Études sur le développement des cryptes intestinales chez la souris
Texte intégral
Figure



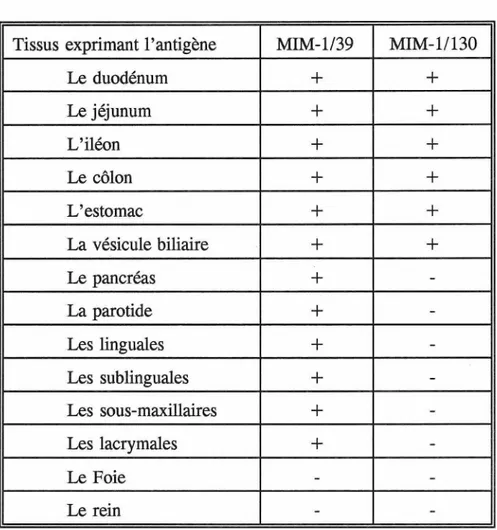
Documents relatifs
Nos résultats immunohistochimiques ont montré que les niveaux de YAP et TAZ (totaux) sont exprimés dans le mésenchyme de.. Toutefois, les niveaux de pYAP sont exprimés seulement
Chez le lapin nous montrons une réorganisation (formation d’amas) des marques d’histones spécifiques de l’hétérochromatine après l’EGA précédée par
Cette stratégie hybride apparait particulièrement efficace du fait que les récepteurs 5-HT 1A ciblés par la buspirone semblent également impliqués dans la récupération
Il ressort des résultats que, au cours de la période allant de cinq ans avant la perte d’emploi à neuf ans après, les travailleurs ayant fait des études et suivi
Après un rappel des connaissances actuelles relatives au développement de la thyroïde embryonnaire et fœtale humaine et aux greffes de tissus humains chez la souris nude, nous
Cette stratégie hybride apparait particulièrement efficace du fait que les récepteurs 5-HT 1A ciblés par la buspirone semblent également impliqués dans la
Les veines prélevées au membre supérieur semblent donner des résultats encourageants avec des taux de perméabilité et de sauvetage de membre assez comparables à ceux obtenus
Plus qu’une simple réorganisation municipale, la réforme proposée suggère des changements majeurs en matière de gouvernance, de développement économique et communautaire,
