Behavioral and Immunological Features Promoting the Invasive Performance of the Harlequin Ladybird Harmonia axyridis
Texte intégral
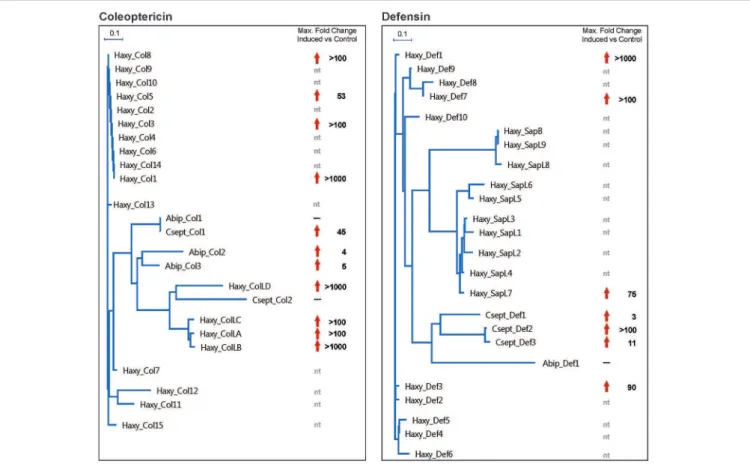
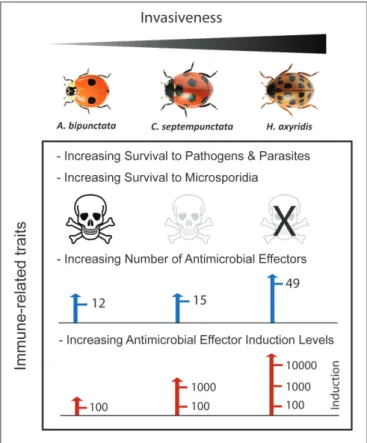
Figure
Documents relatifs
Specific numbers of fertilized and unfertilized eggs, and larvae from these lab samples (n = 5, 10, 20 and 50; with two replicates of each), were used for the purpose of
« Une délégation du service publique et un contrat par lequel une personne de droit publique confie la gestion d’un service publique dont elle a la responsabilité à un
laperrinei populations from South Algeria and Niger is conducted with two main objectives: (1) to analyse the genetic structure of populations distributed in four isolated
( فلاث نم ظئاٚضلا ٜكٚم ڗڤكتي 03 :ڗيتحلصمڣ غضابقڣ غيسيئٙ حلاصم ) 1 - يئٚلا غحلصلما :رييستلل غيس :ب لفكتتڣ - غيئابجلا غبقاٚلماڣ ، اعڤلا
In contrast, aside from slightly negative net diversification during the Late Cretaceous, there is little evidence from the fossil record for a significant K-Pg mass extinction
Using a molecular phylogenetic framework for the New World Troidini of the genera Battus, Euryades and espe- cially Parides, this study aims to: (i) use a DNA-based delimitation
Global variation in climbing palm species richness can be partly explained by past and present-day climate and rain forest canopy height, but regional differences in residual
رثأتت تامولعلما هدوج ةيبساحلما ريراقتلا يف ةيلالما .يف اهصيخلت نكمي لماوع هدعب 1 - ةئيب( ةيئيبلا لماوعلا ةبساحلما :) نإ فورظلل ةيعامتجلاا ةيداصتقلااو
