Toxicité aigüe du cadmium chez Donax trunculus L. : variation de la composition biochimique des gonades en fonction du sexe et de l’activité sexuelle au cours de l’exposition aigüe et de la dépuration
191
0
0
Texte intégral
Figure
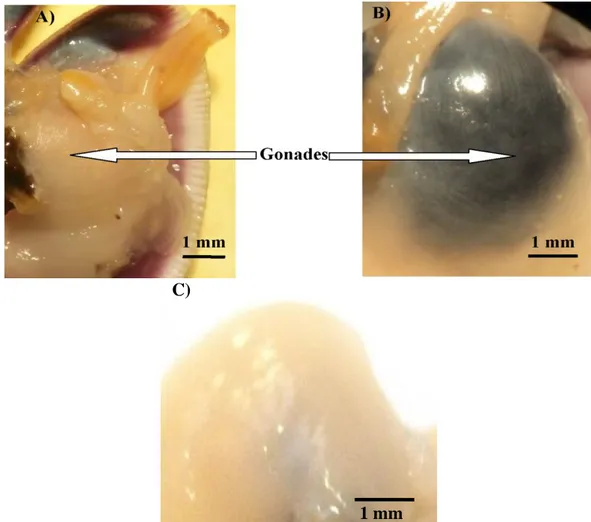


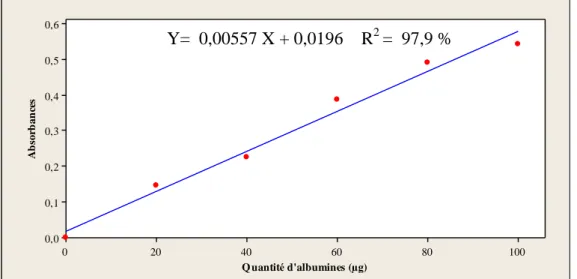
+7
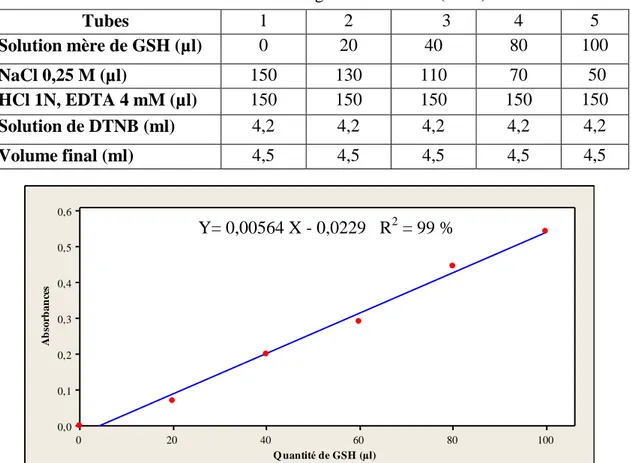
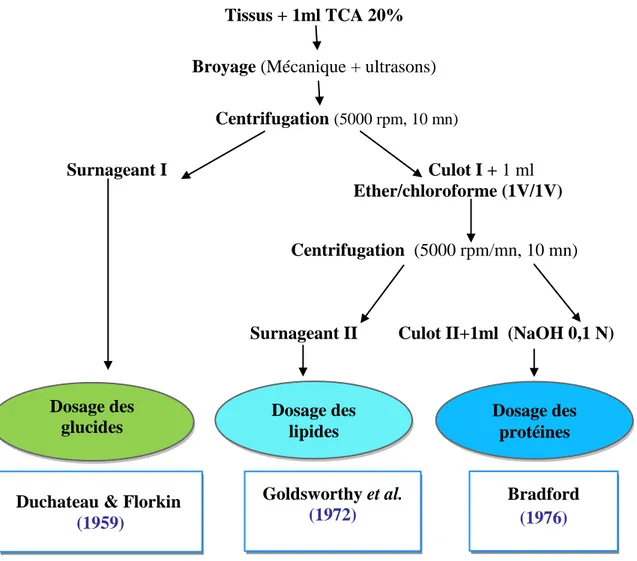

Documents relatifs
