© Joanie Lemieux, 2019
Validation d'échantillonneurs d'air et biais sur la
diversité
Mémoire
Joanie Lemieux
Maîtrise en microbiologie - avec mémoire
Maître ès sciences (M. Sc.)
Validation d’échantillonneurs d’air et biais sur la diversité
Mémoire
Joanie Lemieux
Sous la direction de :
iii
Résumé
Différents types d’échantillonneurs d’air existent pour récolter les bioaérosols. Ils ont tous leurs avantages et leurs inconvénients, mais de ces types, un en particulier est susceptible d’introduire des biais dans le traitement et l’analyse des résultats. Les échantillonneurs de type liquide voient une portion de leur liquide d’échantillonnage s’évaporer pendant leur fonctionnement, ce qui favoriserait la perte de bioaérosols par ré-aérosolisation ou leur concentration dans le liquide. Peu de connaissances sont acquises sur la ré-aérosolisation, la concentration et leurs effets sur les résultats. Cette étude avait pour but de documenter comment l’évaporation du liquide de collecte impacte l’échantillonnage de l’air. Des expérimentations in vitro, où les récipients de collection de deux échantillonneurs liquides (Coriolisµ® et BioSampler®) étaient inoculés avec des consortiums bactériens connus, ont permis de conclure que la ré-aérosolisation et la concentration des bactéries sont des phénomènes complexes. En effet, ils sont difficilement prédictibles et semblent influencés par l’évaporation, le genre bactérien, l’hydrophobicité de la membrane bactérienne, l’interaction avec les autres bactéries, la composition du liquide de collection, le débit et le mécanisme de capture de l’échantillonneur. De plus, des expérimentations en milieu naturel ont permis de comparer la diversité récoltée par les échantillonneurs de type liquide et de type filtre, par méthodes de séquençage à haut débit. Une des particularités de cette étude était qu’une cassette contenant un filtre était branchée à la sortie d’air du BioSampler® pour récolter les bactéries ré-aérosolisées pendant l’échantillonnage. Plusieurs genres bactériens sont totalement ré-aérosolisés du récipient de collection du BioSampler®. Plus de la moitié des genres bactériens récoltés par le Coriolisµ® diffèrent de ceux du BioSampler®, et inversement. Les échantillonneurs filtres ont tous deux récolté une diversité bactérienne très similaire. Ces résultats constituent un apport important au domaine scientifique puisqu’ils prouvent les biais potentiels induits par les échantillonneurs de type liquide.
iv
Abstract
Different types of air samplers are available to harvest bioaerosols. They all have their advantages and disadvantages, but of these types, one in particular is likely to introduce bias in the treatment and analysis of the results. Liquid-based samplers see a portion of their collection fluid evaporate during operation, which would favor either the loss of bioaerosols by re-aerosolization or their concentration in the fluid. Very little knowledge is known about re-aerosolization, concentration and their effects on results. The main purpose of this study was to document how the evaporation of the collection fluid impacts air sampling. In vitro experiments, in which collection vessels from two liquid samplers (Coriolisμ® and BioSampler®) were inoculated with known bacterial consortia, concluded that re-aerosolization and concentration are complex phenomena. Indeed, they are difficult to predict and seem influenced by evaporation, the bacterial genus, the hydrophobicity of the bacterial membrane, the interaction with other bacteria, the composition of the collection fluid, the flow and capture mechanism of the sampler. In addition, experiments in a natural environment have made it possible to compare the diversity harvested by the liquid-based and filter-based samplers by high throughput sequencing methods. One of the peculiarities of this study was that a cassette containing a filter was connected to the BioSampler® air outlet to collect the re-aerosolized bacteria during sampling. The results are unequivocal, several bacterial genera are totally re-aerosolized from the BioSampler® collection vessel. More than half of the bacterial genera harvested by the Coriolisμ® differ from those of the BioSampler®, and vice versa. The filter samplers both harvested a similar bacterial diversity. These results are an important contribution to the scientific field since they prove the biases induced by liquid type samplers.
v
Table des matières
Résumé ... iii
Abstract ... iv
Table des matières ... v
Liste des figures ... vii
Liste des tableaux ... ix
Liste des abréviations et des sigles ... x
Remerciements ... xi Avant-propos ... xii Introduction ... 1 Les bioaérosols ... 1 L’échantillonnage de l’air ... 5 Échantillonneurs d’air... 5
Facteurs influençant la ré-aérosolisation ... 15
Facteurs influençant l’échantillonnage ... 22
Méthodes de traitement et d’analyse des échantillons ... 23
Méthodes de traitement ... 23
Méthodes d’analyses ... 25
Problématique, hypothèse et objectifs des travaux de recherche ... 28
Problématique ... 28
Hypothèse ... 28
Objectifs ... 28
Chapitre 1 : Re-aerosolization in liquid-based air samplers induces bias in bacterial diversity ... 30
1.1 Résumé ... 31
1.2 Abstract ... 32
1.3 Introduction ... 33
1.4 Material and methods ... 36
1.4.1 In vitro experiments ... 36
vi
1.4.3 Statistical Analysis ... 43
1.4.4 Data Availability ... 45
1.5 Results ... 46
1.5.1 In vitro Experiments ... 46
1.5.2 Sequencing from Field Assays ... 48
1.6 Discussion ... 56
1.7 Conclusion ... 60
1.8 Acknowledgments ... 60
1.9 Supplemental Information ... 61
1.10 Bibliography ... 62
Chapitre 2 : Matériel et méthodes additionnels non présentés dans l’article ... 70
2.1 Conditions de croissance des bactéries de l’étude ... 70
2.2 Détermination de l’hydrophobicité de la paroi cellulaire ... 70
2.3 Détermination de l’auto-agrégation bactérienne ... 71
2.4 Échantillonnage in vitro : liquide de collection supplémenté de glycérol et de saccharose ... 71
Chapitre 3 : Résultats additionnels non présentés dans l’article ... 72
3.1 Pourcentages d’hydrophobicité relative ... 72
3.2 Pourcentages d’auto-agrégation relative ... 73
3.3 Liquide de collection supplémenté de glycérol et de saccharose ... 74
3.4 Analyse taxonomique ... 81 3.4.1 Échantillonneurs liquides ... 81 3.4.2 Échantillonneurs filtres ... 82 Chapitre 4 : Discussion ... 84 Conclusion ... 87 Bibliographie ... 88 Annexe ... 104
vii
Liste des figures
Figure 1. Processus d’assèchement d’une gouttelette en noyau de gouttelette. Figure 2. Diamètre aérodynamique d’une particule irrégulière.
Figure 3. Cassette 37 mm.
Figure 4. Le SASS 3100 Dry Air Sampler et son filtre électrostatique. Figure 5. BioSampler®.
Figure 6. Trajet des particules selon leur inertie dans le récipient de collection du Coriolisµ®. Figure 7. Schéma du récipient conique du Coriolisµ®.
Figure 8. Bulle éclatant en plusieurs gouttelettes à la surface de l’eau. Figure 9. Schéma de la paroi cellulaire des bactéries à Gram positif. Figure 10. Schéma de la paroi cellulaire des bactéries à Gram négatif.
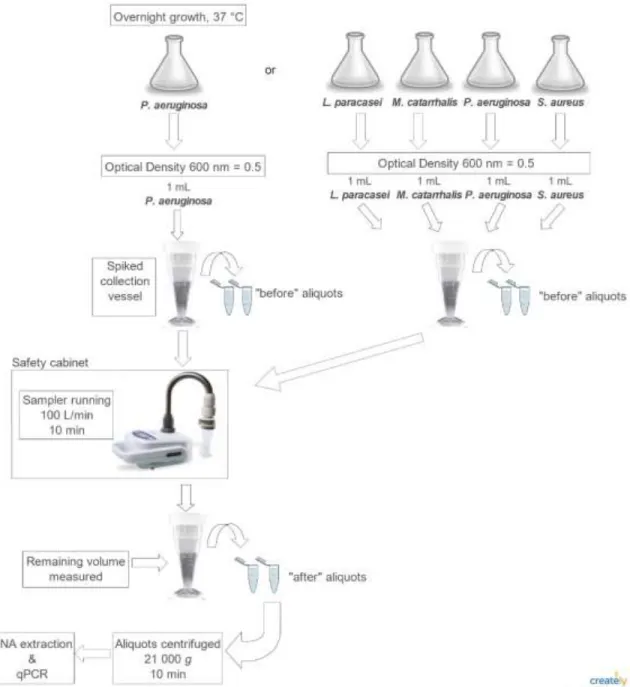
Figure 1-1. Experimental protocol for sampling each of the four bacteria strains and the bacterial consortium using Coriolisµ® and the BioSampler®.
Figure 1-2. The 37 mm cassette filter connected to the BioSampler®.
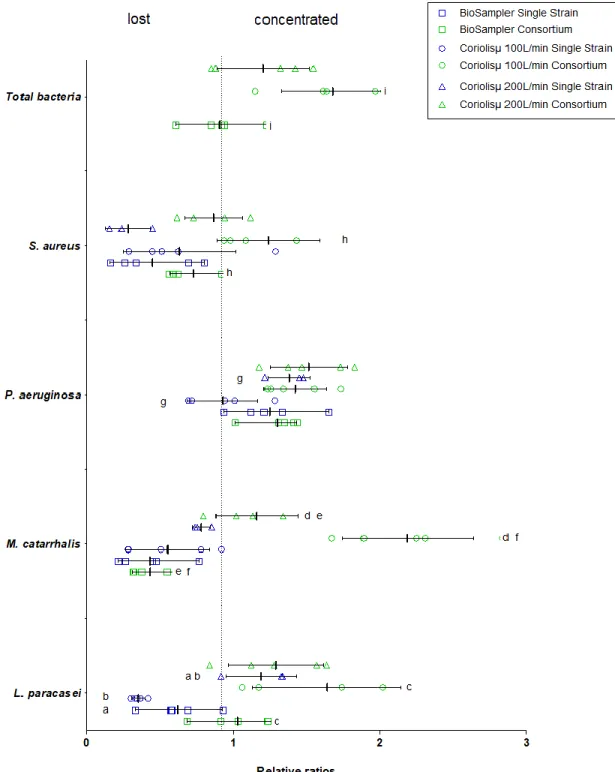
Figure 1-3. Relative ratios of genome copy numbers of each single strain and the bacterial consortium when added to the BioSampler® and the Coriolisµ® collection fluid.
Figure 1-4. Comparisons of alpha-diversity metrics of observed OTUs (a) and Chao1 (b) for natural bioaerosols collected with two sampler types (liquid and filter).
Figure 1-5. Principal-coordinate analysis of air samples collected with two different types of air samplers.
Figure 1-6. Relative abundance of the bacterial phyla collected using liquid- and filter-based samplers.
Figure 1-7. Relative abundances of bacterial genera collected using liquid and filter type samplers.
Figure 1-8. Venn diagram of the genera (relative abundance) collected by the BioSampler® and those re-aerosolized from the collection fluid captured by the BioSampler® cassette. Figure 1-9 The 20 most abundant genera collected with the air samplers used in this study. Figure 3-1. Pourcentage d’hydrophobicité relative des quatre bactéries utilisées dans cette étude.
Figure 3-2. Pourcentage d’auto-agrégation relative des quatre bactéries utilisées dans cette étude.
viii
Figure 3-3. Ratios relatifs de L. paracasei lorsqu’inoculé dans le Coriolisµ® et le BioSampler® avec comme liquide de collection du PBS 1X (référence), du PBS 1X + Glycérol 20% et PBS 1X + Saccharose 20%.
Figure 3-4. Ratios relatifs de M. catarrhalis lorsqu’inoculé dans le Coriolisµ® et le BioSampler® avec comme liquide de collection du PBS 1X (référence), du PBS 1X + Glycérol 20% et PBS 1X + Saccharose 20%.
Figure 3-5. Ratios relatifs de P. aeruginosa lorsqu’inoculé dans le Coriolisµ® et le BioSampler® avec comme liquide de collection du PBS 1X (référence), du PBS 1X + Glycérol 20% et PBS 1X + Saccharose 20%.
Figure 3-6. Ratios relatifs de S. aureus lorsqu’inoculé dans le Coriolisµ® et le BioSampler® avec comme liquide de collection du PBS 1X (référence), du PBS 1X + Glycérol 20% et PBS 1X + Saccharose 20%.
Figure 3-7 Ratios relatifs des génomes totaux lorsque le mélange bactérien est inoculé dans le Coriolisµ® et le BioSampler® avec comme liquide de collection du PBS 1X (référence), du PBS 1X + Glycérol 20% et PBS 1X + Saccharose 20%.
Figure 3-8. Diagramme de Venn des genres récoltés dans le liquide de collection du BioSampler® et du Coriolisµ®.
Figure 3-9. Diagramme de Venn des genres récoltés par la cassette 37 mm et le SASS 3100. Figure A-1. Nombres d’UTO obtenues par le Coriolisµ®, le SASS 2300 et le SASS 3100 dans un environnement de centres de traitements des eaux usées (CTEU).
Figure A-2. Nombres d’UTO obtenues pour le Coriolisµ® et le SASS 3100 dans un environnement de bio méthanisation.
Figure A-3. Nombres de copies d’ARN 16S par mètre cube d’air dans des centres de traitement des eaux usées entre le Coriolisµ®, SASS 2300 et le SASS 3100.
Figure A-4. Nombres de copies d’ARN 16S par mètre cube d’air en environnement de bio méthanisation entre le Coriolisµ® et le SASS 3100.
ix
Liste des tableaux
Tableau 1. Microorganismes communs causant des infections ou des problèmes de sensibilisation.
Table 1-1. Bacteria examined in this study. Table 1-2. Primers and probes used in this study.
x
Liste des abréviations et des sigles
ADNr : Acide désoxyribonucléique ribosomique
BHI : Infusion cœur-cervelle (Brain Heart Infusion broth) LPS : Lipopolysaccharide
PBS : Solution saline tamponnée au phosphate (Phosphate Buffered Saline)
PUM : tampon phosphate sulfate d’urée et de magnésium (phosphate urea magnesium sulfate
buffer)
qPCR: Réaction en chaîne par polymérase en temps réel (Real-time Polymerase Chain
Reaction)
TSB : Bouillon de soja tryptique (Tryptic Soy Broth) UTO : Unité Taxonomique Opérationnelle
xi
Remerciements
Chèvres festives Accomplissements Remarquable Organique Luxuriante Iconique Nourricière Enigmatique Durable Unificatrice Cohésive Hors du commun Attachante Incomparable Nomenclature Eblouissante Merci pour tout, infiniment.Merci Marc pour ton support technique et moral et les quelques blagues que tu lances ici et là.
Merci Val et Nath pour votre support technique, votre présence et votre bonne humeur. Merci à Marie-Eve, Maria, Jodelle, Karine, Nathan et Jon d’avoir partagé une partie de ces deux années.
Merci à Michel et Yan pour votre soutien et votre aide annuelle.
Merci à ma p’tite Pameloooooooooo de me faire rire autant et de toujours me déranger pour une bonne raison. Je suis choyée de t’avoir dans ma vie, Je t’aime.
Merci au séquençage de m’avoir ouvert les yeux sur la magnificence d’Hamza, Je t’aime. Merci à Sarah-Jeanne, meine pretté, de toujours être là pour moi et de rendre ma vie tellement plus belle. Je t’aime vraiment beaucoup.
Merci à Steph ma cop pref d’être présente depuis si longtemps dans les bons et mauvais moments, Je t’aime trop.
Merci à mes parents d’exister et d’être là.
xii
Avant-propos
Ce mémoire est divisé en six chapitres. Premièrement, une introduction présente les notions importantes à l’aérobiologie, aux échantillonneurs d’air et à leurs limitations, ainsi qu’aux différentes méthodes de traitements et d’analyses. Le chapitre 2 comporte la majorité du contenu des sections méthodologie et résultats, présenté sous forme d’article scientifique ayant été soumis le 10 avril 2019 dans le journal Applied and Environmental Microbiology. L’article inséré dans ce mémoire varie de la version soumise pour satisfaire aux exigences du mémoire. La section des résultats se trouve après la section matériel et méthodes, le nom des figures a été modifié, ainsi que le format des références. Les chapitres 3 et 4 contiennent respectivement les méthodes et les résultats non présentés dans l’article scientifique. Le chapitre 5 est une discussion générale des résultats présentés dans l’article et le chapitre quatre. Finalement, le chapitre 6 présente les conclusions générales de ce projet de maîtrise ainsi que des perspectives intéressantes à la poursuite de ce sujet de recherche.
1
Introduction
Les bioaérosols
Un aérosol représente toute particule solide ou liquide en suspension dans l’air d’un diamètre variant de 0,002 microns à 200 microns (Hinds, 1999). Dès que l’origine de l’aéorosol est biologique, il s’agit d’un bioaérosol. Ceux-ci représentent toutes particules d’origine biologique en suspension dans l’air pouvant contenir des microorganismes vivants ou morts, leurs fragments et composantes tels que des bactéries, des moisissures, des virus, des spores et des toxines. Ce sont des particules hétérogènes, elles peuvent donc contenir d’autres particules biologiques provenant de fragments de plantes (pollen), d’animaux (acariens, protéines aviaires) aussi bien que des composés non biologiques (poussières, sels et minéraux) (Schenker et al., 1998; Hinds, 1999; Miller et al., 1999; Stetzenbach, 2007).
Les bioaérosols proviennent de diverses sources telles que les plantes, les animaux, le sol et l’eau et se retrouvent en suspension dans l’air à la suite de divers processus d’aérosolisation. Ces processus peuvent être naturels comme par exemple, l’évaporation et le bullage d’un liquide, l’action de parler, d’éternuer, de tousser ou la friction de deux surfaces. Certains processus sont industriels tels que l’élevage d’animaux par le brassage de la litière et la moulée, le bullage et l’agitation mécanique de certaines stations dans les usines de traitement des eaux usées, le retournement du compost en andain, l’écorçage des arbres dans les scieries ou l’ensachage de la tourbe. La taille d’un bioaérosol varie, entres autres, selon le processus d’aérosolisation et a une influence directe sur son comportement subséquent dans l’air (Schenker et al., 1998; Hinds, 1999; Miller et al., 1999; Stetzenbach, 2007).
2
Un bioaérosol peut être sec ou humide dépendant de sa source et de sa durée de séjour dans l’air. Un bioaérosol est sec s’il provient d’une source peu humide et est mis en suspension dans l’air sous forme de poussière. Un bioaérosol humide, nommé gouttelette, de taille initiale inférieure à 100 µm, s’assèche graduellement pour devenir un noyau de gouttelette et adopter subséquemment le comportement d’un bioaérosol sec (Figure 1). L’humidité relative ambiante peut ralentir ou accélérer la vitesse et le niveau d’assèchement des gouttelettes et donc influencer la taille finale du noyau de gouttelette (Hinds, 1999; Stetzenbach, 2007; Verreault et al., 2008).
Figure 1 Processus d’assèchement d’une gouttelette en noyau de gouttelette. Figure tirée de Verreault et al., 2008.
Tout bioaérosol de taille supérieure à 100 µm, qu’il soit sec ou humide, va sédimenter sur une surface peu de temps après son aérosolisation si les conditions aérauliques le permettent. Le comportement de ce type de bioaérosol est principalement régi par la force gravitationnelle. Une des lois de Stokes décrit ce phénomène de déposition des particules en fonction de la gravité : une particule en suspension dans l’air est soumise simultanément à la force gravitationnelle (qui l’attire vers le bas) et à la force de résistance de l’air (égale et inverse à la gravité) ce qui résulte en une vitesse de chute (déposition) constante. Par exemple, les particules de 100 µm ont une vitesse de déposition de 2,49 x 10 -1 m/s, celles de 10 µm ont une vitesse de déposition de 3,06 x 10 -3 m/s, alors que celles de 0,001 µm ont une vitesse de déposition de 6,75 x 10 -9 m/s. Les petites particules sont sous l’influence du flux d’air et non uniquement de la force gravitationnelle comme c’est le cas pour les plus grosses particules (Xie et al., 2007; Engelstaedter et al., 2006).
3
La forme d’une particule influence aussi son comportement dans l’air. Les bioaérosols ne sont pas parfaitement sphériques, ils ont plutôt des formes irrégulières. La description de leur comportement se fait grâce au concept de diamètre aérodynamique. Il s’agit du diamètre qu’aurait une particule homogène fictive, de forme sphérique, si elle avait la même vitesse de déposition que la particule irrégulière et la masse volumique de l’eau (1 g/cm3) (Figure 2). Par exemple, si une particule irrégulière et une particule sphérique d’eau d’un diamètre de 8,6 µm ont une vitesse de déposition de 2,2 m/s, le diamètre aérodynamique équivalent de la particule d’eau sera établi à 8,6 µm. La particule irrégulière se comportera donc comme une particule d’eau de 8,6 µm, peu importe sa forme, sa densité et sa taille (Hinds, 1999). Il devient alors possible de prédire le comportement des bioaérosols dans l’air.
Figure 2 Diamètre aérodynamique d’une particule irrégulière. Figure adaptée et traduite de Hinds, 1999, par Caroline Duchaine, dans le cadre du cours Bioaérosols et aérobiologie de l’Université Laval.
4
De ce fait, les petites particules, telles que les noyaux de gouttelettes, peuvent demeurer en suspension dans l’air pendant des temps relativement longs et parcourir des distances variables avant de se déposer sur une surface. Les distances parcourues varient d’environ deux mètres pour les bioaérosols de quelques dizaines de micromètres, alors que les bioaérosols de quelques microns et submicrométriques ont la capacité de parcourir jusqu’à des centaines de kilomètres (Hinds, 1999). Plus la distance parcourue par un bioaérosol est grande, plus les risques d’inhalation, d’ingestion, de contact et donc de dissémination d’infections sont grands (Bovallius et al., 1978; Hurst et al., 2007; Polymenakou et al., 2008; Smith et al., 2012).
Toutefois le temps de passage dans l’air est un événement stressant pour les bioaérosols. En effet, qu’ils soient pathogènes ou non, infectieux ou pas, les microorganismes qui se retrouvent trappés dans les bioaérosols à la suite de leur aérosolisation sont susceptibles d’être endommagés par le taux d’oxygène, la température, l’humidité relative et les radiations (Marthi et al.,1990; Handley, & Webster, 1995; Mohr, 2007).
Il existe plusieurs évidences d’infections ou de problèmes de sensibilisation du système respiratoire, digestif et cutané causés par les bioaérosols. Le Tableau 1-1 dresse une liste non exhaustive d’exemples de microorganismes impliqués dans certaines infections ou maladies respiratoires pour lesquelles l’air est un vecteur (Ibach et al., 1954; Riley et al., 1959; Ehrenkra, & Kickligh, 1972; Nicholson, 1972; Dondero et al., 1980; Leclair et al., 1980, Bloch et al., 1985; Dick et al., 1987; Gurney, & Conces, 1996; Cole, & Cook, 1998; Hinds, 1999; Wang et al., 2005; Ismail et al., 2006; Vonberg, & Gastmeier, 2006; Brankston et al., 2007; Fernstrom, & Goldblatt, 2013).
5
Tableau 1. Microorganismes communs causant des infections ou des problèmes de sensibilisation.
Microorganisme (s) Infection/ Maladie
Influenza A Grippe
Rhinovirus Rhume
Virus de la rougeole Rougeole
Virus varicelle-zona Varicelle
Mycobacterium tuberculosis Tuberculose
SARS-coronavirus Syndrome respiratoire aigu sévère (SRAS)
Legionella pneumophila Légionellose
Bactéries et moisissures (substances antigéniques)
Alvéolite allergique extrinsèque
Aspergillus fumigatus/ flavus Aspergillose
Histoplasma capsulatum Histoplasmose
Pour pouvoir étudier les bioaérosols, il faut être en mesure de les récolter sous des conditions représentatives. Il est donc primordial de disposer d’échantillonneurs d’air efficaces pour récolter et concentrer le contenu microbien de l’air afin de poursuivre la caractérisation de certains environnements et la documentation de l’exposition humaine aux bioaérosols.
L’échantillonnage de l’air
Échantillonneurs d’air
L’échantillonnage de l’air est nécessaire pour récolter et concentrer le contenu microbien de l’air. Les différents types d’échantillonneurs d’air disponibles récoltent les bioaérosols en mettant à profit une ou plusieurs forces physiques pour les soustraire au flux d’air. Ces forces comprennent la sédimentation gravitationnelle, l’impaction inertielle, l’interception, la diffusion et l’attraction électrostatique.
6
La sédimentation gravitationnelle représente le processus de déposition naturel des bioaérosols d’une taille supérieure à 1 μm sur une surface (Hinds, 1999).
L’impaction inertielle s’applique à une particule ayant une certaine inertie et suivant une ligne du flux d’air. Étant incapable de s’ajuster rapidement aux changements brusques de direction de ces lignes, causés par la présence d’un obstacle, la particule s’y impacte. L’inertie dépend de la masse de la particule. Plus la masse est élevée et plus les forces appliquées sur la particule doivent être grandes pour la faire dévier de sa trajectoire afin d’éviter les collisions (Hinds, 1999).
L’interception se définie lorsqu’une particule suivant une ligne du flux d’air se trouve dans le rayon de surface d’un obstacle, le rencontre et s’y impacte. Lors de la collision, la particule est capturée en raison de sa taille finie. Certaines lignes du flux d’air entraînent la capture de particules alors que d’autres lignes du flux d’air ne le font pas. Il est supposé que les particules suivent parfaitement les lignes du flux d’air et donc que leur inertie, leur capacité de déposition et leur mouvement brownien sont négligeables. Ainsi, l’interception est le seul mécanisme n’étant pas le résultat d’une particule s’écartant de sa ligne du flux d’air d’origine (Hinds, 1999).
La diffusion se décrit lorsque le mouvement brownien des très petites particules (< 0,2 μm) est suffisant pour augmenter considérablement leur probabilité d’entrer en contact avec un obstacle, et ce, même si la ligne du flux d’air dans laquelle elles se trouvent ne rencontre pas un obstacle (Hinds, 1999).
La précipitation électrostatique s’applique à toutes particules chargées, attirées par une surface de charge contraire. Une particule chargée positivement sera attirée par une surface chargée négativement, et inversement. Des particules neutres peuvent aussi être attirées par des surfaces chargées (Hinds, 1999).
7
Évidemment, de la panoplie d’échantillonneurs d’air existant depuis les débuts de l’aérobiologie (impacteurs, impingers, cyclones et membranes filtrantes), quelques-uns sont encore utilisés avec leur design d’origine, alors que d’autres sont des versions plus récentes et performantes et d’autres encore sont le fruit de nouvelles technologies (condensation, micro fluidique et précipitation électrostatique) (Hering, & Stolzenburg, 2005; Liu et al., 2018; Imani et al., 2019). Chaque principe d’échantillonnage possède ses avantages et ses inconvénients et le choix d’un échantillonneur doit en tenir compte. De nombreuses études ont décrit divers échantillonneurs et ont permis leur comparaison (May, & Harper, 1957; Thorne et al., 1992; Grinshpun et al., 1997; Pahl et al., 1997; Willeke et
al., 1998; Lin et al., 1999; Lin et al., 2000; Duchaine et al., 2001; Agranovski et al., 2002;
Agranovski et al., 2005; Yao et al., 2009; Zhen et al., 2009; Coccia et al., 2010; Kesavan
et al., 2010a; Griffin et al., 2011; Springorum et al., 2011; Yamamoto et al., 2011; Han, &
Mainelis, 2012; Sanchez-Munoz et al., 2012; Su et al., 2012; Dybwad et al., 2014; Amato
et al., 2015; Kesavan, & Sagripanti, 2015; Wubulihairen et al., 2015; Haig et al., 2016; Yu et al., 2016; Chen, & Yao, 2018; Kim et al., 2018). Les conclusions formulées sont
généralement que l’efficacité de capture est variable d’un échantillonneur à l’autre, selon les objectifs de l’étude, les conditions testées et les microorganismes ciblés. Il est attendu que, dans un contexte particulier, un type d’échantillonneur d’air montrera des aptitudes supérieures à un autre et pourra ainsi être préféré selon les buts et les attentes d’une étude. Il est important de ne pas négliger ce dernier point et d’être critique avant de transposer des conclusions tirées d’une étude à une autre (Benbough et al., 1993; Dybwad et al., 2014; Haig et al., 2016; Sanchez-Munoz et al., 2012; Thorne et al., 1992). L’échantillonneur d’air idéal répondant à toutes les attentes, convenant à toutes les situations (environnements plus ou moins contaminés, présence de poussières, air turbulent, température ambiante, etc.) et capturant efficacement tous les différents types de bioaérosols, n’existe pas (Grinshpun et al., 2007; Haig et al., 2016). La création de nouveaux échantillonneurs d’air plus performants demeure justifiable et nécessaire, de même que l’utilisation de certains échantillonneurs plus classiques. Il est important d’être conscient des limites de chaque
8
échantillonneur pour éviter l’introduction de biais pendant l’expérimentation et lors des analyses subséquentes.
Échantillonneurs de type filtre
Les méthodes d’échantillonnage à l’aide de filtres reposent sur leur capacité à capturer des particules électriquement neutres et de différents diamètres. Une pompe est nécessaire pour faire passer activement l’air au travers du filtre. Il y a trois catégories principales de filtres : les filtres fibreux, les filtres à membrane et les filtres plats. Ils sont tous faits de matériaux différents qui influencent la grosseur des pores, variant de 0,01 µm à 10 µm (NIOSH, 2016b; Hinds, 1999). Les cassettes 37 mm (SKC Inc., Eighty-Four, PA, US) qui permettent d’utiliser diverses membranes et le SASS 3100 Dry Air Sampler (Research International, WA, É-U), mettant à profit une membrane électret, sont les deux échantillonneurs de type filtre d’intérêt pour ce projet.
Les cassettes 37 mm fermées sont faites d’un support en plastique hermétique contenant un filtre d’un matériel et d’une porosité variable. Le filtre est déposé sur une membrane de cellulose dans la partie inférieure de la cassette et elle est ensuite scellée avec la partie supérieure (Figure 3). L’air est aspiré par une pompe, généralement bas débit, et traverse le filtre. L’entrée d’air se trouve dans la partie supérieure de la cassette et fait face au filtre, alors que la sortie d’air se trouve dans la partie inférieure de la cassette, sous la membrane de cellulose (SKC Inc).
9
Figure 3 Cassette 37 mm. Figure tirée de Verreault et al., 2008 et adaptée par Caroline Duchaine dans le cours Bioaérosols et aérobiologie de l’Université Laval.
Le SASS 3100 possède un filtre en microfibres électrostatiques (ou filtre électret), où chaque fibre contient un champ électrique dans sa structure. Cela induit une charge aux particules passant au travers du filtre et augmente l’efficacité de capture. Le filtre électrostatique s’appose directement sur un ventilateur centrifuge servant à aspirer l’air (Figure 4). Le débit du SASS 3100 peut aller jusqu’à 300 L/min (Kesavan et al., 2010b; Research International).
10
Figure 4 Le SASS 3100 Dry Air Sampler et son filtre électrostatique. Figure fournie par Research International (https://www.resrchintl.com).
La sédimentation gravitationnelle, l’interception, l’impaction inertielle, le mouvement brownien et l’attraction électrostatique sont les forces impliquées dans la capture des bioaérosols par les filtres. Les filtres sont capables de capturer plusieurs tailles de particules selon la dimension de leurs pores, mais aussi qui sont plus petites que la grosseur de leurs pores en raison des différentes forces physiques mises à profit et par la structure des fibres. Leur efficacité de capture est élevée et peut atteindre environ 100% pour les particules d’une taille supérieure à la grosseur des pores (Grinshpun et al., 2007). La grosseur des pores, la présence d’un orifice critique ainsi que le pouvoir d’aspiration de la pompe déterminent le débit adéquat. Les filtres peuvent récupérer les particules de grands volumes d’air puisqu’ils ont peu de contraintes relatives à la durée de l’échantillonnage. Ils peuvent servir à échantillonner sur de longues périodes (heures). Toutefois, la capacité de rétention physique du filtre lui-même peut limiter l’échantillonnage, un filtre surchargé risquant de colmater. Aussi, le flux d’air constant passant à travers la membrane filtrante accentue la dessiccation des bioaérosols et risque d’affecter la viabilité et l’intégrité des microorganismes. Pour cette raison, les méthodes de culture sont généralement peu recommandées puisque la cultivabilité est altérée lors de l’échantillonnage sur membrane filtrante (Grinshpun et al., 2007). Les méthodes
11
moléculaires, telles que les réactions en chaîne par polymérase en temps réel (qPCR) cibant des microorganismes spécifiques, et le séquençage à haut débit sont alors favorisées (Stafford, & Ettinger, 1972; Cox, & Wathes, 1995; Hinds, 1999; Dybwad et al., 2014; Haig
et al., 2016; NIOSH, 2016a; NIOSH, 2016b, Soo et al., 2016).
Échantillonneurs de type liquide
Les échantillonneurs de type liquide tels que le BioSampler® (SKC Inc., Eighty-Four, PA, USA) et le Coriolisµ® (Bertin Technologies, Montigny-le-Bretonneux, France) sont couramment utilisés et représentent deux des échantillonneurs d’intérêt pour ce projet.
Le BioSampler® est un barboteur qui comporte un récipient de collection de 20 mL fait de verre contenant un liquide, une entrée et une sortie d’air (Figure 5). Une pompe branchée à la sortie d’air crée un vide qui permet à l’air d’être aspiré par l’orifice d’entrée. Celui-ci est un tube courbé se terminant en trois ajutages soniques (ou nozzles) à quelques millimètres au-dessus du liquide. Les ajutages, orientés en angle par rapport à la surface du récipient, agissent comme orifice critique et accélèrent les particules à la vitesse du son afin de permettre l’impaction des particules dans le liquide de collection. Le débit est donc fixé et limité à 12,5 L/min. L’orientation tangentielle des ajutages crée un mouvement circulaire (tourbillon) qui permet au liquide de monter contre les parois du récipient et de récolter les bioaérosols impactés (SKC Inc.).
12
Figure 5 BioSampler®. Les lignes rouges représentent le trajet du flux d’air entrant dans l’échantillonneur. Les lignes bleues représentent le trajet du flux d’air sortant de l’échantillonneur. Figure tirée de Verreault et al., 2008 et adaptée par Caroline Duchaine dans le cours Bioaérosols et aérobiologie de l’Université Laval.
Le fonctionnement du Coriolisµ® est basé sur une technologie cyclonique humide qui permet la récolte des particules selon le principe de centrifugation (Figure 6). Un vortex est créé lorsque l’air, aspiré par une pompe intégrée, est récupéré dans un récipient de collection conique de 15 mL contenant le liquide (Figure 7). Cela permet aux particules présentes dans l’air d’être déposées selon leur inertie sur les parois du récipient et d’être récoltées par le liquide en mouvement. Les particules ayant une grande inertie vont s’impacter rapidement contre la paroi du récipient de collection, alors que les particules ayant une moins grande inertie vont s’impacter plus loin sur la paroi. Les débits du Coriolisµ® peuvent varier de 100 L/min à 300 L/min (Bertin Technologies).
13
Figure 6 Trajet des particules selon leur inertie dans le récipient de collection du Coriolisµ®. Figure tirée de Willeke et al., 1998.
Figure 7 Schéma du récipient conique du Coriolisµ®. Les lignes rouges représentent le trajet du flux d’air entrant dans l’échantillonneur. Les lignes bleues représentent le trajet du flux d’air sortant de l’échantillonneur. Figure tirée et adaptée de Verreault et al., 2008.
L’impaction dans un liquide est un avantage en lui-même puisqu’il s’agit d’un processus relativement doux pour les bioaérosols. La vitesse d’impaction de même que le stress d’impaction sont amoindris, ce qui favorise la conservation de l’intégrité cellulaire
14
et donc de la viabilité des bactéries, des moisissures et de certains virus (Amato et al., 2015; Coccia et al., 2010; Cox, & Wathes, 1995; Grinshpun et al., 1997; Haig et al., 2016; Kesavan et al., 2010a; Lin et al., 1997; Wubulihairen et al, 2015). L’impaction dans un liquide permet la dilution et l’étalement sur des milieux de culture, l’analyse par microscopie et le décompte cellulaire (Juozaitis et al., 1994; Terzieva et al., 1996; Lin et
al., 1999; Agranovski et al., 2002; Hogan et al., 2005; Hermann et al., 2006; Rule et al.,
2007; Verreault et al., 2008; Coccia et al., 2010; Riemenschneider et al., 2010; Griffin et
al., 2011; King, & MacFarland, 2012; Ahmed et al., 2013; Dybwad et al., 2014; West, &
Kimber, 2015; Zheng, &Yao, 2017). Les analyses par méthodes moléculaires telles que les qPCR spécifiques et le séquençage à haut débit sont aussi possibles (An et al., 2006; Cayer
et al., 2007; Kim et al., 2011; Yamamoto et al., 2011; Lecours et al., 2012; Dybwad et al.,
2014; Hoisington et al., 2014; Bonifait et al., 2017; Mbareche et al., 2017a; Brisebois et
al., 2018; Mbareche et al., 2018; Veillette et al., 2018; Zhen et al., 2018).
Cependant, malgré les avantages procurés par les échantillonneurs mettant à profit un liquide de collection, il existe des désavantages notables. Le premier désavantage concerne l’évaporation d’une portion du liquide de collection pendant l’échantillonnage. Cette évaporation représente environ 10% du volume initial, mais peut parfois atteindre les trois quarts du volume (Henningson, & Ahlberg, 1994; Cox, & Wathes, 1995; Haig et al., 2016). Le taux d’humidité relative et la température ambiante sont des facteurs influençant le taux d’évaporation (Springorum et al., 2011). Le deuxième désavantage réside dans le mouvement continuel du liquide de collection qui favorise la ré-aérosolisation (re-suspension) de certains bioaérosols. Les bioaérosols ré-aérosolisés de l’échantillonneur retournent dans le flux d’air et sont perdus du récipient de collection (Agranovski et al., 2002; Griffin et al., 2011; Grinshpun et al., 1997; Han, & Mainelis, 2012; Lin et al., 1999; Riemenschneider et al., 2010; Springorum et al., 2011; Willeke et al., 1998). Malgré des études ayant révélé l’existence de l’évaporation et de la ré-aérosolisation, l’impact réel de ces deux phénomènes sur de possibles biais quant à la quantification et la biodiversité microbienne récoltée avec les échantillonneurs de type liquide demeure flou (Cox, & Wathes, 1995, Haig et al., 2016, Kesavan et al., 2010a, Lin et al., 1997). Pourtant, une
15
seule étude connue (Han, & Mainelis, 2012) a tenté de caractériser les deux phénomènes d’intérêts de ce projet, mais les réponses obtenues ne permettent pas de résoudre l’entièreté des questions adressées. De plus, des résultats préliminaires montrent que les échantillonneurs liquides sous-estiment la biodiversité microbienne récoltée comparativement à un échantillonneur filtre (Annexe A) (Veillette et al., 2017). Il est possible d’affirmer que des microorganismes se trouvent dans la portion du liquide évaporée. La question concernant les effets de cette perte par rapport à ce qui demeure dans le liquide de collection de l’échantillonneur demeure entière et est le cœur de ce travail de maîtrise. Trois scénarios sont possibles et peuvent influencer les biais causés par l’évaporation du liquide de collection : 1) de tous les bioaérosols récoltés dans le liquide de collection, une ou plusieurs populations sont complètement ré-aérosolisées, 2) de tous les bioaérosols récoltés dans le liquide de collection, une portion d’une ou plusieurs populations est ré-aérosolisée et 3) il y a seulement une portion du liquide de collection qui s’évapore donc les bioaérosols récoltés ne sont pas ré-aérosolisés. Dans le premier cas, la population n’aura été que de passage dans le récipient de collection de l’échantillonneur et ne sera pas identifiée. Dans le deuxième cas, la concentration réelle de la population sera sous-estimée puisqu’une portion seulement demeurera dans le récipient de collection. Dans le troisième cas, la population sera surestimée puisque l’unique évaporation du liquide de collection mènera à la concentration des bioaérosols dans le récipient. Dans le cas d’études d’exposition aux bioaérosols ou de présence/absence de certains pathogènes, les biais peuvent avoir des conséquences dommageables sur les résultats obtenus et sur l’évaluation du risque associé à l’exposition aux bioaérosols.
Facteurs influençant la ré-aérosolisation
Ce projet s’intéresse plus spécifiquement au comportement des bactéries pendant l’échantillonnage de l’air. Les prochaines sections vont cibler celles-ci et quelques genres en particulier (Lactobacillus spp., Moraxella spp., Pseudomonas spp. et Staphylococcus spp).
16
Transfert air-eau
Les océans représentent un bassin important d’aérosols et de bioaérosols (Kientzler
et al., 1954; Blanchard, & Woodcock, 1957; Fahlgren et al., 2015). Les études s’intéressant
à l’aérosolisation des microorganismes provenant des océans ont mis en évidence leur enrichissement à l’interface air-liquide, favorisant ainsi leur transfert dans l’air. Des bulles d’air montant à la surface d’un liquide, lorsqu’elles éclatent, vont relâcher une multitude de gouttelettes (Stuhlman, 1932). L’âge de la bulle influence aussi le nombre de gouttelettes qui seront produites. Une bulle plus vieille produira beaucoup plus de gouttelettes qu’une jeune et la taille de ces gouttelettes sera beaucoup plus petite (Poulain & Bourouiba, 2018). Celles-ci sont environ dix fois plus petites que la bulle de laquelle elles originent. Les bactéries ont tendance à se concentrer dans les couches de surface de l’eau si bien que lorsque les bulles éclatent en plusieurs gouttelettes, elles éjectent les bactéries dans l’air (Figure 8). La concentration des bactéries dans les gouttelettes peut être plus élevée que la source d’où elles proviennent, par exemple d’un facteur 100 fois plus concentré dans les gouttelettes que dans la bulle. Certaines des bulles montant à la surface vont accrocher des bactéries qui vont s’ajouter à celles déjà présentes à l’interface air-eau. Du matériel biologique présent dans le liquide s’adsorbe à la surface des bulles et est nécessaire à l’attachement des bactéries. Il semblerait être un des facteurs déterminant dans la réussite d’une bulle à collecter des bactéries, puisqu’une eau naturelle contient plus de matériel actif qu’une eau distillée. Une bulle provenant d’une eau naturelle aurait aussi une durée de vie moins longue (Poulain, & Bourouiba, 2018). Plus une bulle est grosse, plus elle produit un grand nombre de gouttelettes. Il y a aussi une corrélation positive entre le degré d’enrichissement des bactéries à la surface et leur hydrophobicité. Les charges portées à la surface membranaire des bactéries influencent aussi leurs interactions à l’interface air-eau (Blanchard, & Syzdek, 1970; Blanchard, & Syzdek, 1972; Blanchard, 1978; Blanchard, & Syzdek, 1978a; Dahlback et al., 1981). Ainsi, si une bulle qui éclate et qui produit des gouttelettes à la surface de l’océan relâche des bactéries dans l’air, les bulles produites par le mouvement du liquide de collection d’un échantillonneur d’air sont aussi susceptibles de reproduire le même phénomène.
17
Figure 8. Bulle éclatant en plusieurs gouttelettes à la surface de l’eau. Figure tirée de Lhuissier, & Villermaux, 2012.
Caractéristiques de la paroi bactérienne
Les bactéries possèdent différentes caractéristiques selon la composition de leur membrane cellulaire, ce qui influence leur interaction entre elles et avec leur environnement. Les bactéries sont classifiées en deux groupes selon les composantes membranaires de leur paroi cellulaire. Les bactéries à Gram positif possèdent une épaisse couche de peptidoglycane où des acides teichoïques et lipoteichoïques sont insérés. L’espace périplasmique et la membrane cytoplasmique sont situés sous la couche de
18
peptidoglycane (Figure 9). Les bactéries à Gram négatif possèdent une mince couche de peptidoglycane située sous une membrane externe, absente chez les bactéries à Gram positif. Dans cette membrane externe se trouve des lipopolysaccharides (LPS) et d’autres protéines (Figure 10). Parmi les bactéries à Gram positif se trouvent les genres
Staphylococcus spp. et Lactobacillus spp. et parmi les bactéries à Gram négatif, Moraxella
spp. et Pseudomonas spp. C’est entre autres la nature de ces différentes composantes extracellulaires qui détermine les propriétés externes des bactéries telles que la charge cellulaire, l’hydrophobicité et la capacité d’agrégation.
Figure 9 Schéma de la paroi cellulaire des bactéries à Gram positif. Figure tirée de http://www.ebiologie.fr/cours-La_paroi_bacterienne.html et adaptée par Caroline Duchaine, dans le cours Bioaérosols et aérobiologie de l’Université Laval.
19
Figure 10 Schéma de la paroi cellulaire des bactéries à Gram négatif. Figure tirée de http://www.ebiologie.fr/cours-La_paroi_bacterienne.html et adaptée par Caroline Duchaine, dans le cours Bioaérosols et aérobiologie de l’Université Laval.
Les bactéries possèdent généralement une charge nette négative en raison de la présence d’ultrastructures à leur surface cellulaire. Ces structures contiennent différents groupements chargés (négativement ou positivement) qui contribuent à la polarité en périphérie cellulaire. Les charges permettent entre autres aux bactéries d’interagir entre elles ou avec des substrats comme un liquide ou une surface. Les interactions entre bactéries peuvent mener à la formation d’agrégats. L’agrégation varie selon le pH puisqu’elle est liée aux charges portées et définie l’affinité entre les bactéries selon la force ionique du milieu (Hamadi et al., 2005). Les charges portées par les bactéries contribuent aussi à l’hydrophobicité de leur membrane cellulaire. (Dickson, & Koohmaraie, 1989; Mozes, & Rouxhet, 1990; Saito et al., 1997; Wilson et al., 2001).
L’hydrophobicité est la tendance des molécules apolaires de s’associer avec d’autres molécules apolaires plutôt qu’avec des molécules polaires comme l’eau. En effet, une molécule d’eau est réfractaire au fait de sacrifier un de ses liens hydrogène au détriment d’une molécule apolaire. Cela cause inévitablement une réorientation des molécules d’eau autour de la molécule apolaire, l’excluant ainsi de leur réseau (Doyle, 2000). Certaines
20
bactéries sont considérées hydrophobes, d’autres hydrophiles. Le degré d’hydrophobicité de la surface cellulaire bactérienne est affecté par la présence de protéines de surface ou d’autres composés extra membranaires, spécifiques à certains genres et espèces. Par exemple, Staphylococcus aureus est généralement hydrophobe en raison de deux protéines de surface, la protéine A et les protéines de surface se liant à la fibronectine. Les acides téichoïques et lipotéichoïques contribuent aussi à son hydrophobicité (Jonsson, & Wadstrom, 1983; Ljungh et al., 1985; Mitik-Dineva et al., 2009). Pseudomonas
aeruginosa peut être hydrophile en raison d’une composante des LPS à sa surface
cellulaire, mais peut devenir plus hydrophobe s’il y a sécrétion de surfactant neutralisant l’effet des LPS (Al-tahhan et al., 2000; Makin, & Beveridge, 1996; Mitik-Dineva et al., 2009). De même, Lactobacillus spp. peut être hydrophile ou hydrophobe dépendamment de la présence de polysaccharides ou de glycoprotéines dans certaines structures à la surface cellulaire (Reid et al., 1992; Cuperus et al., 1993; Andreu et al., 1995; Pelletier et
al., 1997). Moraxella spp. tend à être hydrophobe en raison d’un type particulier de LPS à
sa surface cellulaire et la présence de phospholipides (Gotoh et al., 1989; Tsujimoto et al., 1999).
Toutefois, toutes ces caractéristiques sont variables selon l’environnement (pH et température), la niche écologique, le stade et les conditions de croissance des bactéries et les différentes espèces d’un même genre (Gilbert et al., 1991; Ljungh, & Wadstrom, 1995; Hamadi et al., 2005; Foster et al.,2014; Di Ciccio et al., 2015).
Liquide de collection
Le principal désavantage de l’échantillonnage de l’air dans un liquide est l’évaporation d’une portion du liquide. Certaines alternatives sont possibles pour tenter de réduire le taux d’évaporation en modifiant la tension de surface du liquide grâce à l’ajout d’un composé visqueux (glycérol, huile minérale, agarose, etc.) (Lin et al., 2000). L’ajout de ces substances modifie la composition du liquide de collection et est donc susceptible d’influencer le comportement des bactéries. En effet, comme décrit précédemment, une
21
eau naturelle ne va pas enrichir les bactéries dans une bulle de la même façon qu’une eau distillée. La présence d’éléments biologiques et chimiques semble favoriser l’interaction des bactéries avec ce liquide. Les liquides de collection utilisés dans certains échantillonneurs sont majoritairement des solutions tamponnées à base d’eau (Grinshpun
et al., 2007). Par exemple, une solution saline tamponnée au phosphate (PBS) contient des
sels, donc des ions chargés positivement ou négativement, susceptibles d’interagir avec les charges à la surface des bactéries et d’influencer leur propre interaction dans le liquide. L’ajout de glycérol au tampon modifie la dynamique chimique du liquide puisqu’il contient des groupements hydroxyles (chargés négativement), favorisant la formation de liens hydrogène (Sigma Aldrich, Oakville, Ontario, Canada). Les interactions bactériennes sont encore susceptibles d’être modifiées et par le fait même d’influencer la ré-aérosolisation ou la concentration des bactéries.
Mécanisme de capture de l’échantillonneur et débit
Le BioSampler® et le Coriolisµ® mettent à profit deux mécanismes de capture des bioaérosols différents. Le mouvement du liquide pendant l’échantillonnage de l’air n’est pas le même dans un barboteur (BioSampler®) que dans un cyclone (Coriolisµ®). Le tourbillon produit par l’aspiration d’air dans le BioSampler® est susceptible de produire beaucoup de bulles et de favoriser ainsi la ré-aérosolisation. L’effet de la force centrifuge utilisée dans le Coriolisµ® peut favoriser les particules ayant une plus grande inertie à demeurer dans le liquide de collection, alors que celles ayant moins d’inertie ont plus de probabilités d’être ré-aérosolisées (Haig et al., 2016). Aussi, sachant que le débit influence le taux d’évaporation et que celle-ci est susceptible d’être un moteur à la production de bulles et d’influencer la ré-aérosolisation, plus le débit est élevé, plus il y a d’évaporation et plus il y a de possibilités de ré-aérosolisation.
22
Facteurs influençant l’échantillonnage
Aérosolisation préférentielle
Un microorganisme enrichi dans l’air comparativement à la source de laquelle il provient se définit comme préférentiellement aérosolisé. Ce phénomène est intrinsèquement lié à l’enrichissement des bactéries à l’interface air-eau des océans. Toutefois, une des premières évidences de l’enrichissement préférentiel de certaines bactéries dans l’air concerne une bactérie potentiellement pathogène, Mycobacterium
intracellulare (Parker et al., 1983). Malgré l’importance relative de ce phénomène sur la
dispersion possible de microorganismes pathogènes, peu d’études s’y sont intéressées avant tout récemment. En effet, Groulx et al. ont mis en évidence l’aérosolisation préférentielle de Pseudomonas fluorescens comparativement à E. coli à partir d’une solution mixte de ces deux genres (Groulx et al., 2015). Perrott et al. ont démontré l’aérosolisation préférentielle de Pseudomonas aeruginosa comparativement à
Staphylococcus aureus, Streptococcus suis (Sérotype 2), Moraxella catarrhalis et Lactobacillus paracasei, à partir d’une solution combinant les cinq genres (Perrott et al.,
2017). Gauthier-Levesque et al. ont montré que l’aérosolisation préférentielle de la souche
S. suis sérotype 2 ST1, à partir d’une source liquide, est un processus « souche dépendante
» (Gauthier-Levesque et al., 2016). Fahlgren et al. ont mis en évidence l’enrichissement de certaines bactéries marines dans les aérosols produits comparativement à leur source liquide (Fahlgren et al., 2015). Ces études ont évalué le phénomène in vitro alors que Moletta et al., Moletta-Denat et al., ainsi que Wang et al. ont mis en évidence l’aérosolisation préférentielle de certaines bactéries dans les environnements de bio-méthanisation et de fluides de coupe de métaux (Moletta et al., 2007, Wang et al., 2004; Moletta-Denat et al., 2010).
Diamètre particulaire (D
50)
Le D50 représente le diamètre particulaire précis où 50% des bioaérosols ayant cette taille seront captés efficacement par l’échantillonneur et où 50% des bioaérosols ayant une taille inférieure ne seront pas captés. C’est la limite de taille minimale des particules
23
pouvant être échantillonnées (Hinds, 1999). La valeur du D50 varie d’un échantillonneur d’air à un autre, il est donc susceptible d’influencer le contenu microbien récolté par chacun d’eux. Par exemple, le D50 du BioSampler®est de 0,3 µm (Kesavan et al., 2010a ; SKC Inc), 0,5 µm pour le Coriolis® (Bertin Technologies) et le SASS 3100 (Research International). Le D50 est toutefois inconnu pour la cassette 37 mm (SKC Inc).
Méthodes de traitement et d’analyse des échantillons
Méthodes de traitement
Les méthodes de traitement et d’analyse des échantillons peuvent aussi introduire des biais. En effet, à la suite de l’échantillonnage, des étapes de traitement des échantillons sont nécessaires et varient selon le type d’échantillonneur utilisé. Comme pour les protocoles d’échantillonnage, il n’existe pas non plus de protocoles standardisés de traitement des échantillons. Des étapes de centrifugation, de filtration ou d’élution sont appliquées pour récupérer les bioaérosols d’un échantillon liquide ou d’un filtre avant l’extraction de l’ADN (Jacobsen, & Rasmussen, 1992; Lucore et al., 2000; Cullison, & Jaykus, 2002; Cayer et al., 2007; Nehme et al., 2008; Nehme et al., 2009; Gendron et al., 2010; Lecours et al., 2012; Mbareche et al., 2017a; Triado-Margarit et al., 2017; Turgeon
et al., 2017; Zhen et al., 2018).
La centrifugation sert principalement à concentrer dans un culot les bioaérosols provenant d’un échantillon liquide ou d’un éluat. Les échantillons d’air récupérés par un échantillonneur de type filtre doivent être élués dans un liquide et peuvent faire l’objet d’une étape de filtration supplémentaire ou de centrifugation. Une récente étude a montré que la centrifugation n’arrive pas à concentrer efficacement certaines moisissures dans le culot en raison de propriétés hydrophobes (Mbareche et al., 2019). Elles demeurent plutôt dans le surnageant et sont perdues en même temps que ce dernier est retiré. Les résultats obtenus sont donc biaisés puisqu’il y a une sous-estimation de la quantité réelle de ces moisissures dans l’air. Toutefois, une méthode alternative a été proposée où les
24
échantillons liquides sont filtrés sur une membrane de polycarbonate (0,22 µm) à l’aide d’une unité de filtration. Cette méthode donne un rendement significativement supérieur à la centrifugation. Si certaines moisissures sont perdues lors de la centrifugation, il n’est pas exclu que certaines bactéries ayant des caractéristiques hydrophobes le soient aussi.
L’élution des filtres n’est pas standardisée et dépend de l’échantillonneur utilisé ou du support choisi. Par exemple, il est recommandé d’éluer les filtres du SASS 3100 avec l’extracteur de particules (SASS 3010 Particle Extractor, Research International) et la solution d’extraction (Extraction Solution, Research International), prévus à cette fin. Le fabricant garantit une efficacité de première extraction de 70% à 80%, mais trois extractions supplémentaires sont nécessaires pour obtenir une efficacité de près de 100% (Research International). Ces tests d’efficacité d’extraction ont été exécutés par le fabricant avec des billes de polystyrènes et non avec des particules biologiques. Il est donc fort probable que les bioaérosols récoltés sur les filtres électrostatiques ne soient pas extraits en totalité. Par contre, pour ce qui est des cassettes, les méthodes d’élution varient. Les bioaérosols peuvent être extraits en injectant une solution directement sur le filtre, dans la cassette encore scellée, et agiter sur plaque agitatrice pendant un temps donné (Blomquist
et al., 1984a; Blomquist et al., 1984b; Palmgren et al., 1986; Nielsen et al., 1995; Heldal et al., 1996; Wang et al., 2001). Une autre méthode consiste à retirer le filtre de la cassette
et le déposer dans un tube stérile contenant un certain volume de solution d’extraction. Les particules sont extraites du filtre en plaçant le tube sous agitation, sur un vortex ou encore dans un bain à ultrasons. Il est toutefois impossible d’extraire en totalité tous les bioaérosols captés, peu importe la méthode utilisée. La composition (tampon phosphate, eau peptonée) et le volume des solutions d’extraction (20 à 50 mL) varient, en plus du temps de contact entre le filtre et la solution d’extraction (10 à 30 minutes). Finalement, le type de filtre utilisé varie et ne donne pas le même rendement d’extraction des particules, peu importe la méthode (Wang, 1999; Burton et al., 2005; Wang et al., 2015). Bien que les échantillonneurs de type filtre aient une efficacité de capture des bioaérosols élevée, les méthodes d’extraction sont susceptibles de biaiser les résultats.
25
Les méthodes d’extraction d’ADN sont aussi variées et n’ont pas toutes les mêmes efficacités d’extraction dépendamment des microorganismes ciblés. Des biais peuvent donc s’introduire à cette étape du traitement des échantillons d’air puisqu’aujourd’hui, une multitude de trousses commerciales sont disponibles selon les mediums (sol, tissus, sang, fèces, eau, air, etc.) et les analyses subséquentes ciblées. La grande majorité des trousses procèdent à l’extraction d’ADN sur colonnes, selon une séquence d’étapes préétablies, incluant une lyse cellulaire mécanique (Tan, & Yiap, 2009). Par exemple, l’efficacité d’extraction d’ADN des bactéries du sol est variable selon la trousse utilisée. Il y a un enrichissement de certains genres tout au long du processus, ce qui mène inévitablement à un biais au niveau de la composition de l’échantillon résiduel. La paroi des bactéries à Gram positif est plus difficile à briser que celle des bactéries à Gram négatif (Pitcher et al., 1989; Martin-Laurent et al., 2001). Le même scénario est observé pour les moisissures aéroportées moins sensibles que d’autres à certaines étapes de l’extraction d’ADN. La robustesse des spores et des conidies peut rendre plus difficile la lyse mécanique et leur caractère hydrophobe rendre certains tampons moins efficaces. La sous-estimation de certaines espèces lors d’analyses par réaction en chaîne par polymérase en temps réel (qPCR) ou séquençage est observée (Haugland et al., 1999; Fredricks et al., 2005; Rittenour et al., 2012).
Méthodes d’analyses
La caractérisation et la description des bioaérosols présents dans certains environnements d’intérêt sont possibles par des analyses de quantification et de biodiversité microbienne. La quantification peut se faire par culture sur milieu sélectif ou à l’aide de méthodes moléculaires. La culture peut grandement sous-estimer le nombre total de microorganismes présents en raison de la notion de viable, mais non cultivable. En effet, il y aurait moins d’un pour cent des espèces bactériennes présentes dans l’environnement pouvant être cultivées (Bloomfiled et al., 1998; Oliver, 2005; Kim et al., 2011). Il est vrai que les différents stress subis par les bioaérosols lors de l’échantillonnage, et parfois même les conditions de croissance plus restrictives de certains microorganismes, biaisent les méthodes d’analyses par culture. Le nombre et la diversité des
26
microorganismes présents sont grandement sous-estimés. La culture sert, en règle générale, à évaluer la portion des bactéries et des moisissures viables et représentant un risque de santé lors d’études d’évaluation de l’exposition, par exemple. (Amann et al., 1995; Heidelberg et al., 1997; Stewart et al., 1995; Rinttila et al., 2004; Kim et al., 2011). Pour pallier aux limitations de la culture et tenter de détecter et quantifier l’ensemble des communautés microbiennes présentes, des méthodes moléculaires telles que la qPCR et le séquençage à haut débit ont été mises au point.
La PCR quantitative (qPCR) permet de quantifier l’ensemble des génomes bactériens en ciblant la région V6-V8 du gène de l’ADNr 16S, commune et conservée parmi toutes les familles bactériennes (Woese, 1987; Bach et al., 2002). Selon la spécificité des amorces utilisées, cette méthode permet de quantifier l’ensemble des bactéries présentes ou certains groupes, espèces ou souches spécifiques. D’autres régions du génome, spécifiques à un genre ou à une espèce, peuvent également être ciblées et permettre l’identification et la quantification d’une bactérie en particulier (Greiner et al., 2003; Spilker et al., 2004; Haarman, & Knol, 2006). La région ciblée est amplifiée à l’aide d’amorces spécifiques pendant plusieurs cycles consécutifs, en présence d’un fluorophore spécifique à la double hélice d’ADN, ce qui permet de déduire la quantité d’ADN produite à chaque cycle (Bach
et al., 2002). La qPCR peut détecter jusqu’à 100 fois plus de bactéries que la culture (Cruz,
& Buttner, 2007; Rinttila et al., 2004; Nehme et al., 2008; Kim et al., 2011), mais elle possède certaines limites. Par exemple, le nombre de copies du gène de l’ADNr 16S varie d’un genre, d’une espèce et d’une souche à l’autre. En effet, certaines bactéries n’ont qu’une copie du gène alors que d’autres en ont plus (Acinas et al., 2004; Rastogi et al., 2009). La quantité d’ADN produite est relative et donne une approximation de l’ordre de grandeur de la charge bactérienne présente. L’abondance des différentes bactéries variant d’un environnement à un autre, il est alors possible que la concentration d’un microorganisme soit sous la limite de détection. Dans ce cas-ci, le microorganisme ne sera pas détecté (Gilbert et al., 2010). La présence d’inhibiteurs présents dans les échantillons naturels peut aussi réduire la sensibilité et le seuil de détection de la réaction et mener à de faux négatifs (Bach et al., 2002; Cruz, & Buttner, 2007).
27
Pour sa part, le séquençage à haut débit permet une analyse qualitative par la présence/absence de bactéries plutôt que quantitative comme la qPCR. Le séquençage cible le gène de l’ARNr 16S bactérien et permet l’amplification de millions de copies des gènes (séquences) présents dans un échantillon (An et al., 2006; Mardis, 2008; Reis-Filho, 2009; Behjati, & Tarpey, 2013). La quantité et la complexité des données sont telles que la bio-informatique est nécessaire pour trier et convertir les séquences en identités bactériennes. La volonté d’éviter l’introduction de biais par des chimères (mauvaises amplifications et recombinaisons de séquences) et par un protocole bio-informatique inapproprié peut tout de même mener à d’autres biais. En effet, le protocole bio-informatique subséquent peut nuire à la représentation des bactéries moins abondantes et rares dans l’échantillon. Les différentes étapes de nettoyage et de tri des séquences mènent inévitablement à un retranchement des bactéries peu abondantes et rares puisque souvent leurs séquences sont de petites tailles et présentent dans un nombre insuffisant. Ces bactéries peuvent demeurer dans l’ombre de celles plus abondantes (Morgan et al., 2010; Peccia et al., 2011; Xiong et
al., 2010; Logares et al., 2014; Mbareche et al., 2017a).
La qPCR et le séquençage ne permettent pas de distinguer les microorganismes viables des non viables ni d’évaluer le potentiel infectieux. La culture permet parfois la détection de bactéries rares qui ne le seraient pas par méthodes moléculaires ou qui voient leur abondance sous-estimée en raison de leur absence des bases de données (Shade et
al.,2012). Les méthodes moléculaires peuvent être utilisées en complément à la culture
28
Problématique, hypothèse et objectifs des travaux de
recherche
Problématique
Selon une récente étude du laboratoire Duchaine, la divergence entre la diversité microbienne récoltée par un échantillonneur de type filtre et celle d’un type liquide est évidente, mais difficilement explicable. L’évaporation d’une portion du liquide de collection est observée lors de l’échantillonnage de l’air à l’aide d’échantillonneurs liquides. Cette évaporation pourrait mener à la ré-aérosolisation ou à la concentration de microorganismes captés et représente un phénomène susceptible de favoriser et d’engendrer un biais dans l’analyse des bioaérosols. En effet, certaines bactéries pourraient être entièrement ou partiellement ré-aérosolisées du liquide de collection ou être concentrées. Cela représente un biais non négligeable puisque que certaines bactéries peuvent ne pas être identifiées, alors que la concentration de certaines peut être grandement sous-estimée ou surestimée, dans un environnement ou un contexte précis. Les conséquences de ces biais sur l’issue de conclusions émises par différentes équipes de recherche peuvent être importantes. Pourtant, jusqu’à ce jour, peu d’intérêt de la part de la communauté scientifique n’a été démontré concernant la ré-aérosolisation et l’évaporation.
Hypothèse
L’évaporation observée lors de l’utilisation des échantillonneurs de type liquide est liée à la ré-aérosolisation ou à la concentration des bactéries présentes dans le liquide et elle induit un biais quant à la quantification des bactéries totales présentes dans l’air et leur diversité.
Objectifs
Ce projet de maîtrise a pour but de documenter l’évaporation et les phénomènes de concentration et de ré-aérosolisation conséquents afin de mieux comprendre les biais qu’ils
29
sont susceptibles d’engendrer lors de l’utilisation d’échantillonneurs de type liquide. L’étude comporte deux volets, un volet in vitro où la prédictibilité et la constance des comportements de ré-aérosolisation et de concentration seront évaluées et un volet en condition naturelle permettant de documenter les biais engendrés par l’évaporation.
1. Évaluer les phénomènes de ré-aérosolisation et de concentration, en inoculant le récipient de collection du BioSampler® et du Coriolisµ® avec un consortium bactérien connu et suivre son évolution pendant le fonctionnement des échantillonneurs.
2. Évaluer le biais induit par l’évaporation du liquide des échantillonneurs en les comparant au type filtre et en captant les particules perdues lors de l’évaporation, dans un environnement naturel.
30
Chapitre 1 : Re-aerosolization in
liquid-based air samplers induces bias in
bacterial diversity
Article accepté le 23 juillet 2019 dans le journal Aerosol Science and Technology. (numéro du manuscrit AST-MS-2019-073.R1)
Joanie Lemieux12, Marc Veillette2, Hamza Mbareche12 and Caroline Duchaine1 2
1. Centre de recherche de l’Institut universitaire de cardiologie et pneumologie de Québec, Quebec City (Qc), Canada.
2. Département de biochimie, de microbiologie et de bio-informatique, Faculté des sciences et de génie, Université Laval, Quebec City (Qc), Canada
Keywords: Bioaerosols, liquid-based air samplers, diversity, high throughput sequencing, re-aerosolization.