m UNIVERSITÉ DE
U SHERBROOKE
Faculté de génieDépartement de génie civil
FORMATION D'UNE STRUCTURE POLYMERIQUE POUR LA
STABILISATION DE LA LACCASE ET D'AGREGATS DE LACCASE
RETICULES
Mémoire de maîtrise Spécialité : Génie civil
Thanina HASSANI Jury : Hubert Cabana (directeur)
Jay Lacey (Rapporteur) Alexandre Cabrai
Library and Archives Canada Published Heritage Branch Bibliothèque et Archives Canada Direction du Patrimoine de l'édition 395 Wellington Street Ottawa ON K1A0N4 Canada 395, rue Wellington Ottawa ON K1A 0N4 Canada
Your file Votre référence ISBN: 978-0-494-90969-0 Our file Notre référence ISBN: 978-0-494-90969-0
NOTICE:
The author has granted a
non-exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
telecommunication or on the Internet, loan, distrbute and sell theses
worldwide, for commercial or non-commercial purposes, in microform, paper, electronic and/or any other formats.
AVIS:
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque et Archives Canada de reproduire, publier, archiver, sauvegarder, conserver, transmettre au public par télécommunication ou par l'Internet, prêter, distribuer et vendre des thèses partout dans le monde, à des fins commerciales ou autres, sur support microforme, papier, électronique et/ou autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur conserve la propriété du droit d'auteur et des droits moraux qui protege cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement
reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
While these forms may be included in the document page count, their removal does not represent any loss of content from the thesis.
Canada
Conformément à la loi canadienne sur la protection de la vie privée, quelques
formulaires secondaires ont été enlevés de cette thèse.
Bien que ces formulaires aient inclus dans la pagination, il n'y aura aucun contenu manquant.
Résumé
L'utilisation d'enzymes dans différentes applications biotechnologiques et industrielles s'est avérée prometteuse durant les dernières décennies. La laccase est une enzyme qui suscite l'attention, notamment, grâce à sa capacité à dégrader une vaste gamme de polluants présents dans l'environnement, à son apparente stabilité, et au fait qu'elle peut d'être utilisée dans une variété d'applications comme : les cosmétiques, l'industrie du papier, ou encore le traitement des eaux et la bioremédiation de sols. Il apparait donc pertinent d'améliorer le potentiel d'utilisation de cette enzyme en augmentant sa stabilité et sa capacité à être réutilisée.
À ces fins, plusieurs méthodes l'immobilisation/insolubilisation sous forme de cross-linked enzyme agregates (CLEAs) ou encore la formation d'une structure polymérique autours des biocatalyseurs se sont avérés efficaces. Cette étude vise à stabiliser la laccase libre et sous forme de CLEAs via la formation de réseau polymérique organique/inorganique formé de chitosane et de silane. Cette technique a permis de 1) stabiliser significativement l'activité enzymatique face aux dénaturants chimiques et thermique; 2) augmenter la résistance thermique de la laccase et des CLEAs modifiées; 3) augmenter l'efficacité catalytique de la laccase et CLEAs modifiées ainsi que leur affinité pour le substrat utilisé. De plus, les biocatalyseurs formés s'avèrent insolubles ce qui permettrait leur utilisation/réutilisation dans des bioréacteurs opérés en continu.
Mots clés : Laccase, CLEAs, Chitosane, Structure polymérique organique/inorganique.
Remerciements
Je tiens à sincèrement remercier monsieur Hubert Cabana, sans qui, ce projet n'aurait pu être entrepris et mené à bien. Je remercie également Djazia Liamini et Serge Bérubé pour leur collaboration et support technique qu'ils m'ont apportés. Un grand merci à toutes les personnes qui ont rendu agréable la réalisation de ce projet : Alexandre Arsenault, Jean-François Leduc, Sidy Ba, Halim, et Amina.Finalement, je tiens à exprimer ma reconnaissance au Conseil de Recherches en Sciences Naturelles et en Génie du Canada pour son aide financière.
Table des matières
Résumé i
Remerciements i
Table des matières ii
Liste des figures iv
Liste des tableaux vi
Liste des abréviations vii
Chapitre 1
1.1 Perturbateurs endocriniens 1
1.1.1. Nonylphénol 2
1.1.2. Triclosan 3
1.1.3. Bisphénol A 5
1.2. Similitudes structurales entre les PE 6
1.3. Techniques d'élimination des composés phénoliques 8
1.2. Objectifs 12 1.2.1. Objectif général 12 1.2.2. Objectifs spécifiques 12 1.3. Contributions originales : 12 1.3.1. Articles 12 1.3.2. Présentations orales 13
1.3.3. Présentations par affiche 13
1.4 Structure du manuscrit 14
Chapitre 2
2.1. La laccase 15
2.1.1. Origine : champignons lignivores 15
2.1.2. Propriétés de la laccase 18
2.2. Stabilisation enzymatique 22
2.2.1 Immobilisation enzymatique 23
Immobilisation par liaisons covalentes 23
Immobilisation par adsorption physique 25
Insolubilisation par réticulation d'enzymes (sans support) 26 2.2.2. Stabilisation des enzymes par la formation de structures polymériques 29
Chapitre 3
Avant-propos 33
Formation of enzyme polymer engineered structure for laccase and cross-linked
laccase aggregates stabilization 34
Résumé 34
Abstract 34
3.1. Introduction 35
3.2. Materials and methods 37
3.2.1. Chemicals 37
3.2.2. Preparation of chitosan solution 37
3.2.3. Preparation of the biocatalysts 37
3.2.4. Enzymatic activity assay 39
3.2.5. Stability of the biocatalysts 39
3.2.6. Biocatalysts kinetics 39
3.2.7. Optimal pH and temperature 40
3.2.8. Scanning electron microscopy of the biocatalysts 40
3.3. Results and discussion 40
3.3.1. EPES formation 40
3.3.2. Thermal stability of free laccase, CLEAs, EPES-lac and EPES-CLEAs 43
3.3.3. Stability against chemical dénaturants 45
3.3.4. Michaelis-Menten kinetic parameters 47
3.3.5. Optimum pH 48
3.3.6. Optimal temperature 49
3.3.7. SEMs of EPES-lac and EPES-CLEA 51
3.4. Conclusion 51 3.5. Acknowledgments 52 Chapitre 4 Conclusion 53 Annexe 1 56 Annexe 2 57 Annexe 3 60 Références 62
Liste des figures
Figure 1-1 : Structure du nonylphénol 2
Figure 1-2: Structure du triclosan 3
Figure 1-3 : Structure du bisphénol A 5
Figure 2-1 : Pleurotus ostreatus 15
Figure 2-2 : Trametes versicolor 15
Figure 2-3 : Pourriture blanche sur de bois provoquée par le WRF 15 Figure 2-4 : Le cycle catalytique de la LiP montrant les trois états de l'enzyme représentés par le centre heme. AH représente le substrat de l'enzyme, et A°, le substrat oxydé par l'enzyme (Gold et Alic, 1993) 17 Figure 2-5: Le cycle catalytique de la MnP montrant les trois états de l'enzyme représentés par le centre hème, et montrant le couplage avec le complexe Mn(III)-acide organique. Les demi-cercles représentent des acides organiques, AH le substrat et A° le substrat oxydé par l'enzyme (Lôpez et
al., 2002) 18
Figure 2-6 : Cycle catalytique de la laccase (d'après (Wesenberg et al, 2003)) 20 Figure 2-7 : Représentation schématique de l'oxydation du substrat par un système
laccase-médiateur (Cantarella et al., 2003) 21
Figure 2-8 : Schéma des configurations attendues du biocatalyseur en utilisant différentes procédures. Agents d'agrégation : le_Glutaraldehyde (Glu^— ) et le Glyoxal (Gly —). Proteines : Laccase ((_ ), Bovine serum albumine
(( .-D (Cabana et al., 2009) 24
Figure 2-9 : Les différentes approches pour la production d'enzymes immobilisées sans support : préparation par a) cristallisation (CLEC), b) agrégation (CLEA), c) séchoir atomiseur (CSDE), d) réticulation directe (CLE). (Adaptée de Cao et al.,
(2003)) 29
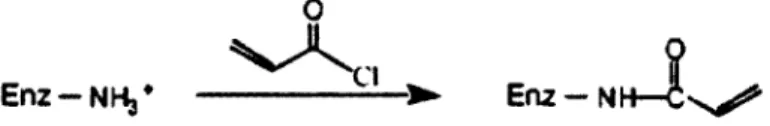
Figure 2-10 : Étapes de synthèse de SENs (adapté de Hegedus et Nagy, 2009) 29 Figure 2-11: modification de la surface de l'enzyme avec du chlorure d'acryloyle.
(adapté de Kim et Grate, 2003) 29
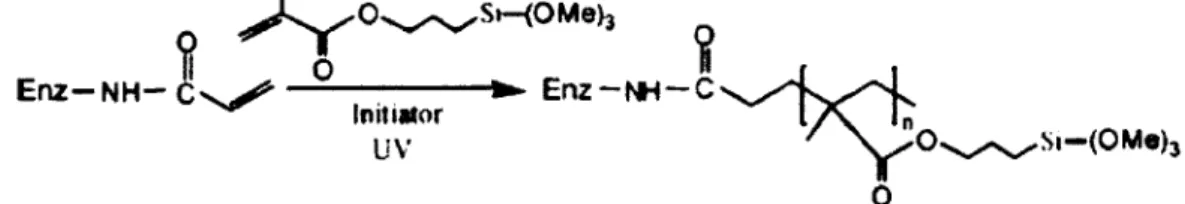
Figure 2-12 : Polymérisation linéaire sous rayonnement ultraviolet.(adapté de Kim et
Grate, 2003) 30
Figure 2-13 : Étapes de synthèse de SENs formés de chitosane et d'APTES (adapté de
Yadav et al. 2011 A) 32
Figure 3-1: Schematic (a) diagram for the preparation of EPES-lac and EPES-CLEA
and (b) the chemical reactions involved 42
Figure 3-2: Residual activity of
(i
) free laccase, (#) EPES-lac, ( •) CLEAs and (•) EPES-CLEAs at pH 3 and 40 °C (the values are means of triplicates ± StdFigure 3-3: Residual activity of ( £ ) free laccase, (• ) EPES-lac, ( • ) CLEAs and (+) EPES-CLEAs after a 4 h incubation with different chemical dénaturants at pH 3(the values are means of triplicates ± Std dev.) 46 Figure 3-4: The effect of pH on the activity of (a) ( | ) free laccase, ( #) EPES-lac, (b) (•) CLEAs and ( +) EPES-CLEAs (the values are means of triplicates ±
Std dev.) 49
Figure 3-5: The effect of temperature on the activity of (a) ( I) free laccase, (#) EPES-lac, (b) ( •) CLEAs and ( •) EPES-CLEAs (the values are means of
triplicates ± Std dev.) 50
Liste des tableaux
Tableau 3-1: Apparent Michaelis-Menten kinetic constants of free laccase, CLEAs, EPES-lac, and EPES-CLEAs for the oxidation of ABTS at pH 3 and
Liste des abréviations
Ao, t*room activité de la laccase au temps 0, au temps t, et à température pièce, ; ABTS, 2, 2'-azino-bis-(3-ethylbenzthiazoline-6-sulfonic acid);
APTES, 3-aminopropyltriethoxysilane; BPA, bisphénol A;
CA, carbonic anhydrase; CLE, cross-linked enzymes',
CLEA, cross-linked enzyme aggregate; CLEC, cross-linked enzyme crystal,
ED AC, l-ethyl-3-(3-dimethyl aminopropyl)-carbodiimide; EPES, enzyme polymer engineered surface;
EPES-CLEA, enzyme polymer engineered surface of CLEA; EPES-lac, enzyme polymer engineered surface of laccase; GLU, glutaraldéhyde;
GLY, glyoxal;
k^at, taux de réaction maximale par unité massique d'enzyme (coefficient catalytique); Km, constante d'affinité de Michaelis-Menten;
LiP, lignine peroxydase; LME, lignin modifying enzyme; MnP, manganèse peroxydase; NPE, nonylphénols polyéthoxylés ; NP, nonylphénol;
PE, perturbateurs endocriniens;
SEMs, scanning electron micrographs; SENs, single enzyme nanoparticle; TCS, triclosan;
U, quantité d'enzyme formant l^mol d'ABTS + par minute.
UV, ultra-violet;
VP, versatile peroxydase; WRF, white-rot fungi;
Chapitre 1
Les processus de fabrication industriels et les activités anthropogéniques génèrent un grand nombre de substances et molécules qui potentiellement peuvent polluer l'environnement (incluant l'air, le sol et l'eau) en induisant des impacts négatifs aux écosystèmes et la santé humaine.
L'environnement aquatique est l'ultime récepteur pour la majorité des composés chimiques émis par des activités anthropogéniques et ce, malgré les efforts d'épuration effectués à l'aide d'ouvrages de traitement des eaux. De nombreuses substances se retrouvent dans les effluents et les boues des stations d'épuration des eaux usées (Tan et al, 2007; Auriol et al, 2006). Parmi ces substances, on retrouve des composés qui ont le pouvoir de perturber le système endocrinien.
1.1 Perturbateurs endocriniens
Une substance perturbatrice du système endocrinien est un composé chimique exogène ou un mélange de ces composés qui altère une ou des fonctions du système endocrinien des organismes et qui, par conséquent, cause des effets néfastes sur la santé des organismes vivants, sur leur progéniture ou sur une partie de la population (Vos et al, 2000).
Ces perturbateurs endocriniens (PE) peuvent exercer leurs effets par différents mécanismes (Vos et al, 2000) :
En mimant ou en ayant comme antagoniste l'effet des hormones,
- En changeant le mécanisme de synthèse et de métabolisme des hormones, - Et, en modifiant les niveaux des récepteurs d'hormones.
À l'heure actuelle, les PE qui retiennent le plus l'attention sont ceux qui imitent les estrogènes naturels (comme le 17(3 estradiol qui est l'un des estrogènes les plus actifs) (Lagana et al, 2004), et qui sont suspectés d'être responsables de l'augmentation des perturbations dans la reproduction et le développement de l'homme, mais aussi de la vie sauvage (Saito et al, 2004). Ces perturbations se sont traduites, par exemple, par un retard de mue chez les nouveaux nés d'une espèce de crustacé (Daphnia magna) (Zou et Fingerman, 1997), la
féminisation de certains poissons (Sumpter, 2005), ou encore l'augmentation du risque de développer un cancer mammaire chez l'homme (Acevedo et al, 2005).
Ces PE peuvent se lier au récepteur à estrogène et réguler l'activité des gènes responsables de ces hormones. Ainsi, de tels effets ont augmenté les inquiétudes vis-à-vis des perturbations causées par une exposition prolongée à ces substances et ce, malgré leurs faibles concentrations dans l'environnement (Cooper et Kavlock, 1997). Une centaine de composés ont été identifiés comme perturbateurs endocriniens probables (Fromme et al., 2002; Fang et al, 2001). Parmi ces substances on retrouve le bisphénol A (figure 1-3), le triclosan, et le nonylphénol qui sont parmi les xénobiotiques organiques les plus détectés dans les zones urbanisées (Boyd et al, 2004; Boyd et al., 2003; Kolpin et al., 2002) et sur lesquels se concentrent les activités de notre groupe de recherche.
1.1.1. Nonylphénols
Les nonylphénols (NP) sont des composés organiques synthétiques (Figure 1-1) précurseurs dans la fabrication des nonylphénols polyéthoxylés (NPE). Les NPE sont synthétisés pour leurs propriétés tensioactives et sont largement utilisés dans l'industrie textile, la production de papier et sont également utilisés dans les peintures, la production de pétrole, et les nettoyants domestiques (Soares et al, 2008). Lors de leur biotransformations, les organismes qui les dégradent produisent des métabolites plus estrogéniques que les NPE (dont ultimement le NP) et qui sont de 2 à 200 fois plus toxiques que les NPE (Soares et al, 2008) . Les produits intermédiaires et finaux du métabolisme sont plus persistants que les NPE parents, et ils s'en retrouvent donc dans l'environnement notamment dans les sols, les boues d'épandage, les eaux souterraines, les rivières et les sédiments (Soares et al, 2008). Les NP sont connus pour s'accumuler dans les tissus des poissons, et d'autres organismes vivants, et se retrouvent ainsi dans la chaîne alimentaire (Soares et al, 2008).
Les NP présentent une activité œstrogènique (bien qu'il soit 1000 à 10 000 fois moins oestrogénique que l'œstradiol) (Sumpter et Jobling, 1995) qui peut notamment : 1) affecter la santé reproductive en entraînant la féminisation des poissons (induction de vitellogénine chez la truite mâle) (Sumpter et Jobling, 1995), 2) accroître significativement le risque de cancer mammaire chez la souris (Acevedo et al, 2005), ou encore 3) détériorer l'ADN dans le sperme humain (Harréus et al, 2002).
D'une façon générale, il y a peu de données disponibles concernant l'efficacité des techniques de traitement d'effluents domestiques et industriels sur les nonylphénols. De façon générale, les éléments suivants caractérisent le traitement des NP présent dans les eaux usées (Danish Environmental Protection Agency, 2002):
A. Le rendement d'élimination des NP de la phase aqueuse par les stations d'épuration des eaux usées, se situe typiquement entre 20 et 80 %, en fonction du type de traitement (les meilleurs rendements étant réalisés dans les filières les plus complètes) ; B. Des rendements supérieurs à 90 % peuvent être obtenus mais ils rendent nécessaire le
recours à des techniques extrêmement coûteuses (ex. charbon activé) ;
C. L'élimination des NP des eaux usées se traduit vraisemblablement par son transfert dans vers les boues (Environnement Canada et Santé Canada, 2001). Ceci se traduit par un transfert de la contamination,
1.1.2. Triclosan
Le triclosan (TCS) (Figure 1-2) est un agent antifongique et antibactérien à large spectre (Mezcua et al, 2004). On le trouve dans des produits de soins corporels (ex. savons, dentifrices) et beaucoup de produits de consommation courante en sont imprégnés tels que des: ustensiles de cuisine, des jouets, de la literie, etc. (Mezcua et al, 2004).
cr ^
^ xi
Peu d'éléments sont connus à propos du potentiel de perturbations endocriniennes associées au TCS. Ses effets ont notamment été observés chez les amphibiens (Rana catesbeiana, Xenopus laevis, grenouille-taureau Nord-américaine) (Jackson et Sutton, 2008) (Veldhoen et ai, 2006) et des poissons (Medaka orziyas latipes) (Ishibashi et al., 2004) (Foran et al., 2000). Malgré ce manque d'informations, ce composé s'avère d'intérêt car sa structure chimique 1) est similaire à celle d'autres perturbateurs endocriniens (ex. le BPA) et 2) ressemble à d'autres substances extrêmement toxines telles les dioxines
D'autre part, l'Association médicale canadienne a demandé au gouvernement canadien d'interdire l'usage du triclosan dans des produits ménagers car il crée une résistance bactérienne et génère des produits dangereux (chloroforme) (Rule et ai, 2005). En effet des études ont rapprté que le TCS peut se combiner au chlore dans l'eau pour former du gaz de chloroforme (Rule et al., 2005), classé cancerigène par l'Agence de Protection Environnementale des États-Unis d'Amérique. Le TCS peut aussi réagir avec le chlore libre pour produire de plus petites quantités d'autres composés, tel que 2,4-dichlorophénol (Rule et al., 2005), La plupart de ces intermédiaires se convertissent en dioxine lors d'exposition aux rayonnements UV. Bien que seules de petites quantités de dioxines sont produites un grand intérêt leur est prêté en raison de l'extrême toxicité de certaines d'entre elles, ainsi que leur potentiel à jouer le rôle de perturbateur endocrinien.
La plupart des études traitant de l'élimination du TCS dans les stations de traitements des eaux usées se sont focalisées sur les concentrations en phase aqueuse. Les concentrations trouvées dans ces études suggèrent des efficacités d'élimination de 58-97% pour des installations de filtration et de 95-98% pour les usines de boues activées (Bester, 2003; McAvoy et ai, 2002 ;Thomas et Foster, 2005). D'autre part, un traitement aux UV a permis de réduire de 22% le TCS présent dans les effluents des usines de traitement des eaux (Hua et ai, 2005). Durant les traitements conventionnels des eaux usées, les molécules de TCS peuvent être éliminées de différentes manières. La volatilisation est considéré comme négligeable dûe à la faible pression de vapeur du TCS. (Sabaliunas et al., 2003). De même, l'hydrolyse (et autres transformations chimiques) et la photodégradation n'ont pas lieu de manière appréciable en raison, respectivement, de la stabilité thermodynamique du TCS (Sabaliunas et ai, 2003), et de la turbidité des eaux usées (Latch et ai, 2003).
Finalement, des études ont montré que le TCS demeure extractible des boues, laissant de ce fait la transformation biologique comme principal processus régissant le taux et l'ampleur de l'élimination du TCS (Sabaliunas et al, 2003). D'où l'intérêt de développer des technologies de remédiation permettant d'éliminer ce polluant des eaux usées.
1.1.3. Bisphénol A
Le bisphénol A (BPA) est un composé organique aromatique (Figure 1-3) utilisé comme monomère et plastifiant pour la fabrication de plastiques de type polycarbonate et des résines époxy. Le BPA est un xénoestrogène qui peut jouer un rôle dans l'altération de la fonction de reproduction, et du développement d'organes comme le cerveau ou le système cardio vasculaire (Calafat et al, 2005).
Massivement produit (environ trois millions de tonnes de BPA sont produites chaque année dans le monde (Calafat et al, 2005) et dispersé dans l'environnement depuis quelques décennies, il est déjà couramment retrouvé dans l'organisme d'une large majorité de la population américaine. En effet, 95 % des échantillons d'urine collectés auprès d'adultes américains contenaient des niveaux quantifiables de BPA (Calafat étal,, 2005).
Les premières études sur l'action oestrogénique du BPA datent des années 1930 (Takeuchi et al, 2004). Cette molécule aurait une affinité pour le récepteur humain d'oestrogènes elle serait environ 1 000 fois inférieure à celle de l'œstradiol (Routledge et Sumpter, 1996). Malgré ce fait, ses effets pourraient entraîner des problèmes de santé chez l'homme, tels que des anomalies de fonctionnement de certains organes, comme le pancréas (Ropero et al, 2008) ou la thyroïde (Moriyama et al, 2002).
D'autre part, des études ont montré (sans que d'éventuels liens de causalité directe soient déjà identifiés et compris) qu'un taux urinaire élevé de ces produits était corrélé avec un risque plus élevé notamment :
- de maladies cardiovasculaires (Melzer et al, 2010) i>n3
Bon nombre d'études ont été menées concernant les possibilités d'élimination du BPA des eaux usées. Ainsi, différents traitements physiques et chimiques peuvent être envisagés (adsorption sur boues activées et inactivée (Clara et al, 2003), oxydation électrochimique (Boscolo Boscoletto et al., 1994).
Actuellement, les usines de traitement n'étant pas conçues pour l'élimination du BPA, et devant le potentiel danger que représente cette substance, il apparait nécessaire de développer des méthodes de remédiation pour l'éliminer.
1.2. Similitudes structurales entre les PE
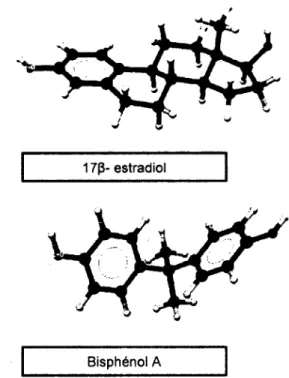
En principe, l'activité biologique d'un produit chimique est déterminée par sa structure (Fang et al., 2001). Une étude effectuée en 2001 a établi une relation entre la structure et l'activité (SAR) des PE (Fang et al, 2001). En comparant les différentes classes de perturbateurs endocriniens par une étude SAR, les auteurs ont pu établir 5 caractéristiques structurelles essentielles permettant à ces contaminants de se lier au récepteur humain d'estrogène. Pour ce faire, ils ont utilisés le 17P-œstradiol (estrogène humaine) comme molécule modèle. Il a été démontré que les éléments structuraux suivants favorisent la liaison des perturbateurs endocriniens au récepteur humain : (i) la capacité d'établir des ponts hydrogènes du noyau phénolique imitant le OH, (ii) le donneur H imitant le 17P-OH et la distance O-O entre le 3-OH et le 17|3-3-OH, (iii) centre hydrophobique stérique précis imitant les substituants stériques 7a- et 17P-, (iv) l'hydrophobicité, (v) la structure en cycles (Fang et al, 2001). La Figure 1-4 montre la similitude structurale existant entre le BPA et le 17p-œstradiol.
Les travaux de Fang et al. (2001) illustrent clairement la nécessité qu'il y ait présence d'un noyau phénolique dans la structure des PE afin de leur permettre de se lier aux récepteurs à oestrogenes. Ce faisant, la réduction de la capacité de perturbation endocrinienne de ces substances peut se faire via l'oxydation de ces structures phénoliques.
17(3- estradiol
Bisphénol A
Figure 1-4 : Similitudes structurales entre le bisphénol A et le 17p- œstradiol (adapté de Fang et al, 2001)
Devant cet état de fait, il s'avère essentiel de développer des stratégies de traitements des eaux usées novatrices permettant de transformer cette structure de par le fait que :
1) Les eaux usées domestiques et industrielles représentent une source importante d'émission de ces substances dans l'environnemen (Batt et al, 2007);
2) Les procédés conventionnels de traitement des eaux usées, tels que les boues activées, ne sont pas conçus pour l'élimination de contaminants à de faibles concentrations et sont donc considérés inadéquats pour l'élimination des sources d'émissions de PE dans l'environnement (Batt et al., 2007);
3) Une modification structurelle de ce groupement chimique permettra d'éliminer le potentiel des ces contaminants à ce lier au récepteur humain d'estrogène et par extension réduira les risque pour la santé associés à la présence de ces molécules dans l'environnement.
1.3. Techniques d'élimination des composés phénoliques
Mondialement, le phénol est produit à raison de 6 millions de tonnes/an en moyenne (Busca et al., 2008), et ce chiffre ne cesse d'augmenter. En raison de son usage en industrie, le phénol est présent dans les eaux usées à différentes concentrations, et ce selon l'industrie de laquelle il est issu (Busca et al., 2008).
Ainsi, et en raison du danger que représentent ces composés phénoliques, les eaux usées ne peuvent rejoindre l'environnement aquatique récepteur sans traitement préalable. Des normes de purification des eaux usées vis-à-vis de ces substances ont été établies. Ainsi, l'Union européenne a fixé comme limite de concentration en phénol 0,5 jxg/1, et 0,5 mg/1 respectivement pour l'eau potable et les eaux usées (Busca et al., 2008). D'autre part, le Canada d'ici 2012 interdirait le rejet de BPA provenant des effluents industriels à des concentrations supérieures à 1,75 ng/L (Gazette du Canada, 2010), et le Comité scientifique de l'alimentation humaine, a limité une dose quotidienne admissible provisoire du bisphénol A à 10 ng/kg pc/jour (CSAH, 2002).
Différentes approches peuvent être envisagées pour l'élimination du phénol. On peut ainsi adopter des techniques de séparation du phénol des effluents (adsorption, distillation, etc.), ou bien encore sa dégradation (oxydation à l'ozone, au peroxyde d'hydrogène, réduction biochimique, etc.). De manière générale ces techniques présentent des inconvénients dues au transfert de pollution (adsorption, distillation), aux coûts élevés associés à ces techniques (ex : distillation), ou encore à l'utilisation et production de substances plus toxiques que la substance d'intérêt (ex : réaction homogène conventionnelle de fenton) (Busca et al., 2008). Depuis quelques décennies, des efforts ont été déployés afin de mettre en place des stratégies de bioremédiation pour le traitement de différentes matrices contaminées. Ces efforts ont été motivés par le désir, et la nécessité de :
Fiabiliser, et faciliter la mise en œuvre des procédés de décontamination, ainsi que la réduction des coûts qui y sont associés (Karam et Nicell, 1997);
- Générer des produits environnementalement plus acceptables, et limiter la production de composés issus de réactions parrallèles (Rodriguez Couto et Toca Herrera, 2006; Ahuja et al., 2004).
Toutefois, les stratégies biologiques actuelles de traitement des eaux usées pour les perturbateurs endocriniens ciblés précédemment s'avèrent inefficaces ; il en résulte un transfert de pollution (ex : des eaux vers les boues) ou une inefficacité d'élimination de la phase aqueuse (Auriol et al., 2006).
Face à ces constats, le développement de bioprocédés voués à l'élimination des PE par l'action d'enzymes s'avère extrêmement prometteur (Auriol et al., 2008; Cabana et al., 2007A). La biocatalyse enzymatique présente différents avantages car ces protéines sont caractérisées par la spécificité des substrats qu'elles peuvent utiliser, par leur fonction spécifique de catalyseur, par leur capacité à être utilisées sur une large gamme de conditions environnementales (pH, température, salinité, concentrations, présence de composés toxiques), par leur faible sensibilité à la présence de prédateurs, d'inhibiteurs et de variation des conditions du milieu, et par la simplicité et la facilité de contrôle des procédés utilisant des enzymes (Ahuja et al., 2004; Gianfreda et Rao, 2004; Karam et Nicell, 1997; Nannipieri et Bollag, 1991). De plus, certains facteurs spécifiques à un site peuvent favoriser l'utilisation d'enzymes pour la bioremédiation, tels que l'absence ou la faible densité de microorganismes capables d'éliminer la substance d'intérêt, la température, l'humidité, le conditions d'oxygénation, la présence d'éléments nutritifs, la biodisponibilité des contaminants et l'écologie microbienne du milieu (Ahuja et al., 2004).
Dans ce contexte, la biocatalyse enzymatique constitue un traitement alternatif pour éliminer différents xénobiotiques qui peut s'avérer attractif pour différents motifs. On peut citer à ce titre que :
les avancées techniques permettent la production plus rapide et à moindre coût d'enzymes grâce aux percées dans le domaine du génie génétique, à un meilleur isolement/sélection de souches, et à de meilleures procédures de purification (Karam et Nicell, 1997; Elisashvili et Kachlishvili, 2009) ;
les composés formés par biocatalyse peuvent s'avérer environnementalement plus acceptables que les produits formés par des procédés conventionnels (Rodriguez Couto et Toca Herrera, 2006; Ahuja et al, 2004) ;
effet, ces microorganismes sécrètent des enzymes oxydatives ayant une grande versatilité vis-à-vis du spectre de contaminants qu'ils peuvent transformer.
Parmi les enzymes lignivores un intérêt grandissant a été porté sur la laccase (EC 1.10.3.2). Cette dernière est une phenoloxydase catalysant l'oxydation d'un grand nombre de composés organiques tels que des méthoxyphénols, des phénols, des o- et p- diphénols, des aminophénols, des polyphénols, des polyamines, des molécules provenant de la lignine et certains ions inorganiques (Burton, 2003; Call et Mucke, 1997). De plus, cette enzyme peut catalyser la déméthylation de la lignine, des acides méthoxyphénols et des méthoxyaromatique. La laccase a également été identifiée comme un biocatalyseur impliqué dans la dégradation de perturbateurs endocriniens et d'autres contaminants tel que : les hydrocarbures aromatiques, les colorants polycycliques (Haglund et al, 2002), mais aussi le bisphénol A, le triclosan, et le nonylphénol (Cabana et al. 2009; Hundt et al., 2000), qui sont caractérisés par la présence d'un groupement phénolique dans leur structure chimique, qui rend possible l'utilisation de la laccase pour oxyder et bio-éliminer ces composés.
Malgré les nombreux avantages que présente la catalyse enzymatique, son utilisation reste limitée pour différentes raisons. L'un de ces facteurs est le coût élevé associé à la production, l'isolement, et la purification de ces enzymes. Toutefois, certains auteurs prétendent que les coûts associés à l'utilisation de ces protéines dans des biotechnologies environnementales peuvent s'apparenter aux coûts des technologies de traitement dites ex situ (Trombly, 1995). L'instabilité des protéines, qui résulte en une durée de vie de l'activité biocatalytique relativement faible dans différentes matrices contaminées, constitue elle aussi un frein à leur utilisation en raison de l'augmentation des coûts engendrés par de telles stratégies.
Diverses méthodes ont été élaborées à des fins de stabilisation enzymatique. Ainsi, diverses formes d'immobilisation d'enzymes sont possibles (physiques, chimiques). Parmi elles, on retrouve une technique d'insolubilisation par la formation d'agrégats d'enzymes réticulés [cross-linked enzyme aggregates (CLEAs)]. Cette approche, de plus en plus utilisée, ne nécessite pas de support solide et produit des biocatalyseurs avec une grande activité spécifique (Sheldon, 2007). En marge des avantages que peuvent offrir ces méthodes, l'immobilisation enzymatique présente aussi des inconvénients. On peut citer à ce titre :
Diminution du transfert de masse entre la solution et le site catalytique de l'enzyme (Krajewska, 2004) ;
- Baisse de l'activité enzymatique comparativement à l'enzyme libre (Krajewska, 2004);
- Difficulté potentielle de stabilisation vs des dénaturants chimiques (Krajewska, 2004). De nouvelles techniques utilisant des structures polymeriques greffées de manière covalente à la surface de l'enzyme ont été mises au point afin d'améliorer les performances en matière de stabilité enzymatique (Kim et ai, 2008). Cette approche prometteuse a permis une importante stabilisation des enzymes utilisées tout en ne limitant pas le transfert du substrat de la solution vers le site actif de l'enzyme (Yadav et al, 2011A; Hegedus et Nagy, 2009; Kim et Grate, 2003).
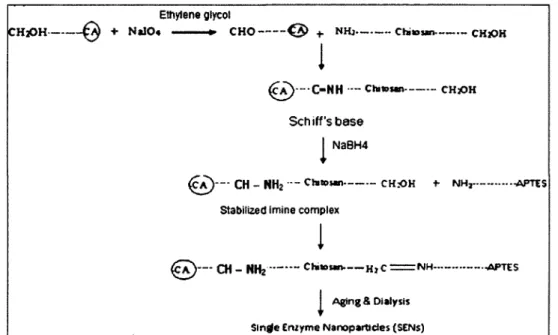
Ainsi, Kim et Grate (2003) ont mis au point une structure polymérique de taille nanométrique contenant des enzymes individuelles en utilisant la chymotrypsine et la trypsine comme enzymes modèles. En clair, chaque enzyme est entourée d'une structure organique/inorganique poreuse de taille nanométrique. La synthèse de cette structure nommée SENs (single enzyme nanoparticles) requière trois étapes : i) modification de la surface de l'enzyme ; ii) polymérisation linéaire des monomères de vinyle et de trimethoxysilane ; iii) polymérisation orthogonale pour relier entre eux les groupes trimethoxysilane pendants (Kim et Grate, 2003). Les SENs formés ont permis d'augmenter la demi-vie de la chymotrypsine de 12h à 140 jours.
Une structure semblable à celle des SENs formée de chitosane et de 3-aminopropyltriethoxysilane (APTES), a été mise au point par Yadav et al. (201 la, 201 lb). Ce groupe a rapporté que l'anhydrase carbonique ainsi modifiée est restée stable pendant plus de 100 jours à -20°, 4° ou 20°C.
Au vu des différents avantages que présentent les CLEAs et la technique des SENs, il apparaît fort intéressant de développer des biocatalyseurs combinant ces deux approches. Ceci permettrait de tirer profit des avantages complémentaires que présentent les CLEAs et les SENs.
1.2. Objectifs
1.2.1. Objectif général
L'objectif général de ces travaux est d'augmenter la stabilité de la laccase vis-à-vis des dénaturants potentiellement présents dans des matrices environnementales (pH, température, sels, solvants organiques, etc.), et qui pourraient mettre en péril leur efficacité à éliminer des xénobiotiques.
Particulièrement, l'objectif de mon mémoire est de synthétiser et caractériser une structure stabilisatrice (enzyme polymer engineered surface (EPES)) semblable à celle des SENs, formée à partir de laccase de Trameîes versicolor qui sera entourée d'un réseau polymérique formé de chitosane et d'APTES.
1.2.2. Objectifs spécifiques
De manière plus spécifique ce projet vise à :
- Démontrer la possibilité de synthétiser des EPES de laccase (EPES-lac) et de CLEAs formées de laccase (EPES-CLEA).
- Caractériser des EPES formés (paramètres cinétiques, morphologie, résistance aux dénaturants, etc.)
1.3. Contributions originales :
Les travaux réalisés lors de ce projet ont été présentés à différentes occasions dans des colloques et conférences, et ont également été publié sous forme d'articles en 2011 et 2012.
1.3.1. Articles
T. Hassani, S. Ba, H. Cabana (2012). Formation of Enzyme Polymer Engineered Structure for Laccase and Cross-Linked Laccase Aggregates Stabilization. (Article soumis à Bioresource Technology)
S. Ba, A. Arsenault, T. Hassani, J.P. Jones , H. Cabana (Article sous presse). Laccase immobilization and insolubilization; from fundamentals to applications in wastewater bioremediation. Critical Reviews in Biotechnology.
1.3.2. Présentations orales
H. Cabana, T. Hassani, J.-F. Leduc (2011). La présence de substances perturbatrices du système endocrinien dans l'environnement aquatique : tout un défi à relever pour l'épuration des eaux usées et potables. 79e Congrès de l'ACFAS. Sherbrooke, Qc, Canada. 9-13 May.
H. Cabana, A. Arsenault, T. Hassani, J. P. Jones (2010). Élimination de micropolluants des eaux usées : Et si Champignac avait la solution? 33e symposium sur les eaux usées, Réseau
Environnement. Saint-Hyacinthe, Qc, Canada, 26-27 octobre.
1.3.3. Présentations par affiche
T. Hassani, H. Cabana (2011). Développement d'un biocatalyseur enzymatique pour l'élimination de polluants émergents phénoliques : stabilisation de laccase via la formation d'un réseau polymérique nanométrique (SEN's). 79e Congrès de l'ACFAS. Sherbrooke, Qc,
Canada. 9-13 May.
T. Hassani, H. Cabana (2011). Free laccase and cross-linked laccase aggregates armored by an organic/inorganic polymeric network : Toward the formation of robust biocatalysts. 4th
International Conference Enzymes in the Environment, Activity, Ecology & Applications. Bad Nauheim, Germany. 17-21 July.
T. Hassani, H. Cabana (2011). Stabilisation de biocatalyseurs versatiles pour l'élimination de polluants émergents phénoliques via la formation d'un réseau polymérique nanométrique (SEN). 26 th Eastern Canadian Symposium on Water Quality Research, Canadian association on Water quality . Québec (Qc), Canada, 7 octobre.
1.4 Structure du manuscrit
Le chapitre 2 comporte une revue de littérature en lien avec les enzymes ligninolytiques, la laccase, l'immobilisation enzymatique, la technique de formation des SENs et son application pour former des EPES de laccase (EPES-lac) et des EPES de CLEAs (EPES-CLEA). Le chapitre 3 est présenté sous forme d'article qui résumé les travaux effectués ainsi que les résultats de caractérisation des EPES et leur discussion. Finalement, le manuscrit se termine par une conclusion qui apporte quelques pistes de travaux futurs.
Chapitre 2
2.1. La laccase
2.1.1. Origine : champignons lignivores
La laccase est une enzyme principalement trouvée dans les plantes, les champignons (Baldrian, 2006), et certains travaux mentionnent également l'existence de laccases bactériennes (Alexandre et Zhulin, 2000). Des protéines avec des caractéristiques typiques de laccases ont récemment été, aussi identifiées chez des insectes (Kramer et al., 2001). Cette enzyme est, entre autres, sécrétée par des champignons (Figures 2-1 et 2-2) responsables de la pourriture blanche du bois (Figure 2-3) (white rot fungi, WRF). Bien que connues depuis longtemps (depuis la fin du 19ème siècle) (Baldrian, 2006), la laccase a attiré une attention
considérable seulement après le début des études de la dégradation enzymatique du bois (Baldrian, 2006). Récemment, la capacité de ces champignons à biodégrader une variété de xénobiotiques et des polluants récalcitrants a généré un intérêt considérable dans le domaine de la recherche en microbiologie industrielle/environnementale.
Figure 2-1 : Pleurotus ostreatus Figure 2-2 : Trametes Versicolor
Les WRF sont des mycètes uniques de par leur aptitude à sécréter un système d'enzymes oxydatives ayant la capacité de dégrader la lignine. Second composé contribuant à la biomasse (Schimmel et al., 1994), la lignine est un polymère aromatique hétérogène présentant une très grande résistance aux agents chimiques, et à la dégradation biologique. Cette résistance extrême est due à sa structure très complexe, il s'agit du composé naturel organique le plus difficilement biodégradable (Hammel, 1997).
Système enzymatique lignivore
La structure hétérogène de la lignine implique que les enzymes ligninolytiques possèdent la capacité d'oxyder des substrats de structures chimiques variées possédant des potentiels redox élevés et ce, de façon non spécifique (Evans et Hedger, 2001 ). Cette capacité oxydative non-spécifique ainsi que leur potentiel redox élevé représentent des caractéristiques intrinsèques de ces protéines favorisant leur utilisation dans des biotechnologies.
Ce système enzymatique extracellulaire est généralement constitué de deux peroxydases, la lignine peroxydase (LiP) et la manganèse peroxydase (MnP) et d'une phénoloxydase, la laccase (Wesenberg et al, 2003). Certaines souches de WRF sécrètent également de la peroxydase versatile (PV) qui a la capacité d'oxyder les substrats types de la LiP et de la MnP (Ruiz-Duefias et al, 1999).
Alors que la LiP clive les liens aromatiques non phénoliques de la lignine, la MnP et les laccases fongiques peuvent seulement oxyder les composés phénoliques de la lignine. Contrairement aux enzymes lignolytiques LiP et MnP qui requièrent du peroxyde d'oxygène pour être activées, la laccase requière, elle, de l'oxygène comme récepteur final d'électrons. Cette capacité à dégrader la lignine leur permet aussi de transformer une grande gamme de polluants ayant des structures similaires à celle de la lignine: HAP, phénols chloreux, dioxines, pesticides et des colorants.
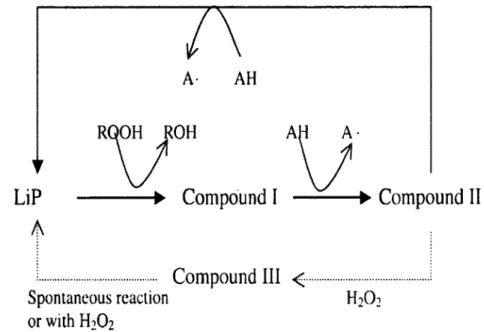
a- Lignine peroxydase
La LiP est une glycoprotéine excrétée dans le milieu (Tien et al, 1986), non-spécifique et capable d'oxyder des composés aromatiques à haut potentiel d'oxydo-réduction (voir figure 2-4). Elle est capable de catalyser une dépolymérisation partielle de la lignine (Jaouani et al, 2003).
La LiP qui peut être impliquée dans le clivage de lien C-C de la lignine, agit à distance en utilisant un médiateur, qui va envahir la superstructure de la lignine et y amorcer une série de réactions radicalaires aboutissant au démantèlement du polymère (Galliano et al, 1991). Elle pourrait également se lier à la lignine et y initier des processus de transfert d'électrons à grande distance (Jaouani et al., 2003).
A- AH
LiP
A
ROOH ROHCompound I
> Compound II
Compound III <•
Spontaneous reaction H2O2
or with H2O2
Figure 2-4 : Le cycle catalytique de la LiP. AH représente le substrat de l'enzyme, et A°, le substrat oxydé par l'enzyme (Shah et Nerud, 2002).
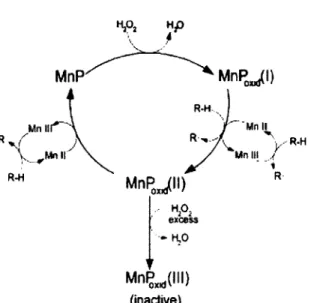
b- Manganèse peroxydase
La MnP est une glycoprotéine secrétée sous de multiples isoformes (Leisola et al, 1987) et dont la fonction est de génerer du Mn3+ à partir de Mn2+. Le cation Mn3+, qui est un oxydant
fort, s'attaque préférentiellement aux portions aromatiques de la lignine (Galkin et al., 1998; Wariishi et al., 1992). Il y a production d'acides organiques qui vont stabliser le Mn3+, et ce
complexe va s'insérer dans la structure et provoquer ainsi un clivage (Kishi et al., 1994; Glenn etal., 1986).
Hp MnP" MnP^I) Mn II r MnP0JH) 'Mn II
^ V
Mn III /R
-
H
" t R HA excess - K.0 Mn^dH) (inactive)Figure 2-5: Le cycle catalytique de la MnP. RH représente le substrat et R° représente le substrat oxydé par l'enzyme (Lôpez et al, 2002).
Le VP est capable d'oxyder le Mn2+ en Mn3+ et de dégrader les composés non phénoliques
modèles mais avec une affinité moindre que celle de la LiP. Elle est également capable d'oxyder les hydroquinones et les phénols substitués qui ne sont pas oxydés efficacement ni par la LiP ni par la MnP en absence de l'alcool vératrylique et de Mn2+ respectivement
(Jaouani et al, 2003).
2.1.2. Propriétés de la laccase
Les laccases appartiennent au groupe des phenoloxydases contenant des atomes de cuivre dans le centre catalytique et sont souvent appelées oxydases à multiples atomes de cuivre. Elles contiennent généralement trois types d'atomes de cuivre (Baldrian, 2006).
À ce jour plusieurs dizaines de laccases issues de souches fongiques ont été purifiées (Baldrian, 2006). Ces protéines à cuivre sont caractérisées par un poids moléculaire, une température optimale et un point isoélectrique moyens respectivement de 66 kDa, 3.9 et 55°C (Baldrian, 2006).
Biochimiquement, la laccase est une enzyme qui oxyde une variété de composés aromatiques donneurs d'hydrogène. Ainsi, elle catalyse l'extraction d'un électron et d'un proton aux hydroxyles phénoliques ou aux groupes amino aromatiques pour former des radicaux phenoxy libres et des radicaux amino, respectivement. D'ailleurs, cette enzyme ne fait pas qu'oxyder
les acides phénoliques et les acides methoxyphenoiiques, mais agit aussi en les decarboxylant, elle attaque leurs groupes methoxyl à travers la démethylation (Leonowicz et al, 1984),ou la démethoxylation (Potthast et al., 1995). Ces réactions peuvent représenter une étape importante dans la transformation initiale du polymère de lignine.
La large spécificité vis-à-vis des donneurs d'électrons lui confère des possibilités intéressantes à être utilisées pour l'élimination de composés aromatiques, notamment phénolique, des eaux usée. L'oxydation de composés phénoliques génère des radicaux phenoxy, et des quinoides intermédiaires, qui sont, plus tard, transformés en dimères et polymères insolubles (Nannipieri et Bollag, 1991; Liu etal., 1985).
Les laccases sont constituées d'au moins un atome de cuivre de type 1 (Tl) avec 3 atomes de cuivre additionnels : le premier de type 2 (Antorini et al, 2002), et les deux derniers de type 3 (D'Annibale et al, 2000) arrangées en un groupe de 3 noyaux. Les cuivres sont toujours attachés à l'histidine, à la leucine et à la cystéine. Les cuivres de type 2 et 3 forment un site de stabilisation de l'oxygène lors de sa réduction en eau. Le cuivre de type 1 est impliqué dans la réduction du substrat et les électrons générés sont alors transférés aux cuivres de type 2 et 3 (Call et Mucke, 1997).
L'intermédiaire natif de l'enzyme possède ses quatre cuivres sous forme oxydée Cu2+ (voir
Figure 2-6). Une première molécule de substrat réduit le cuivre de type 1. Le groupement trinucléaire peut alors être réduit suivant deux mécanismes possibles. Dans le premier mécanisme, le cuivre de type 1 transfert son électron au cuivre de type 2. Le premier est alors à nouveau réduit par une molécule de substrat. Ensuite, les cuivres de type 1 et 2 réduisent en association les cuivres de types 3. Une troisième molécule de substrat réduit le cuivre de type 1 qui passe son électron au cuivre de type 2 avant d'être une dernière fois réduit par une quatrième molécule de substrat. Dans le deuxième mécanisme, le cuivre de type 1 passe séquentiellement ses électrons au groupement trinucléaire tandis que quatre molécules de substrat le réduisent au fur et à mesure. Quel que soit le mécanisme utilisé, l'enzyme se trouve entièrement réduite après ces transformations (Lê, 2004).
La réduction de l'oxygène en eau permet de régénérer l'enzyme native et se passe en deux étapes. Chacune de celle-ci est caractérisée par un échange de deux électrons. La première
étape est déterminante de la vitesse. Les cuivres de types 2 et 3 agissent d'abord et il y a formation de peroxyde intermédiaire. Dans la deuxième étape, le peroxyde est utilisé pour réduire le cuivre de type 2 (Cole et al, 1990). Il est manifeste que le cuivre de type 2 est indispensable à la réactivité du dioxygène. On remarque aussi que la réduction du dioxygène se produit en absence du cuivre de type 1. Le cuivre de type 1 n'est ainsi clairement pas nécessaire pour la réactivité du dioxygène ; en son absence, un intermédiaire sera quand même formé. Ceci démontre que le groupement cuivrique trinucléaire de type 2/3 représente le site actif pour la liaison et la réduction du dioxygène (Cole et al, 1990).
reduced _ „ laccase yPCiT TypeS Cu+0 ciAyp0 3 resting co-enzyme + peroxide-level Cu intermediate ,H,0 Cu "r Cu 0 Cu i ^2* native intermediate
Figure 2-6 : Cycle catalytique de la laccase (d'après Wesenberg et al, 2003)
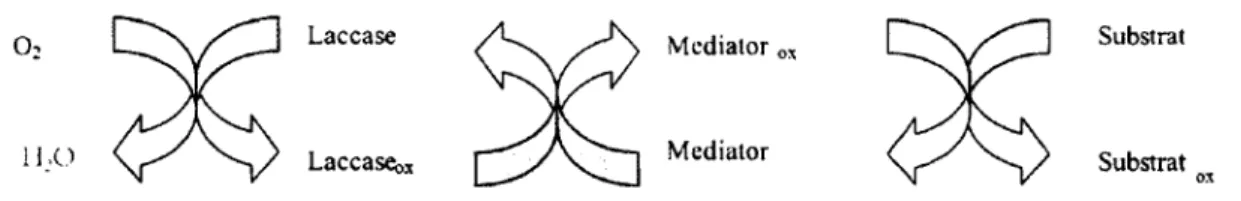
Le potentiel redox de laccases (450-800mV) ne permet pas d'oxyder des composés à haut potentiel redox tels que certains HAP et colorants synthétiques. L'utilisation de médiateurs redox a permis de passer outre cette limitation. Un médiateur est une molécule de faible poids moléculaire qui agit comme une sorte de « transporteur d'électron», une fois qu'il est oxydé par l'enzyme, il diffuse du site catalytique de l'enzyme et à son tour oxyde des substrats qui grâce à leur taille ne peuvent pas directement entrer dans ce site (Fabbrini et al, 2002). Une représentation schématique du système laccase-médiateur est proposée à la Figure 2-7.
Laccaseoi Laccase
Mediator
Mediator 0, Substrat
Substrat
Figure 2-7 : Représentation schématique de l'oxydation du substrat par un système laccase-médiateur (adapté de Cantarella et al, 2003)
Dans le cas de la dégradation de la lignine, des restrictions de taille gênent la pénétration de l'enzyme dans la structure du polymère. Les médiateurs oxydés par la laccase sont capables de diffuser dans la structure de la lignine et agissent comme de forts agents oxydants.
Parmi les médiateurs du système laccase-médiateur, citons le 2-2'-azino bis(3-ethylbenzthiazoline-6-sulpnonate) (ABTS), et 1-Hydroxy-benzotriazole (HBT).
En présence de médiateurs tels que l'ABTS ou le 1-HBT, la laccase peut oxyder des composés non phénoliques comme l'alcool vératrylique (substrat naturel de la LiP) (Bourbonnais et Paice, 1990).
Malgré l'efficacité de la laccase à éliminer des perturbateurs endocriniens tel que le NP, TCS, et le BPA, l'utilisation d'enzymes libres dans des applications de biotechnologie environnementales s'avère très limitée. En effet, une telle approche nécessite l'utilisation d'une grande quantité d'enzymes libres pour le traitement d'effluents liquides. De plus, ces enzymes, sous leur forme soluble, sont sensibles à l'effet de différents dénaturants physiques, chimiques et biologiques présents (Rodriguez Couto et Toca Herrera, 2006) dans ces effluents réels et s'avèrent impossibles à confiner dans un espace restreint de façon à les utiliser de manière continue.
L'immobilisation de laccases, via des liaisons covalentes, provenant de différentes souches de WRF a permis d'augmenter grandement la stabilité de ces enzymes. C'est ainsi que la stabilité thermique (Luterek et al., 1998), la stabilité au pH (Al-Adhami et al, 2002), et aux agents dénaturants chimiques(Luterek et al, 1998), et biologiques (D'Annibale et al., 2000) ont pu être améliorées par différentes méthodes d'immobilisation covalentes.
Une récente étude (Forde et al., 2010) s'est intéressée à la stabilisation de deux différents laccases (Myceliophthora thermophila et Trametes hirsuta). Ces enzymes ont été soumises à
éthylène-glycol-JV-hydroxy succinimide (EGNHS) et glutaraldehyde et (ii) par de l'anhydride citraconique monofunctionnel, et les formes non traitées et chimiquement modifiées des deux enzymes ont été alors immobilisées sur trois types différents de particules de silice mésoporeuse.
Une augmentation de la stabilité thermique et de l'activité ont été observées sur la souche M. thermophila respectivement lorsque traiteées avec de l'EGNHS et l'anhydride citraconique, et lors d'immobilisation sur le MPS, la plus grande augmentation de la stabilité a été enregistrée pour la souche M. thermophila. traitée avec du glutaraldehyde. Des observations similaires ont été faites sur la souche T. hirsuta, mais avec des gains moins importants en stabilité et activité. (Forde et al., 2010). Cette augmentation de la résistance à la dénaturation offre un outil prometteur pour augmenter l'applicabilité, la stabilité que la réutilisabilité de laccase dans l'industrie biotechnologique (Forde et al., 2010).
2.2. Stabilisation enzymatique
Afin de promouvoir l'usage de la catalyse enzymatique dans des applications bio-environnementales différentes approches ont été développées en matière de stabilisation d'enzymes (D'Annibale et al, 2000).
L'utilisation d'enzymes libres s'avère très limitée, car une telle approche nécessite l'utilisation d'une grande quantité d'enzymes solubles pour le traitement d'effluents liquides de par le fait qu'il s'avère impossible de les confiner dans un bioréacteur utilisé en continu. De plus, ces enzymes, sous cette forme libre, sont sensibles à l'effet de différents dénaturants physiques, chimiques et biologiques présents dans ces effluents réels et s'avèrent impossibles à confiner dans un espace restreint de façon à les utiliser de manière continue (Duran et al, 2002). Néanmoins, ces contraintes peuvent être contournées via l'immobilisation de l'enzyme sur un support solide ou via la réticulation des agrégats d'enzymes qui permettent de produire des biocatalyseurs réutilisables et stabilisés. De plus, depuis peu la synthèse d'un réseau polymérique englobant individuellement l'enzyme a permis de développer des biocatalyseurs extrêmement stables (Kim et Grate, 2003).
2.2.1 Immobilisation enzymatique
L'immobilisation enzymatique consiste dans le confinement physique des protéines dans une matrice donnée à des fins de catalyse continue.
En effet, elle permet de générer des biocatalyseurs réutilisables et, par extension, de les utiliser en continu dans des bioréacteurs. De plus, l'immobilisation d'enzymes permet de stabiliser ces protéines contre l'effet dénaturant de différents sels, solvants et enzymes protéolytiques (O'Fâgâin, 2003). Par conséquent, cette approche s'avère avantageuse pour des applications bio-environnementales.
Diverses formes d'immobilisation enzymatique sont possibles. Ces modes d'immobilisation peuvent être divisés en fonction du type de liens impliqués. En ce sens, il y a des moyens chimiques et physiques. Les supports utilisés peuvent être d'origine organique ou inorganique, et être une substance naturelle ou synthétique.
Il n'y a pas de concept systématique pour trouver la méthode d'immobilisation la plus appropriée pour les différents biocatalyseurs.
La principale tâche est de définir : i. Un support adéquat ;
ii. Des conditions appropriées (pH, température, etc.) ;
iii. Une enzyme adaptée afin de rencontrer les attentes du biocatalyseur immobilisé (Cao, 2005).
Pour des applications enzymatiques où la valeur ajoutée est faible, il est impératif d'utiliser un support peu onéreux, facile à manipuler, stable chimiquement et mécaniquement, non altérable par l'action de microorganismes, avec une surface spécifique élevée et pouvant être recyclé (Cao, 2005).
Immobilisation par liaisons covalentes
L'immobilisation par liaisons covalentes est une méthode chimique qui repose sur la formation de liens covalents formés entre un groupement fonctionnel présent sur le support
d'acides aminés non-essentiels est la méthode la plus utilisée pour immobiliser des enzymes. Les résidus d'acides aminés qui ne sont pas impliqués dans le site actif ou le site de liaison au substrat de l'enzyme à immobiliser peuvent être utilisés pour des liaisons covalentes avec différents supports (Aehle, 2004).
Lors de l'immobilisation de l'enzyme par la formation de liens covalents entre l'enzyme et le support, il est impératif que la protéine maintienne sa conformation active qui est assurée par sa structure tridimensionnelle. En effet, les liens définissant les structures secondaires, tertiaires et quaternaires de la molécule dépendent de la structure en acides aminés de la protéine. Cette structure tridimensionnelle est assurée par des interactions non covalentes à l'exception des liens disulfures.
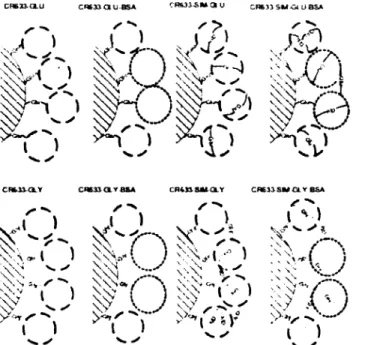
L'immobilisation d'enzymes sur des supports insolubles peut être effectuée via deux étapes : (i) le support est activé de façon à réagir directement avec les groupes fonctionnels de la protéine, (ii) un agent de liaison (agent réticulant tel que le glutaraldéhyde ) est utilisé pour lier la protéine au support (voir figure 2-8).
CH63J-OLU CW330HJ-BSA CR633-S»*au CR631 SM-Gi 0 BSA
• C>
mm
•Of
CP
s. •CR&iVOLY C«W3iGLVaSA CR4»SAKXY CRÉÎSSMCir BSA
f i ' " i
O
w ^ ^ -v <\\ h /- ^ yN <-< < v ; s V-l 1 v^'(
î(\
)
\v' \>y \J
\V \>/ A. ) y<*r S y*' * >>". J ^ s Y ;>V '-**^ J
w wFigure 2-8 : Schéma des configurations attendues du biocatalyseur en utilisant différentes procédures. Agents d'agréggtion : le Glutaraldehyde (GIut^— ) et le Glyoxal (Gly —*).
Ces liens sont des interactions dites fortes et procurent ainsi une fixation de l'enzyme stable et dans certains cas, réduisent la vitesse de désactivation de l'enzyme et peuvent altérer utilement la spécificité de l'enzyme.
Cependant, l'immobilisation enzymatique via des liaisons covalentes tend à réduire l'activité et l'affinité de l'enzyme, puisque les liens formés peuvent perturber la structure native de la protéine.
Cette diminution de l'affinité vis-à-vis de certains substrats peut, dans certains cas, être aussi expliquée par un problème d'encombrement stérique résultant de l'immobilisation, qui cause un problème de substrat vers le site actif de l'enzyme. De plus, cette diminution de l'affinité peut être associée à une torsion de la structure tridimensionnelle de l'enzyme (Durante et al., 2004).
Immobilisation par adsorption physique
L'immobilisation par adsorption est basée sur le contact de l'enzyme avec la surface d'un adsorbant. En fonction du type de surface, l'immobilisation fait intervenir des forces physiques telles que des interactions ioniques, des liaisons hydrophobes, des liaisons de Van Der Waals ou encore une combinaison de ces forces (Kennedy et al., 1990).
La présence de liens faibles entre l'enzyme et le support induit peu de changements de conformation du biocatalyseur. De ce fait, l'activité initiale de l'enzyme est en grande partie conservée.
L'adsorption se fait sur des supports insolubles tels que du carbone poreux, de l'argile, du verre et des résines aromatiques polymériques.La procédure est basée sur : (i) un mélange de l'enzyme avec l'adsorbant adéquat sous les conditions appropriées de pH et de force ionique, (ii) temps de contact suffisant et (iii) lavage des enzymes légèrement liées ou non liées.
Cette méthode physique d'immobilisation perturbe beaucoup moins la structure tridimensionnelle de la protéine et offrent la rétention des propriétés enzymatiques ressemblant à celles de la molécule en solution (Zaborsky, 1974). En contrepartie, la stabilité de l'activité enzymatique est typiquement moindre que celle observée lorsque des liaisons
s'avère sensible aux fluctuations des conditions du milieu. En ce sens, des variations du pH ou de la conductivité du milieu peuvent induire le relargage des enzymes immobilisées par adsorption.
Insolubilisation par réticulation d'enzymes (sans support)
L'insolubilisation d'enzymes via la réticulation est une méthode attractive puisque la préparation finale résulte essentiellement en de la protéine avec une haute concentration en activité par unité de volume et une petite quantité d'agents réticulants (Mateo et al., 2004; Lôpez-Serrano et al., 2002). Ces biocatalyseurs solides sont préparés par réticulation directe des enzymes par des agents tel que le glutaraldehyde (Mateo et al., 2004).
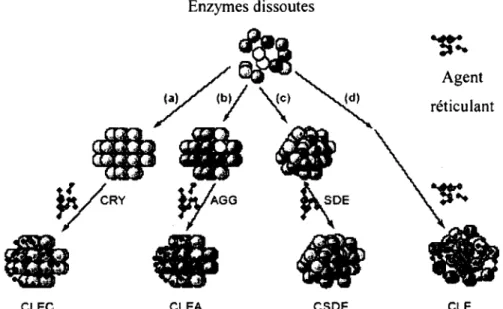
De façon générale, les enzymes en solution sont insolubilisées par différentes méthodes physico-chimiques (voir figure 2-9) à savoir par cristallisation (formation de CLEC), agrégation (formation de CLEA) et séchage (formation de CLSD). Ces enzymes sont par la suite réticulées de façon à conserver leur caractère insoluble. De plus, il est également possible de réticuler directement les enzymes en solution et ce, sans étape préalable de précipitation (formation de CLE).
Le mode d'immobilisation utilisé sans support influence grandement les différentes propriétés du biocatalyseur solide formé. De façon générale, ce type d'immobilisation augmente la stabilité thermique et au pH des enzymes. Il y a toutefois des différences au niveau de la rétention de l'activité enzymatique et de la stabilité mécanique des biocatalyseurs formés (Cao et al., 2003). Ainsi, la formation de CLEs a permis d'augmenter la stabilité thermique des enzymes, mais présentait un inconvénient en lien avec leur faible résistance mécanique, et le gel qu'elles forment qui rend difficile leur manipulation. De plus, les expériences étaient difficiles à répéter car les conditions d'opération faisaient beaucoup varier les résultats.
D'autre part, les cristaux d'enzymes réticulés (CLECs) ont montré une très grande stabilité lorsque soumis à des forces de cisaillement, des pH extrêmes et des températures élevées (Roy et Abraham, 2006). Cependant, la préparation de cristaux d'enzymes s'est avérée couteuse et laborieuse car elle requière une solution enzymatique d'une très haute pureté et un contrôle
Enzymes dissoutes
réticulant
CLEC CLEA CSDE CLE
Figure 2-9 : Les différentes approches pour la production d'enzymes immobilisées sans support : préparation par a) cristallisation (CLEC), b) agrégation (CLEA), c) séchoir atomiseur
(CSDE), d) réticulation directe (CLE). (Adaptée de Cao et al., (2003))
La formation de CLEA repose sur une technique d'immobilisation simple : la précipitation d'enzymes par l'ajout de différents précipitants (sels, surfactants, solvants organiques, etc.) est combinée à la réticulation (simultanée ou non) des enzymes présents (Mateo et al., 2004). Les catalyseurs ainsi formés présentent des caractéristiques semblables à celles des CLEC en termes de rétention de l'activité enzymatique et de la stabilité au pH, à la température et des propriétés mécaniques. Contrairement aux CLECs, la formation de CLEAs ne nécessite pas de cristallisation (Lopez-Serrano et al., 2002). En ce sens, la précipitation enzymatique est une technique simple qui est utilisée dans la majorité des protocoles de purification des enzymes. Ainsi, il est possible d'utiliser des solutions protéiques non purifiées et de récupérer des biocatalyseurs composés uniquement de protéines.
Globalement, la formation de CLEAs présente plusieurs attraits pour le développement d'applications industrielles car elle repose sur une technologie simple et facilement optimisable. Ceci se traduit par: 1) des coûts de développement faibles et une utilisation rapide; 2) une applicabilité sur de nombreuses enzymes, incluant des solutions enzymatiques non purifiées; 3) elle permet de produire des biocatalyseurs stables, réutilisables présentant des activités massiques élevées (Sheldon et al., 2005).
L'agent réticulant le plus utilisé pour former des CLEAs est le glutaraldéhyde (GLU) (Sheldon, 2007b). Cet agent bifonctionnel réagit avec le groupement amine des résidus lysine constituant l'enzyme pour former des liens amides. Les mécanismes chimiques impliqués ne sont pas très bien compris, et sous des contraintes mécaniques, les CLEAs pourraient relâcher du glutaraldéhyde dans le milieu réactif (Betancor et al, 2006), et de ce fait, il pourrait être associé à des problématiques environnementales. En ce sens, des études démontrent qu'il réduit le taux d'éclosion des oeufs de certaines espèces aquatiques (Leung, 2001; Raikow et al, 2007; Emmanuel et al, 2005), et qu'il aurait aussi certains effets chroniques sur des embryons de poissons et certains phytoplanctons (Sano et al, 2005). Il a été démontré que le GLU peut causer chez l'homme un asthme occupationnel, de l'eczéma et des irritations de la peau et des voies respiratoires (Takigawa et Endo, 2006). Son potentiel cancérigène est également évalué puisqu'il provoque des liaisons entre l'acide désoxyribonucléique (ADN) et les protéines de cellules mammifères (Speit et al, 2008). Devant ce constat, il apparait évident qu'il est recommandable de réduire l'impact environnemental qu'a la formation des CLEAs en trouvant une alternative au GLU.
Plusieurs alternatives au GLU sont possibles, on retrouve notamment le glyoxal, les hydrates de carbone fonctionnalisés, la génipine, et le chitosane. Ce dernier est un biopolymère dérivé de la chitine qui se distingue des autres car il possède plusieurs avantages en ce qui concerne l'immobilisation enzymatique. Comme c'est un polycation, cela facilite l'adsorption des enzymes sur le chitosane (Yang et al, 2006). Il est peu coûteux, abondant et son hydrophilicité facilite sa manipulation pour l'immobilisation d'enzymes hydrosolubles (D'Annibale et al, 1999). Le chitosane est également résistant à la dégradation par des microorganismes ce qui en fait un support de choix pour des enzymes destinées à traiter des milieux contenant une flore microbienne importante comme le sol ou les eaux usées (Yang et al, 2006). Il est également biocompatible et n'est pas considéré comme étant toxique autant pour l'environnement que pour la santé humaine (Void et Christensen, 2005).
D'autre part, plusieurs travaux ont démontré la possibilité d'immobiliser avec succès la laccase sur ce biopolymère (Cabana et al, 2011 ; Lu et al, 2007; Yang et al, 2006; Delanoy et al, 2005; Palmieri et al, 2005; D'Annibale et al, 1999). Ce faisant, le chitosane a été sélectionné comme agent réticulant pour la formation de la structure des EPES de CLEAs et