FACULTÉ DES SCIENCES
QUELQUES ASPECTS
DE DYNAMIQUE DES POPULATIONS
DE BIOLOGIE GÉNÉRALE
ET DE BIOMÉTRIE
DU GARDON
(RUTILUS RUTILUS L.)
DANS 4 LACS
DU PLATEAU SUISSE
THESE
présentée à la faculté des Sciences
de l'Université de Neuchâtel
pour obtenir le grade
de docteur es Sciences
par
B L A I S E Z A U G G
Licencié es Sciences
OU GARDON (Rutilus r u t i l u s L.) DANS 4 LACS DU PLATEAU SUISSE
1 9 8 7
Par Blaise Zaugg
Institut de Zoologie
Faculté des S c i e n c e s
Université de Neuchâtel
Chemin de Chanteme rie 22
CH-2000 N E U C H A T E L 7
Quelques..aspects...de dynamique.J.e.s...popu]a,.tipns,
de b i o l o g i e générale e t de b i o m é t r i e du
gardon ( R u t i l u s r u t i l u s L.) dans quatre lacs
du Plateau suisse
de Monsieur.Bl a i se ZAUGG
UNIVERSITÉ DE NEUCHATEL
FACULTÉ DES SCIENCES
La Faculté des sciences de l'Université de Neuchâtel,
sur le rapport des membres du jury,
Messi eu.rs..,J
r-c. ....Pie^droll^^^^^^^Mermpd^^et
R. M ü l l e r (Kastanienbaum)
autorise l'impression de la présente thèse.
Neuchâtel, le ?...Mçembre....1987
Le doy_en:
T A B L E D E S M A T I E R E S * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * *
I.
I N T R O D U C T I O N l i . . 1 . . 2 . . 3 . I . 1 . 1 . 1 . 2 . 1 . 3 . 1 . 4 . 1 . 5 . 1 . 6 . IV. I V . 1 . I V . 2 . I V . 2 . 1 . I V . 2 . 2 . I V . 2 . 2 . 1 D E S C R I P T I O N G E N E R A L E D U G A R D O N SITUATION TAXONOHIQUEGENERALITES SUR LA BIOLOGIE
D I S T R I B U T I O N D E S C R I P T I O N D E S M I L I E U X GENERALITES LAC DE NEÙCHATEL LAC DE BIENNE LAC DU LOCLAT
LAC DES QUATRE-CANTONS
.CANAL DE LA T H I E L L E B I O M E T R I E I N T R O D U C T I O N M E T H O D E S M e s u r e s C a r a c t è r e s d é n o m b r a b l e s D e n t s p h a r y n g i e n n e s
IV.2.2.2.2.
IV.2.2,3.
IV.3.
IV.3.1.
IV.3.2.
IV.3.3.
IV.3.3.1 .
IV.3.3.2.
IV.3.4.
IV,3.5.
IV.3.6.
IV.4.
IV.5.
IV.5.1.
IV.5.2.
IV.5.2.1.
IV.5.2.2.
IV.5.2.3.
IV.6.
Transversa L
Décomptes des rayons des nageoires
RESULTATS GARDONS
Relations "longueur totale/longueur du corps'
Relations "Longueur totale/poids"
Décomptes des écailles
Longi tudina L
Transversal
Décomptes des' rayons des nageoires
Dents pharyngiennes
Résumé des comparai sons entre les caractères
bi omet riques
DISCUSSION GARDONS
RESULTATS HYBRIDES
Proportiond'hybrides
Types d"hybri des
Lac de Neuchâtel
Lac de Bienne
Lac dû Loclat
DISCUSSION HYBRIDES
V . V . 1 . V . 2 . P O P U L A T I O N INTRODUCTION LA PECHE DU GARDONV . 2 . 3 . 1 . S i t u a t i o n en Suisse
V . 2 . 3 . 2 . S i t u a t i o n d e s lacs é t u d i é s
V . 2 . 3 . 3 . Détail d e s c a p t u r e s
V . 3 . A N A L Y S E D'UNE SERIE DE P A R A M E T R E S DES
P O P U L A T I O N S
V . 3 . 1 . I n t r o d u c t i o n
V . 3 . 2 . M é t h o d e s
V . 3 . 2 . 1 . E c h a n t i l l o n n a g e
V . 3 . 2 .
2 .
M e s u r e s
V . 3 . 2 . 3 . D é t e r m i n a t i o n s d ' â g e V . 3 . 2 . 4 . C r o i s s a r i c e V . 4 . RESULTATSV . 4 . 1 . A b o n d a n c e et b i o m a s s e
V . 4 . 2 . M a t u r i t é s e x u e l l e
V . 4 . 3 . S e x - r a t i o
V . 4 . 4 . S t r u c t u r e d e s p o p u l a t i o n s en p l a c e
V . 4 . 4 . 1 . Lac de N e u c h S t e L
V . 4 . 4 . 2 . Lac de B i e n n e
V . 4 . 4 . 3 . Canal de la T h i e l l e
V . 4 . 4 . 4 . Lac du Loclat
V . 4 . 4 . 5 . . Lac d e s Q u a t r e - C a n t o n s
V . 4 . 5 . Croi s s a n c e
V . 4 . 5 . 1 . R e l a t i o n s " â g é / longueur t o t a l e " basées
sur Les r é t r o c a l c u l a t i o n s
V,
li.
5 . 1 . 1 . Lac de N e u c h S t e L
V. 4 . 5 . 1 . 2 . Lac de B i e n n e
V, 4. 5 . 1 . 3 . Lac des Q u a t r e ^ C a n t o n s
V . 4 . 5 .
2 .
R e l a t i o n s "âge
I
longueur t o t a l e " basées
sur les r é s u l t a t s bruts
V . 5 .
D I S C U S S I O N
VI.
S Y N T H E S E E T C O N C L U S I O N
V I I . R E S U M E / Z U S A M M E N F A S S U N G /
S U M M A R Y
V i l i .
B I B L I O G R A P H I E
A N N E X E S
R E M E R C I E M E N T S
P a r m i s Les plus anciens Vertébrés de notre planète, se
trouvent les cyclostomes et tes poissons. Ces derniers
peuplent depuis des millions d'années Les eaux les plus
diverses et constituent des stocks de nourri ture
consi dérab les.
Cette abondance ainsi que les qualités nutritives des
poissons ont depuis des temps immémoriaux incité la
convoi ti se des hommes qui ont déve loppé des méthodes de
capture d'une très grande diversité.
Si jusqu'au début du XXème siècle, on pouvait considérer
La pêche comme une activité artisanale, où L'action de
l 'homme sur les populations de poissons était modérée du
.point de. vue biologi que; actuel lenient, le perfect ion ne ment
des engins', de pêche et la mécanisation permettent aux
pêcheurs professionnels d'effectuer une exploitation
intense, où le nombre d"espèces comestibles mises à
con-tribution est étonnamment faible. En effet, les rares
poissons indi gènes que les Suisses daignent consommer,
sont principalement constitués par les corégones
CCoregonus s p ) , les truites (Sa Imo trutta ssp L . ) , Les
ombles chevaliers CSa l ve L i nus a Lpinus L . ) , les perches
(Perca
i
luviati lis L.) et les brochets (Esox Lucius L.î.
Cette intensification des prélèvements, jumelée à la
pollution croissante des milieux est à La base de nombreux
problèmes occupant Largement les chercheurs,
particu-lièrement Les ichtyologues, dont l'intérêt principal est
par La force des choses voué aux poissons dit "nobles".
Dans l'optique de favoriser ces peuplements et surtout de
freiner la prolifération des espèces peu ou pas
exploi-tées, dont l'expansion est de surcroît favorisée par
l'eutrophisation des lacs, Les autorités compétentes en
matière de pêche ont encouragé L'élimination des poissons
dits " i n d é s i r a b l e s " , c o n s t i t u é s en m a j o r i t é par des C y p r i
-n i d é s , par L ' i -n t r o d u c t i o -n r é c e -n t e de s u b v e -n t i o -n s v e r s é e s
aux p ê c h e u r s p r o f e s s i o n n e l s Lors de La c a p t u r e de ces
e s p è c e s .
L ' o c c a s i o n nous a semblé o p p o r t u n e a la r é a l i s a t i o n d ' u n e
é t u d e touchant à d i f f é r e n t s a s p e c t s de (a b i o l o g i e g é n é
-r a l e , a v e c un a c c e n t p a -r t i c u l i e -r p o -r t é su-r La d y n a m i q u e d e
p o p u l a t i o n , de L'espèce la p l u s t o u c h é e par ces c a m p a g n e s
d ' é l i m i n a t i o n s , Le g a r d o n CRuti Lus ruti Lus L . ) , dont Les
p e u p l e m e n t s p a s s e n t d a n s c e r t a i n s L a c s , d'un statut p e u
ou pas p é c h é à une s i t u a t i o n d ' i n t e n s e e x p l o i t a t i o n .
N o u s e x a m i n o n s donc en p r e m i e r Lieu q u e l q u e s a s p e c t s
s y s t é m a t i q u e s et b i o m é t r i q u e s de c e t t e e s p è c e ; en s e c o n d
Lieu, n o t r e a t t e n t i o n se p o r t e sur l ' é v o l u t i o n de la p ê c h e
du g a r d o n en S u i s s e et sur les é v e n t u e l l e s v a r i a t i o n s
q u ' e l l e o c c a s i o n n e à une s é r i e d e p a r a m è t r e s de leurs
p o p u l a t i o n s . N o u s t e r m i n o n s par une mise en garde à p r o p o s
de L'Impact des c a m p a g n e s d'éLimi n a t i o n s sur c e r t a i n e s
e s p è c e s dont les s t a t u t s sont p r é c a i r e s .
Nous nous contentons ici de donner une description sommaire du gardon.
Figure 1 : Gardon
II.1. SITUATION TAXONOMIOUE Selon HOLCIK & A_L_.C1971) et STEITZ & A L . ( 1 9 8 4 ) . Embranchement Sous-Embrancheraent Super-Classe Classe Sous-C lasse Super-Ordre Ordre Sous-Ordre Famille Genre Espèce CORDES VERTEBRES GNATHOSTOMES OSTEICHTHVENS ACTINOPTERYGIENS TELEOSTEENS CYPRINIFORMES CYPRINOIDEA CYPRINIDES RUTILUS (RAFINESQUE, 1820) R. RUTILUS (LINNAEUS, 1758)
Synonymies CSPILLMANN, 1961) : Cyprinus rutilus CL., 1758)
i
Cyprinus jaculus CJUR., 1825)
I
Leusciscus prasinus
(AG, 1835)
I
Leusciscus selysi (HECKEL, 1842)
I
Leusciscus
ruti loïdes CSEL. LONGCH., 1842)
I
Leusciscus p a U e n s
(BLANCH., 1866) / Leusciscus ruti Lus (GUENTHER, 1868).
Noms français : gardon, gardon commun, gardon blanc.
Noms vernaci! lai res : vengeron, vangeron, blanchet, rousse,
rosse, roche, f ago, francai s, echat out, etc..
II.2. GENERALITES SUR LA BIOLOGIE OU GARDON
Poisson de lacs ou de rivières à courant lent, son
exigence en oxygène est assez modeste. On le trouve
également dans les eaux saumâtres près de t'embouchure des
grands fLeuves.
PEDROLI 8 AL., (en préparation) définissent son habitat en
Suisse principalement en plaine, dans Les Lacs de toutes
tailles, ainsi que dans les cours d'eau d'une Largeur
minimale d'environ 10 m (sauf exceptions).
D'instinct assez grégaire, les gardons se déplacent en
bancs épars près des rives, ils regagnent souvent la
"profondeur" en hi ver,
La péri ode du frai se situe au printemps, Lorsque la
température de l'eau avoisine 10 'C, soit aux environs du
mois d'avril. Les feme Lies déposent alors leurs ovules
(selon nos mesures par volumetrie : 7350 ovules/IOOgr de
femelle, écart-type = 1320, nombre d'individus = 1 3 ) , d'un
diamètre approximatif de 1 a 1,5 mm, sur la végétation
aquatique à faible profondeur. Les ma Les, portant des
tubercules nuptiaux en forme de petits cônes, fécondent
alors ces ovules, qui après une incubation de 7 à 15 jours
se Ion La température, donnent naissance aux alevins qui
demeurent quelques temps cachés dans La végétation
aquatique, puis se réunissent en bancs immenses pour
longer Les côtes. CFATIO, 1882; DOTTRENS, 1951; SPILLMANN,
1961; QUARTIER, 1980).
Le gardon est omnivore, son régime alimentaire passe,
selon Les conditions, les saisons ou l'âge du poisson, du
phytoplancton aux macrophytes, du zooplancton au
macrobenthos, sans oublier tous les détritus et particules
susceptibles d'être assimilés. (LEFEVRE, 1940; CHEVALIER
S AL., 1974; BUESSER S M.., 1987; BRABAND, 1985; HAMMER,
1985).
II.3. DISTRIBUTION DU GARDON
Sa répartition très vaste (figure 2) va de l'Angleterre à
l'Ouest, ô la Rivière Lena à L'Est; et de la mer de Kara
au Nord, jusqu'aux affluents des mers Caspienne et d'Aral
dans le Sud. WHEELER(1969) donne comme limite altitudinale
maxima le, 914 m.
Pour l'Europe, dans l'ensemble,
on
ne trouve pas cette
espèce au-delà des Pyrénées et des Alpes, ni en Ecosse et
Norvège, ni dans les rivières qui se jettent dans La Baie
de Mourmansk ou dans l'Adriatique. On trouve d'autres
espèces voisines au Sud des Alpes et des Pyrénées ainsi
qu'en Turquie. (BERG, 1932; BANARESCU, 1960).
En Suisse, PEDROLI S M..(en préparation) le recensent au
Nord des Alpes, principalement dans Le bassin du Rhin
(figure 3) et jusqu'à 1780 m d'altitude (introduit),
affinant par là Les travaux de FATI0C1882).
Figure 2 : Distribution générale du gardon
Figure 3 : Distribution en Suisse du gardon
• I L « oc 1[PBiIItIiIi cti mtsoa ci tcurisscl CE LB «HISSE
I-I I-I I-I . D E S C R I-I P T I-I O N D E S M I-I L I-I E U X
I I I . 1 . GENERALITES
( f i g u r e 4Ï
N o t r e é t u d e a pour cadre q u a t r e Lacs et un canal s i t u é s
i n t é g r a l e m e n t en t e r r i t o i r e h e l v é t i q u e ; il s ' a g i t , par
o r d r e de g r a n d e u r , d e s ' lacs de N e u c h â t e l , des Q u a t r e
-C a n t o n s , de B i e n n e et du L o c L a t ; ainsi que du canal de la
T h i e l l e r e l i a n t les lacs de N e u c h â t e l et de B i e n n e .
Tous ces m i l i e u x font p a r t i e d'une vaste région à lacs
d ' E u r o p e qui était a u t r e f o i s r e c o u v e r t e par les g r a n d s
g l a c i e r s a l p i n s .
Les lacs de N e u c h â t e l et de B i e n n e , g r o u p é s avec celui de
Mo r a t , f o r m a i e n t un tout a p r è s le retrait des g l a c i e r s .
S u i t e à un a b a i s s e m e n t n a t u r e l des e a u x , ces lacs se sont
i n d i v i d u a l i s é s , b i e n q u ' o c c a s i o n n e l l e m e n t e n c o r e unis
lors de f o r t e s c r u e s . A c t u e l l e m e n t , d e p u i s 1 8 8 9 , des
c o r r e c t i o n s du s y s t è m e f l u v i a l font q u e ces t r o i s Lacs
s u b j u r a s s i e n s , n o m m é s ainsi p a r c e q u e s i t u é s au pied de la
c h a î n e m o n t a g n e u s e du Jura tout à L ' e x t é r i e u r de L'arc
a l p i n , e n c o r e Largement r e l i é s e n t r e eux par les canaux de
La T h I e L I e ( N e u c h â t e l — > B i e n n e ) et de La B r o y é C M o r a t — >
N e u c h â t e l ) , sont b i e n s é p a r é s ( Q U A R T I E R , 1 9 4 8 ) .
L ' o r i g i n e du L o c l a t , si tué à p r o x i m i t é de l ' e x t r é m i t é
N o r d - E s t du lac de N e u c h â t e l est s i m i l a i r e , iL s'agit en
fait d ' u n bras de ce d e r n i e r isolé s e c o n d a i r e m e n t à la fin
de la p é r i o d e g l a c i a i r e (BAER, 1 9 6 2 ) .
Lac de La S u i s s e C e n t r a l e , Le lac des Q u a t r e - C a n t o n s est
d ' o r i g i n e g l a c i a i re.
III.2. LAC DE NEUCHATEL
(f i gure 5)
Avec une surface de 214,6 km2, il est le plus grand lac
situé entièrement en Suisse. Son altitude est de 429,3 m.
Ses longueur et Largeur maxima les sont respectivement de
38,3 km et de 8,1 km. Son volume est de 13,77 kra3, sa
profondeur maximale de 153 m et sa profondeur moyenne de
64 m. Le bassin versant du lac représente une surface de
2672 kro'2 et ses trois principaux affluents CTh iel le(Orbe),
Broyé et Areuse) ont chacun un débit moyen d'environ 13
m3/s, L'émissaire unique est La ThîelLe avec un débit
d'environ 53 m3/s. Les mouvements globaux (58 m3/s)
provoquent un séjour des eaux de 8 ans et 82 jours
CSOLLBERGER, 1974).
Le taux de saturation en oxygène des eaux Les plus
pro-fondes, ainsi que la concent rat ion en phosphore tota L
(actuellement en légère diminution) (graphe 1, annexe 1,
page 111) permettent de qualifier ce lac de mésotrophe
(POKORNI, 1983 et 1984; SERVICE CANTONAL DE LA PROTECTION
DE L'ENVIRONNEMENT, 1984, 1985 et 1986, comm. pers.).
Toute la rive Sud ainsi que L'extrémité Nord-Est du lac
sont caractérisées par des zones peu profondes et
sableuses avec une végétât ion constituée de très vastes
roselières, avec notamment Phragmites communi s ( L . ) , et de
beaucoup de joncs des tonneLiers (Schoenoplectus lacustris
(TRIN.)), relevons de plus que Potamogeton pectinatus (L.)
est la principale espèce formant les herbiers; ces régions
de la zone littorale (selon QUARTIER (1948), 27
X
de la
surface du lac, c'est-à-dire 58 km2) représentent un
biotope idéal pour La fraie de nombreuses espèces de
Cypri nidés.
La faune piscicole de ce Lac compte, selon l'Atlas de
répartition des poissons et écrevisses de la Suisse
(PEDROLI 8 A_L., en préparation), 32 espèces de poissons.
III.3. LAC DE BIENNE
(figure 6)
Sa surface est de 39,3 km2, sa longueur maximale de 15 km
et sa largeur de 4,2 km. Avec des profondeurs maxima Le et
moyenne de respectivement 74 m et 30,5 m, il atteint 1,24
km3 de volume. Son altitude est de 429,15 m. Depuis 1878,
suite au remaniement fluvial, son bassin versant a passé
de 3165 km2 à 8305 km2 120
X
du territoire Suisse), dès
lors la circulation de ses eaux est de 240 m3/s
(ancienne-ment 55 m3/s) et leur séjour théorique moyen de 58 jours
(anciennement 253 jours), avec comme conséquence
importante l'évolution de son degré d'eutrophisation. En
effet, depuis les années 60, ce lac peut être considéré
comme eutrophe. (graphe 2, annexe 1, page 111).
La zone Littorale, 8,6 km2, représente le 22
X
de la
surface. CTSCHUMI S ^ L . , 1982; TSCHUMI, 1986).
32 espèces de poissons y sont recensées (PEDROLI fi A_L., en
préparât ion).
III.4. LAC DU LOCLAT
(figure 7)
En forme de cuvette, avec un fond très plat et un pourtour
constitué de talus très inclinés, ce petit lac, 0,046 kn2,
est profond d'environ 9,5 m et représente un volume de
0,0003 km3. Ses dimensions maximales avoisinent 470 m de
Longueur et 115 m de largeur. Son altitude est de 432 m et
la surface de son bassin versant est de 0,68 km2. Soumis
dans les années 50 à divers travaux d'irrigation et de
correction, son écoulement est actuellement très faible et
il est considéré comme eutrophe (CLUB DES AMIS DE LA
NATURE DE NEUCHATEL, 1907; BAER, 1962).
Du mois d'avril au mo is d'octobre, on observe une
disparition totale de l'oxygène dès 5 m de profondeur. Le
phytoplancton y est "abondant", par contre il est "très
pauvre" en zooplancton (ARAGNO : communi cation orale,
1986; SCHWEIZER 8 AL., 1975).
Les pêcheurs à La ligne prétendent qu'environ 6 espèces de
poissons vivent dans ce lac (communications orales,
1984/86).
III.5. LAC DES QUATRE-CANTONS
(figure 8)
Il s'agit d'un lac mésotrophe tendant à nouveau à
l'oligo-trophie (graphe 3, annexe 1, page 111). Le renouvellement
de ses eaux est de 3 ans et 142 jours (110 m3/s). Son
volume est de 11,8 k m 3 et son bassin d'alimentation de
2124 km2. Il est situé à une altitude de 433,6 m. Sa
surface est de 113,6 km2, sa longueur maximale de 39,8 km
et sa largeur maximale de 4,2 km. Ses profondeurs maxi ma le
et moyenne sont respectivement de 214 m et 104 m (DOLDER,
1977; S01LBERGER, 1974; STADELMANN, 1984; AMBLIEHL, 1986;
EAHAG, communi cation personnel le 1987).
Selon le recensement effectué par l'Atlas de répartition
des poissons et écrevisses de La Suisse (PEOROLI S
AL^
., en
préparation), 29 espèces de poissons y vivent.
III.6. CANAL DE LA THIELLE
Reliant, artificiellement depuis La première correction
des eaux du Jura, le Lac de Neuchätel au lac de Bienne, le
canal de la ThieLle a une longueur de 8 km, sa Largeur â
La surface de L'eau est de 48 à 52 m et sa profondeur
avoisine Les 5 m. Malgré d'occasionnels refoulements
estivaux, Le débit moyen des eaux en direction du lac de
Bienne est de 53 m3/s et La vitesse du courant varie de 0
à 0,5 m/s (QUARTIER, 1948; S0LLBERGER, 1974).
La Thielle, dont Les rives sont constituées d'enrochements
et le fond de vase, appartient à La région cyprinicole.
LAC DE NEUCHÂÏEL LAC DE BIENNE LAC DU LOCLAT CANAL DE LA THIELLE
FiGURE 7 : L A C DU L O C L A T , - 9 m
v>---->
F I G U R E 8 : I_AÇ DES Q U A T R E - C A N T O N S BRUNMEN ALPNACHSTADIV. B I O M E T R I E
IV.1. INTRODUCTION
D'une manière générale, La systématique des Cyprinidés est
re Lati ve ment complexe. Les différences taxonomi que s sont
habituellement basées sur des relevés morphométriques, de
nombreux caractères biométriques dénombrables ainsi que
sur la structure des dents pharyngiennes.
Le gardon (Rut i lus rut i tus L.) est L'un des Téléostéens
Les plus répandus et communs de La zone Eurasienne et L 'on
constate, en examinant La littérature, qu'il existe de
nombreuses petites différences concernant Les cri teres
de détermination Les plus courants, selon les auteurs
consultés; citons notamment BERGC1949) qui en décrit 11
sous-espèces. Cependant, HOLCIK
&
SKOREPA¢1971), après
avoir constaté des variâtions selon les facteurs
de l'environnement ainsi qu'un clivage géographique
Nord/Sud, regroupent toutes ces formes dans une espèce
polymorphe, RutiLus rutitus ( L
1) .
IL nous a par conséquent paru intéressant d'établir pour
chacun des milieux étudiés et sur La base d'un matériel
abondant. Les caractères de déterminât ion types,
ut ili sables en déterminât ion courante, des gardons; ceci
afin de déceler d'éventuels écotypes et d
1effectuer des
comparaisons avec Les critères établis ailleurs par
d'autres auteurs.
Les cas naturels d'hybridation, au niveau spécifique et
même générique, sont Largement répandus chez les
Cyprinidés, mais n'ont jusqu'ici pas suscité beaucoup
d'intérêt chez Les ichthyologistes. Par conséquent, chaque
fois que nous rencontrions des difficultés à classer un
cyprin selon Les clés et les caractères discriminatoires
connus, un examen biometrique était entrepris.
IV.2. METHODES
Le matériel provient essentiellement de pêches
expéri menta Les effectuées au moyen de fi lets mai liants
multimailles (voir point V.3.2.1, page 5 0 ) , dans les lacs
de Neuehâtel, Bienne, Loclat et des Quatre-Cantons. De
plus, un certain nombre d'individus provenant des prises
des pêcheurs professionnels du Lac de Neuchâtel ont
également été examinés.
IV.2.1. Mesures
(figure 9)
Pour chaque individu, outre la provenance, La longueur
tota le CLT) avec La nageoire caudale normaLement déployée,
La longueur du corps <ou longueur standard) (LC) et Le
poids du poisson frais entier (PDS) ont été relevés.
Nous avons choisi d'utiliser la Longueur totale dans nos
résultats et c'est afin de permettre des comparaisons avec
les travaux d'autres auteurs, que nous avons calculé
(régression par la méthode des moindres carrés), les
équations de conversion "Longueur tota Le/longueur du
corps", ceci séparément pour les populations de chacun des
milieux. Pour le même motif, nous donnons en annexe 2 La
relation "longueur tota Le/longueur maxima Le" (nageoire
caudale rabattue) établie en Suisse pour les gardons par
MUELLER (1984).
Un calcul identique, basé sur les logarithmes des données,
nous a fourni Les regressions relati ves aux relations
"longueur tota le/poids". Ces différentes droites ont été
comparées se Ion la méthode décrite par HALD(1969).
Fi gure 9 : M e s u r e s de Longueurs
, LWCUEm AJ COBPS (LC) UWXCLR «!«LE ( U )
' U M J j E m DWIIWLC <LTI)
I V . 2 . 2 . C a r a c t è r e s d é n o m b r a b l e s
Les critères u t i l i s é s couramment dans Les cLés de d é t e r m i n a t i o n ont été r e t e n u s , ainsi La s t r u c t u r e des dents p h a r y n g i e n n e s a été examinée et les caractères s y s t é m a t i q u e s d é n o m b r a b L e s . ont été r e l e v é s , puis Les r é s u l t a t s furent soumis à une anaLyse s t a t i s t i q u e sommaire destinée à fournir m o y e n n e s et é c a r t s - t y p e s a La m o y e n n e . Un test d ' h o m o g é n é i t é a e n s u i t e permis de juger si les v a r i a t i o n s entre Les valeurs o b t e n u e s pour les d i f f é r e n t s m i l i e u x p o u v a i e n t être c o n s i d é r é e s corame étant s i g n i f i c a t i v e s <seuil de sécurité de 9 9 X ) , ( L A M O T T E , 1 9 7 1 ; H E L L E R , 1 9 7 A ) ; Le cas é c h é a n t , le coefficient de d i f f é r e n c e d e MAYR C 1 9 6 9 ) , évaluant Le taux de non
recouvrement de deux é c h a n t i l l o n s , était c a l c u l é .
I^V.2^2^1^ £en.tj5 £h3.rZn£''£.nrie£
Les dents p h a r y n g i e n n e s , o r g a n e s de m a s t i c a t i o n • e t de b r o y a g e p a r t i c u L i e r s aux C y p r i n i d é s , ont été p r é l e v é e s après a b l a t i o n des o p e r c u l e s et des arcs b r a n c h i a u x . ELLes ont e n s u i t e été e x a m i n é e s à la loupe b i n o c u l a i r e .
(figure 10)
Trois d é c o m p t e s d ' é c a i l l é s ont été e f f e c t u é s .
1 . ^ 2 ^ 2 ^ 2 ^ 1 ^ L_on_gj_ti£d_i_na_l : comptage des é c a i l l e s cana L i-culées de la ligne latérale (ex : L L A T = 4 2 ) .
1 . ^ 2 ^ 2 ^ 2 ^ 2 ^ Xra_njLvJLr.§,al. : comptage des é c a i l l e s d o r s o
-l a t é r a L e s , re-liant Le p r e m i e r rayon de -la n a g e o i r e d o r s a -l e à la ligne l a t é r a l e , et v e n t r o - l a t é r a l e s , reliant le p r e m i e r rayon de la n a g e o i r e ventrale à la Ligne latérale (ex : DLAT=8 . et V L A T = I + 5 , Le I r e p r é s e n t e l'écaillé c a n a l i c u l é e de La ligne l a t é r a l e ) .
ly^sJ-^ì. U,é.ç_o£p.t_e.s_ d_es_ _ra^o^s_d£s_na_g£o_i_re^ (figure 10)
D é n o m b r e m e n t des rayons mous des n a g e o i r e s d o r s a l e s C D ) , c a u d a l e s C C ) , anales ( A ) , v e n t r a l e s CV) et p e c t o r a l e s C P ) . La lettre désigne La n a g e o i r e , ainsi par e x e m p l e 0 = 1 1 1 + 1 0 , s i g n i f i e que la n a g e o i r e d o r s a l e présente 3 rayons d u r s p r é c é d a n t 10 rayons m o u s .
F i g u r e 10 : Localisat ion des d é c o m p t e s d ' é c a i l l é s et de rayons des nageoires
IV.3. RESULTATS GARDONS
IV.3.1. Relations "Longueur totale / longueur du corps" (fi gure 11)
Les équations exprimant le plus fidèlement les conversions "longueur totale CLT>/longueur du corps (LC)" sont Les suivantes :
Lac de Neuchâtel : LC = 0,8215 LT + 0,757 r=0,999 n=l824
Lac de Bienne : LC = 0,8354 LT - 1,340 r=0,999 n= 200
Lac du LocLat : LC = 0,7980 LT + 1,101 r=0,989 n= 234
Lac des 4-Cantons : LC = 0,8150 LT + 1,804 r=0,997 n= 94
Figure 11 : Relations "longueur totale /• longueur du corps"
ICB m 2ËB 2 5 9 3BB 356HH.
LOMEUI! TOTALE
'!HEUCHSIEL EJBIEKNE IMlAX «iqDATBE-CfiHIOKS
Pour les trois grands Lacs, la relation "longueur du corps/longueur totale" est identique ( P < 0 , 0 1 ) . Les gardons du. LocLat présentent quant à eux, à Longueur du corps égale, une Longueur tota Le légèrement supérieure, autrement dit leurs nageoires eauda les sont proport i on-neLLement plus longues que celles des individus provenant des autres lacs < P < 0 , 0 1 ) .
IV.3.2. Relations "Longueur totale
I
poids"
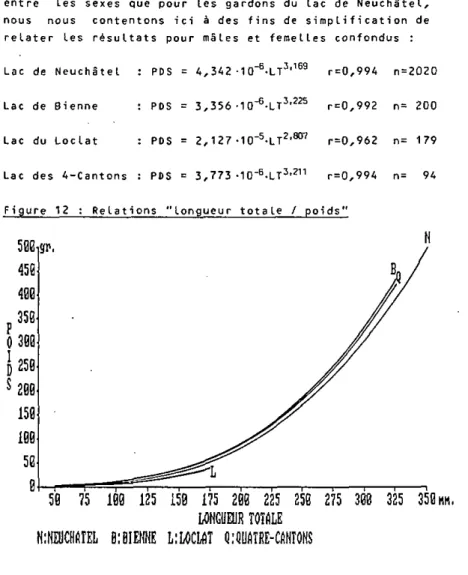
(figure 12)
Les tests stati s tiques convent ionne Is n'ayant re Levé, pour
cette relation, des différences significatives (P<0,01)
entre Les sexes que pour les gardons du Lac de Neuchitel,
nous nous contentons ici à des fins de simplification de
relater les résultats pour mâles et feme Lies confondus :
Lac de Neuchàtel : PDS = 4,342 -1 O
-^LT
3'
169r=0,994 n=2Û20
Lac de Bienne : PDS = 3,356-10^-LT
3'
225r = Ü,992 n= 200
Lac du Loclat : POS = 2,1 27-lO^-LT
2,807r=0,962 n= 179
Lac des 4-Cantons : PDS = 3,773 -10"
6.LT
3'
211r=0,994 n= 94
Figure 12 : Relations "Longueur totale / poids"
50 75 1 0 0 125 150 175 2 0 0 225 250 275
LONGUEUR TOTALE
NîNEUCHAÎEL BlBIENNE LïLOCLfiT QlQUflTßE-CftNTOHS
325 350HM.
Proportionnellement, Les individus du Loclat sont
significativement les plus légers, viennent ensuite ceux
de NeuchâteL, puis ceux des Quatre-Cantons. Les plus
Lourds sont Les gardons capturés à Bienne < P < 0 , 0 1 ) .
IV.3.3. Décomptes des écailles
n ^ ^ ^ U L_ongvtudj_n£l
(figure 13 / tableau 1)
On observe une stabilité évidente du caractère entre les
individus provenant des 4 Lacs étudiés. La moyenne est
néanmoins Légèrement inférieure pour Les gardons du Loclat
C P < 0 , 0 1 ) .
Tableau 1 : Résultats du décompte des écailles
canaliculées de La Ligne latérale
nombre moyen
écart type
nombre d'individus
Neuchâte L
44,01
1,48
105
Bi enne
44,06
1,37
68
Figure 13 : Fréquence des nombres
d'écailLes canaliculées de la Ligne Lat
Loc Lat
43,13
0,64
113
éra Le
4-Cantons
44,02
1,27
93
40 4 1 4 2 43 4 4
HUMERI D'ECAILLES
CAHHL[CULEES
DE Lfl
I NEUCHATEL OBIEHNE B LOCWT (B QUATRE
tfcI05 N:fiB N=1S2 H=93
45 4 6 4 7
LATEHALE
(figures 14 et 15
I
tableau 2)
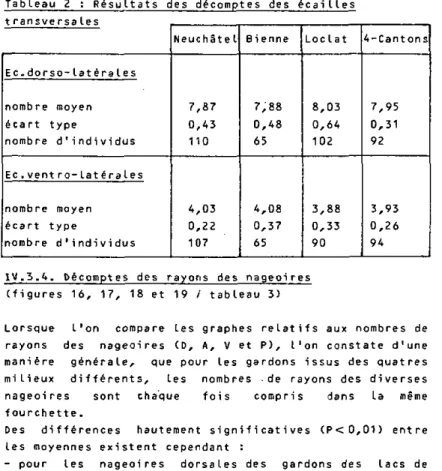
Les moyennes des nombres d 'écailles dorso-latérales des
échantillons sont "similaires" à un degré de certitude de
99X pour les 4 localités, néanmoins une différence
significative à 95% de ce caractère existe entre les
gardons issus du Lac de NeuchStel et ceux du Loclat .
Seules les moyennes des écailles vent ro-latérales des
populations des lacs de NeuchStel et de Bi enne ainsi que
celles des lacs du LocLat et des Quatre-Cantons ne
diffèrent pas si gni f i cati'vement ( P > 0 , 0 5 ) .
Tableau 2 : Résultats des décomptes des écailles
t ransversales
Ec.dorso-latérales
nombre moyen
écart type
nombre d ' i ndi vi dus
Ec.ventro-Latérales
nombre moyen
écart type
nombre d'individus
Neuchâtel
7,87
0,43
110
4,03
0,22
107
Bienne
7,88
0,48
65
4,08
0,37
65
Loclat
8,03
0,64
102
3,88
0,33
90
4-Cantons
7,95
0,31
92
3,93
0,26
94
IV.3.4. Décomptes des rayons des nageoires
(figures 16, 17, 18 et 19 / tableau 3)
Lorsque L'on compare les graphes relatifs aux nombres de
rayons des nageoires (D, A, V et P ) , l'on constate d'une
manière générale, que pour les gardons issus des quatres
milieux différents, Les nombres -de rayons des diverses
nageoires sont chaque fois compris dans La même
fourchette.
Des différences hautement significatives (P<0,01) entre
Les moyennes existent cependant :
- pour les nageoires dorsales des gardons des lacs de
Bienne et de Neuchâtel ainsi que de Bienne et du LocLat.
F i g u r e 14 : F r é q u e n c e des n o m b r e s d ' é c a i l l é s d o r s o - l a t é r a l e s
100
l n
l
Qu 50
£
N
E
20
110
B
p ü ™
7 8 9 10
N M D'ECAILLES DORSO-LfiTOftLES
lNEUCHfllEL DBIENNE B LOCLflTffi
QUfllRE-H=110 H=65 N:113 N=93
Figure 15 : Fréquence des n o m b r e s d'écaillés v e n t r o - l a t é r a l e s
F
K
K
f]
II
E
N
C
L
y
70
fiR<
Mi
46
30
20
IU
JL
Jl
7 8 9
NOHBREI)
1ECAILLESIlENTRU-LfIIEHALES
INEUCHAIEL DBIENNE BLOCLAI
DDQUflTRE-N=IB? N:65 N:90 N=94
- pour Les nageoires anales des individus issus des Lacs
du LocLat et des Quatre-Cantons, du LocLat et de Bienne
ainsi que des Quatre-Cantons et de Neuchâtel.
- pour Les nageoires ventraLes des poissons provenant des
lacs de Neuchâtel et du LocLat,
- pour les nageoires pectorales des gardons du lac des
Ouatre-Cantons avec celles des poissons issus des trois
autres mi Lieux. Citons de plus la différence
significa-tive à 95X observée pour ce caractère entre les poissons
du lac de Bienne et ceux du LocLat.
Tableau 3 : Résultats des décomptes des rayons des
Rayons nag.dorsales
nombre moyen
écart type
nombre d'individus
Rayons nag.caudaLes
nombre moyen
écart type
nombre d'individus
Rayons nag.ana Les
nombre moyen
écart type
nombre d'individus
Rayons nag.vent rales
nombre moyen
écart type
nombre d ' i ndi vi dus
Rayons nag.pectorales
nombre moyen
écart type
nombre d'individus
Neuchâtel
9,92
0,40
113
19
0
46
10,16
0,49
115
7,92
0,27
115
15,94
0,80
115
Bi enne
10,06
0,30
77
19
0
41
10,31
0,59
77
7,97
0,23
76
16,05
0,73
75
Loclat
9,91
0,33
139
19
0
25
10,10
• 0,53
160
7,99
0,15
139
15,77
0,81
133
4-Cantons
10,01
0,47
95
19
0
54
10,36
0,58
95
7,98
0,21
95
16,85
0,82
95
Figure 16 : Fréquence des nombres
de rayons de La nageoire dorsale
9 10 I i
NOMBRE DE R A » DE LfI NAGEOIRE DORSALE
INEÜCHAIEL OBIENHE 8WCLAI
fljflUAÏHE-«=113 f t ? ? N:139 N=95
Figure 17 : Fréquence des nombres
de rayons de la nageoire anale
J N
E 60
QU 5ß
N «
C 30
28
>> 1 0
JCEL
G
g î e i l 12 13
NOMBRE DE RAVONS DE LA NAGEOIRE AMLE
INEUCHAIEL G BIENNE S LOCLAT Ol QUAIRE-CANIOf)S
K=115 N:?? N:160 N=95
F i g u r e 18 : F r é q u e n c e des n o m b r e s de rayons des n a g e o i r e s v e n t r a l e s
.168
98
88
l
™
R
î 60
Q
Il M
S «
0
f 38
E
28
V
10
8
jT-rm
:
7 B S
KOKBHE DE BAVONS DES IiSGEOIRtS UOURALIS
iNEUCHflïEL D B « BLÖCLAI
[DQUATRE-H=IlS H=7G H=I39 rfc95
Figure 19 : Fréquence des nombres de rayons des n a g e o i r e s p e c t o r a l e s
75.8
67,5
F 52.5
E 4 5 . 0
Q
jj 37.5
l 38,8
E
22,515.0
X7,5
il
J":
i
i
14 15 16 17 18
KOKBBE DE RAVOKS DES NAGEOIHES PECTORALES
IKHJCHfIIEL GBIENNE 8 LOCLAI (D QUATRE
I V . 3 . 5 . Dents p h a r y n g i e n n e s (figures 2D et 21 / tableau 4 )
La stucture des dents p h a r y n g i e n n e s des individus d i f f è r e s e n s i b l e m e n t en fonction de leur p r o v e n a n c e ; en e f f e t , la majori té (552) des p o i s s o n s du Loclat présente 5 d e n t s , alors que pour les gardons des trois autres m i l i e u x , elle se situe à 6 dents (51 à Ó 1 X ) . La m o y e n n e des dents des é c h a n t i l l o n s du Loclat d i f f è r e d'ailleurs s i g n i f i c a -ti vement (P < 0,01) de celles des individus provenant des a u t r e s lacs.
T a b l e a u 4 : R é s u l t a t s du décompte des dents p h a r y n g i e n n e s
n o m b r e moyen écart type n o m b r e d'individus N e u c h â t e L 5,49 0,71 88 Bi enne 5,53 0,63 77 Loc lat 5,15 0,66 124 4 - C a n t o n s 5,57 0,56 89
Figure 20 : Fréquence des n o m b r e s de dents p h a r y n g i e n n e s
Figure 21 : S t r u c t u r e des dents p h a r y n g i e n n e s
4 5 6 7
NONBRE CE CENTS FKARVtIG[EKHES
I NIUCHfìTEL 0 BIENNE 0 LOCLAT Q]
I V . 3 . 6 . R é s u m é des comparai sons e n t r e tes c a r a c t è r e s
bi o m é t r i q u e s / t a b l e a u 5
R a p p e Ions que c'est s e u l e m e n t Lorsque Le test de signi f i
-c a t i o n était p o s i t i f , que Le -c o e f f i -c i e n t de d i f f é r e n -c e de
M A Y R C 1 9 6 9 ) a été c a l c u l é .
T a b . 5
LC/LT
LT/PDS
LLAT
DLAT
VLAT
D
A
C
V
P
DENTS
NS. CAS
POSITIFS
NEU
SIG
+ + + + + + + + + + + + +7
LOC
CD
-BIE
SIG
+ + + + + + + + + + ++ + + +8
LOC
CD
-4CA
SIG
+ + + + + + + + + + + +ó
LOC
CD
-NEU
SIG
+ + + + + +3
BIE
CD
-NEU
SIG
+ + + + + + •t +4
4CA
CD
-BIE/
SIG
+ + + + + +3
4CA
CD
-S I G = t e s t de signi fi c a t i o n : ++ = d i f f é r e n c e h a u t e m e n t
s i g n i f i c a t i v e ( P < 0 , 0 1 ) .
+ = d i f f é r e n c e s i g n i f i c a t i v e
( P < 0 , 0 5 > .
C D = c o e f f i c i e n t de d i f f é r e n c e : - - taux de non r e c o u v r e m e n t
des deux d i s t r i b u t i o n s
i n f é r i e u r à 7 5 X.
I V . 4 . D I S C U S S I O N G A R D O N S
Q u e l q u e s c o n c l u s i o n s p r é l i m i n a i r e s , r e l a t i v e s à nos
p r o p r e s r é s u l t a t s s'imposent :
- les r e l a t i o n s e n t r e La l o n g u e u r et le poids ne sont pas
d i r e c t e m e n t p r i s e s en c o m p t e d a n s les c o n s i d é r a t i o n s
b i o m é t r i q u e s , ceci pour des rai sons qui seront d i s c u t é e s
p l u s Loin.
- l'examen des g r a p h e s r e l a t i f s aux r e l a t i o n s " L o n g u e u r
t o t a l e / l o n g u e u r du c o r p s " , ainsi que de ceux r e l a t i f s
aux c a r a c t è r e s d é n o m b r a b L e s , m o n t r e à L ' é v i d e n c e , m a l g r é
de légères d i f f é r e n c e s , une f o r t e s i m i l i t u d e e n t r e les
g a r d o n s p r o v e n a n t des d i f f é r e n t s l a c s .
- r a p p e l o n s que Les c a l c u l s des r e l a t i o n s " L o n g u e u r t o t a l e
/ l o n g u e u r du c o r p s " n ' a v a i e n t pas comme but e x c l u s i f la
d i f f é r e n c i a t i o n en é c o t y p e s des g a r d o n s pré Levés d a n s
tel où tel m i l i e u . Ces é q u a t i o n s d e v a i e n t é g a l e m e n t
p e r m e t t r e La c o m p a r a i son de nos m e s u r e s avec c e l l e s
d ' a u t r e s a u t e u r s . N é a n m o i n s , la d i f f é r e n c e s i g n i f i c a t i v e
e n t r e La r e l a t i o n é t a b l i e pour Les g a r d o n s du L o c l a t et
c e l l e s , s i m i l a i r e s e n t r e - e l L e s , c a l c u l é e s pour les
a u t r e s l a c s , m é r i t e d ' ê t r e r e t e n u e ( P < 0 , 0 1 ) . Les
p o i s s o n s du LocLat d i f f è r e n t des indivi dus c a p t u r é s d a n s
les a u t r e s m i l i e u x , par une r e l a t i o n " L o n g u e u r - corps /
l o n g u e u r - n a g e o i r e c a u d a l e " s e n s i b l e m e n t p l u s é l e v é e .
- des d i f f é r e n c e s s i g n i f i c a t i v e s e x i s t e n t é g a l e m e n t
p a r f o i s e n t r e Les c a r a c t è r e s d é n o m b r a b L e s m o y e n s des
p o p u l a t i o n s de p o i s s o n s p r o v e n a n t de ces d i f f é r e n t s
m i l i e u x . Dans ces c a s , le c a l c u l du c o e f f i c i e n t de
d i f f é r e n c e de M A Y R C 1 9 6 9 Ï a été e f f e c t u é a f i n de d é f i n i r
si les p o p u l a t i o n s p o u v a i e n t ê t r e , sur La base des
c a r a c t è r e s r e t e n u s , d i s t i n g u é e s Les unes des a u t r e s . Il
ressort de ces c a l c u l s q u ' a u c u n e d i s t i n c t i o n f i a b l e
n'est possi ble (¾ de non r e c o u v r e m e n t m a x i m u m i n f é r i e u r
à 7 5 X ) C v o i r point I V . 3 . 6 . ) .
- les v a l e u r s , parfois légèrement inférieures (caractères d é n o m b r a b l e s ) , t r o u v é e s pour Les p o i s s o n s du L o c l a t , ont p r o b a b l e m e n t été a c c e n t u é e s par les d i f f i c u l t é s é p r o u v é e s lors des c o m p t a g e s ; en e f f e t , les individus p r o v e n a n t de ce Lac s o n t , comme nous le v e r r o n s plus t a r d , d'une taille notablement inférieure à ceLle des i n d i v i d u s issus des autres milieux i n v e s t i g u é s .
IL ressort n é a n m o i n s que : Les g a r d o n s issus des lacs d e N e u c h â t e L et de B i e n n e ne d i f f è r e n t que faiblement par deux c a r a c t è r e s . Les d i f f é r e n c e s entre ces p o i s s o n s et ceux du Lac des Q u a t r e - C a n t o n s sont un peu plus p r o n o n c é e s , citons notamment le fai b Le nombre d'écaillés v e n t r o - l a t é r a l e s et surtout le grand nombre de rayons des n a g e o i r e s p e c t o r a l e s des individus provenant de ce lac. Les g a r d o n s du Loclat divergent des autres par p l u s i e u r s c a r a c t è r e s , 5 d i f f é r e n c e s sont constatées avec les p o i s s o n s issus du lac des Q u a t r e - C a n t o n s , 6 avec ceux de N e u c h â t e L et 7 avec les individus provenant du Lac de Bi e n n e .
Les " a n c i e n s " a u t e u r s décrivaient de n o m b r e u s e s s o u s -e s p è c -e s d-e gardons (FATIO, 1 8 8 2 ; B E R G , 1 9 4 9 ) . Actu-e L-l e m m e n t , Les c h e r c h e u r s s ' a c c o r d e n t , ci tons notamment S P I L L M A N N C 1 9 6 1 ) , HOLCIK S A L . ( 1 9 7 1 ) , pour regrouper toutes ces formes dans une espèce p o l y m o r p h e . Rut i lus rut ilus ( L . ) , fortement influencée par les facteurs de l ' e n v i r o n n e m e n t . C'est pour cette raison que nous n'avons pas jugé utile d ' e n t r e p r e n d r e une a n a l y s e s t a t i s t i q u e m u l t i v a r i é e dont Le but aurait été de d i f f é r e n c i e r a b s o l u m e n t Les p o p u l a t i o n s é t u d i é e s .
On peut n é a n m o i n s f a c i l e m e n t , sur La base de nos r é s u l t a t s , d é f i n i r trois p o l y m o r p h e s , à savoir : un regroupant les p o p u l a t i o n s des lacs de N e u c h â t e L et de B i e n n e q u i , rappelons Le, sont très p r o c h e s et largement reliés e n t r e - e u x . Un autre écotype concerne tes g a r d o n s du L o c l a t , lac qui bien que très proche g é o g r a p h i q u e m e n t des deux lacs cités précédemment p o s s è d e des c a r a c t é r i s -t i q u e s m o r p h o l o g i q u e s , biologiques e-t b i o c h i m i q u e s -très
différentes. Enfin la troisième forme comprend les
individus du lac des Quatre-Cantons, lac géographiquement
éloigné des précédents.
Fort des conclusions ci-dessus émises, il nous est
possible d'établir des caractéristiques systématiques
types, ut ili sab les en détermination courante (la relation
LT/LC n'est pas considérée car la comparai son de droites
de régression nécessite de nombreux critères rarement
cités par les auteurs), valables pour les gardons
provenant des 4 lacs étudiés :
-é c a i l l e s c a n a l i c u l -é e s de La
é c a i l l e s d o r s o - l a t é r a l e s
é c a i l l e s v e n t r o - l a t é r a les
r a y o n s n a g e o i r e s d o r s a l e s
r a y o n s n a g e o i r e s c a u d a l e s
r a y o n s n a g e o i r e s ana Les
r a y o n s n a g e o i r e s v e n t r a l e s
r a y o n s n a g e o i r e s p e c t o r e Les
d e n t s p h a r y n g i e n n e s (sur un
ligne
r a n g )
latéra
e
I
III
III
I
I
+
+
• t t40
7
3
8
19
8
7
14
4
-48
10
5
11
13
9
18
7
Il apparaît que la stabilité régionale observée, ainsi que
la comparaison avec les autres auteurs suisses (FATIO,
1882; QUARTIER, 1980; PEDROLI, 1983) - la fourchette de
nos résultats eng Lobant Largement leurs descriptions
-permet d'extrapoler la description systématique ci-dessus
aux gardons (Rut i Lus ruti lus L.) présents dans Les eaux
he Lvét iques.
La provenance du matériel est importante, beaucoup
d'auteurs L'ont obtenu dans des milieux différents ou
des musées. Les résultats de FATI0(1882) ainsi que ceux
de QUARTIERO980) sont basés sur l ' examen de poissons
récoltés en Suisse; PEDROLI (1983) a obtenu ses gardons
dans un petit lac du Plateau suisse (Seedorf FR) et
SPILLMANN C1961) quant à Lui, ti re ses critères
d'individus issus "d'eaux françaises". Ceux de LIBOVARSKY
& AL.(1984) proviennent de gardons vivant dans un lac de
retenue situé dans Le cours inférieur de La rivière Dy je
en Tchécoslovaquie. Les critères d'HOLCIK S M..(1971)
résultent d'une synthèse de prélèvements effectués sur
toute L'aire de répartition de L'espèce et représentent par c o n s é q u e n t Les critères e x t r ê m e s du gardon (Rutitus r u t i L u s L. ) .
T a b L e a u 6 : Synthèse des d e s c r i p t i o n s b i o m é t r i q u e s du gardon
LLAT DLAT VLAT
D
C
A
V
P
DENTS FATIO (1882) 40-46 7-8(9)3-4
9-1119
9-138-9
(14)15-185-6
QUARTIER (1980) 41-45 -8-11 -8-11 -PEDROLI (1983) 41-46 8(9) 4(5) -10-12-5-6
SPILLMANN (1961) 42-45(46)7-8
(3)4-5 9-11 (18)19 9-117-8
15-185-6
HOLCIK (1971) 36-47 -7-11 -7-13 -LIBOV. (1984) 39-437-9
3-4
10-11 18-20 10-127-9
12-155-6
PRESENT TRAVAIL 40-48 7-103-5
8-1119
8-137-9
14-184-7
La c o m p a r a i s o n entre Les d e s c r i p t i o n s faites par Les d i f f é r e n t s auteurs conduit aux réflexions s u i v a n t e s :
- il est d o m m a g e que Le calcul des critères m o y e n s ne soit pas plus g é n é r a l i s é . L'examen des moyennes p e r m e t t r a i t des comparai sons b i o m é t r i q u e s plus fines entre les d i f f é r e n t e s descript i o n s , et p e r m e t t r a i t ainsi de d i s t i n g u e r les éventuels é c o t y p e s .
- avec 48 é c a i l l e s le long de la ligne l a t é r a l e , ou seulement 4 dents p h a r y n g i e n n e s , nous avons même d é p a s s é Les e x t r ê m e s " c o n n u s " . IL faut n é a n m o i n s remarquer q u ' H O L C I K S A L . ( 1 9 7 1 ) , en résumant Leurs d e s c r i p t i o n s et celles de d i f f é r e n t s a u t e u r s , citent de rares cas où les Lignes latérales comptent également 48 é c a i l l e s .
- Les c r i t è r e s des g a r d o n s f r a n ç a i s ( S P I L L M A N N , 1 9 6 1 ) sont
i d e n t i q u e s aux n ô t r e s , m a l g r é d ' o c c a s i o n n e l s cas où la
n a g e o i r e c a u d a l e ne p r é s e n t e que 18 r a y o n s .
- si l'on c o m p a r e aux c r i t è r e s o b t e n u s par les a u t e u r s
s u i s s e s et f r a n ç a i s , qui d'une m a n i è r e g ê n é r a i e sont
c o m p r i s dans les m ê m e s f o u r c h e t t e s , ceux de L I B O V A R S K I 8
A L . ( 1 9 8 4 ) , on r e m a r q u e que les n o m b r e s d ' é c a i l L e s de La
Ligne l a t é r a l e et de r a y o n s des n a g e o i r e s p e c t o r a l e s
sont p l u s f a i b l e s . La f o u r c h e t t e du n o m b r e de r a y o n s des
n a g e o i r e s c a u d a l e s quant a e l l e , est plus l a r g e . Il
a p p a r a î t , lors de la c o m p a r a i s o n avec les c r i t è r e s
e x t r ê m e s p r é s e n t é s par H O L C I K & A L . ( 1 9 7 1 ) , que L e u r s
mini m a s sont n e t t e m e n t i n f é r i e u r s à ceux des a u t r e s
a u t e u r s .
Les d i v e r g e n c e s c o n s t a t é e s e n t r e ces d i f f é r e n t e s
d e s c r i p t i o n s p e u v e n t ê t r e a t t r i b u é e s au p o l y m o r p h i s m e du
g a r d o n et au fait que Les é c h a n t i l l o n s p r o v i e n n e n t ,
s u r t o u t pour la d e s c r i p t i o n d ' H O L C I K S A L . ( 1 9 7 1 ) , de
s t a t i o n s très é l o i g n é e s e t / o u d i f f é r e n t e s . Ces a u t e u r s
a f f i r m e n t par e x e m p l e que les b r a n c h i o s p i n e s et le n o m b r e
d ' é c a i l l é s de La Ligne l a t é r a l e a u g m e n t e n t au N o r d et
B A R L 0 W ( 1 9 6 1 ) s o u l i g n e d e s v a r i a t i o n s N o r d / S u d du n o m b r e de
v e r t è b r e s .
L ' u t i l i s a t i o n de la r e l a t i o n " L o n g u e u r t o t a l e / p o i d s " c o m m e
c r i t è r e de d é t e r m i n â t ion spéci fi que n'est é v i d e m m e n t pas
e n v i s a g e a b l e . C e p e n d a n t c e t t e r e l a t i o n , d i r e c t e m e n t
d é p e n d a n t e du m i l i e u , est s u s c e p t i b l e de f l u c t u e r e n t r e
é c o t y p e s d'une m ê m e e s p è c e .
L ' é t a b l i s s e m e n t d e s m e i l l e u r e s r é g r e s s i o n s r e l a t i o n n e l l e s
e n t r e La L o n g u e u r et le p o i d s sont pour d i f f é r e n t e s
r a i s o n s , v a r i a b l e s et d i s c u t é e s . C i t o n s H A N N ( 1 9 7 3 ) et
B U R R O U G H & A L . ( 1 9 7 9 ) qui a p r è s a v o i r é t a b l i des r e l a t i o n s
l o n g u e u r / p o i d s (frai s) s é p a r é m e n t pour les i m m a t u r e s , les
m â l e s et Les feme I L e s , ceci pour d i f f é r e n t e s p é r i o d e s
a n n u e l l e s , c o n c l u e n t q u ' i l n ' e x i s t e a u c u n e d i f f é r e n c e
significative <P > 0,05) entre ces divers groupes de
gardons et entre Les différentes périodes de mesures; ils
calculent dès Lors des équations uniques. A l'opposé,
G0LDSPINK(1979) prétend qu'une régression commune n'est
pas représentât i ve, iL opte pour des prélèvements à date
fixe et distingue mâles et femelles. HUELLER S AL.(1986)
séparent également Les sexes et concluent à un
accroissement pondéral supérieur chez les feme I les.
A ces diverses méthodes, s"ajoute le fait que Les mesures
de longueurs ne sont pas standardisées (longueur du corps,
à la fourche ou totale). Les comparaisons entre les
données des différents auteurs sont donc fort compromises.
Nos courbes relationnelles entre Le poids et La Longueur
totale (figure 12, p.18) sont établies sur des données
brutes prenant en compte le poids frai s à la capture,
cette mesure étant se Ion RICKER(1980) la plus fréquemment
utilisée en recherche piscicole. Néanmoins, pour pouvoir
apporter quelque crédit a ces relations, il convient de
les utiliser précautionneusement, en tenant compte des
faits suivants :
- le poids est susceptible de varier avec le sexe. L'état
de maturité sexuelle, La saison, Le milieu, l'état de
nutrition, e t c . . De la période d'échantillonnage dépend
. donc dans une large mesure le poids moyen pour une
longueur donnée.
- nous avons exclu L'examen des différences pondérales
entre sexes. Nos courbes et équations s'appliquent donc
aux sexes réunis, cependant La structure particulière du
sex-ratio, qui comme nous le verrons plus loin, comporte
une proportion dérisoire de mâles, fait que ces
relations "longueur tota le/poids" sont établies presque
exclusivement sur des individus femelles.
Les mesures pour le lac de Neuchâtel sont faites sur la
base de nombreux prélèvements effectués en toutes sai sons
sur un laps de temps de plusieurs années, la relation
obtenue permet donc une approche assez fidèle de La
moyenne pondéra Le annuel le pour une longueur donnée.
Les p é r i o d e s d ' é c h a n t i l l o n n a g e s ( p r i n t a n i è r e s e t / o u
e s t i v a l e s ) dans les lacs de B i e n n e et des Q u a t r e - C a n t o n s
sont sans d o u t e à la base de la légère d i f f é r e n c e
p o n d é r a l e d'avec Les g a r d o n s du lac de N e u c h â t e l . En
e f f e t , co rum e nous Le v e r r o n s plus loin (figure 4 4 , p . 6 8 ) ,
la c r o i s s a n c e est i d e n t i q u e d a n s ces t r o i s Lacs et iL est
peu p r o b a b l e q u ' u n e d i f f é r e n c e r é e l l e m e n t s i g n i f i c a t i v e de
poi ds ex i s t e .
Eté comme hiver des p r é l è v e m e n t s ont été e f f e c t u é s dans Le
L o c l a t , la c o u r b e " l o n g u e u r t o t a l e / p o i d s " semble donc
r é a l i s t e . La p a u v r e t é en r e s s o u r c e s a l i m e n t a i r e s ( A R A G N O ,
c o m m u n i c a t i o n o r a l e , 1 9 8 6 ; S C H W E I Z E R S A L . , 1975) de ce
lac e x p l i q u e la faible c r o i s s a n c e des g a r d o n s Le p e u p l a n t .
Des c o u r b e s b a s é e s sur le p o i d s n o m i n a l ou sur Le p o i d s
f r a i s de p o i s s o n s c a p t u r é s à une p é r i o d e " i d e n t i q u e "
p e r m e t t r a i e n t de comparer les p o p u l a t i o n s issues des
d i f f é r e n t s m i l i e u x a v e c , plus d ' e f f i c a c i t é . On c o n s t a t e
c e p e n d a n t que cette r e l a t i o n met é g a l e m e n t en é v i d e n c e la
p r é s e n c e d ' é c o t y p e s , p a r t i c u l i è r e m e n t pour Les g a r d o n s du
IV.5. RESULTATS HYBRIDES
IV.5.1 . Proportion d'hybrides
Sur L'ensemble des pré lèvements de gardons effectués dans
le cadre de nos recherches, Le pourcentage d'hybrides de
Cyprinidés par rapport aux captures globales, varie
considérablement d'un lac à L'autre. La proportion
d'hybrides semble augmenter "proport ionne Llement" avec la
diminution de la dimension du Lac.
Tableau 7 : Proportion d'hybrides
total des captures
nombre de gardons
nombre d'hybri des
X
d'hybrides
surface du lac <km2)
Neuchâtel
1785
1764
21
1,1 8%
2 U , 6
Bi enne
2OA
200
4
1,96SS
39,3
Loclat
255
234
21
8,24%
0,046
4-Cantons
94
94
0
0¾
113,6
IV.5.2. Types d'hybrides
FATI0C188Z), NIK0LJUKIN(1948>, BERGC1949), DOTTRENS(1951),
SPILLMANN(1961), décrivent quantités d'hybridations
possibles entre Cyprinidés. L'anaLyse des critères
morphologiques dénombrables des hybrides issus de nos
propres échantillonnages permet d'émettre quelques
hypothèses quant à leurs origines.
15 individus (71,4X des hybrides correspondent à La
description de la brème de Buggenhagen résultat du
croi sèment intergénérique entre la brème franche (Abrami s
brama L.) et Le gardon (Ruti Lus rut ilus L . ) .
Tableau 8 : Caractères biométn'ques des brèmes de
Buggenhagen du Lac de Neuchâtel
No.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
-écai
LLAT
50
48
48
52
49
50
49
50
52
49
50
50
50
O7
Lles-TRANS
10/5
13/4
10/6
10/5
10/?
10/5
10/6
10/6
11/5
11/5
11/5
11/5
10/5
7
7
D
10 .
10
9
9
10
9
10
10
10
9
10
10
10
10
10
C
19
19
19
19
19
1
O1
7
O7
• >7
7
7
jeoi re
A
17
16
16
17
15
16
17
16
15
15
16
15
15
16
16
V
8
8
8
8
8
8
8
8
8
8
8
8
8
8
8
P
17
17
16
17
17
16
15
17
16
16
17
17
17
16
16
DENTS
5
5
5
6
6
5
5
6
5
6
5
6
6
5
5
GONADES
7
7
ovules
ovules
7
ovules
7
ovLf Les
7
7
7
laitance
• >ovu les
7
Les 6 hybrides restants (28,6X), dont les origines n'ont
pu être clairement définies, ont été cLassés en 3
catégories.
Tableau 9 : Caractères biométriques des hybrides
i ndéterminés du lac de Meuchgtel
Catégorie
1 I 5 1
Catégorie
1
2
3
4
7
7
?
7
Catégori e
1
7
A
B
C
9/5
i7
7
7
• >8
10
10
9
10
9
19
7
7
7
7
19
22
17
15
17
16
21
8
8
8
8
8
8
16
13
14
14
14
13
5
7
5
4
4
5
Laitance
7
7
?7
iC a t é g o r i e A : c a r a c t é r i s t i q u e s types de La brème b o r d e
-Ii ère (BLicca bjoerkna L . ) , à l'exception des dents p h a r y n g i e n n e s q u i , chez cette d e r n i è r e , p r é s e n t e n t une s t r u c t u r e sur 2 rangs ( 5 + 1 - 3 ) .
C a t é g o r i e B : out re Le nombre trop faible de rayons des n a g e o i r e s p e c t o r a l e s , les c a r a c t é r i s t i q u e s s y s t é m a t i q u e s des p o i s s o n s classés dans cette catégorie sont celles de La brème de B u g g e n h a g e n ; cependant le nombre de p a r a m è t r e s m a n q u a n t s est trop important pour garant ir l'ident i f i cat i o n .
C a t é g o r i e C : le manque de critères ainsi que la n a t u r e de ceux o b t e n u s , empêchent de définir les o r i g i n e s de cet i n d i v i d u .
Les 4 hybrides (100X) issus de ce lac présentent les c a r a c t é r i s t i q u e s de La brème de B u g g e n h a g e n .
T a b l e a u 10 : Caractères b i o m é t r i q u e s des brèmes de B u g g e n h a g e n du lac de Bi enne No.
1
2
3
4
é c a i l l e s -LLAT TRANS 46 10/4 49 10/5 52 10/6 53 ?D
9
10 109
C A V 19 16 8 19 14 8 19 16 8 19 16 8P
17 16 16 17 DENTS5
6
6
5
G O N A D E S -j 1 1 lai tanceSur les 21 h y b r i d e s , 16 (76,2Z) sont des brèmes de B u g g e n h a g e n .
Tableau 11 : C a r a c t è r e s bioniétriques des brèmes de B u g g e n h a g e n du lac du Loclat No.
1
Z3
4
5
6
7
8
9
10 11 12 13 14 15 16 -écai LLAT 49 47 47 48 48 47 47 49 49 45 49 47 497
7
7
l l e s -TRANS 11/5 11/5 11/5 11/4 10/4 12/5 10/5 10/4 10/4 9/4 10/47
7
11/4 9/57
D
109
9
10 10 10 109
9
9
10 109
9
9
10C
19 19 19 19 197
1 1•>
17
7
7
7
7
?
geoi reA
16 15 15 15 14 16 16 17 17 15 15 17 16 15 15 17V
8
8
6
8
8
8
8
8
8
8
8
8
8
8
8
8
P
16 16 14 14 16 17 16 14 16 15 17 16 16 15 15 16 DENTS5
4
5
5
7
7
6
6
4
5
7
5
5
5
7
5
G O N A D E S ovules7
7
7
7
7
lai tance7
7
7
7
7
7
7
7
7
Le solde des hybrides du LocLat (5 ind. = 2 3 , 8 X ) a pu être classé en 3 c a t é g o r i e s . T a b l e a u 12 : C a r a c t è r e s b i o m é t r i q u e s des h y b r i d e s i n d é t e r m i n é s du lac du Loclat C a t é g o r i e






![Figure 15 : Fréquence des n o m b r e s d'écaillés v e n t r o - l a t é r a l e s F K K f] II E N C L y 70 fiR< Mi 46 30 20 IU JL Jl 7 8 9 NOHBREI) 1 ECAILLESIlENTRU-LfIIEHALES](https://thumb-eu.123doks.com/thumbv2/123doknet/2209980.13518/28.600.67.365.69.867/figure-frequence-ecailles-ii-mi-nohbrei-ecaillesilentru-lfiiehales.webp)

