Université de Neuchâtel Institut de Zoologie
Station fédérale de Recherches en production végétale Changins, Nyon
Les tordeuses (Lepidoptera, Tortricidae) des vergers:
analyse uni- et multidimensionnelle de leur
distribution, des échanges avec les écosystèmes
adjacents et des processus de recolonisation
(Bassin lémanique)
Par
Philippe Jeanneret
Thèse présentée à la Faculté des sciences de l'Université de
¡Neuchâtel pour l'obtention du grade de docteur es sciences
Travail réalisé avec le soutien du Fonds national suisse de la
Les tordeuses (Lepidoptera, Torticidae) des
vergers: analyse uni-et mutidimensionnelle de
leur distribution, des échanges avec les
écosystèmes adjacents et des processus de
recolonisation (Bassin lémanique)
de M. Philippe Jeanneret
UNIVERSITE DE NEUCHATEL
FACULTÉ DES SCIENCES
La Faculté des sciences de l'Université de
Neuchâtel sur le rapport des membres du jury,
Mme M. Rahier (directrice de thèse), MM. J.-M. Gobât,
A. Buttler, P.-J. Charmillot (co-directeur de thèse, Nyon),
F. Burel (Rennes) et J. Casas (Tours)
autorise l'impression de la présente thèse.
Neuchâtel, Ie 1er avril 1998
Le doyen:
Table des matières
Table des matières
CHAPITRE 1 - Introduction1.1 Etat des connaissances
1.1.1 Généralités 1 1.1.2 Le mouvement et la dispersion 2
1.1.3 Le verger traditionnel et la culture d'arbres fruitiers en tant qu'écosystème
île 3 1.1.4 Les tordeuses (Lep. Tortricidae) dans les vergers et autres habitats . . . 4
1.1.5 La dispersion des tordeuses et les échanges avec les milieux adjacents 6
1.2 But du travail 7
CHAPITRE 2 - Les vergers traditionnels d'arbres à haute tige, les cultures intensives d'arbres à basse tige et les écosystèmes adjacents
2.1 Introduction 10 2.2 Les vergers traditionnels d'arbres à haute tige et les milieux adjacents
2.2.1 Situation générale 13 2.2.2 Les vergers traditionnels étudiés 14
2.3 Les cultures de pommiers et les milieux adjacents
2.3.1 Les cultures de pommiers étudiées 31
CHAPITRE 3 - Matériel et Méthodes
3.1 L'échantillonnage et l'élevage des chenilles 35 3.2 L'application d'un insecticide dans les vergers d'arbres à haute tige 36
3.3 L'interception des imagos avec le piège Malaise 37 3.4 L'identification des chenilles, des chrysalides et des imagos 39
CHAPITRE 4 - Les tordeuses phyilophages des vergers d'arbres à haute tige dans Ie Bassin lémanique
4.1 Introduction . . . " 45
4.2 Résumé des éléments importants de la biologie des tordeuses phyilophages et espèces
proches capturées 49 4.3 La densité des populations de tordeuses phyilophages et espèces apparentées dans les
vergers d'arbres à haute tige du Bassin lémanique 52 4.4 La distribution des tordeuses phyilophages et espèces apparentées dans les vergers:
interprétation selon l'analyse factorielle des correspondances 58 4.5 L'influence des descripteurs environnementaux sur la distribution des tordeuses
phyilophages et espèces apparentées: interprétation selon l'analyse canonique des correspondances
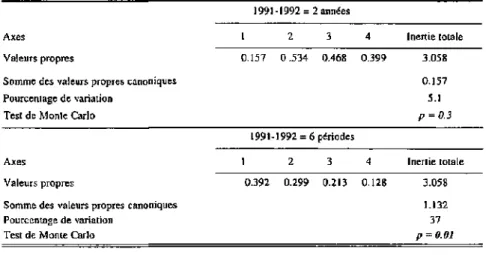
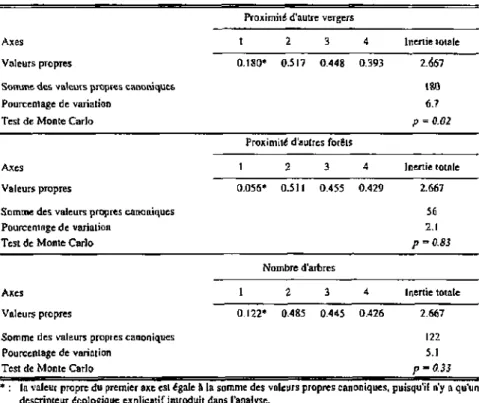
4.5.1 Introduction 63 4.5.2 L'effet temporel 64 4.5.3 L'effet spatial 74 4.5.4 L'effet des descripteurs paysagers et autres descripteurs environnementaux
sur les tordeuses et leur diversité 76 4.5.5 L'importance relative des descripteurs environnementaux et synthèse des
résultats 85
L'importance relative du temps et de l'espace décrite par une série
d'analyses canoniques partielles 86
4.6 Résumé du chapitre 4: Les tordeuses phyilophages des vergers d'arbres à haute tige
dans Ie Bassin lémanique 90
CHAPITRE S - L'activité de vol des tordeuses à l'interface des vergers/cultures et des écosystèmes adjacents
5.1 Résumé de la biologie des tordeuses capturées avec le piège Malaise 91 5.2 Le mouvement global à l'interface entre les vergers/cultures et les écosystèmes
adjacents
5.2.1 Analyse globale du peuplement de tordeuses capturées et comparaisons
entre les 2 années de piégeage 95 5.2.2 Analyse de la structure des mouvements et distribution dans les six vergers
Table des matières
5.3 Le mouvement différencié à l'interface entre les vergers/cultures et les écosystèmes adjacents
5.3.1 Introduction 105 5.3.2 L'activité du peuplement complet de tordeuses à l'interface des
vergers/cultures et des écosystèmes adjacents: interprétation selon l'analyse
factorielle des correspondances et le groupement 105 5.3.3 L'activité à l'interface des vergers/cultures et des écosystèmes adjacents de
la guilde des tordeuses inféodées: interprétation selon l'analyse factorielle
des correspondances et le groupement 119 5.4 L'influence des descripteurs environnementaux sur le mouvement des tordeuses à
l'interface entre les vergers/cultures et les écosystèmes adjacents
5.4.1 Introduction 127 5.4.2 Les descripteurs environnementaux 131
5.4.3 L'influence des descripteurs environnementaux sur le peuplement complet de tordeuses: interprétation selon l'analyse canonique des correspondances
Analyse canonique 1 (peuplement) 136 ACC descripteur par descripteur et sélection pas à pas des descripteurs
environnementaux multiclasses (peuplement) 139 Diagrammes d'ordination ACC, interprétation des axes et comparaison
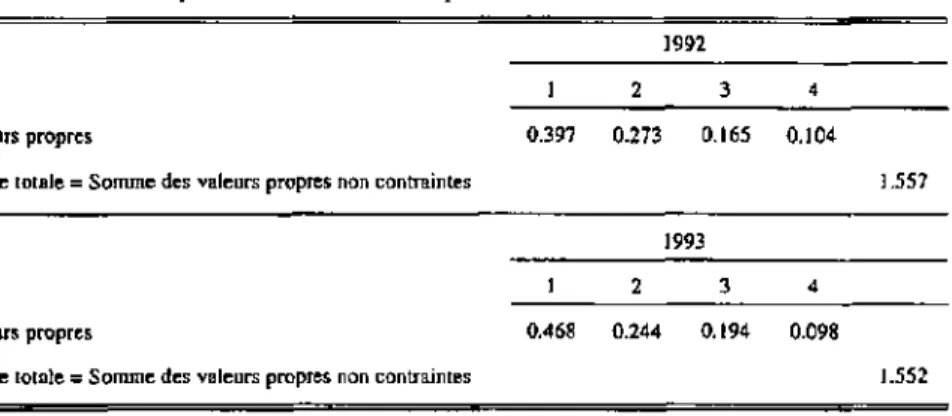
avec les résultats de VAFC (peuplement) 147 Relations espèces-environnement (peuplement) 161 Analyses canoniques partielles 1992 (peuplement) 170 Analyses canoniques partielles 1993 (peuplement) 181
5.4.4 L'influence des descripteurs environnementaux sur la guilde des tordeuses inféodées au pommier: interprétation selon l'analyse canonique des correspondances
Analyse canonique 1 (guilde) 189 ACC descripteur par descripteur et sélection pas à pas des descripteurs
environnementaux multiclasses (guilde) 190 Diagrammes d'ordination ACC, interprétation des axes et comparaison
avec les résultats de VAFC (guilde) 196 Relations espèces-environnement (guilde) 206 Analyses canoniques partielles 1992 (guilde) 211 Analyses canoniques partielles 1993 (guilde) 216
5.5 Résumé du chapitre 5: L'activité de vol des tordeuses à l'interface des vergers/cultures
CHAPITRE 6 - LE processus de recolonisation des vergers non cultivés d'arbres à haute tige après l'application d'un insecticide
6.1 Introduction 221 6.2 Analyse temporelle de l'effet d'un traitement insecticide sur la guilde des "chenilles
de printemps" 222 6.3 Analyse temporelle et multidimensionnelle de l'effet d'un traitement insecticide et
d'autres descripteurs environnementaux sur la recolonisation des vergers par la guilde des tordeuses phyllophages et espèces apparentées
6.3.1 Introduction 227 6.3.2 Analyse temporelle et multidimensionnelle de l'effet d'un traitement
insecticide sur la recolonisation des vergers par la guilde des tordeuses
phyllophages et espèces apparentées 228 6.3.3 Analyse temporelle et multidimensionnelle de l'effet des descripteurs
environnementaux sur la recolonisation des vergers par la guilde des
tordeuses phyllophages et espèces apparentées 233 6.4 Résumé du chapitre 6: Le processus de recolonisation des vergers non cultivés
d'arbres à haute tige après l'application d'un insecticide 239
CHAPITRE 7 - Discussion et Conclusions
7.1 Discussion 240 7.2 Conclusions 248
CHAPITRE 8 - Remerciements 250
CHAPITRE 9 - Résumé - Summary - Zusammenfassung
9.1 Résumé 252 9.2 Summary 254 9.3 Zusammenfassung 256
Table des matières
Annexe 1 : Captures des chenilles de tordeuses (Lep. Tortricidae) du feuillage en 1991
et 1992.
Annexe 2: Captures des tordeuses (Lep. Tortricidae) avec le piège Malaise (1992):
espèces associées au pommier.
Annexe 3: Captures des tordeuses (Lep. Tortricidae) avec le piège Malaise (1992):
espèces non associées au pommier.
Annexe 4: Captures des tordeuses (Lep. Tortricidae) avec le piège Malaise (1993):
espèces associées au pommier,
Annexe 5: Captures des tordeuses (Lep. Tortricidae) avec le piège Malaise (1993):
espèces non associées au pommier.
Annexe 6: Captures des chenilles de tordeuses (Lep. Tortricidae) du feuillage (1993 et
1994) dans 5 vergers non traités.
Annexe 7: Captures des chenilles de tordeuses (Lep. Tortricidae) du feuillage (1993 et
1994) dans 5 vergers traités.
CHAPITRE 1
Introduction
1.1 Etat des connaissances
1.1.1 G é n é r a l i t é s
La distribution des organismes vivants est hautement dépendante de leur capacité à se disperser et la façon dont ils se dispersent est un sujet de recherche important en écologie fondamentale et appliquée. La dispersion permet aux organismes de coloniser et de recoloniser les milieux. Ce rôle est prépondérant à l'échelle temporelle géologique, comme à l'échelle écologique, plus petite, et conduit aux processus d'évolution et de distribution d'abondance.
Les études menées sur les processus de colonisation utilisant les îles comme exemple montrent que le nombre d'espèces habitant une île atteint un équilibre, les extinctions compensant les immigrations (MAC ARTHUR & WILSON, 1967). Cette théorie appliquée aux interactions insectes-plantes a été proposée la première fois par JANZEN (1973) et considère les plantes hôtes comme des îles "dans une mer de végétation hostile". La distribution en tache des habitats est un facteur déterminant pour la dispersion des organismes et les îles "réelles" peuvent être comparées avec les "îles" existant sur le continent. Plusieurs auteurs ont tenté d'appliquer Ia théorie de la biogéographie des îles comme concept pour le contrôle de ravageurs dans les cultures. Mais ils ont souligné que les cultures n'étaient pas à l'équilibre (dans le sens biogéographique) et que la communauté y vivant était largement dominée par les migrations (REY & McCOY, 1979; PRICE, 1976; LISS et al, 1986; LEVINS & WILSON, 1980).
2 Introduction
1.1.2 Le m o u v e m e n t et la dispersion
PIELOU (1979) et KREBS (1985) ont synthétisé trois modes de dispersion des espèces:
- la diffusion, mouvement graduel qui se déroule sur plusieurs générations. - le "jump dispersal", mouvement sur de longues distances, suivi d'un
établissement réussi.
- la migration séculaire, dispersion à l'échelle de l'évolution.
A l'échelle locale, les barrières naturelles peuvent affecter ou même interdire la dispersion des espèces. Mais pour ANDREWARTHA & BIRCH (1954), les animaux ont une tendance innée à la dispersion.
Les entomologistes ont tenté de classer les mouvements dans différentes catégories en utilisant comme critères les causes et les moyens du déplacement. Dans son introduction, JOHNSON (1969) résume les concepts de migration et de dispersion. Particulièrement, la migration est définie comme un transfert des adultes d'un habitat vers d'autres habitats, ce qui est similaire à la dispersion adaptative et distincte de la dispersion accidentelle ou non intentionnelle. D'anciens concepts représentent la migration comme une tentative désespérée de fuir la pression de la population. Mais sa principale fonction est de permettre le déplacement d'un environnement défavorable vers des milieux plus propices (DINGLE, 1972 et 1989). SOUTHWOOD (1962) souligne que les migrations sont plus souvent observées de ou vers des milieux "temporaires" ou éphémères (à évolution rapide), par comparaison avec les écosystèmes "stables" et prévisibles (à évolution lente).
La distribution spatiale et temporelle des plantes est déterminante pour la colonisation et l'établissement des insectes phytophages, comme le mouvement et la dispersion. La distribution des plantes et leur diversité influence la densité des insectes phytophages, principalement parce qu'elles influencent le mouvement et le
comportement de recherche (KAREIVA, 1982 et 1990). Mais le problème réside dans la mesure et la quantification du mouvement (par ex. TURCHIN et ai, 1991; WIENS
et ai, 1993). Dans les agroécosystèmes pour lesquels Ia concentration des ressources
est très grande, le problème des ravageurs est aggravé (CROMARTIE, 1981; TONHASCA & BYRNE, 1994). Cette évidence a conduit plusieurs auteurs à soulever la possibilité de manipuler l'organisation spatiale des cultures pour contrôler les ravageurs (par ex. PERRIN & PHILIPS, 1978; PERRIN, 1980). RISCH et ai (1983), résumant les effets de Ia diversité sur la stabilité des agroécosystèmes, montre que la diversification botanique abaisse fréquemment les populations de ravageurs. On peut imputer ces observations à l'effet conjoint de la dilution des ressources et de la prédation. A plus large échelle, la distribution en tache1 d'une plante hôte ou de plusieurs plantes hôtes influence également directement la dynamique des populations d'insectes phytophages polyphages (GOULD & STINNER, 1984; C R I S T e / a l , 1992).
1.1.3 Le verger traditionnel et Ia culture d'arbres fruitiers en tant qu'écosystème île
Pour la culture d'arbres fruitiers2, CROFT & HULL (1983) ont développé une approche écologique de l'agroécosystème. Considérant le verger comme un écosystème (!), ils ont souligné six composantes agissant sur la diversité des arthropodes: (1) l'arbre (variété, architecture); (2) la strate herbacée; (3) le sol; (4) l'environnement du verger; (5) la phénologie de l'arbre; (6) la conduite de la culture (si cultivé).
1 par analogie avec le terme de "patch" utilisé par les anglo-saxons, nous employerons le terme de "tache" pour définir la surface occupée par une plante hôte.
2 les observations faites s'appliquent également aux vergers traditionnels. Le terme de
4 Introduction
Toujours selon CROFT & HULL (1983), la théorie de la biogéographie des îles peut aussi s'appliquer au verger. Dans ce cas, le degré d'immigration et d'émigration dépend de: (1) la diversité et la capacité à se disperser des espèces colonisatrices; (2) la distance entre le ou les écosystèmes sources et le verger; (3) la taille du verger. Les concepts généraux de la diversité des insectes phytophages à l'échelle locale développés par STRONG et al. (1984) peut s'appliquer à la faune des vergers. En particulier, la diversité locale est influencée par la distribution locale et l'abondance des plantes hôtes (taille des surfaces occupées, densité), l'isolation des taches (controversée en dépit de la théorie de la biogéographie des îles, SIMBERLOFF,
1978; REY, 1981; WILLIAMSON, 1981) et le mode de culture.
L'étude de la communauté complète d'arthropodes vivant dans le pommier a révélé l'importance conjointe de la conduite de la culture et de son environnement (BROWN1 1993; BROWN & WELKER, 1992; RATHMAN & BRUNNER, 1988). Notamment, la diversité des arthropodes du verger semble fortement dépendre des écosystèmes alentours (SZENTKIRÁLYI & KOZAR1 1991).
Dans les vergers suisses, l'approche systémique a essentiellement été développée pour les acariens ravageurs (ZAHNER, 1985; KLAY, 1987; GENINI, 1987; JUVARA-BALS, 1992), mais fait peu de cas de l'environnement du verger.
1.1.4 Les tordeuses (Lep. Tortricidae) dans les vergers et autres habitats
La famille des tordeuses contient de nombreuses espèces à la dynamique communément ou occasionnellement explosive et qui sont devenues des ravageurs connus sur Ia terre entière. Les plus fameuses espèces sont certainement Zeiraphera
diniana Gn. (la tordeuse grise du mélèze), Chorisioneura fumiferana Clem. (la
tordeuse du pin), Cydia pomonella L. (le carpocapse des pommes) et Tortrix viridana L. (Ia tordeuse verte du chêne).
tordeuses causant des dommages dans les vergers européens sont bien développées. La distribution, le voltinisme, la biologie, les plantes hôtes, les habitudes alimentaires (l'organe préféré: feuillage, fruit, bourgeon, fleur, écorce) sont en général bien connus (BALACHOWSKY 1966; BOVEY, 1979; CHAMBÓN, 1986; VAN DER GEEST & EVENHUIS, 1991).
Trois groupes de ravageurs sont définis pour la lutte intégrée dans les vergers (CROFT & HULL, 1983): (1) les ravageurs clefs (espèces persistantes, perennes); (2) les ravageurs sporadiques ou occasionnels; (3) les ravageurs secondaires (ravageurs sporadiques qui ne se nourrissent pas du fruit). Malgré leur dynamique parfois explosive, les tordeuses sont plutôt des stratèges K que r, mais des habitats aussi différents que des cultures fruitières intensives et des arbres "sauvages" isolés peuvent causés des changements dans le statut r-K (CROFT & HULL, 1983).
STRICKLER & WHALON (1985), étudiant la composition faunistique des microlépidoptères dans 17 vergers de pommiers, ont trouvé que le groupe d'espèces occupant les cultures était un sous-groupe de celui des vergers abandonnés. Rejoignant WHALON & CROF7T (1984), ils expliquent que les espèces les plus largement distribuées sont aussi celles qui se dispersent le plus facilement avec une haute probabilité de colonisation réussie.
Dans les cultures européennes, la diversité des tordeuses varie à l'échelle biogéographique, régionale et locale. Les groupes d'espèces causant des dégâts sont souvent similaires, mais les densités de population peuvent différer. Excepté le carpocapse des pommes qui est un ravageur clef partout, le statut de ravageur des autres tordeuses varie selon l'espèce, la région et l'année. Dans une revue sur la question, AUDEMARD (1986) considère 10 espèces comme ravageurs clefs et 8 espèces comme ravageurs sporadiques, mais souligne que 1 à 3 espèces seulement sont des ravageurs locaux. Ces ravageurs sont essentiellement liés au pommier puisque parmi 27 espèces communément piégées dans les cultures fruitières (pommes, poires, pêches et prunes), 17 créent des dégâts dans les cultures de
6 Introduction
pommiers. Nous préciserons que les méthodes de capture utilisées (pièges appâtés de phéromones sexuelles, observation visuelle des chenilles) limitent ie nombre des espèces aux tordeuses inféodées au pommier. En Suisse, dans un verger de pruniers, GIGON (1978) a capturé pas moins de 51 espèces de tordeuses avec un piège Malaise, et en Hongrie, MESZAROS et al. (1984) ont recensé 88 espèces dans un verger de pommiers en utilisant différentes méthodes de piégeage. Nous précisons que le terme "inféodé" est utilisé dans cette étude alors même que les tordeuses ne sont pas exclusivement dépendantes du pommier mais notoirement polyphages.
1.1.5 La dispersion des tordeuses et les échanges avec les milieux adjacents
La dispersion des ravageurs a été étudiée pour les espèces économiquement importantes. Le carpocapse des pommes se déplace généralement à courte distance (de l'ordre de 100 à 200m), à l'intérieur du verger et dans les milieux adjacents (WORTHLEY, 1932; COUTIN, 1959; WILDBOLZ & BAGGIOUNI, 1959; NAGY & JERMY, 1972; HOWELL & CLIFF, 1974; CAUSSE & FERRON1 1979). Mais des expériences de lâché-recapture ont permis de mesurer des distances parcourues beaucoup plus grandes (STEINER, 1940), allant jusqu'à 11 km (MANI & WILDBOLZ, 1977)1. Les adultes de la principale tordeuse de la pelure2, Adoxophyes
orana F.v.R., se disperse essentiellement à courte distance (approx. 100 m) mais
BAREL (1973) a montré qu'un déplacement des chenilles au bout de leur fil était possible, d'arbre en arbre en tout cas.
En Europe, une carte de distribution établie par FASSOTTE et al. (1986) pour
Clepsis spectrana Tr. et Pandemis heparana Den.&Schiff. montre que les deux
il s'agit en !'occurence d'un individu mâle recapturé à l'aide d'un piège appâté d'attractif sexuel.
appelée ainsi car l'espèce est phyllophage mais les chenilles mangent également la pelure des fruits.
espèces occupent aussi les environs du verger. En Australie, les travaux sur Epiphyas
postvittana Walk, ont démontré l'importance des écosystèmes adjacents comme
source possible de réinfestation (THOMAS & SHAW, 1982), malgré que les populations extérieures aux cultures soient confinées dans de petites poches de plantes hôtes favorables (GEIER & BRIESE, 1980). L'importance des milieux adjacents aux vergers a aussi été révélée par ALTIERI & SCHMIDT (1986), soulignant ses effets sur la colonisation par les insectes (malheureusement sans les lépidoptères): ayant capturé les insectes avec un piège Malaise placé à l'interface du verger et de l'écosystème adjacent, cette étude révèle des échanges importants entre le verger commercial classique et son environnement forestier, mais que par contre Ie mouvement global entre un verger abandonné et la forêt est faible. ALTIERI & SCHMIDT (1986) concluent que plus la différence structurale entre le verger et Ie milieu adjacent est grande plus les échanges sont importants. Nous soulignerons, sans vouloir diminuer en rien la valeur de l'étude, que les conclusions sont tirées de l'examen d'une seule interface par catégorie (i interface verger commercial - forêt; I interface verger organique - forêt; I interface verger abandonné - forêt).
1.2 But du travail
Dans ce travail, nous avons étudié Ia distribution régionale des tordeuses habitant les vergers traditionnels non cultivés de La Côte, ainsi que les mouvements entre les vergers, les cultures, et les milieux qui leur sont adjacents. Nous nous sommes également intéressés aux processus de recolonisation des vergers traditionnels après l'application d'un traitement insecticide, afin de mieux définir l'échelle temporelle du phénomène. Les trois thèmes développés dans cette étude ont pour but principal de mieux définir, pour les tordeuses, le rôle paysager de l'écosystème verger.
8 Introduction
sexuelle, vims de la granulose, régulateurs de croissance et inhibiteurs de chitine) contre les ravageurs les plus importants, le carpocapse des pommes et Ia tordeuse de Ia pelure, d'autres tordeuses augmentent leur pression dans les cultures fruitières. Ce récent changement provient sans nu! doute du fait que ces espèces étaient par le passé éliminées par l'application d'insecticides à très large spectre. D'un point de vue agronomique, il est intéressant d'étudier la composition en espèces des vergers traditionnels pour évaluer leur éventuel rôle comme source d'infestation pour les cultures. L'échelle temporelle de la recolonisation d'un verger traditionnel après un traitement est également importante agronomiquement puisqu'elle permettra de définir si par exemple le traitement annuel des cultures avant la fleur est justifié pour les tordeuses (même si ce traitement est plus ou moins obligatoire à cause d'autres ravageurs comme les noctuelles et les géométrides).
D'un point de vue écologique, il est intéressant d'étudier la composition faunistique des vergers traditionnels et surtout de tester quantitativement l'influence des facteurs environnementaux caractérisant le verger sur la distribution des tordeuses. La mesure du taux d'échanges entre le verger et son environnement, ainsi que l'influence de cet environnement sur le taux d'échange, permet de mieux définir le degré "d'ouverture" de l'écosystème au paysage alentour. Le suivi de la recolonisation après traitement, mis en relation avec les facteurs environnementaux rend également possible l'analyse de l'influence paysagère globale sur le phénomène. Les trois questions ou thèmes suivants sont abordés dans cette étude et sont directement liées aux chapitres de résultats:
(a) Quelle est la distribution régionale des tordeuses phyllophagcs inféodées au pommier dans les vergers traditionnels de !a région de Nyon - La Côte (Bassin lémaniqtie) et quelle est l'influence des descripteurs environnementaux caractéristiques de ces vergers et en particulier l'entourage du verger, sur cette distribution (chapitre 4) ? (b) Quelles est l'intensité des immigrations et émigrations des tordeuses
phyllophages entre les vergers et les milieux adjacents, et en particulier, cette intensité dépend-elle des milieux adjacents (chapitre 5) ? (c) A quelle échelle temporelle la recolonisation des vergers traditionnels
après un traitement insecticide s'effectue-t-elle et quelle est l'influence des milieux adjacents sur cette recolonisation (chapitre 7) ?
IO Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
CHAPITRE 2
Les vergers traditionnels d'arbres à haute tige, les
cultures intensives d'arbres à basse tige et les
écosystèmes adjacents
2.1 Introduction
Le paysage du bassin lémanique dans la région de Nyon - La Côte est fait de petites villes (Nyon: 12000 ha.), de nombreux villages, de cultures de céréales, de cultures fruitières, de vergers traditionnels non cultivés, de vignes et d'écosystèmes naturels (forêts, haies, etc.). A l'échelle régionale, la distribution des ressources potentielles pour les tordeuses est par conséquent caractérisée par une structure en tache.
Dans Ie bassin lémanique, nous pouvons considérer trois différents types de milieux favorables aux tordeuses inféodées au pommier:
(a) Les cultures intensives de pommiers (b) Les vergers traditionnels
(c) Les écosystèmes naturels contenant des plantes hôtes favorables
Nous avons concentré notre étude sur deux types de milieux abritant les tordeuses, les cultures de pommiers (a) et les vergers traditionnels (b). Les écosystèmes naturels n'ont pas été étudiés en soi. Selon ZWYGART (1983), nous parlerons donc de "verger" pour une plantation d'arbres à haute tige non traitée et de "culture" pour les autres. La composante commune aux vergers et aux cultures est la présence du pommier, Pyrus
de l'utilisation qu'on en fait.
Les trois différences de structure qui nous viennent d'emblée à l'esprit sont : Io ia taille des arbres de chacun de ces deux milieux; 2° la prairie sous-jacente sur laquelle les arbres sont plantés; et 3° la densité des arbres.
Dans le cas du verger, les arbres à haute tige atteignent facilement 5 m avec un tronc d'environ 1.8 m et une couronne très large. Dans la culture de pommiers, les arbres atteignent 3.5 m au plus, en fin de "vie commerciale", soit vers l'âge de 15-20 ans. Mais les cultures ont majoritairement des arbres plus jeunes donc plus petits. Le tronc des pommiers cultivés est très court, au plus 0,5m et leur couronne est généralement étroite1. Dans les vergers traditionnels, les arbres sont plantés sur une prairie de fauche ou un pâturage, plus ou moins intensivement exploités. En général, l'agriculteur met le bétail au printemps, pratique une fauche en été et remet éventuellement le bétail en automne ou fait une deuxième fauche. Dans les cultures d'arbres fruitiers, la prairie devient gazon par la régularité et le rythme soutenu des fauches (toutes les 2 semaines). Le nombre d'arbres par unité de surface peut avoisiner 100 arbres par hectare dans un verger traditionnel, mais ce chiffre est très rarement atteint (HORNER1 1988). Dans les cultures, la densité peut atteindre 8000 arbres par hectare (voir plus) en très haute densité, mais chez nous il se situe plutôt entre 2000 et 4000 (MONNEY & BLASER, 1993).
Les vergers traditionnels et les cultures étudiés sont distribués dans la région de La Côte le long du lac Léman sur une longeur de 25 km et une largeur d'environ 9 km, du bord du lac au pied du Jura (Fig. 1, reproduit avec l'autorisation de l'office fédéral de la topographie du 29.10.1998). Le verger le plus au sud est celui de Châtaigneraie, le verger le plus à l'ouest celui de La Rippe, le verger le plus à l'est et le plus au nord celui de St Livres.
La forme est dépendante du mode de taille mais généralement les arbres des cultures sont maintenus dans un plan.
12 Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
Fig. I. Carte de distribution des vergers traditionnels et cultures de pommier étudiés dans la région de La Côte. Numéros 1 à 14: 14 vergers d'arbres à haute tige (descriptions p. 16-30). Cl à C3: 3 vergers commerciaux (descriptions p. 32-34).
'!V
A
¿tÂ
'P
l ' i * VKi
?~Y-8&H'Jí-^" / /J'y*
-.Ay '.'2.2 Les vergers traditionnels d'arbres à haute tige et les milieux
adjacents
2.2.1 Situation générale
La surface couverte par les vergers traditionnels en Suisse est en constante diminution depuis 1960. C'est la conséquence, au départ (en 1970}, des campagnes d'abattage subventionnées par les pouvoirs publics, car on voulait remplacer les vergers d'arbres à haute tige par des cultures d'arbres à basse tige (rentabilité pour la production de fruits). Au cours de ces 40 dernières années, plus du trois quarts des arbres fruitiers à haute tige ont disparu en Suisse; leur nombre est ainsi passé de 20 mio. à 5 mio (MÜLLER, 1988)'.
Après les campagnes d'abattage, le facteur le plus important causant la disparition des vergers traditionnels est l'élargissement inconsidéré des surfaces construites. En effet, les vergers traditionnels se trouvant majoritairement aux abords immédiats des fermes et des villages, dans ce qu'on appelle aujourd'hui les zones à bâtir, leur disparition était et est encore programmée à courte et moyenne échéance. Les mesures prises depuis 1994 et qui tentent d'enrayer les influences néfastes de l'agriculture à l'égard de l'environnement (production biologique, production intégrée, surfaces de compensation écologique) ne peuvent qu'être bénéfiques aux vergers traditionnels, puisqu$e ceux-ci rentrent dans le cadre d'indemnisations pour l'agriculteur (surface de compensation écologique type 8 selon l'ordonnance sur les contributions écologiques). Pourtant, le mode actuel de subventionnement permettra peut-être d'arrêter l'hémorragie mais ne suffira pas à la création nouvelle de vergers traditionnels2.
Le rôle écologique de l'écosystème verger traditionnel n'est pourtant plus à
1 On peut estimer que depuis lors (1988), on en a encore perdu 1 ou 2 mio.
2 Comment voulez-vous résister quand on vous propose plus de 600.-Sfr. Ie m2 pour voire verger en zone à bâtir, alors que la confédération vous donne 15.-/arbre + éventuellement 3000.-/ha pour la prairie pour le garder à titre de surface de compensation écologique ?!!
14 Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
démontrer. En effet, le verger offre une place intermédiaire entre la forêt et les prés ouverts, dépourvus d'éléments de structure (BLAB11986; MÜLLER et al., 1988). L'offre structurelle du verger traditionnel permet l'enrichissement spécifique à l'échelle d'un paysage, en particulier pour les oiseaux (BROGGI & SCHLEGEL, 1990; LUDER, 1981 ). Comme espèces typiques, on cite en particulier la chouette chevêche, la huppe, le torcol fourmilier et la pie-grièche à tête rousse (SERVICE DE PROTECTION DES OISEAUX,
1983).
2.2.2 Les vergers traditionnels étudiés
Les vergers traditionnels dans lesquels nous avons travaillé correspondent aux descriptions faites dans le sous-chapitre précédent. Après avoir examiné la composition botanique de la prairie sous-jacente de tous les vergers, nous avons pu conclure à une variation faible et compte tenu du sujet de notre étude, nous donnerons une liste générale des espèces rencontrées le plus fréquemment. Les éventuelles informations complémentaires sont données dans la description de chaque verger s'il y a Heu. et notamment en ce qui concerne les environs immédiats du verger.
Tab. I. Composition botanique des prairies sous-jacentes (inventaire non exhaustif).
Achilca Millefolium L- Lolium perenne L. Agrostis alba L. Medicago saliva L. Arrhenaterum elati us L. Plantago major L. Bellis perennis L. Planiamo media L. Capsela Bursa-pas tori s L. Prunella vulgaris L. Carduus sp- Ranunculus accr L. Cynosurus cristatus L. Rumex Acetosa L. Dactylis glomerata L. Rumex sanguineus L. Gallium molugo L. Taraxacum officinale L-Geranium roiundifolium L. Trifolium pratense L. Heracleum sphondylium L. Trifolium repens L. Holcus lanatus L. Trisetum flavescens L. Lolium multiflorum Lam. Urtica dioica L.
La description des environs de chaque verger se divise en trois parties ou zones. La première fait référence aux abords immédiats du verger, soit Ie milieu directement adjacent. Cette zone est décrite dans la première ligne du tableau de la description des vergers. Elle sera reprise comme descripteur écologique paysager et nommée paysage 1 dans les analyses. La deuxième ligne du tableau considère le milieu observé à une distance de 50 à 100m du verger. Les milieux plus éloignés (dès 100m) sont décrit dans la troisième ligne. Les descriptions contenues dans les deuxième et troisième lignes forment le descripteur écologique paysager paysage II utilisé dans les analyses.
Tous les extraits de cartes sont reproduit avec l'autorisation de l'office fédéral de la topographie du 29.10 1998.
16 Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
Verger 1: Genolier (VD)
Coordonnées géographiques: 506 280/143 400 Pommiers: 22
Autres essences: cerisiers
Exploitation: bétail au printemps, 1 fauche en été Environnement du verger:
Altitude: 550 m Superficie: env. 0.8 ha Pente: légère, exp. Est
Nord Est Sud Ouest
prairie de fauche type sous- prairie de jacentc fauche type
sous-jacente
route, habitation, jardin complexe potager avec ¡tubus scolaire, gazon spp.
bosquet (Acer platartoides, cordon boisé de idem
Fraxinus excelsior, Sorbus 20m de large aucuparia, Rubus sp., (type bosquet Cornus mas, Aegopodium Nord) et rivière podagraria
village
village idem verger mi-tige
intensif
prrq Verger traditionnel étudié I v.- ) Autres vergers traditionnels
Forêt
I I Grandes cultures, prés à fourrage et pâturages E Vigne
Verger 2: Malesseri (VD)
Coordonnées géographiques: 516 820/147 380 Pommiers: 13
Autres essences: cerisiers, poiriers Exploitation: moutons toute l'année Environnement du verger:
Altitude: 440 m Superficie: env. 1 ha Pente:
-Nord Est Sud Ouest
haie de Crataegus culture de pommiers,
oxyacantha L. poiriers vigne
prairie artificielle, fauchée,
engraissée
vigne cordon boisé de 20 m de autoroute, vigne
large {Acer pseudopìatanus, grande culture
Fagus sylvatica, Crataegus monogyna, Rosa spp., Rubus fruticosus)
vigne vigne culture d'arbres vigne
fruitiers
t'.v I Verger traditionnel étudié I v.* I Autres vergers traditionnels I ••••-• I Cultures fruitières
I 1 Grandes cultures, prés à fourrage et pâturages
Mn
vig"e
- i i i_
18 Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
Verger 3 : Céligny
Coordonnées géographiques: 504 700/133 620 Pommiers: !6
Autres essences: cerisiers, poiriers Exploitation: bétail toute l'année Environnement du verger:
Altitude: 390 m Superficie: env. 0.7 ha Pente:
-Autre: taillé régulièrement Nord-Ouest route, voie ferroviaire bosquet (Acer platanoides, Fraxinus excelsior, Sorbits aucuparia) village Nord-Est cordon boisé de 20m de large (Acer pseudoplatanus,
Fraxinus excelsior, Ulmus campestris, Crataegus monogyna, Corylus avellana) villas, jardins grandes cultures Sud-Est prairie artificielle, fauchée, engraissée prairie artificielle, fauchée, engraissée villas, jardins Sud-Ouest ferme grande culture grandes cultures
I vWl Verger traditionnel étudié WW Forêt
I I Grandes cultures, prés à fourrage et pâturages Ü ^ i Lac Léman
Verger 4: Trélex (VD)
Coordonnées géographiques: 505 200/141 60 Pommiers: 15
Autres essences: cerisiers, poiriers
Exploitation: bétail au printemps, 1 fauche en été Environnement du verger: Altitude: 500 m Superficie: env. 0.6 ha Pente: -Nord village, jardins village Est jardins, villas, autre verger villas Sud voie ferroviaire grande culture Ouest village autres vergers
village villas, grandes
cultures grandes cultures prairies artificielles fauchées et engraissée
¿¿Oír * X 3
í b u é/r in • . *'-.*V''-•'-•II ^kS '' / / '•''•$:•:•' 'il:-¿*.. £ a | f t « u w w o j "¿TX I ^
;1'.-."I Verger traditionnel étudié l-v I Autres vergers traditionnels
1 I Grandes cultures, prés à fourrage et pâturages EMD Vigne
N
20 Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
Verger 5: Petit Bois (VD) Coordonnées géographiques: Pommiers: 28
503 250/135 600 Autres essences: cerisiers, poiriers, cognassiers Exploitation: bétail au printemps, 1 fauche en été Environnement du verger:
Nord-Ouest
forêt (Quercus robur, Acer
campestre, Carpinus betulus, Crataegus
monogyna et C. oxyacantha, Corylus avellana. Rosa sp., Prunus spinosa, Corylus avellana. Cornus mas, Fagus sylvatica
Ligustrum vulgare. Lonicera xylosteum, Cornus sanguineum, Rubus spp., Euphorbia amygdaloïdes, Nord-Est idem Nord-Ouest mais Crataegus monogyna et C. oxyacantha, Rubus spp., Rosa sp., Prunus spinosa forme la lisière
forêt idem Nord-Ouest Sud-Est habitation prairie pâturée, fauchée Altitude: 470 m Superficie: env. 1.1 ha Pente: -Sud-Ouest haie (1.5m de large, 1.2m de haut) avec Crataegus monogyna et C. oxyacantha, Rubus spp., Rosa sp., Prunus spinosa Corylus avellana, Cornus mas, prairie pâturée, fauchée, cultures de cerisiers mi-tige
grandes cultures forÊt idem
Nord-Ouest forêt idem Nord-Ouest et grandes cultures grandes cultures
I y :-., I Verger traditionnel étudié I---' I Vergers cultivés Forêt I I Grandes cultures, prés à fourrage et pâturages
IMD
V iê
n e 100 500 1000 m22 Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
Verger 6: Givrins (VD)
Coordonnées géographiques: 505 40/142 620 Pommiers: 13
Autres essences: cerisiers
Exploitation: bétail au printemps, 1 fauche en été Environnementdu verger:
Altitude: 560 m Superficie: env. 0.4 ha Pente:
-Nord-Ouest Nord-Est Sud-Est Sud-Ouest
village, jardins centre scolaire voie ferroviaire village, jardins avec pommiers, pruniers, noyers, poiriers village, jardins village, jardins avec villas, jardins
poiriers, pruniers, noyers, pommiers, cerisiers
village, jardins
village, jardins grandes cultures grandes coteau avec verger (30 cultures, culture pommiers, qq. poiriers, de pommiers cerisiers, noyers,
cordon boisé, rivière
I v./ I Verger traditionnel étudié I •-••• l Autres vergers traditionnels
Forêt
I I Grandes cultures, prés à fourrage et pâturages
Verger 7: St Livres (VD)
Coordonnées géographiques: 519 500/151 350 Pommiers: 10
Autres essences: cerisiers, poiriers
Exploitation: bétail au printemps, 1 fauche en été Environnement du verger:
Altitude: 600 m Superficie: env. 0.5 ha Pente:
-Nord-Ouest Nord-Est Sud-Est Sud-Ouest
village, jardins cordon boisé de 20m de large
(Fraxinus excelsior, Crataegus monogyna, Corylus avellana, Rubus sp., Tilia platyphyllos, A cer pseudoplatanus, Ligustrum vulgare, Rosa spp.,)
prairie pâturée, fauchée prairie pâturée, fauchée village, jardins, autres vergers
grande culture idem grande
culture grandes cultures grandes cultures et verger de
cerisiers
grandes cultures, village, prairie artificielle, jardins engraissée, fauchée
l-v.'l Verger traditionnel étudié I'---'I Autres vergers traditionnels
Forêt
1 I Grandes cultures, prés à fourrage et pâturages [ Ü 3 Vigne
24 Les vergers traditionnels d'arbres à haute tige, ies cultures intensives ...
Verger 8: Au bonne (VD)
Coordonnées géographiques: 518 750/150 350 Pommiers: 20
Autres essences: cerisiers, poiriers Exploitation: bétail toute l'année Environnement du verger:
Altitude: 540 m Superficie: env. 1 ha
Pente: moyenne, exp. Nord-Est
Nord-Ouest Nord-Est Sud-Est Sud-Ouest
grande culture grande culture
et prairie pâturée fauchée prairie pâturée, fauchée prairie pâturée, fauchée, verger de pruniers, cerisiers, habitation grande culture et prairie pâturée, grande culture prairie pâturée, prairie pâturée, fauchée fauchée fauchée forêt (Fagus sylvatica, Fraxinus
excelsior, Crataegus monogyna
et C. oxyacantha, Corylus
avellana, Acerpseudoplatanus, Ligustrum vulgare, Rosasp., Prunus spinosa, Cornus mas)
idem forêt Nord-Ouest idem forêt Nord-Ouest idem forêt Nord-Ouest
ivsK
äK
^
I T A ^ V ^ ^ ^ È
f^^-XME^Äv^r^l^vY*^r-vœt J^M¿,
• | / ^ V > S S J ! S & * Ä : •''
T\V^A¿^^'
^s^^^^
wvJviSiiftrBBJ/lrs—^> JL ~jE!u^±sfiflaftuiÊ!vaBSPPai
P R Ï B
ríP$£CT M'A?
t •.••• I Verger traditionnel I *•'•* I Autres vergers traditionnels ItfMW ForêtI I Grandes cultures, prés à fourrage et pâturages
EMJ Vigne
N
Verger 9: Gingins 1 (VD)
Coordonnées géographiques: 502 980/140 280 Pommiers: 8
Autres essences: cerisiers, poiriers
Exploitation: bétail au printemps, 1 fauche en été Environnement du verger: Altitude: 540 m Superficie: env. 0.2 ha
Pente:
-Nord-Ouest village, jardins prairie pâturée, fauchée Nord-Est prairie pâturée, fauchée village, jardins Sud-Est prairie pâturée, fauchée grande culture Sud-Ouest prairie pâturée, fauchée village, jardins grandes cultures, petite forêt (idem Nord-Ouest Petit-Bois village, jardins, autres vergersgrandes cultures village, jardins, autres vergers
I v.*.I Vergers traditionnels étudiés I '•'•• I Autres vergers traditionnels
I I Grandes cultures, prés à fourrage et pâturages (D Verger 9
© Verger 10
26 Les vergers traditionnels d'arbres à haute tige, les cultures intensives...
Verger 10: Gingins 2 (VD)
Coordonnées géographiques: 503 00/140 280 Altitude: 540 m Pommiers: 8 Superficie: env. 0.2 ha Autres essences: cerisiers, poiriers Pente:
-Exploitation: bétail au printemps, 1 fauche en été Environnement du verger: Nord-Ouest prairie pâturée, fauchée grande culture Nord-Est
village, jardins, autres vergers grande culture Sud-Est village, jardins village, jardins Sud-Ouest
village, jardins, autres vergers
grande culture, petite forêt (idem Nord-Ouest Petit-Bois grandes cultures grandes cultures et grandes grandes cultures
prairies pâturées et cultures fauchées
Verger 11: La Rippe (VD)
Coordonnées géographiques: 501 190/137 380
Pommiers: 10
Autres essences: cerisiers
Exploitation: bétail toute l'année
Altitude: 510 m
Superficie: env. 0.3 ha
Pente:
-Autre: taillé régulièrement
Environnement du Nord-Ouest village, jardins, autres vergers idem verger: Nord-Est village, jardins, autres vergers idem Sud-Est grande culture idem Sud-Ouest prairie pâturée, fauchée, autre verger grande culture et prairie pâturée fauchée grandes cultures grandes cultures et petite forêt (idem idem
et prairies prairies pâturées, Nord-Ouest Petit pâturées, fauchées fauchées Bois)
|:,:.:| Verger traditionnel étudié I •-••• I Autres vergers traditionnels
\ I Grandes cultures, prés
à fourrage et pâturages
28 Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
Verger 12: Coinsins (VD)
Coordonnées géographiques: 507 580/142 100 Pommiers: 16
Autres essences: cerisiers, poiriers, pruniers, cognassiers Exploitation: bétail toute Tannée
Environnement du verger:
Altitude: 480 m Superficie: env. 0.2 ha Pente:
-Autre: taillé régulièrement Nord-Ouest village, jardins, autres vergers idem grandes cultures et prairies pâturées, fauchées Nord-Est grande culture et prairie pâturée fauchée idem forêt (Bois de Chênes) Sud-Est village, jardins vigne idem Sud-Ouest village, jardins grande culture et prairie pâturée fauchée idem
I v:,:| Verger traditionnel étudié [v.* I Autres vergers traditionnels ü § Forêt
I 1 Grandes cultures et pâturages
\mi Vigne
Verger 13: Châtaigneraie (VD)
Coordonnées géographiques: 502 280/132 380 Pommiers: 15
Autres essences: poiriers Exploitation: bétail toute l'année Environnement du verger: Altitude: 450 m Superficie: env. 0.5 ha Pente: -Nord village, jardins culture d'arbres fruitiers, vigne Est village, jardins vigne Sud village, jardins vigne Ouest village, jardins grande culture et prairies pâturées, fauchées grandes cultures grandes cultures et idem
et prairies prairies pâturées, pâturées, fauchées fauchées
vigne
ipa- \'.'.'\ Verger traditionnel étudié
I '•'•* I Autres vergers traditionnels ^ H Vergers cultivés
I I Grandes cultures, prés à fourrage et pâturages OU] Vigne
30 Les vergers traditionnels d'arbres à haute tige, les cultures intensives ...
Verger 14: Grens (VD) Coordonnées géographiques: Pommiers: 7
504 380/139 460 Autres essences: cerisiers, poiriers
Altitude: 495 m Superficie:
Pente:
-Exploitation: bétail au printemps, 1 fauche en été Environnement du verger:
Nord-Ouest grande culture
grande culture et prairie pâturée, fauchée forêt (Quercus robur,
Fraxinus excelsior, Crataegus monogyna et C. oxyacantha, Corylus avellana, Acer pseudoplaianus, Ligustrum
vulgare. Rosa sp., Prunus spinosa. Cornus mas)
Nord-Est grande culture grande culture culture d'arbrcs fruitiers Sud-Est grande culture grande culture grandes cultures, cultures d'arbres fruitiers env. 0.1 ha Sud-Ouest grande culture grande culture village, jgrdins, autres vergers, cultures d'arbres fruitiers _l I 1_
| v . ) Verger traditionnel étudié |v.' I Vergers cultivés Forêt [ I Grandes cultures, prés à fourrage et pâturages l-frvyj Vigne 100 500 1000 m
2.3 Les cultures de pommiers et les milieux adjacents
2.3.1 Les cultures de pommiers étudiées
Les trois cultures de pommiers étudiées sont conduites selon les principes de la production intégrée, avec notamment l'application de techniques de lutte contre les ravageurs le moins néfastes possible pour l'environnement. En particulier, la technique de la confusion sexuelle est appliquée contre le carpocapse des pommes dans les cultures de Begnins et Allaman, alors que la cultures de Crassier est protégée contre ce ravageur par l'application du virus de la granulose. Les tordeuses phyllophages, considérées avec les noctuelles et les géométrides comme "chenilles de printemps" sont éliminées avec un régulateur de croissance ou un inhibiteur de chitine au printemps avant l'apparition de la fleur.
32 Les vergers traditionnels d'arbres à haute lige, les cultures intensives ...
Culture de pommiers 1: Allaman (VD) Coordonnées géographiques: 519 00/146 600 Pommiers: env. 11500
Autres essences:
Exploitation: production intégrée Environnement du verger:
Altitude: 375 m Superficie: env. 5.7 ha Pente;
-Nord-Ouest Nord-Est Sud-Est Sud-Ouest
route, habitation
grande culture forêt (Robinia
Pseudo-Acacia, Fagus sylvatica, Tilia piatyphyllos, Sambucus nigra, Acer pseudoplatanus, Jugions regia, Quercus robur)
cultures de pommiers, habitations et forêt idem Sud Est
vigne idem lac Léman lac Léman
idem villas, jardins lac Léman lac Léman
Culture étudiée
I '•'•' I Autres cultures fruitières Forêt I I Grandes cultures, prés à fourrage et pâturages Lac
EM3
v"gne
Verger de kiwi 100 500 1000 mCulture de pommiers 2: Crassier (VD)
Coordonnées géographiques: 502 600/136 800 Pommiers: env. 4600 '
Autres essences:
Exploitation: production intégrée Environnement du verger:
Altitude: 470 m Superficie: env. 2.3 ha Pente:
-Nord-Ouest Nord-Est Sud-Est Sud-Ouest
vigne grande culture,
pépinière de pommiers
route, habitation, prairie pâturée, fauchée autre verger, forêt (Quercus
robur, Fraxinus excelsior, Fagus sylvatica, Tilia platyphyllos, Crataegus monogyna et oxyacanlha, Acer platanoïdes et campestre, Ligustrum vulgare, Corylus avellana, Viburnum ¡anfana
grand cultures grande culture
grande culture
grandes cultures villas, jardins cordon boisé village (idem
Nord-Ouest)
I v / I Culture étudiée
\ •.'•' I Autres cultures fruitières
B l Vergers traditionnels
I H Forêt
I I Grandes cultures, prés à fourrage et pâturages B Vigne 100 500 1000 m34 Les vergers traditionnels d'arbres à haute tige, les cultures intensives...
Culture de pommiers 3: Begnins (VD) Coordonnées géographiques: 502 600/136 800 Pommiers: env. 16000
Autres essences:
Exploitation: production intégrée Environnement du verger:
Altitude: 470 m Superficie: env. 8 ha Pente:
-Nord-Ouest Nord-Est Sud-Est Sud-Ouest
habitation cordon boisé (Fraxinus vigne
excelsior, Robinia Pseudo-Acacia, Rubus spp., Urtica dioica grande culture vigne grandes cultures, vigne grande culture grandes cultures et cultures fruitières grande culture forêt (Quercus robur,
Robinia Pseudo-Acacia, Tilia platyphyllos, Sambucas nigra, Acer pseudoplatanus)
vigne cultures fruitières
FvTl Culture étudiée
I '•'•' I Autres cultures fruitières Forêt
Uu0 I I Grandes cultures, prés à fourrage
et pâturages
FÏÏ^Î Vigne
Jardins avec arbres à haute tige
i — i — • ' •
CHAPITRE 3
Matériel et Méthodes
3.1 L'échantillonnage et l'élevage des chenilles
Nous avons échantillonné les chenilles des tordeuses phyllophages dans les vergers de pommiers non traités de la région de Nyon - La Côte selon la même méthode en 1991, 1992, 1993 et 1994. En 1991 et 1992, années d'étude de la distribution des espèces (Chapitre 4), 11 et 8 vergers respectivement ont été échantillonnés (Tab. 2). En 1993 et 1994, années d'étude du processus de recolonisation (Chapitre 7), nous avons échantillonné les chenilles dans 10 vergers.
Nous avons défini le bouquet foliaire (ensemble des feuilles issues d'un même bourgeon) ou floral (ensemble des fleurs issues d'un même bourgeon) comme unité d'échantillonnage et 150 unités, choisies au hasard, ont été examinées par arbre en 1991. En 1992 et les années suivantes, nous avons augmenté le nombre d'unités échantillonnées par arbre à 200. Le nombre d'arbres échantillonnés par verger est proportionnel au nombre total d'arbres du verger: 1-5 arbres, 1 arbre échantillonné; 6-10, 2 arbres échantillonnés; 11-15, 3 arbres échantillonnés; 16-20, 4 arbres échantillonnés; 21-25, 5 arbres échantillonnés. Nous avons effectué l'échantillonnage à 4 reprises en 1991 et 1992, à 6 reprises en 1993 et 2 reprises en 1994.
Les chenilles obtenues ont été déterminées provisoirement et mises individuellement en élevage dans des cellules climatisées à 250C, 75% d'humidité relative et 18 heures de lumière. En 1991, nous avons nourri les chenilles tout au long de leur développement avec du feuillage provenant de vergers non cultivés. Nous avons néanmoins remarqué que le feuillage séchait ou pourrissait trop rapidement et, en 1992 et les années suivantes, les chenilles ont été nourries avec du
36
Matériel et Méthodes
milieu artificiel. La détermination des imagos éclos confirme ou infirme
l'identification des chenilles.
Tab. 2. Résumé des échantillonnages de bouquets foliaires et floraux effectués en 1991 et
1992 pour l'étude de la distribution (chapitre 4) et, en 1993 et 1994, pour l'étude de la
recolonisation après un traitement insecticide (chapitre 7).
Vergers Genolier Malesseri Ceïigny Trélex Peiii Bois Givrins St Livres Aubonne Gingins 1 La Rippe Coinsins Gingins2 Châtaigneraie Grens Total Nombre d'arbres 22 13 16 16 28 13 10 20 8 8 16 8 15 7 200 Nombre d'arbres Échantillonnés 5 3 4 3 5 3 3 5 2 3 4 2 3 2 47
Nombre de bouquets échantillonnés
Distribution 1991' 3000 1800 2400 1800 1800 1200 1800 2400 1200 1800 1200 20400 1992' 4000 2400 3200 2400 4000 2400 2400 4000 24800 Recolonisat 19931 6000 3600 4800 3600 6000 3600 3600 6000 3600 2400 43200 ¡on 1994' 2000 1200 1600 1200 2000 1200 1200 2000 1200 800 14400 4 Échantillonnages: fin avril, mi-mai, fin mai, mi-juin.
6 échantillonnages: fin avril, mi-mai, fin mai. mi-juin, début septembre début octobre. 2 échantillonnages: début mai et fin mai.
3.2 L'application d'un insecticide dans les vergers d'arbres à
haute tige
Dans le but d'étudier le processus de recolonisation du verger d'arbres à haute tige
par les tordeuses, nous avons échantillonnés les chenilles dans 10 vergers de la
région de Nyon-La Côte à la fin du mois d'avril 1993, puis nous avons appliqué un
insecticide dans 5 des 10 vergers. La recolonisation a ensuite été observée par échantillonnage des chenilles tout au long de l'année 1993, du début du mois de mai à la mi-septembre, puis au printemps 1994 par 2 échantillonnages au mois de mai. P o u r les vergers de pommiers, le p r o g r a m m e de lutte intégrée élahoré par les scientifiques de Changins, et périodiquement publié dans la Revue suisse de
viticulture, d'arboriculture et d'horticulture, conseille une application au printemps
avant l'apparition des fleurs pour lutter contre les "chenilles de printemps"'. Le produit utilisé, le Zolone®, fait partie de la gamme des insecticides polyvalents traditionnellement appliqués en arboriculture fruitière. L'action de cet insecticide est directe (contact) et indirecte (ingestion). Nous avons répandu le produit à l'aide d'un pistolet gicleur couramment appelé "gun". L'efficacité à 100% d'un traitement dans les vergers d'arbres à haute tige peut être mis en doute, car le feuillage important et dense, caractéristique des pommiers de ces vergers, ne peut être atteint dans son intégralité par la méthode utilisée. L'efficacité est également dépendante du groupe d'insectes étudié puisque les chenilles des espèces se déplaçant beaucoup augmentent leur risque d'ingérer du feuillage traité.
3.3 L'interception des imagos avec le piège Malaise
Pour la capture des imagos actifs à l'interface entre le verger et les milieux adjacents, nous avons utilisé un piège Malaise (dit "tente Malaise") transformé. Le modèle sur lequel nous nous sommes basé est décrit par TOWNES2 (1972). Le piège a 3m de haut et 2m de long (Fig. 2). Il intercepte les insectes entre Im et 2m du sol (surface d'interception lmx2m, soit 2m1) ce qui est légèrement en dessous de la
Cette expression, fréquemment utilisée par les professionnels de l'arboriculture désigne l'ensemble des tordeuses, géométrides, noctuelles, géléchides, argyrestides, piutellides, etc.
38 Matériel et Méthodes
hauteur de la couronne d'un pommier. Le tissu tendu entre les piquets d'aluminium qui forment la structure du piège est fait de mailles de 1mm1. Nous avons placé 2 collecteurs au sommet de la structure pour séparer les individus emigrants des individus immigrants. Nous avons donc attribué au piège une interception bidirectionnelle. Chaque collecteur contient 250 ml d'alcool à 75°. Le piège est Fixé au sol à l'aide de cordelettes et de sardines.
Fig. 2. Plan du piège Malaise bidirectionnel.
Collecteur double 1.60m Toit (blanc) \ Surface d'interception (noir) Armature en aluminium Nous avons placé un piège bidirectionnel Malaise sur chacun des 4 côtés de 6 vergers de pommiers, en respectant si possible l'orientation selon les 4 points cardinaux. Le collecteur représente l'unité d'échantillonnage. En 1992, nous avons collecté 48 unités (4x6x2) durant 14 semaines successives, du début du mois de
juillet au début du mois d'octobre {soit 672 unités). En 1993, les pièges ont été disposés du début du mois de juin au début du mois de septembre et ont été relevés chaque semaine durant 15 semaines (soit 720 unités).
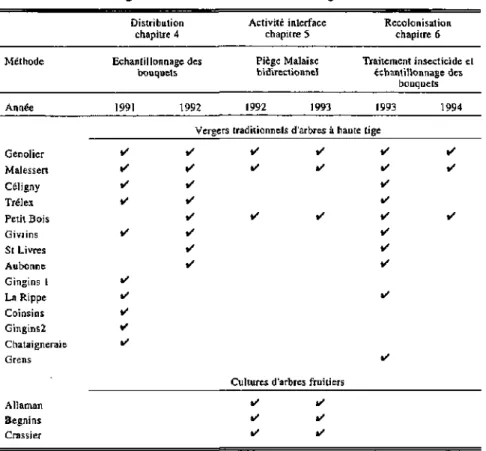
Tab. 3. Résumé des vergers étudiés et des méthodes d'investigations.
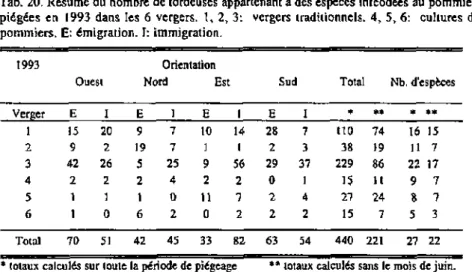
Méthode Année Distribution chapitre 4 Echantillonnage des bouquets 1991 1992 Activité interface chapitre 5 Piège Malaise bidirectionnel 1992 1993 Recolonisation chapitre 6 Traitement insecticide et échantillonnage des bouquets 1993 1994 Genolîer Malesseri Céligny Trélex Petit Bois Givrins St Livres Aubonne Gingins I La Rippe Coinsins Gingins2 Châtaigneraie Grens • • • • • i / • • • • Allaman Begnins Crassier
3.4 L'identification des chenilles, des chrysalides et des imagos
L'identification des chenilles est peu sûre en particulier lorsque celles-ci sont aux premiers stades de leur développement. Les ouvrages de détermination font le
Vergers traditionnels d'arbres à haute tige
• • • • • • • • • •
• • • • •
Cultures d'arbres fruitiers
40 Matériel et Méthodes
plus souvent référence aux derniers stades (SWATSCHEK, 1958; BEEKE & DE JONG, 1991; CHAMBÓN, 1986). Seuls GUILBOT & GOUJET (1978) proposent une clef d'identification des jeunes stades, mais reconnaissent que la fiabilité des déterminations est faible. Il est donc impératif d'élever les chenilles capturées, afin de les conduire le plus loin possible dans leur développement. L'identification des chrysalides est possible et relativement simple pour un petit nombre d'espèces fréquemment rencontrées dans les vergers comme ravageurs (BEEKE & DE JONG,
1991). Néanmoins, la détermination de l'imago reste le moyen d'identification le plus sûr. Les couleurs et la structure du dessin des ailes permettent d'identifier les espèces sans trop de difficultés (HANNEMAN, 1961; GRAAF BENTINK & DIAKONOFF, 1968; KUZNETSOV, 1989). Il est néanmoins indispensable de pouvoir comparer les individus à une collection de référence (en l'occurrence, nous avons utilisé la collection de M. HÄCHLER, collaborateur à Changins).
Dans notre cas, les imagos élevés avaient souvent perdu une partie ou la totalité des écailles recouvrant leurs ailes, car sitôt éclos ils s'activaient et cherchaient à sortir de leur boîte d'élevage. Les imagos capturés par le piège Malaise ne sont pas dans un très bon état car dès leur interception, leurs ailes frottent contre le tissu et les bord du collecteur avant qu'ils ne tombent dans l'alcool. Par conséquent, nous avons procédé à la détermination par l'extraction des pièces génitales, et à leur éclaircissage dans un bain chaud (9O0C) de KOH.
L'ouvrage le plus complet pour l'identification des genitalia est celui de KUZNETSOV (1989) mais il est également nécessaire d'utiliser HANNEMAN (1961) et GRAAF BENTINK & DIAKONOFF (1968). L'identification des genitalia est généralement applicable aux imagos mâles car peu de travaux ont été accompli sur les genitalia femelles. Néanmoins, nous avons déterminé les femelles grâce à l'ouvrage de GRAAF BENTINK & DIAKONOFF (1968), qui comporte un assortiment très complet de photos, ainsi que les dessins présentés par KUZNETSOV
(1989)
1. Il est clair que nous avons le pius souvent utilisé le recoupement des
observations faites sur les ailes et les genitalia.
3.5 Les techniques d'analyse et traitements statistiques
Les trois chapitres de résultats sont traités sur le même schéma d'analyse. La première partie considère les totaux d'individus et d'espèces capturés. Les techniques appliquées sont du domaine de la statistique classique unidimensionnelle. L'essentiel de l'information est obtenu à partir des moyennes et des écart-types (statistique descriptive). Dans cette partie, le traitement statistique inférentiel des données utilise le test U de Wilcoxon-Mann-Whitney ou le test de Wilcoxon pour échantillons appariés (SCHERRER, 1984).
La deuxième partie des analyses, beaucoup plus conséquente dans cette étude, considère simultanément l'ensemble des espèces et leurs abondances. Nous parlons alors d'analyse multivariable et de statistique multiâimensionnelle.
La bibliographie de base2 que nous avons utilisée fait référence aux ouvrages suivant (par ordre alphabétique):
BORCARD, D., LEGENDRE, P. & DRAPEAU, P. (1992). BORCARD, D. & BUTTLER, A. (1993).
GAUCH, H.G. (1982).
JONGMAN, R.H.G., TER BRAAK, C.J.F. & VAN TONGEREN, O.F.R. (1987).
LEGENDRE, L. & LEGENDRE P. (1984a).
LEGENDRE, L. & LEGENDRE
1P. (1984b).
LEGENDRE, P. & VAUDOR, A. (1991).
' Dans cet ouvrage, les genitalia femelles ne sont pas présentées pour toutes les espèces .
42 Matériel et Méthodes
TER BRAAK
1, C.J.F. (1986).
TER BRAAK, C.J.F. (1987a).
TER BRAAK, CJ.F. (1987b).
TER BRAAK, C.J.F. (1990a).
• TERBRAAK, C.J.F. (1991).
Les applications pratiques des analyses statistiques utilisées sont intimement liées à l'utilisation des logiciels qui ont été développés sur la base des théories mathématiques. Pour les analyses unidimensionnelles, noua avons utilisé le logiciel SYSTAT® (Course Technology, Cambridge) et l'ouvrage s'y référant (BERK, 1994). Pour les analyses multidimensionnelles, nous avons employé les logiciels suivants: - CANOCO® (Microcomputer Power, Ithaca) et les ouvrages de TER BRAAK
(1986, 1987a, 1987b, 1990a, 1991).
- PROGICIEL "R" (Université de Montréal) et l'ouvrage de LEGENDRE & VAUDOR (1991).
Trois phases caractérisent cette deuxième partie de notre analyse:
1. Phase exploratoire: diagrammes d'ordination obtenus grâce à l'analyse
factorielle des correspondances (AFC), et superposition des résultats d'un groupement. Dans notre étude, les ordinations donnent une excellente
image des similarités et dissimilarités entre les objets, mais nous utiliserons également les groupements pour confirmer certaines structures observées et pour affiner les liens qui existent entre ces objets2.
2. Phase d'interprétation des données, en trois étapes:
analyse indirecte dans laquelle on tente d'expliquer a posteriori les
1 11 faut préciser que cet article fait intégralement partie de fa thèse de TER BRAAK (1987a). 3 Cette méthode est largement recommandée par la littérature spécialisée. Elle permet
structures de la phase exploratoire obtenues par une AFC à l'aide de facteurs explicatifs (descripteurs environnementaux). Techniquement, nous avons utilisé les coefficients de corrélations intrasets (corrélations entre les facteurs explicatifs et les axes de l'ordination, définis selon TER BRAAK, 1986, p. 1170), ainsi que les coefficients de régression multiple des coordonnées des sites (ou relevés) sur les facteurs explicatifs (la régression est calculée après l'extraction des coordonnées des espèces et des sites de l'AFC).
analyse directe dans laquelle les structures sont dégagées compte tenu des facteurs explicatifs introduit a priori. Dans notre étude, il s'agit de
l'analyse canonique des correspondances (ACC). Nous utilisons alors les coefficient de corrélations intrasets et les coefficients canoniques
de régression multiple des axes environnementaux (coordonnées des sites calculés comme combinaison linéaire des facteurs explicatifs) sur les données.
partition de variance dans lequel on extrait la part explicative de
chacun des facteurs explicatifs introduits. Celte analyse est réalisée par une série à'ACCpartielles.
3. Tests d'hypothèses: tests statistiques permettant de vérifier l'hypothèse d'une action significative des facteurs explicatifs sur la structure des données. La procédure utilisée est un test par permutations dit de Monte Carlo (HOPE, 1968).
Les données brutes ont été transformées par la fonction y' = ln(y+l). Cette transformation est typiquement adaptée aux données d'abondance d'espèces. En effet, ce type de données est généralement composé de nombreuses espèces peu représentées et de quelques espèces abondantes sur l'ensemble des stations. Un telle répartition ne correspond pas à une distribution normale. La transformation logaritmique nous permet de remédier à cette particularité et, par son effet,
44
Matériel et Méthodes
d'amenuiser !Impact sur l'analyse des quelques espèces très abondantes (LEGENDRE
et LEGENDRE, 1984a, p. 18-19; JONGMAN et al., 1987, p. 103). Les auteurs cités
ci-dessus conseillent de ne pas intégrer dans l'analyse les espèces avec trop peu
d'individus. Dans notre étude, nous avons fixer la limite à 5.
Nous avons parfois été confrontés au problème d'effet d'arche (GAUCH, 1982)
ou d'effet Guttman (BENZÉCR1, 1973). Dans ce cas, nous avons utilisé la forme
détendencée de l'analyse factorielle des correspondances ou DCA (Detrended
Correspondence Analysis). Nous avons eu recour au "détendançage" par fonction
polynomiale, conseillé si les effets de bord sont faibles (TER BRAAK, 1987a;
BORCARD & BUTLER, 1993).
CHAPITRE 4
Les tordeuses phyllophages des vergers d'arbres à
haute tige dans le Bassin lémanique
4.1 Introduction
La relation entre le nombre d'espèces phytophages la surface occupée par une plante hôte donnée, ainsi que la notion de distribution en tache de cette plante hôte dont on dérive l'idée d'isolation, font partie d'un ensemble plus large de considérations sur l'équilibre écologique des communautés, l'extinction des espèces, la colonisation des milieux et le rapport émigration-immigration. Cet ensemble, mis en évidence par MAC ARTHUR & WILSON (1967)', a été nommé "la théorie de la biogéographie des îles" ("The theory of island biogeography"). Les concepts de base, fondés sur quelques exemples précis tirés d'observations sur les oiseaux, ont été abondamment confrontés à d'autres exemples. L'adaptation au cas des insectes phytophages vivant au dépend de "plantes hôtes îles" dans une "mer" d'autre végétation a été proposée la première fois par JANZEN (1968 et 1973).
Sur la base des travaux de PRICE (1976), et l'appui de la théorie de JANZEN (1968 et 1973), REY & McCOY (1979) explorent la validité du modèle de la biogéographie des îles pour le cas des ravageurs de la culture du soja. Ces auteurs concluent que la démarche est intéressante mais spéculative à cause notamment du manque de connaissance sur la biologie des espèces et leur capacité à se disperser. A la même période, une contreverse oppose d'ailleurs les défenseurs de la théorie (DIAMOND, 1975; DIAMOND & MAY, 1976) à ses détracteurs (SIMBERLOFF &