les organismes etudies 4
2.1 Frankiaspip 5 2.2 Les plantes actinorhiziennes 8
2.3 La symbiose entre Frankia spp. et les aulnes... 11 3. Les aulnes : leur utilisation anterieure et actuelle. 13 3.1 L'aulnenoir {Alnus glutinosa (L.) Gaertn) 16 4. L'effet des metaux sur les organismes vivants 17 4.1 L'effet des metaux sur les microorganismes... .17
4.2 L'effet des metaux sur les plantes 19 4.3 L'effet des metaux sur les symbioses entre les plantes et les microorganismes
fixateurs d' azote 21 5. Hypothese et objectifs du projet de recherche 24
CHAPITRE 1
HIGH-THROUGHPUT SCREENING OF MICROBIAL ADAPTATION TO
ENVIRONMENTAL STRESS ..25
2. Originalite du travail et apport a la science 25
3. Resume de 1'article en francais.. ..26
4. Manuscrit de 1'article 27
4.1 Abstract 27 4.2 Introduction .27 4.3 Results.. 31 4.3.1 Method development 31
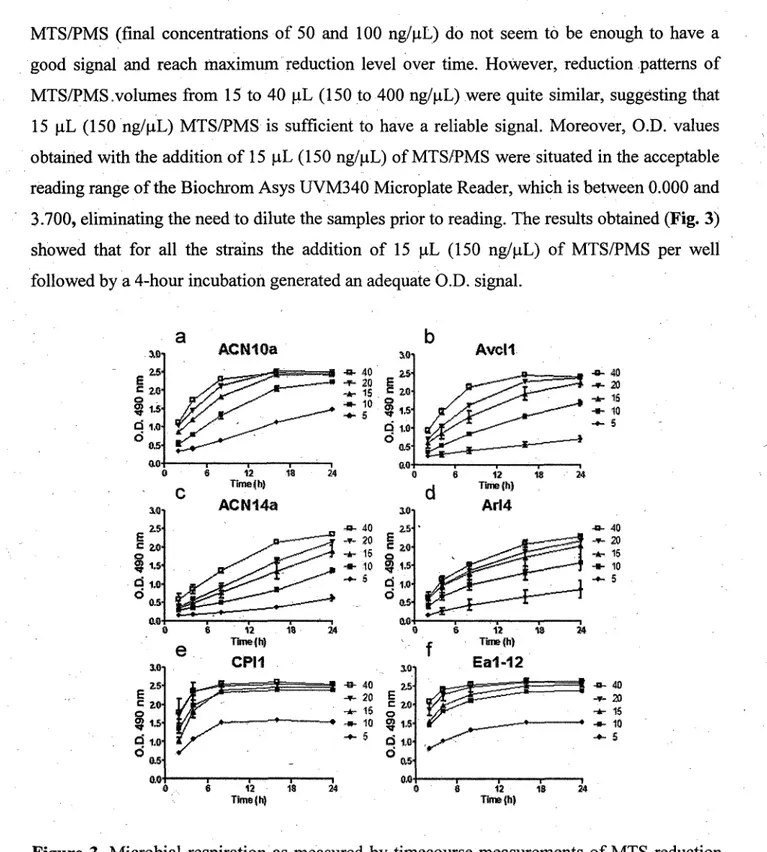
4.3.1.1 Culture medium and inoculation density 32 4.3.1.2 Optimization of MTS reagent concentration used in the revelation step 34
4.3.1.3 Optimization of respiration revelation procedure: incubation time and
temperature 36 4.3.2 Large-scale evaluation of Frankia spp. tolerance to heavy metals 36
4.3.2.1 Metals of acute toxicity: CdCl2, CoCl2, CuCl2, K2Cr04, NiCl2 and ZnCl2... 37
4.3.2.2 Metals of moderate toxicity: A1C13, Pb(N03)2 andNaV03 39
4.3.2.3 Metals of weak toxicity: NaAs02, MgCl2 andNa2Mo04 40
4.4 Discussion 41 4.5 Methods 42 4.5.1 Bacterial strains 42
4.5.2 Microwell plate preparation and incubation 43 4.5.3 Revelation of microbial viability using MTS/PMS coloration.. 44
4.6 Acknowledgements ...C 44
4.7 References 45
CHAPITRE2
L'IMPACT DES METAUX SUR LA SYMBIOSE FRANKIA-AVLNE 50
1. Mise en contexte destravaux , 50
3. Materiel et methodes 54 3.1 Souche bacterienne 54 3.2 Preparation des graines d'aulnes 55
3.3 Preparation du materiel de culture vegetale 56 3.4 Transfer!, entretien et croissance des plantules en sachets de germination 5 6
3.5 Ajout des metaux et de rinoculum ....57 3.6 Recolteet dissection ; 58
3.7 Dosage de Pactivite nitrogenase ; 59
3.8 Bioaccumulation des metaux 60 4..Resultats , ...60 4.1 Evaluation de la methode culturale ; 60
4.2 Examen de l'effet de la premiere serie de metaux: Cu, Ni, Zn, Pb et Cd 61 4.3 Examen de l'effet de la deuxieme serie de metaux : Al, Co-, Cr, Mg, Mn et Mo....64
4.4 Examen de l'effet de la troisieme serie de metaux : As, Se et V ..66
4.5 Bioaccumulation des metaux... 68 4.5.1 Resultats de bioaccumulation, serie 1 : Cu, Ni, Zn, Pb et Cd 69
4.5.2 Resultats de bioaccumulation, serie 2 : Al, Co, Mg, Mn et Mo 69
4.5.3 Resultats de bioaccumulation, serie 3 : As, Se et V :.71
5. Discussion... 72 5.1 Methode culturale..;... 72
5.2 Influence sur la symbiose du Cu, du Ni, du Zn, du Pb et du Cd ,...73 5.3 Influence sur la symbiose de l'Al, du Co, du Cr, du Mg, du Mn et du Mo 74
5.4 Influence sur la symbiose de l'As, du Se et du V 75 5.5 La bioaccumulation des metaux „.. 75
CONCLUSION. ...:... .80 BIBLIOGRAPHIE , ...83
LISTE DES ABREVIATIONS
acide desoxyribonucleique biovar
acide ethylene diamine terra aeetique exemple
force relative de centrifugation gramme
unite de grosseur d'aiguilles (calibre) guanine et cytosine contenues daris 1'ADN gramme par gramme
gramme par litre
Inductively Coupled Plasma-Mass Spectrometry kilogramme (10 gramme)
milligramme par kilogramme Minimal inhibitory concentration minute
millilitre (10"3 litre) • • a
millimetre (10" metre) millimolaire
MTC MTS nmol NNPP' PCV PGPR pH PMS Pr. p/v ppm
maximal tolerable concentration
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt
MTS/PMS solution contenant un melange de MTS et de PMS selon les concentrations recommandees
nanomole (10" mole) nombre de nodules par plant
packed cell volume (volume de culot bacterien en uL) plant growth promoting rhizobacteria
potentiel hydrogene phenazine methosulfate professeur
poids/volume
partie par million (mg/L)
ratio A/R ratio du poids des parties aeriennes d'un plant sur le poids des parties racinaires de ce meme plant
ssp. sous-espece Hg/L microgramme par litre uL microlitre(10'6 litre)
um micrometre (10"6 metre)
LISTE DES TABLEAUX
Tableau 1. Liste des especes actinorhiziennes regroupees selon la pu les regions dans
laquelle ou lesquelles elles se retrouvent.... 10 Tableau 2. Concentrations de metaux auxquelles les aulnes noirs ont ete exposes...58
LISTE DES FIGURES
Figure 1. Morphologies du genre Frankia en microscopie electronique a balayage. 6 Figure 2. Distribution globale de toutes les especes actinorhiziennes combinees 8 Figure 3. Distribution des plantes actinorhiziennes de la famille des Betulaceae.. 9 Figure 4. Resultats de 1'impact du Cu, du Ni, du Zn, du Pb et du Cd sur la symbiose entre
Frankia ACN14a et l'aulne noir .63 Figure 5. Resultats de l'impact de PA1, du Co, du Gr, du Mg, du Mn et du Mo sur la
symbiose entre Frankia ACN14a et l'aulne noir ....65 Figure 6. Resultats de l'impact de 1'As, du Se et du V sur la symbiose entre Frankia
ACNHaet l'aulne noir 67 Figure 7. Bioaccumulation des metaux dans les parties aeriennes des aulnes noirs inocules et
INTRODUCTION
1. Problematique etudiee : la contamination anthropogenique des sols par les metaux
Les metaux font partie integrante de la composition normale des sols. lis proviennent naturellement des processus geologiques et leurs concentrations de fond varient selon les regions. Par contre, suite a la venue de l'ere industrielle, les metaux se retrouvent frequemment en quantites excessives dans les sols, ce qui en fait des contaminants majeurs et preoccupants. •
Les sources principales des contaminations anthropogeniques des sols en metaux sont l'utilisation des combustibles fossiles, les industries minieres ainsi que les fonderies, Tapplication d'engrais et de pesticides tout comme le rejet des eaux usees ou des boues
d'epuration (Garbisu et Alkorta, 2001; Almeida Pereira et al., 2006; Khan et ah, 2009). Les sols peuvent egalement s'enrichir en metaux suite a la dispersion aerienne de fines particules de sols et a la deposition de ces contaminants (Mendez et Maief, 2008; Roy et al., 2005; Dickinson, 2000). II arrive egalement des accidents qui entrainent une contamination aigiie et subite des sols. Le deversement de boues provenant de la mine de pyrite Aznalcollar (sud de l'Espagne) ou 4,5 millions de metres carres de boues contaminees avec au moins douze metaux ont ete repandus sur des terres agricoles en est un exemple (Carrasco et al., 2005).
Contrairement aux polluants organiques dont la concentration dans le sol s'attenue avec le temps grace a des processus biologiques de decomposition, il est impossible de degrader biologiquement les metaux contaminant Penvironnement (Ledin, 2000). Cela represente un reel probleme puisque les metaux sont les contaminants les plus repandus (Dickinson, 2000; Garbisu et Alkorta, 2001; Khan et al., 2009). Cette impossibilite de degradation et
d'elimination rend le temps de retention des metaux dans le sol indetermine (Roy et al., 2005; Khan et al., 2009). Cette incapacity a se debarrasser naturellement des metaux inquiete puisque les metaux.peuvent causer de serieux pfoblemes de sante chez les humains et les animaux en plus d'affecter l'environnement et les microorganismes. Les contaminations anthropogeniques des sols en metaux lourds sont deja bien presentes et il est a prevoir que 1'industrialisation croissante des pays en voie de developpement ne fera qu'accroitre le probleme.
1.1 Metaux frequemment retrouves dans les sols contamines
Plusieurs metaux et metalloides sont essentiels pour les organismes vivants puisqu'ils sont impliques dans le bon fonctionnement de nombreux processus biologiques. Cependant, ils deviennent des contaminants quand ils se retrouvent sous leurs formes toxiques ou en trop grandes quantites dans le sol. Les metaux ou metalloides les plus communement retrouves dans les sols contamines au Canada aihsi qu'aux Etats-Unis incluent l'arsenic (As), le cadmium (Cd), le cuivre (Cu), le chrome (Cr), le plomb (Pb), le mercure (Hg), le nickel (Ni), le selenium (Se), l'argent (Ag) et le zinc (Zn). D'autres metaux ou metalloides tels que l'aluminium (Al), le cesium (Cs), le cobalt (Co), le manganese (Mn), le molybdene (Mo), le strontium (Sr) et l'uranium (U) sont aussi considered comme contaminants, mais sont retrouves moins frequemment (Mclntyre, 2003). La contamination des sols avec un unique metal se rencontre tres rarement. La majprite des sols contamines contiennent un melange complexe de plusieurs metaux ce qui rend leur assainissement d'autant plus complique (Glick,, 2003).
1.2 Methodes d'assainissement des sites contamines
< •
Les methodes d'assainissement traditionnelles des sols contamines font principalement appel a des precedes physicochimiques dispendieux. De nouvelles methodes plus ecologiquement responsables font peu a peu leur apparition et s'averent egalement moins couteuses. Les methodes en general se divisent en deux grandes classes soit ex situ et in situ.
1.2.1 Les methodes ex situ
Les methodes ex situ sont generalement couteuses et plus dommageables pour l'environnement. L'une des methodes mecaniques de decontamination les plus employees consiste a excaver les sols contamines en metaux pour les transporter vers un site d'enfouissement securitaire (Khan et ah, 2009). Le principal inconvenient de ce type de methode, c'est qu'elle implique un deplacement des residus. Elle ne s'applique done qu'aux cas de contamination ou la quantite de materiel a decontaminer est mineure. De plus, cette methode est tres dispendieuse et possede un cout moyen de 1 000 000 $ par acre (Garbisu et Alkorta, 2001). II existe egalement des methodes chimiques comme la lixiviation qui consiste a detacher les metaux des particules de s o l a l'aide de solutions acides ou d'autres agents chimiques puis a retourner le sol ainsi nettoye vers son emplacement initial (Khan et al., 2009; Garbisu et Alkorta, 2001). Cette technique comporte egalement un inconvenient majeur : les traitements chimiques imposes sont si drastiques qu'il y a co-extraction de tous les autres nutriments en plus des metaux. II en resulte un sol extremement appauvri avec un pH acide ne permettant pas l'etablissement facile des especes vegetales.
1.2.2 Les methodes in situ
Les methodes de decontamination in situ sont habituellement moins couteuses, facilement applicables a de grahdes surfaces et plus douces pour l'environnement. Certaines methodes chimiques comme le chaulage permettent d'elever le pH du sol afin de stabiliser les metaux et d'eviter leur dissemination (Dickinson, 2000). II existe egalement des methodes biologiques comme la phytoremediation. Cette approche englobe plusieurs methodes de decontamination in situ relativement recentes qui utilisent principalement les plantes comme agent de detoxification. Les phytotechnologies applicables aux metaux comprennent la phytoextraction (elimination des metaux du sol), la phytostabilisation (sequestration des metaux dans le sol) et la phytovolatilisation (volatilisation des metaux) (Mendez et Maier, 2008; Garbisu et Alkorta, 2001). Lors de la phytoextraction, a la fin du procede, la biomasse vegetale peut etre briilee ou compostee et dependamment de la concentration en metaux, ces derniers peuvent meme etre recuperes (Garbisu et Alkorta, 2001).
2. Les organismes etudies
Ce projet de recherche en microbiologic, s'interesse aux metaux presents dans l'environnement, mais plus specifiquement a l'impact qu'auront ces metaux sur les microorganismes du sol ainsi que sur certaines plantes symbiotiques. Nos travaux se concentrent sur le genre Frankia et sur les aulnes qui representent une des especes hotes des Frankiaspp.. \
2.1 Frankia spp.
Bien que 1'etude' des bacteries du genre Frankia est en plein essor depuis les annees 1980, l'existence de Frankia spp. etait soupcorinee par les biologistes depuis 1829 et des 1866 la premiere description d'un nodule se retrouvait dans la litterature scientifique (Schwencke et Caru, 2001; Normand, 2006). Le nom Frankia spp. fut propose en 1886 par un denomme Brunchorst en l'honneur de son mentor, le microbiologiste Suisse A. B. Frank (Benson et Silvester, 1993; Lechevalier et Lechevalier, 1989). Ce n'est qu'en 1964 que la presence d'actinomycetes est decelee et confirmee par microscopie dans des nodules, mais aucune souche n'avait encore ete isolee (Benson et Silvester, 1993). En 1959, Pommer aurait isole puis perdu une souche de Frankia spp.. Cette malchance designe done comme point tournant de la recherche sur Frankia spp. l'isolement d'une premiere souche en culture pure par Torrey et ses collegues en 1978 (Wall, 2000; Benson et Silvester, 1993).
Les Frankia spp. sont des bacteries a Gram positif ayant un genome tres riche en G+C (variant entre 66 et 77%) (Benson et Silvester, 1993; Pawlowski et Bisseling, 1996; Schwencke et Caru, 2001). Elles ont une croissance tres lente (temps de doublement allant de 15 a 48 h et meme plus) (Benson et Silvester, 1993). Les Frankia spp. sont des bacteries filamenteuses (developpement sous forme d'hyphes) qui se caracterisent par la formation d'agregats plus ou moins compacts en milieu liquide. Lors de sa croissance, Frankia spp. se retrouve sous trois formes principales (Figure 1) : les hyphes vegetatifs, les vesicules permettant la fixation de 1'azote atmospherique ainsi que les sporanges multiloculaires permettant la dissemination via les sporangiospores (Benson et Silvester, 1993). Une quatrieme structure a ete observee chez Frankia spp., il s'agit des hyphes de reproduction toruleux {reproductive torulose hyphae ou RTH) qui seraient une deuxieme forme de propagules (Diem et Dommergues, 1985). Certaines souches de Frankia spp. produisent egalement des pigments intracellulaires ou extracellulaires en milieu de culture. Ces pigments sont de couleurs tres variees passant par le jaune, 1'orange, le rouge, le vert, le brun et allant jusqu'au noir (Benson et Silvester, 1993).
Figure 1. Morphologies du genre Frankia en microscopie electronique a balayage. A) Hyphes avec vesicules (v). B) Vesicules de Frankia attachee a son hyphe (H). C) Sporange multiloculaire de Frankia sp., images tirees de Schwencke et Caru (2001).
Les Frankia spp. appartiennent a la classe des actinomycetes (Normand, 2006). Du point de vue taxonomique, elles appartiennent a l'ordre des Actinomycetales, au sous-ordre des Frankineae et a la famille des Frankiaceae (Benson et Dawson, 2007). La taxonomie des Frankia spp. jusqu'a l'etape du genre est bien definie, mais les criteres a evaluer afin de completer la classification des especes de Frankia spp. font toujours l'objet de controverses. II n'y a done aucune classification claire au niveau des especes chez Frankia spp. a ce jour (Schwencke et Caru, 2001; Lechevalier et Lechevalier, 1989). Pour pallier a cette lacune, l'appellation des souches suit les criteres d'un systeme provisoire de lettres et de chiffres. Ce code alphanumerique attribue a chaque souche represente le laboratoire d'origine, le genre et Pespece de la plante hote en plus de comprendre un nombre unique de six a huit chiffres pour chacune des souches (Benson et Silvester, 1993).
La distribution de Frankia spp. a l'echelle mondiale est extremement vaste. En effet, la nodulation produite par Frankia spp. a ete observee sur tous les continents sauf en Antarctique (Benson et Dawson, 2007). Ce serait la senescence des nodules qui permettrait le maintien de
Frankia spp. dans les sols et 1'initiation de nouveaux nodules (Benson et Dawson, 2007; Benson et Silvester, 1993). Par contre, il est aussi connu que Frankia spp. survit de maniere saprophyte au niveau du sol (Richardson et al., 2000). Des etudes viennent appuyer ce mode de croissance saprophyte en demontrant que Frankia spp. persiste dans le sol apres la disparition de ses plantes hotes ou quelle est presente malgre l'absence totale de plantes hotes (Benson et Dawson, 2007; Nickel et al, 2001; Wall, 2000; Benson et Silvester, 1993). Les spores seraient les responsables majeurs de la dissemination de Frankia spp. via les sediments ou encore dans l'air via les oiseaux (Wall, 2000).
Frankia spp. se caracterise egalement par sa capacite a fixer 1'azote atmospherique. Grace a son enzyme nitrogenase Frankia spp. est en mesure de convertir le diazote (N2) atmospherique en ammoniac ou en ions ammonium (Schwencke et Cam, 2001; Lundquist, 2000; Wall, 2000; Pawlowski et Bisseling, 1996; Valverde et Wall, 2003b). Cette reaction de fixation d'azote a lieu a l'interieur des vesicules de Frankia spp.. Ces vesicules possedent une membrane lipidique multicouche ainsi que des septa internes les isolant des hyphes vegetatifs (Schwencke et Caru, 2001; Pawlowski et Bisseling, 1996; Benson et Silvester, 1993). Ces caracteristiques seraient essentielles afin de proteger 1'enzyme nitrogenase dont Taction est inhibee par la presence d'oxygene (Lundquist, 2000; Benson et Silvester, 1993). Bien que Frankia spp. ne soit pas la seule bacterie capable de transformer l'azote atmospherique en sources d'azote assimilables, elle est particuliere puisqu'en mesure de realiser cette conversion autant en mode saprophyte qu'en mode symbiotique (Schwencke et Caru, 2001; Benson et Silvester, 1993). Suite a la fixation d'azote, les sols contenant Frankia spp. sous forme libre ou en symbiose finissent par s'enrichir en matiere azotee et deviennent elements pour d'autres especes vegetales ayant des besoins nutritionnels plus grands (Normand, 2006; Nickel et al, 2001). Frankia spp. est done l'exemple d'un microorganisme ubiquiste ayant une grande importance au niveau environnemental, qui influence les cycles biogeochimiques globaux et sur lequel les informations sont encore limitees. II est done primordial de continuer les recherches sur cette bacterie afin de parfaire nos connaissances a son sujet et de mieux definir ses exploitations potentielles en biotechnologie.
2.2 Les plantes actinorhiziennes
Les plantes actinorhiziennes represented tous les genres vegetaux capables d'etablir une symbiose racinaire avec le genre bacterien Frankia spp. (Schwencke et Cam, 2001; Benson et Silvester, 1993). Ces plantes englobent environ deux cent especes d'arbres et d'arbustes regroupees en vingt-quatre genres reunis dans huit families (Franche et al., 2009; Normand et al., 2007; Schwencke et Cam, 2001; Gualtieri et Bisseling, 2000). Les plantes actinorhiziennes sont souvent des especes pionnieres initiant la succession vegetale ^et se retrouvent dans pratiquement tous les ecosystemes : la toundra arctique, les dunes de sable cotieres, les forets en plus des regions arides ou alpines (Benson et Silvester, 1993). La grande adaptabilite de ces plantes aux differents ecosystemes fait que leur distribution geographique s'etend a l'echelle mondiale telle que representee par la figure 2.
Figure 2. Distribution globale de toutes les especes actinorhiziennes combinees, image tiree de Benson et Dawson (2007).
Le nombre d'especes actinorhiziennes se retrouvant dans chacune des regions geographiques du globe varie considerablement (Tableau 1). Les Ameriques possedent la diversite de plantes
actinorhiziennes la plus riche suivies par l'Asie et,l'Oceanie. Pour ce qui est de l'Europe et de •l'Afrique, le nombre d'especes et de genres diminuent considerablement. Parmi toutes les plantes actinorhiziennes, seuls deux genres sont distribues sur tous les continents (a 1'exception de l'Antarctique). II s'agit des genres Alnus et Myrica appartenant respectivement aux families des Betulaceae et des Myricaceae.
La famille des Betulaceae comporte six genres, mais seul le genre Alnus fait partie des plantes actinorhiziennes (Normand et ah, 2007; Schwencke et Caru, 2001). Au total, il existe quarante-sept especes d'aulnes et toutes ont ete reconnues comme etablissant une symbiose avec Frankia spp. (Pawlowski et Newton,. 2008). Les aulnes se retrouvent dans un grand nombre d'habitats: depots glaciaires, dunes de sable, milieux humides, sols volcaniques, en bordure des rivieres et dans des zones de glissement de terrain (Nickel et ah, 2001; Benson et Dawson, 2007). La distribution terrestre du genre Alnus qui s'etend sur plusieurs continents demontre bien la grande adaptabilite de ce genre dans les divers ecosystemes (Figure 3).
Figure 3. Distribution des plantes actinorhiziennes de la famille des Betulaceae, image tiree de Benson et Dawson (2007).
Tableau 1. Liste des especes actinorhiziennes regroupees selon la ou les regions dans laquelle ou lesquelles elles se retrouventa
Families Genres et/ou especes Ameriques Europe Asie et Oceanie Afrique
Betulaceae Myricaceae Casuarinaceae Elaeagnaceae Rhamnaceae Rosaceae Coriariaceae Dasticaceae Alnus Alnus acuminata Alnus glutinosa Alnus viridis crispa Alnus incana rugosa Alnus rubra , ssp. ssp. Comptonia peregrina Morella Myrica Allocasuarina Casuarina Ceuthostoma Gymnostoma Elaeagnus Hippophae Shepherdia Ceanothus Colletia Discaria Kentrothamnus Retanilla Trevoa Cercocarpus Chamaebatia Purshia Coriaria Dastica X X xb X X X X X X X X X X X X ^x X X X X X X X X X X X X X X X X X X X X
aCompilation realised d'apres les references suivantes : Normand et al., 2007; Benson et Silvester, 1993; Benson et Dawson,
2007; Richardson et al., 2000; Pawlowski et Newton, 2008
Lorsque la symbiose avec Frankia spp. s'etablit et qu'elle est fonctionnelle, les plantes actinorhiziennes possedent un taux de fixation d'azote s'apparentant a celui des legumineuses (Franche et al, 2009; Schweneke et Cam, 2001; Richardson et al, 2000; Wall, 2000). En tant qu'endosymbionte, Frankia spp. pourvoit aux besoins en azote de son hote (a concurrence de 70 a 90% des besoins) en echange de sources de carbone provenant des photosynthetats (Nickel et al., 2001). Cet apport en azote varie en fonction de Page de la symbiose, des partenaires impliques, des saisons et de l'environnement. Des estimations faites par Dixon et Wheeler en 1986, suggerent meme que la symbiose actinorhizienne est responsable de 25% de la fixation d'azote planetaire (Pawlowski et Newton, 2008). La contribution des plantes actinorhiziennes a 1'augmentation de 1'azote dans les ecosystemes a lieu grace a la formation de litiere, la decomposition des racines et, dans une proportion plus faible, a 1'excretion d'exsudats racinaires dans le sol (Roy et ah, 2007). Cet apport non negligeable dans le cycle de 1'azote demontre bien 1'importance des plantes actinorhiziennes dans Pequilibre de ce cycle biogeochimique (Wall, 2000).
2.3 La symbiose entre Frankia spp. et les aulnes
Les connaissances sur l'etablissement et le d^veloppement de la symbiose actinorhizienne augmentent avec les annees, mais sont limitees lorsque comparees aux informations disponibles potir les symbioses entre Rhizobium et les legumineuses. Par exemple, alors que l'etablissement de la symbiose chez les legumineuses debute par Pechange de signaux entre les deux parties, il n'y a toujours aucune preuve demontrant qu'un phenomene semblable se produit entre Frankia spp. et son hote (Wall, 2000). Par contre, les voies d'infection utilisees par Frankia spp. afin de penetrer a Pinteiieur des racines sont mieUx connues. En effet, Frankia spp. infecte son hote de maniere intercellUlaire ou intracellulaire (Pawlowski et Bisseling, 1996). L'infection intercellulaire implique la penetration de Frankia spp. dans Pespace entre les cellules de Pepiderme et du cortex racinaire pour, par la suite, entrer dans une cellule et commencer a se multiplier a Pinterieur. Lors d'une infection intracellulaire,
Frankia spp. infecte via les poils absorbants pour, par la suite, eritrer dans les cellules et s'y multiplier. Le mode d'infection varie selon la plante note dont les racines sont colonisees par Frankia spp. puisque c'est l'hote qui controle l'infection (Franche et al., 2009; Mastronunzio etal, 2008; Gualtieri et Bisseling, 2000) Chez les aulnes, la voie d'infection intracellulaire est privilegiee (Benson et Silvester, 1993). II est a hoter que peu importe la voie empruntee par la bacterie lors de l'infection, le symbionte est entoure d'une membrane vegetale de sa penetrationjusqu'asa multiplication (Gentilie? a/., 2006; Benson et Silvester, 1993).
Afin que la fixation d'azote par l'enzyme nitrogenase soit optimale, il faut eviter que cette derniere ne soit exposee a d'importantes concentrations d'oxygene. En ce qui a trait a la symbiose entre Frankia spp. et les aulnes, les deux organismes se partagent le role de protection de la nitrogenase. Comme les nodules d'aulnes sont des organes bien aeres (activite metabolique intense), ce serait majoritairement la Frankia spp. qui controlerait la diffusion de l'oxygene. Chez Frankia spp., la plus importante barriere de diffusion bloquant l'entree de l'oxygene dans les vesicules serait la membrane lipidique multicouche des vesicules elles-memes (Lundquist, 2000). En fonction del'augmentation de la pression d'oxygene, le nombre de couches de la membrane vesiculate augmente ce qui la rend davantage impermeable aux gaz. Par contre,, il y aurait egalement des changements au niveau de l'environnement des cellules vegetales (Wall, 2000; Lundquist, 2000). Toujours chez les aulnes, des hemoglobines produites par la plante en concentrations tres faibles, pourraient avoir un role a jouer dans le controle de la diffusion de l'oxygene (Gualtieri et Bisseling, 2000; Pawlowski et Bisseling,
1996).
Les couts energetiques engendres chez la plante hote pour le developpement et le maintien de la symbiose representent une depense d'energie non negligeable. C'est pourquoi le processus de nodulation est regule chez l'hote. Le cout energetique total de la fixation d'azote est considere comme plus eleve que celui permettant l'acquisition de l'azote sous d'autres formes comme les nitrates ou Fammonium (Lundquist, 2005). Cette demande elevee en energie
explique pourquoi les aulnes sont peu tolerants a l'ombre et necessitent une luminosite tres forte pour poursuivre leur developpement (Roy et al., 2007). Par contre, la symbiose demeure puisque l'inconvenient energetique est contrebalance par l'avantage d'avoir une source d'azote facileraent assimilable dans des environnements limites en nutriments (Lundquist, 2005).
Chez les aulnes, la symbiose est particulierement efficace. Une etude realisee avec Alnus incana sous-espece rugosa, a determine que de 85 a 100% de l'azote retrouve dans les feuilles de cette espece derivait de Tatmosphere lorsque la symbiose etait fonctionnelle (Hurd et al., 2001). La fixation d'azote annuelle par les aulnes selon Schwencke et Cam (2001) s'eleverait entre 40 et 200 kg N par hectare alors que Franche et al. (2009) estime cette fixation entre 20 et 300 kg N par hectare. Ce haut taux de fixation d'azote procure certainement un avantage aux aulnes lors de la colonisation de sols difficiles (ex. : suite a une eruption volcanique) (Roy et al., 2007). Une autre propriete des aulnes leur permettant de coloniser des sols pauvres en nutriments est leur association avec des champignons mycorhiziens reconnus pour faciliter l'entree des nutriments chez leurs hotes (Franche et al., 2009; Schwencke et Cam, 2001). Les aulnes etablissent autant des ectdmycorhizes que des mycorhizes vesiculates et arbusculaires; ils possedent done le potentiel de former des symbioses impliquant jusqu'a quatre organismes (Roy et al, 2007).
3. Les aulnes : leur utilisation anterieure et actuelle
Les habiletes symbiotiques des aulnes, telles que vues ci-dessus, leur accordent un net avantage pour croitre dans des environnements hbstiles ou contamines par rhomme. De plus, la capacite des aulnes a croitre dans les climats humides, froids et/ou temperes leur confere un spectre d'utilisation plus large que celui des especes tropicales qui est restreint a certaines regions climatiques (Roy et al., 2007; Schwencke et Cam, 2001). Les aulnes sont reconnus
pour enrichir les sols pauvres en nutriments ce qui en fait une espece vegetale grandement utilisee dans diverses phytotechnologies (Franche et al., 2009; Normand, 2006; Schwencke et Cam, 2001).
En foresterie et en agroforesterie, l'emploi des aulnes est etabli et diversifie (Schwencke et Cam, 2001). En reforestation, les aulnes ont ete utilises comme especes produetrices de bois d'oeuvre, mais egalement pour produire du bois destine a l'industrie des pates et papiers (Normand, 2006; Schwencke et Cam, 2001). Comme ils enrichissent les sols a faible teneur en azote, ils peuvent servir en coculture avec des especes precieuses et exigeantes comme le noyer (Nickel et al., 2001). La coculture des aulnes a egalement ete utilisee dans des sols infectes et causant des maladies racinaires chez les coniferes (Schwencke et Cam, 2001). L'utilisation des aulnes en production de biomasse sur des terres agricoles abandonnees a aussi ete repertoriee dans la litterature (Normand, 2006). Les aulnes sont egalement retrouves sur des sites contenant des residus d'exploitation petroliere en Alberta (Sebastien Roy, communication personnelle). Finalement, les aulnes servent egalement dans des projets d'envergure comme se fut le cas dans les annees 1970 et 1980 ou six millions de Alnus viridis ssp. crispa furent plantes afin de stabiliser les digues erigees suite a la constmction de barrages hydroelectriques au Quebec (Roy et al., 2007).
Les strategies de revegetalisation ou de phytoremediation de sites miniers ou industriels contamines en metaux lourds incluent frequemment les aulnes (Schwencke et Cam, 2001). Plusieurs especes d'aulnes differentes poussant dans des residus miniers, sur des sites industriels abandonnes, ou sur des andains de sediments ont demontre un potentiel encourageant lors d'essais in situ.- Pour l'elimination des contaminants inorganiques, l'avantage des aulnes ne serait pas 1'extraction des metauxy mais plutot 1'enrichissement et
l'aeration du sol en plus d'exercer un controle hydraulique afin d'attenuer la migration des contaminants vers la nappe phreatique (Roy et al., 2007). Une tentative de phytoremediation avec A. glutinosa sur des sediments salins contamines en metaux a demontre la survie de
l'espece malgre une croissance ralentie. De plus, il n'y avait pas de Gd, Cu, Pb et Zn accumules au niveau des feuilles (Mertens et al, 2004). Un autre essai realise par Kramer et al. (2000) aux Etats-Unis dans le but de restaurer des residus de mine de cuivre, a demontre que Alnus viridis ssp. sinuata etait capable de croitre mieux que quatre autres especes indigenes a l'etude et ce avec ou sans amendements. La revegetalisation avec Alnus incana en Suisse s'est termineepar un etablissement reussi de cette espece dans des residus miniers. De plus, l'abserice d'accumulation des metaux dans les parties aeriennes a ete soulignee renfor9ant l'interet de l'utilisation de cette espece en phytostabilisation (Rosselli et al., 2003). Une experience semblable realisee sur le site d'une ancienne raffinerie avec Alnus cordata, A. glutinosa et A. incana demontrait cette fois-ci une accumulation du Cu dans les parties aeriennes de ces especes d'aulne (Dickinson, 2000). Un deuxieme essai, realise cette fois sur des sediments contamines en Angleterre, ravec A. cordata Lois, A. glutinosa L. et A. incana L. a de nouveau trouve du Cu en plus du Pb dans les feuilles (King et al., 2006). Ces resultats contradictoires d'accumulation des metaux dans les parties aeriennes ne sont probablement pas causes par les aulnes eux-memes, mais pourraient plutot etre expliques par la difference de biodisponibilite des metaux dans les sites; laquelle est grandement influencee par la composition du sol (Kabata-Pendias et Pendias, 2000). Toujours dans le domaine minier, les aulnes ont ete utilises avec succes .dans la reclamation de mines a ciel ouvert (Roy et ah, 2007). lis peuvent aussi etre exploites comme culture-abri pour d'autres plantes capables de faire l'extraction des metaux dans le sol, mais cette strategie ne peut etre envisagee que comme une solution a long terme (Roy et al., 2007). Malheureusement, dans les travaux in situ cites ci-haut, il n'etait pas specifiquement vise d'utiliser l'aulne en combinaison avec Frankiaspp..
Afin d'avoir un potentiel de decontamination plus grand, des recherches realisees en 2005 en conditions controlees ont demontre que l'inoculation de Alnus incana ssp. rugosa avec Frarikia sp. et un champignon mycorhizien {Paxillus involutus) produisait un effet synergique positif sur le developpement de la plante dans des residus contamines. Par contre, 1'etablissement de la symbiose etait si energivore qu'il provoquait un retard de croissance des
parties aeriennes (Markham, 2005). II a ete hypothesise qu'un tel retard pourrait etre evite en inoculant les plants avant de les transplanter dans les residus miniers contamines (Markham, 2005). Un effet synergique semblable sur la croissance d'A. glutinosa a ete observe lorsque cette espece etait inoculee avec Frankia spp. et Glomus intraradices BEG 163 (champignon mycorhizien) (Oliveira et ah, 2005). En somme, les donnees de la litterature demontrent la capacite des aulnes et de leur symbiose a survivre dans un contexte de sols contamines (Roy et ah, 2007). Malheureusement, les recherches faites a ce jour ne procurent aucune connaissance fondamentale sur le ou les mecanismes qui seraient affectes par les contaminants inorganiques. Cette connaissance supplemental permettrait egalement de mieux choisir les especes d'aulnes et souches de Frankia spp. aiin de specialiser les strategies a employer dans les contextes de phytostabilisation (Roy etah, 2007).
3.1 L'aulne noir {Alnus glutinosa (L.) Gaertn)
Les aulnes comprennent plusieurs especes d'arbres et d'arbustes. L'aulne noir ou aulne europeen (A. glutinosa) est un arbre. Cette espece originaire de l'Europe a ete introduite en Amerique du Nord en tant que plante ornementale. Sa distribution americaine se concentre dans la zone du Canada et des Etats-Unis entpurant les Grands Lacs (source : USDA). Les premieres recherches sur 1'exploitation des aulnes en phytotechnologies se concentraient majoritairement sur l'aulne noir en revegetalisation de sites miniers (Roy et ah, 2007). On constate notamment plusieurs etudes dans les dernieres annees qui ont utilise les aulnes noirs sur des sites contamines en metaux (King et ah, 2006; Mertens et ah, 2004; Dickinson, 2000). L'utilisation de A. glutinosa sur des sites contenant divers contaminants organiques comme les hydrocarbures demontre egalement la compatibilite de 1'espece pour la decontamination des sols (Roy et ah, 2007; Ridgway et ah, 2004). Une caracteristique interessante de l'aulne noir est sa capacite a produire une nouvelle classe de proteines vegetales liant plusieurs ions metalliques (Zn2+,Ni2+, Co2+, Cu2+, Cd2+ etHg2+); les metallohistines (Guptaetah, 2002). Ces
l'interet d'utilisation de cette espece d'aulne en bioremediation des sols (Gupta et ah, 2002). Grace a cette caracteristique, l'aulne noir serait en mesure de gerer finement et adequatement le transport et la biodisponibilite des metaux dans la plante et peut-etre meme au niveau du sol.
4. L'effet des metaux sur les organismes vivants
Comme les metaux sont tres repandus dans la nature et proviennent principalement de sources geochimiques naturelles, la necessite de developper des mecanismes de resistance chez les organismes s'est fait sentir tres tot dans revolution (Rosen, 2002). Certains metaux sont devenus essentiels ou benefiques pour les plantes, animaux et microorganismes (Ca, Co, Cu, Fe, Mg, Mn, Mo, Ni, Se, V, Zn), alors que d'autres ne possedent aucune fonction physiologique ou biologique connue (Ag, As, Au, Bi, Cd, Hg, Pb, Sb, Te) et sont consideres comme toxiques. Le seul metal dont la necessite ou la toxicite est mitigee est le Cr. En effet, le Cr se retrouve dans les deux groupes dependamment de la litterature (Ramirez-Diaz et ah, 2008; Almeida Pereira et ah, 2006; Trajanovska et ah, 1997; Summers, 2009; Chen et ah, 2005).
4.1 L'effet des metaux sur les microorganismes
Les concepts de tolerance, resistance et sensibilite aux metaux peuvent etre facilement confondus les uns avec les autres ou utilises un peu a toutes les sauces. De plus, les definitions precises des termes tolerance et resistance portent parfois a confusion dans la litterature. Une publication de Gadd (1992) propose done une terminologie plus precise et specifique. Le terme «tolerance » devrait etre employe lorsque les organismes supportent la presence des metaux grace a leurs caracteristiques biologiques intrinseques, alors que le mot « resistance »
devrait etre mentionne lorsque les organismes survivent a la presence de metaux a l'aide de mecanismes de detoxification specifiques induits en presence du contaminant (Gadd, 1992). II faut donc.porter une attention particuliere a l'emploi de ces mots en contexte de contamination ou de decontamination des metaux. De maniere globale, les metaux exercent une pression selective augmentant la tolerance de base de certains microorganismes, mais diminuent la biodiversite des microorganismes retrouves dans ces sols en comparaison avec des sols non pollues (Almeida Pereira et al., 2006). La sensibilite des microorganismes peut se traduire par une diminution de l'expression proteique alors qu'une augmentation de l'expression proteique signifierait une tentative d'adaptation au nouveau contaminant (expression d'un mecanisme de resistance) (Almeida Pereira et al., 2006). Chez les bacteries, les genes codant pour l'expression des proteines impliquees dans ces mecanismes sont principalement plasmidiques, bien que quelques-uns soient chromosomiques (Trajanovska et al., 1997). L'origine plasmidique de certains genes peut resulter en une frequence de transfert plus elevee de ces plasmides en conditions de stress. Un tel exemple a ete demontre chez Rhizobium leguminosarum bv. viciae qui, lorsqu'expose a la presence de metaux, transferait ses plasmides lui procurant sa resistance (Lakzian et al., 2007).
La regulation de l'homeostasie des metaux chez les procaryotes englobe plusieiirs mecanismes agissant communement afin de maintenir une concentration de metaux intracellulaire adequate. Habituellement, ces systemes de regulation se composent de trois sous-systemes : 1- une pompe permettant 1'entree de metaux presents en concentrations faibles dans le milieu exterieur, 2- une pompe a efflux pour eliminer les surplus de metaux dans le cytoplasme et 3- une proteine relais amenant les metaux des pompes vers leurs cibles cellulaires (Summers, 2009). D'autres voies d'entree peuvent aussi etre employees, quelques metaux toxiques comme le chromate ou 1'arsenate penetrent dans le cytoplasme via des systemes de transport du phosphate ou du sulfate. Ces transporteurs non specifiques vont done etre frequemment sous-exprimes et/ou rempiaces par des systemes plus restrictifs afin de limiter l'apport de metal toxique (Ramirez-Diaz et al, 2008). Les principaux mecanismes de tolerance et de resistance retrouves chez les bacteries comprennent: 1- les pompes a efflux dependantes de
l'energie, 2- la precipitation des metaux sous forme de sels insolubles, 3- les alterations de la permeabilite de la membrane plasmique, 4-1 'immobilisation des metaux a la paroi cellulaire, 5- la production d'agents chelateurs dans le milieu extracellulaire et 6- la transformation biochimique des ions metalliques (Haferburg et Kothe, 2007; Figueira et ah, 2005; Nies,
1999). II a egalement ete demontre chez les diazotrophes que des siderophores sont produits afin de reguler l'acquisition de certains metaux essentiels a l'enzyme nitrogenase (Fe, Mo, V) et l'exclusion de metaux toxiques comme le W (Kraepiel et ah, 2009). La presence des siderophores a egalement ete demontree chez les bacteries fixatrices d'azote du genre Frankia; ces siderophores servaient au recrutement de Fe (Boyer et ah, 1999; Aronson et Boyer, 1994). II est important de noter que tous ces mecanismes de tolerance et/ou resistance developpes par les bacteries sont a la base de precedes biotechnologiques commerciaux comme la biosorption, la bioprecipitation et la biolixiviation des metaux (Gadd, 2004).
4.2 L'effet des metaux sur les plantes
La principale source de metaux pour les plantes se situe au niveau du sol, bien que 1'assimilation des metaux suite a une deposition au niveau du feuillage puisse egalement se produire. Dans le sol, 1'absorption d'un metal par les plantes est grandement influencee par certains facteurs comme le pH, le regime hydrique, la composition en argile et en matiere organique, la capacite d'echange cationique et la presence d'autres elements traces (Kabata-Pendias et (Kabata-Pendias, 2000). Les mecanismes d'acquisition different selon les metaux et peuvent §tre passifs (Ni et Pb), mais preferablement actifs (Cu, Mo et Zn) ce qui procure a la plante un meilleur controle de son homeostasie (Kabata-Pendias et Pendias, 2000). Tout comme pour les microorganismes, plusieurs metaux sont essentiels chez les plantes. Parmi les plus importants se retrouvent le Co, le Cu, le Fe, le Mn, le Mo, le Ni, le Se et le Zn. Parmi ceux-ci, le Cu et le Zn sont particulierement necessaires puisqu'ils entrent dans la composition de plusieurs enzymes et de qiielques proteines (Rout et Das, 2003; Hall, 2002). A l'oppose, le Pb, le Cd et le Hg ne sont pas essentiels chez les plantes (Clemens, 2001). Evidemment, les metaux
essentiels presents en concentrations tres elevees deviendront toxiques tout comme les metaux non essentiels.
Generalement, les plantes sont plus sensibles a un manque d'elements traces qu'a l'exces de ces derniers. Toutes les plantes possedent et partagent une resistance de base aux metaux alors que certaines les tolerent en plus grandes concentrations (Clemens, 2001; Kabata-Pendias et Pendias, 2000). Quelques especes vegetales . peuvent meme hyperaccumuler les metaux. L'hyperaccumulation d'un metal se definit par la concentration de ce metal retrouvee chez la plante par rapport au poids sec de 1'echantillon de tissus vegetaux (Clemens, 2001). Par definition, les plantes hyperaccumulatrices, lorsqu'elles sont utilisees en phytoextraction, ont la capacite d'accumuler plus de 1000 mg/kg d'un metal par poids sec (Gleba et ah, 1999; Roy et al., 2005). Chez les plantes sensibles aux metaux, la toxicite peut etre reliee a plusieurs causes : 1- Tinhibition de l'activite ou la deformation structurale d'une proteine, 2- la formation de radicaux libres ou d'especes reactives de Poxygene et 3- la substitution du metal par un analogue toxique dans la proteine cible (Vanassche et Clysters, 1990; Hall, 2002; Clemens, 2001). Afin de prevenir ou de contrer les effets toxiques des metaux, la plante met en place des strategies de defense plutot efficaces. Ces mecanismes cellulaires menant a la detoxification ou a la tolerance des metaux chez les plantes comprennent: 1- la restriction du mouvement des metaux par les mycorhizes, 2- Pattachement des metaux a la membrane cellulaire ou aux exsudats racinaires, 3- la reduction de Tinflux au niveau de la membrane plasmique, 4- l'efflux actif provenant de l'apoplaste, 5- la presence d'agents chelateurs dans le cytosol, 6- la reparation active de la membrane plasmique en conditions de stress, 7- le transport de complexes metalliques (ex.: phytochelatines) vers les vacuoles et 8- le transport etTaccumulation des metaux dans les vacuoles (Hall, 2002). Les plantes possedent done plusieurs strategies pour s'adapter a la presence de metaux dans leur environnement.
4.3 L'effet des metaux sur les symbioses entre les plantes et les microorganismes fixateurs d'azote
i •
Les deux principales symbioses fixatrices d'azote comprennent celle entre les Rhizobium et les legumineuses et celle entre Frankia et les plantes actinorhiziennes. Toutes deux ont une grande importance environnementale pour le cycle de l'azote et sont reliees a des plantes dont la valeur economique est considerable. II est done important de comprendre quels seront les impacts des metaux sur ces deux symbioses.
Les etudes anterieures concernant l'effet toxique des metaux sur les symbioses etaient souvent realisees avec des souches bacteriennes differentes, des especes vegetales variees ou encore dans des conditions non similaires. Toutes ces variables rendent difficile la comparaison des resultats qui sont souvent contradictoires. L'effet toxique des metaux sur la diminution de la fixation d'azote des Rhizobium serait du a l'extinction graduelle de ces bacteries dans le sol puisque ce genre bacterien est plutot sensible aux metaux (Broos et al., 2005). Des etudes ont demontre que plusieurs isolats de Rhizobium leguminosarum bv. trifolii (85%) provenant d'un sol contamine en metaux avaient perdu leur capacite a fixer l'azote en symbiose (Castro et ah, 1997). D'autres travaux en sont venus a des conclusions legerement differentes, e'est-a-dire que des Rhizobium isoles suite a une contamination aigue en metaux conservaient leur pouvoir fixateur d'azote. Par contre, la capacite a etablir la symbiose etait affectee (Carrasco et al., 2005). La presence de plantes notes dans les sols d'ou proviennent les isolats exposes aux contaminations semble egalement avoir un impact sur la capacite fixatrice d'azote de ces souches. Lors d'une etude realisee en terrains contamines, la presence de Rhizobium capables de fixer l'azote a ete observee dans les sols ou les plantes notes pouvaient croitre et a l'oppose, en l'absence de plantes notes, la capacite de nodulation du sol etait abolie (Smith, 1997). Finalement, des travaux realises en conditions controlees ou les metaux etaient ajoutes sous forme de solutions ont demontre que 1'inoculation de pois chiches suivie d'une exposition au Cd et au Pb provoquait une diminution du nombre de nodules chez les plants (Wani et al.,
2007). II a aussi ete demontre que l'exposition de la symbiose entre la souche R. leguminosarum RP 5 et le pois au Cu diminuait le nombre de nodules par plant, alors que l'exposition au Cd et au Cr augmentait le nombre de nodules par plant (Wani et al., 2008). Ce dernier resiiltat est non seulement surprenant, mais aussi en contradiction avec celui obtenu auparavant par la meme equipe pour une autre espece vegetale.
Chez Frankia, bien qu'habituellement plutot toxique, le Co serait essentiel dans la fixation de l'azote puisqu'il sert a la formation d'analogues de la vitamine B12 chez le symbionte (HussDanell, 1997). Malgre le fait que ce metal profite a la symbiose, la plupart des etudes s'interessant aux metaux ont ete realisees dans le but d'etudier l'impact negatif des metaux sur la symbiose entre Frankia et les plantes actinorhiziennes. Dans le cadre de ces etudes, differents parametres sont evalues. Parmi les plus communs se retrouvent l'etablissement de la symbiose, le developpement de la symbiose et l'efficacite de la symbiose. L'etablissement de la symbiose refere au succes de colonisation de Frankia envers sa plante note. Cette colonisation est quantifiable en determinant le nombre de nodules retrouves sur les racines de l'hote; ce qui correspond au nombre d'infections ayant menees a un etablissement reussi de la symbiose. Le developpement de la symbiose sert a determiner si les nodules croissent bien et repondent aux besoins en azote de l'hote. Le developpement de la symbiose est mesure par le poids des nodules s'etant developpes sur une plante hote. L'efficacite de la symbiose s'interesse a savoir si la fixation d'azote est toujours active a Pihterieur d'un nodule. La methode de la reduction de l'acetylene est la plus commune pour verifier l'activite de l'enzyme nitrogenase. Selon les etudes completees a ce jour, il semblerait que Frankia soit sensible a la presence de contaminants inorganiques dans son environnement et que cela influence sa capacite a etablir la symbiose. En effet, des souches de Frankia pre-cultivees en presence d'eau contaminee avaient plus de difficulte a entrer en symbiose avec Casuarina equisetifolia que si elles etaient cultivees dans un milieu de culture approprie puis arrosees avec l'eau contaminee. Dans ce dernier cas, la symbiose s'etablissait correctement (Sayed, 2003). Toujours chez C. equisetifolia, il a ete demontre que la nodulation etait partiellement inhibee suite a l'ajout de MgCb, MnCb, CaCb et de CdCl2 dans le sol. Le CdCk etait le metal
ayant 1'impact negatif le plus important sur l'etablissement de la symbiose (Selim et Schwencke, 1995). L'etablissement de la symbiose entre Frankia et Discaria americana etait aussi affecte negativement en presence de Zn, mais le developpement (biomasse) des nodules suivait les demandes en azote de la plante. Done, le faible nombre de nodules etait compense par une plus grande biomasse nodulaire (Cusato et al., 2007). Les resultats tires des etudes ci-haut, supportent l'hypothese selon laquelle les metaux affectent davantage l'etablissement de la symbiose que son developpement et son efficacite. Par contre, les etudes presentees ci-dessous en viennent a des conclusions differentes: la concentration du metal ajoute semble avoir une influence sur les resultats obtenus. Par exemple, l'exposition de A. glutinosa a une concentration 0,025 mM de NiCi2, bien que reduisant la croissance des plants, n'a pas affecte significativement la nodulation et l'activite nitrogenase a meme double. Par contre, l'exposition a des concentrations plus elevees de NiCl2 a reduit autant la nodulation que l'activite nitrogenase et a meme entraine la mort des plants a de hautes concentrations (0,5 mM NiGfe) (Wheeler etal., 2001).
Certaines etudes arrivent cependant a des conclusions plus positives et encourageantes demontfant la persistance de Frankia spp. dans les sols contamines. En effet, plusieurs isolats de Frankia spp. provenant de sols contamines en metaux et en composes organiques etaient toujours en mesure de fixer 1'azote et ce malgre l'absence de leurs plantes botes sur le site (Ridgway et al., 2004). La meme equipe a egalement employe A. incana dans des essais de capture de souches de, Frankia spp. sur differents sols contamines et ont demontre que chaque site possedait une communaute de Frankia spp. differente (Ridgway et al., 2004). Finalement, l'exposition de A. rubra prealablement inocules a des biosolides contamines en metaux a demontre que la relation entre A. rubra et Frankia n'etait pas affectee par les hautes concentrations de metaux (Gaulke et al., 2006). Suite a l'obtention de resultats si divergents d'une souche a l'autre, d'une espece vegetale a l'autre et dans des conditions experimentales diverses, il est de toute evidence critique de realiser des essais en conditions controlees afih de mieux cerner les mecanismes sous-jacents aux phenomenes observes. Une fois ces mecanismes mis en evidence, il serait plus facile d'expliquer pourquoi une souche bacterienne
ou une espece vegetale ainsi que leur symbiose se montrent plus tolerantes ou sensibles a la presence des metaux dans ces divers environnerhents. De telles decouvertes permettraient ensuite 1'amelioration des connaissances menant au perfectionnement de phytotechnologies en choisissant judicieusement une souche bacterienne et une espece vegetale pour une contamination donnee. Ces connaissances permettront egalement le developpement d'outils de prediction de l'effet de l'activite industrielle sur le cycle biogeochimique deTazote dans les environnements affectes.
5. Hypothese et objectifs du projet de recherche
L'hypothese de ce projet de recherche etait que Frankia spp. et sa symbiose tolerent les metaux dans leur environhement et pourraient eventuellement servir en revegetalisation et/ou en phytoremediation de sites contamines par les metaux lourds. Afin de parvenir a demontrer cette hypothese, deux objectifs ont ete elabores. Premierement, il fallait etablir la tolerance de Frankia spp. face aux metaux qui pourraient se retrouves dans son ehvironnement (Chapitre 1). Cet objectif a ete realise en developpant une methode de criblage miniaturisee et quantitative suivi par revaluation de la tolerance de cinq souches de Frankia sp. envers douze metaux et metalloi'des. Deuxiemement, il fallait evaluer la tolerance de la symbiose entre une souche de Frankia sp. et une de ses plantes hotes (Chapitre 2). Pour ces etudes, la souche Frankia ACN14a, dont la tolerance aux metaux avait ete prealablement determinee, a ete utilisee en combinaison avec l'aulne noir {A. glutinosa). Le developpement d'une methode de culture hydroponique des aulnes noirs en conditions gnotobiotiques s'est avere necessaire afin de completer cet objectif. Par la suite, l'impact de douze metaux sur la symbiose entre Frankia ACN14a et l'aulne noir a ete evalue grace a differents parametres.
CHAPITRE 1
HIGH-THROUGHPUT SCREENING OF MICROBIAL ADAPTATION TO ENVIRONMENTAL STRESS
1. Reference de Particle et apport de Pauteur
Le titre de l'article est le suivant: High-throughput screening of microbial adaptation to environmental stress et les auteurs sont P.-A. Belanger, J. Beaudin et S. Roy. L'apport du premier auteur dans cette publication est majeur puisqu'il comprend le developpement de la methode en entier en plus du criblage de la majorite des souches pour la determination de leur tolerance a plusieurs metaux. Quelques criblages ont egalement et6 realises grace a l'aide de stagiaires du baccalaureat en microbiologic De plus, la redaction de cet article a ete realisee par 1'auteur principal. Julie Beaudin a participe a cet article en aidant a quelques manipulations ainsi qu'en faisant un grand travail de revision du manuscrit. Finalemerit, Sebastien Roy a ete implique dans la supervision des travaux ainsi que dans la revision du manuscrit.
2. Originalite du travail et apport a la science
L'originalite du travail presente dans cet article provient du developpement d'une methode nouvelle et performante amenant une solution au probleme de determination de la tolerance de microorganismes dont la croissance est fastidieuse face a differents contaminants environnementaux. Cette methode vient palier au manque d'efficacite d'autres methodes traditionnelles parfois mal adaptees pour des microorganismes filamenteux ou a croissance lente. Avec la methode developpee, il existe maintenant un dutil versatile et surtout quantitatif permettant de mesurer l'activite metabolique residuelle des microorganismes suite a leur
exposition a un stress. L'autre avantage majeur de la technique reside dans sa miniaturisation qui permet d'evaluer plusieurs concentrations (un gradient) du contaminant d'interet avec un grand nombre de replicats. Bien qu'elaboree dans le but de determiner la toxicite de certains composes sur les microorganismes, cette methode s'est aussi averee efficace a determiner, par exemple, les impacts positifs de sources de carbone ou de divers milieux de culture. Les adaptations possibles de cette methode sont tres nombreuses et ne dependent que de 1'imagination des scientifiques qui l'emploient pour se realiser. Ces travaux sont egalement les premiers, a notre connaissance, a adresser de maniere quantitative et de facon aussi large l'effet des metaux lourds sur des microorganismes filamenteux comme Frankia.
3. Resume de Particle en francais
Nous avons developpe une methode de criblage visant a determiner quantitativement la tolerance de microorganismes filamenteux et/ou ayant une croissance lente face a differents stress dans un format de microplaque de 96 puits. Cette methode permet de tester des gradients de plusieurs composes avec un nombre suffisant de replicats pour generer des analyses statistiques robustes. La viabilite des microorganismes a ete quantifiee en utilisant le MTS, un sel de tetrazolium qui, une fois reduit, produit un formazan soluble dans l'eau qui permet une lecture spectrophotometrique directe. Le microorganisme modele utilise tout au long du developpement etait Pactinobacterie du genre Frankia. L' article qui suit presente les etapes successives du developpement de la methode, ce qui donnera a d'autres chercheurs un bon indice des parametres auxquels s'attarder afin d'adapter la methode pour leurs propres besoins. Cet article se terminera par la presentation des resultats de la tolerance de 5 souches de Frankia sp. face a 12 metaux. Cela represente un criblage de 600 conditions avec de multiples replicats (concentrations x metaux x souches).
4. Manuscrit de 1'article
4.1 Abstract
We developed a microplate, high-throughput, screening method aimed at determining quantitatively the tolerance of filamentous and/or slow-growing microorganisms to various stresses. The method allowed testing of compound gradient with enough replicates to generate statistically robust results. The microorganisms' viability was quantified using MTS; a tetrazolium salt that once reduced produces a water-soluble formazan that allows direct spectrophotometric measurement. The microorganisms used throughout the development process were the actinobacteria Frankia spp.. We present the stepwise development of the method, allowing other researchers to consider which issues they should address to adapt this method to their needs. The paper ends with the presentation of heavy metal tolerance for 5 strains of Frankia and 12 heavy metals. This represents a multi-replicate screen of 600 conditions (concentration x metal x strain).
Keywords : Actinobacteria Frankia, tetrazolium salt, heavy metals, screening
4.2 Introduction c
Adaptation to environmental changes is essential for microbial survival and proliferation. Environmental changes will shift the balance of complex microbial communities by favoring some populations, while restricting others, through mechanisms such as microbial competition (for nutrients), antibiosis and by the selection of those microorganisms best adapted to environmental stress. Thus, much research in microbial ecology focuses on the evolution of microbial populations (both taxonomic and functional groups) submitted to fluctuations in environmental conditions 1_?.
Studying how microorganisms react to environmental stress and stimuli allows us to better understand microbial ecology in specific natural environments subjected to inherent fluctuations. At the same time, measuring the absolute microbial tolerance to environmental stress allows us to design biotechnological applications that exploit these capabilities. Table 1 presents a selection of current research fields that can benefit from the screening method we present herein, and where microbial response studies are a cornerstone.
The standardized high-throughput method we developed was aimed mostly at studying symbiotic, nitrogen-fixing bacteria that belong to genus Frankia. This method is, however, applicable to many other microorganisms. As importantly, this method provides an efficient experimental device that allows us to measure the microbial component response over a gradient of a particular environmental parameter (e.g. a concentration gradient of heavy metals). Microbial response in the face of such a gradient is critical since it allows us to rigorously correlate cause and effect, and determine as well any threshold effects. Unfortunately, and even before considering the necessity of replicates, studying the effect of hundreds of environmental conditions can be fastidious, and sometimes impossible. Our method not only resolves this, but generates statistically robust data.
Frankiae are filamentous slow-growing actinobacteria found in soil. They can fix atmospheric nitrogen as saprophytes or in symbiosis with over 200 plant species24"26. Bacteria belonging to
the genus Frankia are as critical as Rhizobia to the biogeochemical cycling of nitrogen on Earth; it is- estimated that frankiae could be responsible for as much as 25% of the global nitrogen fixation 27. Since frankiae are directly exposed to soil stresses such as high levels of
heavy metals, hydrocarbons, salts and others, studying their tolerance and sensitivity is important and allows us to 1- estimate their survival under these harsh conditions 9, and