Évaluation des impacts de l'illumination du pont
Jacques-Cartier sur l'avifaune et l'entomofaune
Mémoire
Shivia Nankoo
Maîtrise en génie des eaux - avec mémoire
Maître ès sciences (M. Sc.)
Évaluation des impacts de l’illumination du pont
Jacques-Cartier sur l’avifaune et l’entomofaune
Mémoire de maîtrise
Shivia Nankoo
Sous la direction de :
Résumé
Plusieurs lumières ont été installées sur le pont Jacques-Cartier le 17 mai 2017 afin de célébrer le 375ième anniversaire de la ville de Montréal et le 150ième anniversaire du Canada. Les lumières varient en couleur et intensité et sont illuminées toutes les nuits, pour une durée d’au moins 10 ans. L’Université Laval a répondu à l’appel d’offre lancée par l’organisation du pont afin de déterminer l’effet de ces lumières sur les animaux qui vivent à proximité du ou au pont, c’est-à-dire : l’hirondelle à front blanc (Petrochelidon pyrrhonota), le faucon pèlerin (Falco peregrinus), l’engoulevent d’Amérique (Chordeiles minor), le martinet ramoneur (Chaetura pelagica) et l’entomofaune (les insectes). Les observations, effectuées en 2017 et 2018, consistaient à détecter la présence, l’abondance, les activités de nidification et le taux d’activité des espèces d’intérêt. Les résultats ont montré que l’engoulevent d’Amérique et le martinet ramoneur n’ont pas été détectés aux alentours du pont et que l’illumination semble ne pas avoir d’impact sur leur présence. Les observations sur le faucon pèlerin ont montré que l’espèce n’était présente au pont qu’en 2018. Bien que des activités de chasse ont eu lieu, aucune nidification n’a été observée. Les observations sur les hirondelles ont montré que l’illumination a eu peu d’impacts en 2017. En 2018, l’espèce est plus abondante à proximité du pont, surtout dans la zone illuminée, et est plus active le soir lorsque les lumières sont allumées. Bien qu’aucun impact n’ait été observé en 2017, les observations ont montré une abondance plus élevée à proximité de la zone illuminée du pont. Une chaine alimentaire semble connecter les insectes, les hirondelles et les faucons puisque qu’une augmentation de l’un entraine une augmentation de l’autre. Les implications de ces observations sont expliquées et des recommandations sont émises sur la continuation du suivi.
Abstract
Several lights were installed on the Jacques-Cartier Bridge on May 17, 2017 to celebrate the 375th anniversary of the city of Montreal and the 150th anniversary of Canada. Lights vary in color and intensity and are illuminated every night, for a period of at least 10 years. Université Laval applied for the tender issued by the bridge’s organisation to determine the effect of the illumination on species that live near or on the bridge, specifically the cliff swallow (Petrochelidon pyrrhonota), the peregrine falcon (Falco peregrinus), the common nighthawk (Chordeiles minor), the chimney swift (Chaetura pelagica) and the insect fauna. The observations, conducted in 2017 and 2018, consisted of detecting the presence, abundance, nesting behavior and activity rate of the species of interest. The results reported that the common nighthawk and the chimney swift were not detected near the bridge and therefore seem to be unaffected by the illumination. Observations of the peregrine falcon indicated that the species was only seen in 2018. Although the bridge was used as a hunting ground, no nesting occurred. The observations on the cliff swallow showed the bridge had no impact on the species in 2017. However, in 2018, the species was more abundant, especially in the illuminated zone, and was more active in the evening when the lights were on. Although observations of the insect fauna showed no impact from the bridge in 2017, abundance of insects in 2018 was higher near the illuminated part of the bridge. A food chain seems to connect the insects, the swallows and the falcons since it is possible that an increase in one causes an increase in the next. The implications of these observations are explained, and recommendations are made on further follow-up.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des figures ... ix
Liste des tableaux ... xii
Liste des sigles ... xiii
Liste des acronymes ... xiv
Liste des abréviations ... xv
Remerciements ... xvi
Avant-propos ... xvii
Introduction ... 1
Présentation des espèces à l’étude ... 3
Le faucon pèlerin ... 3
L’hirondelle à front blanc ... 5
L’entomofaune ... 6
Problématique ... 7
Objectifs de la recherche ... 8
L’avifaune ... 8
L’entomofaune ... 9
Chapitre 1 « The multiple consequences of urban light pollution on birds » ... 10
1.1 Résumé ... 10
1.2 Abstract ... 10
1.3 Introduction ... 10
1.4 Methods ... 12
1.5 The relation between birds and light ... 12
1.5.1.1 Visible light ... 12
1.5.1.2 Ultraviolet light ... 13
1.5.2 Attraction to Light ... 13
1.5.2.1 Food source ... 13
1.5.2.2 Nesting sites ... 13
1.5.2.3 Protection against predators ... 14
1.5.2.4 Orientation ... 14
1.5.3 Repulsion from Light ... 14
1.6 How studies were performed ... 14
1.6.1 Experiments in the laboratory... 14
1.6.2 Experiments in the wild ... 15
1.6.3 Ethical standards ... 15
1.7 The consequences of artificial light on birds ... 15
1.7.1 Reproduction ... 15
1.7.1.1 First discovery of the consequence ... 15
1.7.1.2 Modification of the timing of reproduction ... 16
1.7.1.3 Modification of the sex apparatus ... 17
1.7.2 Sleep pattern ... 17
1.7.2.1 Modification of the timing and the duration of sleep ... 17
1.7.3 Migration ... 18
1.7.3.1 Attraction causing fatigue, disorientation and death ... 19
1.7.4 Patterns of activity ... 19
1.7.4.1 Modification of the timing of activity ... 19
1.7.5 Communication ... 20
1.7.6 Problem-solving skills ... 21
1.8 Other factors stronger than light ... 21
1.8.1 Noise ... 21
1.8.2 Cloud cover ... 22
1.8.3 Eye size ... 22
1.8.5 Pulsing light ... 23 1.9 Discussion ... 23 1.9.1 Future directions ... 27 1.10 Conclusion ... 28 1.11 Glossary ... 29 1.12 Literature Cited ... 29
Chapitre 2 « The Jacques Cartier bridge impacts nesting behavior and activity of the cliff swallow (Petrochelidon pyrrhonota) » ... 36
2.1 Résumé ... 36
2.2 Abstract ... 36
2.3 Introduction ... 37
2.4 Methods ... 38
2.4.1 Nest abundance and distribution ... 40
2.4.2 Nest activity ... 40
2.5 Results ... 41
2.5.1 Nest abundance and distribution ... 41
2.5.2 Nest activity ... 42
2.6 Discussion ... 47
2.7 Conclusion ... 50
2.8 Acknowledgements ... 51
2.9 Literature Cited ... 51
Chapitre 3 « The impact of the Jacques Cartier bridge illumination on the food chain: from insects to predators » ... 56 3.1 Résumé ... 56 3.2 Abstract ... 56 3.3 Introduction ... 57 3.4 Methods ... 58 3.4.1 Insects ... 58
3.4.2 Cliff swallows ... 60 3.4.3 Peregrine falcons ... 60 3.5 Results ... 62 3.5.1 Insects ... 62 3.5.2 Cliff swallows ... 64 3.5.3 Peregrine falcons ... 65 3.6 Discussion ... 66 3.7 Conclusion ... 71 3.8 Acknowledgements ... 71 3.9 Literature Cited ... 71
Chapitre 4 « Insect relative mortality rate of different types of urban lights » ... 75
4.1 Résumé ... 75 4.2 Abstract ... 75 4.3 Introduction ... 75 4.4 Methodology ... 77 4.5 Results ... 79 4.6 Discussion ... 82 4.7 Conclusion ... 84
4.8 Compliance with ethical standards ... 84
4.9 Literature Cited ... 84
Conclusion ... 88
Recommandations... 89
Récapitulatif des conclusions ... 90
Bibliographie ... 92
Annexe A Suivi du martinet ramoneur et de l’engoulevent d’Amérique ... 106
A.1 Introduction ... 106
A.1.2 L’engoulevent d’Amérique ... 108
A.2 Méthodologie... 109
A.2.1 Martinet ramoneur ... 109
A.2.2 Engoulevent d’Amérique ... 110
A.3 Résultats ... 111
A.3.1 Martinet ramoneur ... 111
A.3.2 Engoulevent d’Amérique ... 114
A.4 Discussion ... 114
A.5 Conclusion ... 116
A.6 Références ... 116
Annexe B Analyse visuelle et génétique de l’entomofaune ... 119
B.1 Introduction ... 119
B.2 Méthodologie... 119
B.3 Résultats ... 120
B.4 Discussion ... 124
B.5 Conclusion ... 126
Annexe C Protocole d’extraction d’ADN pour l’analyse génétique de l’entomofaune (Qiagen, 2006) ... 127
Annexe D Protocole de PCR (polymerase chain reaction, réaction de polymérase en chaine) pour l’analyse génétique de l’entomofaune ... 128
Annexe E Protocole de purification d’ADN pour l’analyse génétique de l’entomofaune (Zymo Research, 2001) ... 129
Annexe F Protocole de préparation des échantillons pour le séquençage SANGER pour l’analyse génétique de l’entomofaune ... 130
Annexe G Descriptions des ordres d’insectes identifiés visuellement ... 131
Liste des figures
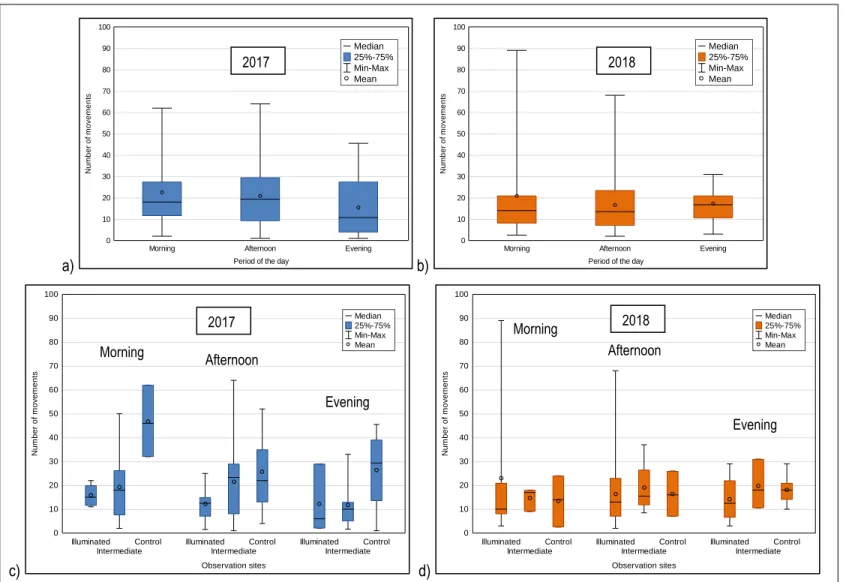
Figure 1. Le pont Jacques-Cartier illuminé (source : PJCCI). ... 1 Figure 2. Faucon pèlerin en vol (https://www.pronatura-ge.ch/Plan_d_action_Pelerin). ... 3 Figure 3. Hirondelle à front blanc en vol (https://www.allaboutbirds.org/guide/Cliff_Swallow/id). ... 5 Figure 4. Picture taken from NASA International Space Station (ISS) of the American continent at night (2017) (https://www.nasa.gov/image-feature/new-full-hemisphere-views-of-earth-at-night). ... 11 Figure 5. Picture of the section of the Jacques Cartier bridge illuminated at night (https://jacquescartierchamplain.ca/community-heritage/structures-and-projects/illumination-of-jacques-cartier-bridge/?lang=en). ... 38 Figure 6. Map of the Jacques Cartier bridge (Montréal, Canada) with its sections (2, 3, 4, 5, 6, 7 and 8) (sourced from Jacques Cartier and Champlain Bridges Incorporated (JCCBI)). ... 39 Figure 7. Location of the three (3) sites used to observe cliff swallows nests on the Jacques Cartier bridge. Locations were changed between the two (2) years to ensure that active nests were present in each zone. Orange circles represent the locations of the zones in 2017 and blue circles the locations of the zones in 2018. ... 40 Figure 8. Total abundance of cliff swallow nests on the Jacques Cartier bridge from 2012 to 2018 regardless of sections. The white columns represent the past data and the dark columns represent the data collected in this study. ... 41 Figure 9. Abundance of cliff swallow nests in a) section 6 and b) section 7 of the Jacques Cartier bridge. These sections are the closest to the bridge’s lights. No observations were made before 2016 in section 7. The blue column shows the data collected in the first year of this study and the orange column the data in the second year of this study. ... 42 Figure 10. Percentage of activity and abundance in parentheses of active cliff swallow nests in each section of the Jacques Cartier bridge for the two years of the study. ... 44 Figure 11. Abundance of cliff swallow movements to and from nests separated by the period of the day (morning, afternoon and evening) in a) 2017 and b) 2018, and number of cliff swallow movements to and from nests separated by the period of the day (morning, afternoon and evening) and by study sites (illuminated, intermediate and control) in c) 2017 and d) 2018. ... 46 Figure 12. Location of the three sites for insect capture in proximity to the Jacques Cartier bridge. The light gray circles represent the study sites in 2017 and the black ones in 2018. ... 59
Figure 13. Pictures of the two types of insect traps used in this experiment: a) Aerial trap (MegaView Science Co. SLAM Trap Large BT1005 and b) Emergence trap (MegaView Science Co. Amphibious Emergence Trap BT2008). ... 59 Figure 14. Location of the sections on the Jacques Cartier bridge. The peregrine falcon observation site is located in section 7 (sourced from Jacques Cartier and Champlain Bridges Incorporated (JCCBI)). ... 61 Figure 15. Insect abundance at the Jacques Cartier bridge in 2017 and 2018 per m3 for aerial traps at a) the control site, b) the intermediate site and c) the illuminated site, as well as insect abundance per m2 for aquatic traps at d) the control site, e) the intermediate site and f) the illuminated site. Loss and breakage of traps might cause discrepancies in the data. ... 63 Figure 16. Cliff swallow nest abundance from 2012 to 2018. The first year of the study (2017) is shown in yellow and the second year (2018) is shown in green. Results from 2012 to 2016 were taken from Services Environnementaux Faucons (SEF). ... 64 Figure 17. Percentage of activity and abundance in parentheses of cliff swallow nests at each section (2, 3, 4, 5, 6 and 7) of the Jacques Cartier bridge in a) 2017 and b) 2018. ... 65 Figure 18. Percentage occurrence of peregrine falcon behaviors observed in 2018 at the Jacques Cartier bridge. ... 66 Figure 19. Diagram of the food chain connecting insects, insectivorous birds (including the cliff swallow) and peregrine falcons (the top predator species). Full arrows show a direct impact and dotted arrows show an indirect impact. ... 70 Figure 20. Pictures of the four (4) types of urban lights used in this experiment: a) the custom-made LED spotlight b) the custom-made LED light strip c) the LED lamp post and d) the sodium lamp post and the custom-made wood supporting apparatus for e) the LED spotlight f) the LED light strip g) the LED lamp post h) the sodium lamp post. ... 78 Figure 21. Experimental protocol for the determination of the insect mortality rate in relation to the type of urban light used. ... 79 Figure 22. Relative mortality rate of the four (4) types of urban light sources. ... 81 Figure 23. Precipitation data in millimeters two (2) days before insect collection and percentage of moon light the night before insect collection in relation to insect abundance on each collection day (some precipitation data was not available). ... 82 Figure 24. Martinet ramoneur perché (http://www.thewoodthrushshop.com/news/2016/9/8/bird-bio-chimney-swifts-common-nighthawks). ... 106
Figure 25. Engoulevent d’Amérique au repos (https://www.canada.ca/fr/environnement-changement-
climatique/services/oiseaux-canada/celebrons-100-ans-conservation-internationale/on-est-aux-oiseaux/coralie-daigle-bruno-drolet.html). ... 108 Figure 26. Cheminées se trouvant sur le bâtiment Hélène-de-Champlain sur l’Île Ste-Hélène observées pour leur potentiel de servir de sites de nidification pour les martinets ramoneurs. ... 110 Figure 27. Carte des deux (2) routes suivies lors de l’observation de l’engoulevent d’Amérique aux alentours du pont Jacques-Cartier. Les chiffres montrent l’ordre dans lequel les stations ont été parcourues. ... 111 Figure 28. Cheminées propices au martinet ramoneur visitées lors des inventaires réalisés en 2017 (Pérez 2017). ... 112 Figure 29. Carte de la zone autour du pont Jacques-Cartier sans la présence d’engoulevents. ... 114 Figure 30. Photographie d’un insecte de l’ordre Trichoptera pour l’analyse visuelle. ... 119
Liste des tableaux
Tableau 1. Calendrier du cycle de nidification du faucon pèlerin. ... 4
Tableau 2. Calendrier du cycle de nidification de l’hirondelle à front blanc... 6
Tableau 3. Abundance of active and inactive nests on the whole Jacques Cartier bridge in 2012, 2017 and 2018. The blue line shows the data collected in the first year of this study and the orange line the data from the second year of this study. ... 43
Tableau 4. Insect mortality rates for each collection day, total mortality rate and mean mortality rate for the four (4) different types of urban lights. ... 80
Tableau 5. Impact de l’illumination sur chaque groupe animal observé au pont Jacques-Cartier en 2017 et 2018. ... 88
Tableau 6. Calendrier du cycle de nidification de l’engoulevent d’Amérique. ... 107
Tableau 7. Calendrier du cycle de nidification de l’engoulevent d’Amérique. ... 108
Tableau 8. Sites possibles de nidification du martinet ramoneur observés en 2017 et 2018. ... 113
Tableau 9. Identification et dénombrement des populations d’insectes lors de la collecte d’échantillonnage in-situ d’août 2017. ... 121
Tableau 10. Identification par séquençage SANGER des insectes récoltés aux sites témoin, intermédiaire et illuminé dans la zone du pont Jacques-Cartier en août 2017 représentant les proportions du nombre d’individus les plus élevées. ... 123
Tableau 11. Description des caractéristiques et du cycle de vie des différents ordres d’insectes identifiés visuellement au pont Jacques-Cartier en août 2017. ... 131
Tableau 12. Résultats complets des identifications par séquençage SANGER d’insectes récoltés aux sites témoin, intermédiaire et illuminé dans la zone du pont Jacques-Cartier en août 2017. ... 137
Liste des sigles
ADN : Acide désoxyribonucléique
CRFPQ : Comité de Rétablissement du Faucon Pèlerin au Québec DDT : Dichlorodiphényltrichloroéthane
DNA : Deoxyribonucleic acid
ÉEE : Évaluation des effets environnementaux
ICOAN : Initiative de conservation des oiseaux de l'Amérique du Nord JCCBI : Jacques Cartier and Champlain Bridges Incorporated
LED : Light Emitting Diode LEP : Loi sur les Espèces en Péril
MFFP : Ministère de la Forêt, de la Faune et des Parcs MRNF : Ministère des Ressources Naturelles et de la Faune NCBI : National Center for Biotechnology Information
PCR : Réaction en chaîne par polymérase, Polymerase chain reaction PJCCI : Les Ponts Jacques Cartier et Champlain Incorporée
SEF : Services Environnementaux Faucons UV : Ultraviolet
Liste des acronymes
ALAN : Artificial Light at Night
BLAST: Basic Local Alignment Search Tool CHUL : Centre Hospitalier de l’Université Laval
COSEPAC : Comité sur la situation des espèces en péril au Canada COSEWIC : Committee on the Status of Endangered Wildlife in Canada DEL : Diode électroluminescente
Liste des abréviations
m : mètres mg : milligrammes ml : millilitres
rpm : Rotation par minute μL : microlitre
Remerciements
J’aimerais remercier la professeure Rosa Galvez pour m’avoir permis de réaliser cette recherche et de m’avoir encouragé durant la totalité de mon parcours.
Je suis reconnaissante pour tout le support qui m’a été donné par Les Ponts Jacques Cartier et Champlain Incorporé pour la réalisation de cette recherche, l’accès aux sites et l’aide technique pour l’entreposage et l’installation de l’équipement nécessaire. Je souhaite remercier Emanuel Chênevert, Soufyane Loubar, Sylvie Boulanger et Stéphane Vaquette.
Je veux remercier l’organisation Services Environnementaux Faucons pour leur aide précieuse pour l’observation des espèces aviaires et les techniques de prises de données, notamment Marilou G. Skelling, Marie-Line Fiola et Luana Graham-Sauvé.
J’aimerais aussi remercier le personnel du Centre Hospitalier de l’Université Laval pour leur support et leur conseils précieux pour l’analyse génétique des insectes, plus spécialement Jacques Corbeil, Nancy Boucher, Pier-Luc Plante et Francis Brière.
J’aimerais aussi remercier les entomologistes de l’Université de Milan pour leur aide lors de la récolte d’insectes et l’analyse des données, notamment Matteo Montagna et Giuseppe Lozzia.
Je veux exprimer ma gratitude envers le professeur Maximiliano Cledon ainsi que tous les stagiaires qui ont participé aux collectes de données et aux observations sur le terrain durant la totalité de cette étude : Arnaud Benoît-Pépin, Jasmine Duchesneau, Marie Bourgault et Justine Auquier.
Je suis reconnaissante à Michel Bisping pour m’avoir permis l’accès aux laboratoires de l’Université et m’avoir aidé pour l’utilisation de l’équipement nécessaire à l’analyse des échantillons d’insectes. Je veux aussi remercier Martin Lapointe, Denis Jobin et Vincent Aeschlimann pour leur aide dans l’organisation et l’installation de l’équipement à l’Université pour une majeure partie de mon projet.
Finalement, je veux remercier Sébastien Raymond et Stéphane Boudreau d’avoir accepté de faire partie du jury pour l’évaluation de mon mémoire et de mon séminaire, et pour leurs nombreux conseils par rapport aux analyses et conclusions de mon projet.
Avant-propos
Ce mémoire intègre quatre (4) articles scientifiques :
• « The multiple consequences of urban light pollution on birds » au chapitre 1: o Date de soumission: 5 mars 2019
o Date d’acceptation: NA o Date de publication: NA
o Statut d’auteur: Je suis l’auteur principal. o Rôle : J’ai écrit, corrigé et soumis l’article.
o Coauteurs : Sébastien Raymond, Maximiliano Cledon et Rosa Galvez.
• « The Jacques Cartier bridge impacts nesting behavior and activity of the cliff swallow (Petrochelidon pyrrhonota) » au chapitre 2:
o Date de soumission: 26 avril 2019 o Date d’acceptation: NA
o Date de publication: NA
o Statut d’auteur: Je suis l’auteur principal. o Rôle : J’ai écrit, corrigé et soumis l’article. o Coauteurs : Sébastien Raymond et Rosa Galvez.
• « The impact of the Jacques Cartier bridge illumination on the food chain: from insects to predators » au chapitre 3:
o Date de soumission: 29 mars 2019 o Date d’acceptation: 2 juillet 2019 o Date de publication: 20 octobre 2019 o Statut d’auteur: Je suis l’auteur principal. o Rôle : J’ai écrit, corrigé et soumis l’article. o Coauteurs : Sébastien Raymond et Rosa Galvez.
• « Insect relative mortality rate of different types of urban lights » au chapitre 4: o Date de soumission: 28 mars 2019
o Date d’acceptation: NA o Date de publication: NA
o Statut d’auteur: Je suis l’auteur principal. o Rôle : J’ai écrit, corrigé et soumis l’article. o Coauteurs : Sébastien Raymond et Rosa Galvez.
Les sections qui ne sont pas des articles scientifiques ont été extraits du rapport présenté à Les Ponts Jacques Cartier et Champlain Incorporé en décembre 2018 (Nankoo et al. 2018). Certains volets de la recherche effectuée sont présentés dans les annexes : le suivi de l’engoulevent d’Amérique et du martinet ramoneur en annexe A et l’analyse visuelle et génétique des insectes en annexe B.
Introduction
Dans le cadre des festivités du 375e anniversaire de Montréal et du 150e anniversaire du Canada, le pont Jacques-Cartier est illuminé depuis le 17 mai 2017, et le restera pour une période d’au moins dix ans. Cette mise en lumière met en valeur un repère historique et architectural majeur du paysage urbain de la région montréalaise, contribuant à son rayonnement touristique international. Le promoteur du projet est Les Ponts Jacques Cartier et Champlain Incorporée (PJCCI), société d’État mandataire mère, propriétaire du pont Jacques-Cartier.
Figure 1. Le pont Jacques-Cartier illuminé (source : PJCCI).
L’illumination du pont se trouve à la section 7 (qui inclue les piles 23 à 26) située entre Montréal et l’Île Ste-Hélène. Les zones éclairées incluent les tourelles, les piles, l’intérieur et l’extérieur de la structure du pont (Figure 1). Les luminaires utilisés comprennent 2 000 réglettes et 400 projecteurs. Ces derniers peuvent être ajustés afin d’illuminer un endroit désiré. Tous les luminaires sont de type LED. Les projecteurs sur la structure sont de type RGB; ils peuvent donc afficher n’importe quelle couleur, alors que ceux sur les piles sont blancs (Raymond et al. 2017, PJCCI 2017a, PJCCI 2017b).
Un rapport émis par le consortium WSP Canada Inc. (WSP) / Aecom Consultants Inc. (Aecom) en 2016 a présenté l’évaluation des effets environnementaux (ÉEE) du projet. Selon les conclusions du rapport, l’effet de l’illumination est ‘’non important’’. Cependant, plusieurs recommandations ont été émises afin de limiter les impacts de l’illumination. En ce qui concerne l’avifaune, une des recommandations mentionne : « effectuer un suivi sur les deux (2) espèces aviaires (hirondelle à front blanc et faucon pèlerin) utilisant la structure et ses environs une fois l’illumination entamée afin de quantifier les effets du projet sur les espèces ». C’est dans ce
cadre que la professeure Rosa Galvez de l’Université Laval a été mandatée par PJCCI pour réaliser le suivi environnemental de l’illumination du pont Jacques-Cartier sur l’avifaune, en particulier le faucon pèlerin, le martinet ramoneur, l’engoulevent d’Amérique et l’hirondelle à front blanc, et l’entomofaune (insectes).
La présence de lumière artificielle est reconnue pour avoir un impact sur le mode de vie des animaux vivant à proximité. Chez les oiseaux, la lumière peut encourager la nidification, la protection contre les prédateurs et la recherche de nourriture (Robertson et al. 2010, Tets et al. 1969, Rao et Koli 2017, De Molenaar et al. 2000, Gorenzel et Salmon 1995). Ces aspects peuvent être positifs pour les individus puisqu’ils aident leur survie. Cependant, plusieurs impacts négatifs ont aussi été observés vis-à-vis l’exposition à la lumière urbaine. Celle-ci peut causer une modification de la période adéquate pour la reproduction en accélérant le processus de développement des organes sexuels et la ponte des œufs (Dominoni et Partecke 2015, Dominoni et al. 2015, Dominoni et al. 2013, Kempenaers et al. 2010). Le sommeil des oiseaux peut aussi être négativement affecté par la lumière artificielle. Plusieurs recherches ont trouvé que le moment auquel les oiseaux vont dormir est décalé, c’est-à-dire que les oiseaux s’endorment plus tard dans la nuit et peuvent se lever plus tard le matin. De plus, la quantité de sommeil (en heures) peut diminuer (Sun et al. 2017, Raap et al. 2017, Yorzinski et al. 2015). La migration des oiseaux peut aussi être modifié puisque les oiseaux peuvent être attirés par la lumière artificielle sur les immeubles et dévier de leur trajectoire initiale (Poot et al. 2008, Cochran et al. 2004, Horváth et al. 2009, Longcore et Rich 2004, Stone 2018, Kociolek et al. 2011, Squires et Hanson 1918, Merkel et Johansen 2011, Avery et al. 1976, Jones et Francis 2003). Les activités de chasse peuvent aussi avoir lieu plus tard dans la soirée, même pour les oiseaux diurnes, lorsque de la lumière artificielle est présente (Jong et al. 2016, Lustick 1973, Russ et al. 2015). De plus, elle peut encourager les oiseaux à chanter plus tôt le matin (Miller 2006, Silva et al. 2016, Kempenaers et al. 2010). Tous ces aspects peuvent jouer un rôle sur la survie des espèces aviaires puisqu’ils affectent leur reproduction, leur recherche de nourriture et leur migration. Le premier chapitre de ce mémoire présente ces informations en détail.
La lumière artificielle peut aussi avoir un impact sur les insectes. En effet, plusieurs recherches effectuées ont démontré que la présence de lumière attire les insectes, les rendant ainsi vulnérables à la prédation (par les oiseaux, par exemple), et diminuant leur chance de survie puisqu’ils risquent la mort par brûlure ou épuisement (McDonnell et al. 2015, Justice and Justice 2016, Pawson and Bader 2014, Egri et al. 2017). Une diminution importante d’insectes dans l’environnement est susceptible d’avoir des conséquences désastreuses pour tous les animaux qui en font leur source de nourriture primaire.
Ces informations montrent l’importance de la recherche effectuée dans ce mémoire afin d’évaluer l’impact de la présence des nouvelles lumières sur le pont et d’assurer la préservation des espèces animales vivant à proximité.
Présentation des espèces à l’étude
Les sections suivantes comprennent des informations sur les quatre espèces de l’avifaune étudiées dans le cadre de cette recherche, soit le faucon pèlerin, le martinet ramoneur, l’engoulevent d’Amérique et l’hirondelle à front blanc. Elles ont été choisies dû à leur présence antérieure ou récente à proximité du pont et à leur statut précaire selon différents organismes responsables de la protection des animaux. Les informations relatives au martinet ramoneur et à l’engoulevent d’Amérique sont présentées à l’annexe A.
Le faucon pèlerin
Le faucon pèlerin (Falco peregrinus) est un des oiseaux les plus étudiés au monde (Davis 2008).
Le faucon arbore un ventre rayé et son dos est gris foncé, presque bleu. La disposition des plumes au niveau de sa tête donne l’apparence d’un casque noir avec des longs favoris de la même couleur, alors que son cou est blanc (figure 2). Les individus juvéniles n’ont pas le cou blanc, mais sont striés de la base de la tête au ventre. Leur dos a une coloration brunâtre (Brûlotte 2000, Davis 2008, Vuilleumier 2009).
Figure 2. Faucon pèlerin en vol (https://www.pronatura-ge.ch/Plan_d_action_Pelerin).
Le faucon pèlerin est une espèce d’oiseau migrateur qui niche normalement le long des côtes dans les montagnes. En milieux urbains, on le retrouve sur les hauts édifices et les ponts (COSEPAC 2007, Hémisphère 2011, Brûlotte 2000). Il préfère nicher dans les grands espaces ouverts afin de faciliter sa capacité à chasser ses proies (Davis 2008).
La présence de l’espèce est notée sur tous les continents sauf l'Antarctique (Davis 2008). En Amérique, son territoire comprend le sud du Canada, les États-Unis, le Mexique ainsi qu’une partie de l’Amérique Centrale et du Sud (Cooperation 2000, Davis 2008). Au Québec, des faucons ont été observés dans la région de Montréal et au Nord de la province (Brûlotte 2000). Ils migrent jusqu’à la côte Est des États-Unis ou même jusqu’en Amérique du Sud alors qu’ils suivent les oiseaux migrateurs qui leur servent de proies (CRFPQ 2002).
En temps que prédateurs généralistes, ils se nourrissent des espèces disponibles à l’endroit où ils se trouvent. Leur régime est surtout constitué d’oiseaux, mais peut parfois compter de petits mammifères. Ils sont souvent vu en chasse au-dessus des plans d’eau où ils se nourrissent d’oiseaux marins. Ils chassent en vol, frappant leur proie du dessus pour l’assommer (Davis 2008).
Le vol du faucon se caractérise par des courts, mais puissants battements d’ailes. Ses ailes sont longues et pointues alors que sa queue est carrée (Brûlotte 2000). Il plane souvent. De loin, il peut être confondu pour un corbeau ou un aigle (Davis 2008). Le faucon a un comportement diurne (COSEPAC 2007).
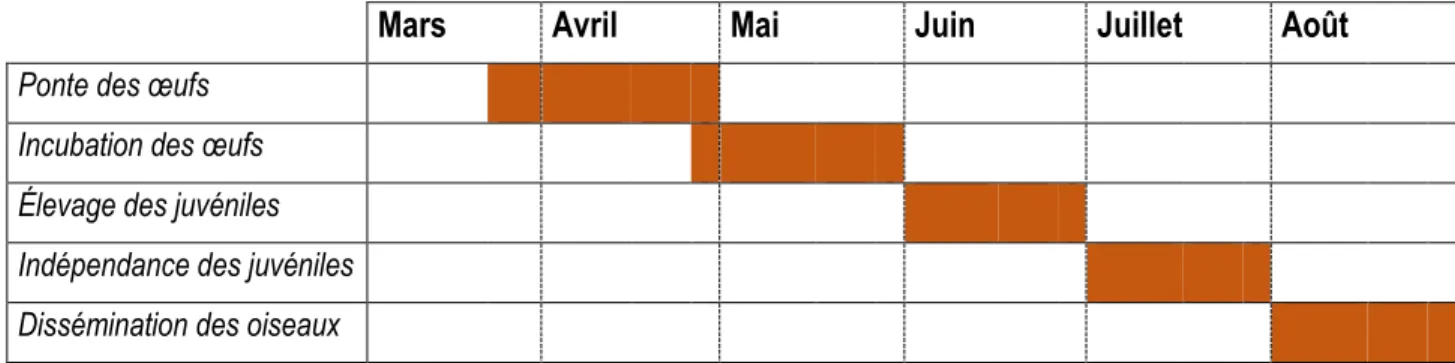
Les faucons nichent dans des dépressions dans la surface des falaises ou des structures urbaines (Davis 2008). Ils pondent 3 à 4 œufs durant l’été et la couvaison dure 28 à 35 jours (Brûlotte 2000, Davis 2008). La période de nidification est séparée en plusieurs parties, tel qu’illustré dans le tableau 1 (Molina et al. 2016).
Tableau 1. Calendrier du cycle de nidification du faucon pèlerin.
Mars
Avril
Mai
Juin
Juillet
Août
Ponte des œufs Incubation des œufs Élevage des juvéniles Indépendance des juvéniles Dissémination des oiseaux
Les faucons ont tendance à réutiliser les mêmes endroits pour nicher pendant plusieurs années (COSEPAC 2007, Davis 2008). Les juvéniles qui reviennent ont tendance à nicher à l’endroit où ils sont nés (Bird 1997). C’est pour cette raison qu’il est important de protéger les nids présents sur les structures anthropiques. Les faucons se sont très bien adaptés aux milieux urbains. Les hauts bâtiments sont des endroits de nidification pour l’espèce. De plus, la forte présence d’oiseaux urbains comme les pigeons assurent leur alimentation. Cependant, plusieurs dangers sont présents dans les villes : les collisions avec les automobiles, les grandes fenêtres qui reflètent la lumière et la présence d’eau sous les ponts où ils nichent augmentent le risque de mortalité des juvéniles (Davis 2008). Plusieurs nids artificiels ont été installés dans la région de Montréal pour répondre à la chute de la population des faucons (Hémisphère, 2011).
Le nombre de faucons a grandement diminué depuis les années 1950. L’insecticide DDT utilisé en grande quantité par les agriculteurs serait l’une des causes de la baisse de leurs populations. L’exposition au pesticide a pour effet secondaire de fragiliser la coquille des œufs des faucons. Les parents brisaient alors leurs œufs de façon accidentelle lors de la couvaison, réduisant ainsi la capacité de reproduction de l’espèce (Vuilleumier 2009). Le DDT a depuis été banni. Des efforts de conservation ont été mis en œuvre et le nombre de faucons
a augmenté (Davis 2008). Entres autres, des individus ont été élevés en captivité et relâchés dans la nature afin d’augmenter la population sauvage (Vuilleumier 2009, CRFPQ 2002). Les populations de cette espèce seraient maintenant stables (Davis 2008, Vuilleumier 2009).
Le faucon pèlerin est désigné vulnérable au Québec par le Ministère de la Forêt, de la Faune et des Parcs (MFFP) et est donc protégé par la Loi sur les espèces menacées et vulnérables. De plus, étant un oiseau de proie, il est protégé par la Loi sur la conservation et la mise en valeur de la faune. Cette loi interdit, en tout temps, de chasser, de piéger ou d’avoir en sa possession un oiseau de proie sauvage (vivant ou mort). Le faucon pèlerin est considéré menacé par le Comité sur la situation des espèces en péril du Canada (Cooperation 2000, COSEPAC 2007, Gauthier et al. 2007). Il est aussi considéré vulnérable par le ministère des Ressources Naturelles et de la faune au Québec (MRNF 2008-2009). Le faucon pèlerin possède aussi le statut d’espèce préoccupante au Canada en vertu de la Loi sur les espèces en péril (LEP).
L’hirondelle à front blanc
L’hirondelle à front blanc (Hirundo pyrrhonota) est une espèce d’oiseau migrateur. Elle peut être trouvée partout en Amérique du Nord (Vuilleumier 2009). Elle y passe l’été pour nicher et élever sa progéniture, et migre ensuite en Amérique du Sud pour y passer l’hiver (Québec Oiseaux 2017a).
Figure 3. Hirondelle à front blanc en vol (https://www.allaboutbirds.org/guide/Cliff_Swallow/id).
L’hirondelle a un dos bleu et noir, un ventre blanc, une gorge brune et une tache blanche sur le front, lui valant ainsi son nom (figure 3) (Brûlotte 2000).
L’hirondelle est insectivore et se nourrit en vol, souvent au-dessus des plans d’eau (Québec Oiseaux 2017a). C’est de cette façon qu’elle est majoritairement détectée.
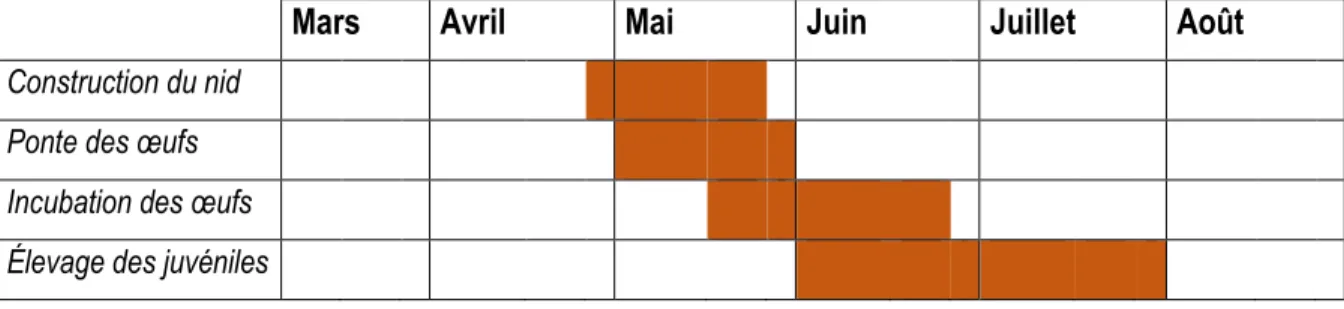
Le nid de l’hirondelle est fait de boules de boue que l’hirondelle récolte et agglutine pour construire un nid en forme de gourde (Hémisphère 2011). En milieu urbain, les hirondelles utilisent les structures anthropiques de façon à minimiser la quantité de boue à utiliser. Elles forment ainsi leurs nids dans les coins des structures formant un angle. Les nids sont souvent regroupés les uns à côté des autres afin de minimiser le temps et l’énergie dépensés à la construction, ce qui est avantageux pour l’espèce. Les hirondelles sont aussi susceptibles de réutiliser les nids qui ont été construits les années précédentes. Cependant, des parasites peuvent être présents dans les nids (Molina et al. 2012). À chaque saison de reproduction, l’hirondelle pond 4 à 5 œufs (Brûlotte 2000). L’oiseau devient territorial pendant sa période de nidification et ne pond généralement qu’une fois dans la saison. La période de nidification de l’hirondelle est présentée au tableau 2 (Molina et al. 2016).
Tableau 2. Calendrier du cycle de nidification de l’hirondelle à front blanc.
Mars
Avril
Mai
Juin
Juillet
Août
Construction du nid Ponte des œufs Incubation des œufs Élevage des juvéniles
Cette espèce étant une espèce migratrice, elle est donc protégée par la Loi de 1994 sur la convention concernant les oiseaux migrateurs. Il est donc illégal de causer des dommages à l’espèce, même de façon accidentelle. De plus, il est interdit de déranger ou détruire le nid et les œufs de ces espèces, et ce, même hors de la période de nidification. L’hirondelle n’a pas de statut selon la Loi sur les espèces en péril au Canada (2002) ou la Loi sur les espèces menacées et vulnérables au Québec (2017).
L’entomofaune
Les insectes forment le plus grand groupe d'organismes vivants au monde en terme du nombre d'espèces et de biomasse. Dans le monde, 850 000 espèces sont identifiées, mais ce nombre ne cesse d'augmenter. Il y a 13 000 espèces d’insectes au Québec, mais le compte total serait autour de 25 000 en estimant celles qui n'ont pas encore été identifiées (Bélanger 1991).
Les insectes se nourrissent surtout de plantes (racines, feuilles, jeunes pousses, fleurs, etc.). Ils se nourrissent en grande partie durant leur stade de développement. Le stade adulte est principalement associé avec la reproduction et la ponte des œufs (Arnett 2000).
La maturation des insectes se fait durant le printemps ou l'été. Ils hivernent pendant l'hiver. La plupart des espèces d'insectes ne migrent pas (Arnett 2000).
Les insectes peuvent vivre dans une variété d'environnements différents: des forêts jusqu'au désert. Peu vivent dans l'océan (Arnett 2000, Bélanger 1991). Au Québec, les forêts boréales, les marécages et les lacs sont des exemples d'écosystèmes favorables aux insectes (Bélanger 1991). Une grande abondance d'insectes est souvent associée aux milieux urbains due à l'importante quantité de lumière émise dans les villes. Cette lumière attire les insectes (Robinson 2005).
Les différents stades juvéniles des insectes sont difficiles à trouver en nature; les espèces sont souvent observées à leur stade adulte (Arnett 2000).
La fragmentation et la destruction des habitats (notamment des milieux humides), l'utilisation excessive de pesticides et la pollution sont des causes du déclin des insectes (Arnett 2000). Par conséquent, plusieurs insectes sont en danger d'extinction, comme certains coléoptères, lépidoptères et odonates (Bélanger 1991). La plupart des informations connues au sujet des insectes traitent de leur activité au stade adulte. Les stades de développement sont beaucoup moins étudiés (Bélanger 1991). Il n'y a aucune donnée sur les espèces présentes dans la région du pont Jacques-Cartier ou même dans la région de Montréal. La grande diversité d’espèces et l’absence de suivis rend difficile une inventorisation historique des espèces. Aucune étude de ce genre n’a été effectuée au pont Jacques-Cartier.
Problématique
Les impacts de l’illumination du pont Jacques-Cartier sur les espèces à l’étude ne sont pas connus. Les premières observations sur l’effet potentiel de l’illumination ont été mentionnées dans le rapport de WSP (2016). À première vue, l’impact potentiel de l’illumination sur le faucon, une espèce territoriale et diurne, apparait faible ou limité. De plus, selon WSP (2016), le faucon pèlerin n’est pas directement touché par l’illumination du pont Jacques-Cartier puisqu’il ne niche plus au pont depuis plusieurs années.
En ce qui concerne le martinet, il est diurne et peu présent autour du pont, tout comme le faucon. Ainsi, l’impact de la lumière est potentiellement faible voir nul sur la nidification de cet oiseau.
Contrairement aux autres espèces à l’étude, l’engoulevent d’Amérique est nocturne. Cependant, selon certaines données historiques, il est peu abondant autour du pont. L’illumination pourrait potentiellement avoir un impact sur son activité.
WSP (2016) affirme aussi que l’hirondelle ne sera pas affectée par l’illumination puisqu’aucun nid n’avait été détecté dans la section de l’illumination. Cependant, des observations récentes ont permis de montrer que des hirondelles nichent effectivement sur la section illuminée du pont, ce qui pourrait avoir un impact sur son comportement.
Finalement, chez les insectes, l’illumination du pont augmentera potentiellement les effets environnementaux liés à la lumière dû à l’attraction des insectes vers les sources lumineuses.
Une des difficultés liées à l’évaluation de l’impact de l’illumination concerne la disponibilité des données antérieures pour comparer les résultats obtenus. En effet, les suivis sont rares en ce qui a trait aux espèces aviaires et sont inexistantes pour les insectes. Un suivi temporel pour la comparaison avant-après l’illumination est donc difficile. Ainsi, cette étude se concentrera sur une comparaison temporelle restreinte et une comparaison spatiale (entre différents sites aux alentours du pont).
Objectifs de la recherche
Afin d’évaluer l’impact de l’illumination du pont Jacques-Cartier sur la faune, les quatre (4) espèces aviaires présentées ont été étudiées, à savoir : le faucon pèlerin, le martinet ramoneur, l’engoulevent d’Amérique et l’hirondelle à front blanc. Le suivi des insectes a également été réalisé dans le cadre de cette étude. Les observations ont été compilées et analysées pour déterminer des tendances à l’aide des deux (2) années de suivi (2017 et 2018) et des données antérieures disponibles. Les objectifs liés à chaque groupe étudié dépendent de la présence antérieure de l’espèce et des impacts potentiels de l’illumination.
L’avifaune
Concernant le faucon pèlerin, les buts spécifiques de cette recherche étaient de :
• Déterminer s’il y a présence/absence de l’espèce à proximité du pont Jacques-Cartier;
• Déterminer les comportements de l’espèce en relation avec le pont, par exemple la chasse ou la nidification.
Pour ce qui est du martinet ramoneur, les buts spécifiques de cette recherche étaient de : • Déterminer s’il y a présence/absence de l’espèce à proximité du pont Jacques-Cartier; • En cas de présence, déterminer si la nidification est présente.
Pour l’engoulevent d’Amérique, les buts spécifiques de cette recherche étaient de :
• Déterminer s’il y a présence/absence de l’espèce à proximité du pont Jacques-Cartier; • En cas de présence, déterminer si la nidification est présente.
Concernant l’hirondelle à front blanc, les buts spécifiques de cette recherche étaient de : • Déterminer s’il y a présence/absence de l’espèce à proximité du pont Jacques-Cartier;
• Mesurer l’activité de cette espèce, c’est-à-dire l’occupation du nid et le nombre de mouvements aux nids;
• Évaluer l’évolution de la répartition spatio-temporelle des nids;
• Évaluer s’il y a des changements d’activités en fonction de la répartition spatio-temporelle.
L’entomofaune
Concernant les insectes, les objectifs de cette recherche étaient de :
• Déterminer l’abondance et la biomasse des insectes pour différents sites répartis spatialement selon leur distance par rapport au pont Jacques-Cartier;
• Déterminer les ordres présents et leurs abondances selon les sites;
• Évaluer le taux de mortalité spécifique causé par les différents types de lumières disponibles au pont. Ce rapport présente les résultats obtenus à la suite des deux (2) années de suivi effectuées sur l’illumination du pont Jacques-Cartier. Il comprend les conclusions produites sur l’effet de l’illumination sur les espèces de l’avifaune et les insectes étudiés.
Chapitre 1 « The multiple consequences of urban
light pollution on birds »
1.1 Résumé
L’augmentation du territoire couvert par l’homme entraîne l’accroissement de l’éclairage artificiel utilisé pendant la nuit. Bien que la lumière nocturne soit essentielle au mode de vie de l'homme, elle a des conséquences importantes sur les animaux vivant en milieu urbain. La présente revue examine les nombreuses recherches publiées à ce jour sur ces conséquences, plus spécifiquement les conséquences pour l’avifaune. La littérature citée a été trouvée en utilisant des bases de données de journaux et en analysant chaque article de manière approfondie. Les informations trouvées dans la littérature sont organisées pour conclure que l’éclairage artificiel a un impact sur les nombreux comportements des oiseaux, notamment la reproduction, le sommeil, la migration, la communication, la quête de nourriture et la résolution de problèmes. Des articles additionnels sont intégrés dans cette revue qui rejettent les hypothèses précédentes, suggérant plutôt que la lumière a un impact secondaire sur les comportements d’oiseaux. Les effets de la lumière artificielle sont ensuite reliés à des conséquences écologiques plus importantes à long terme pour les populations d'oiseaux en termes de survie des populations. Des domaines de recherche futurs sont également explorés.
1.2 Abstract
The expansion of the territory covered by humans brings with it the increase of the extent of artificial lighting used at night. Although light at night is essential to human lifestyle, it is known to have important consequences on animals living within urban environments. The present review looks at the multiple research published up to date on these consequences specifically affecting birds. Literature was found using journal databases and analysing each article thoroughly. The information found in the literature is neatly arranged in a way to conclude that artificial lighting has impacts on many bird behaviors including reproduction, sleep, migration, communication, foraging and problem-solving skills. The research supporting these claims is then rebuffed by further research that show light as a secondary impactor for these behaviors. The effects of artificial light are then related to more important ecological consequences on the longer term for bird populations in terms of their individual fitness and population survival. Future areas of research are also explored.
1.3 Introduction
Throughout history, humans have increased the modification of the environment they use. With rapid population growth, the expansion of these modified areas was also increased. Moreover, the area covered in artificial lights
affects wider stretches of territory (Longcore and Rich 2004) and can be visible from space (Figure 4). Light at night is very useful to humans. It encourages activity later in the evening and is necessary for urban life (Kyba et al. 2015). However, the level of light and noise is the highest it’s ever been in human history (Swaddle et al. 2015). Moreover, light emitted by an urban area can be scattered into the atmosphere increasing the overall light level (Kyba et al. 2015). Around 83% of the population in the world live where the ambient light conditions at night are about 8% higher than natural nighttime conditions (Falchi et al. 2016).
Figure 4. Picture taken from NASA International Space Station (ISS) of the American continent at night (2017) (https://www.nasa.gov/image-feature/new-full-hemisphere-views-of-earth-at-night).
Light can however be harmful to other organisms that live in the urban environments. Although humans depend on artificial lighting to live their lives, animals depend on natural light cycles for many of their physiological behaviors (Navara et al. 2007). This observation has slowly been gaining interest in the field and more research needs to be done to properly assess how artificial light can affect animals living in or close to urban areas (Longcore and Rich 2004, Swaisgood 2007, Lyytimäki et al. 2012).
Some species have been able to adapt to urban environments and make them their home. These species include birds (most songbirds) and small to medium mammals like squirrel (Ditchkoff et al. 2006). In an experiment on European blackbirds, it was found that urban birds of these species were more successful in solving small tasks (they solved the task faster) than birds of the same species in forest habitats (Preiszner et al. 2017). These species can survive in anthropogenic areas, often have access to large quantities of food and predation is less of an issue. However, urban landscapes are not without consequences for them (Ditchkoff et al. 2006). In fact, pollution, light and methods of transport (for example cars) can cause disease, infections, injuries, and death (Kociolek et al. 2011). These dangers can substantially modify animal behaviour like the timing of their activities,
their diet, their reproductive success and their survival (Ditchkoff et al. 2006). Even though cities can be dangerous for animals, it was found that they are a positive factor in determining the density and species richness of birds in an area and their population stability in between seasons (Ciach and Fröhlich 2017). In fact, animals can have multiple reasons to be attracted to cities.
As taken from Longcore and Rich (2004), the type of light examined in the present article is ecological light pollution. It is the artificial light that ‘’alters [the] natural light regimes in terrestrial and aquatic ecosystems’’. This is the type of light that will potentially affect avian populations. In this article, the effect of light pollution on birds will be assessed by investigating the information found throughout multiple articles published on this subject.
1.4 Methods
The articles used in this review were found using the database from the Université Laval library website to get access to the PDF version of the articles. Google Scholar was also used in some instances. Search terms almost always included almost always the term ‘’bird’’ combined with either ‘’artificial light’’, ‘’light’’ and ‘’light pollution’’. Other articles were extracted from the cited references of already found articles.
1.5 The relation between birds and light
1.5.1 Sensitivity to Light
Light plays an important role in a bird’s life. It is useful in communication between conspecifics when the see each other’s colours, avoidance of predators by camouflage and detection of resources (water, food or nesting sites) (Blackwell 2002). Animals can have different sensitivities to different wavelengths and light intensity (Morgan and Tromborg 2007). Birds are known to see light both from the visible and the ultraviolet wavelengths. They are tetrachromatic (some species can be pentachromatic) which means that the four principal colours they can detect are ultraviolet, blue, green and red (Maier 1992, Blackwell 2002). Whether the light is in the visible or the UV range, its effect is different on the bird’s behavior.
1.5.1.1 Visible light
Most articles on the effect of light focus on which lights are the most attractive to birds. For example, Avery et al. (1976) indicated that birds seem to be more attracted to red and white lights. The same observation was done by Poot et al. (2008). This research also highlights that green and blue lights don’t attract birds as much. In contrast, Lustick (1973) indicated that they are sensitive to light in between 500 and 506 nm, which is green.
1.5.1.2 Ultraviolet light
Ultraviolet light is necessary for multiple bird behaviours. It can be used in orientation, foraging, and signaling (for mating, conspecific communication and predation, for example) (Blackwell 2002, Hart and Hunt 2007). It is also necessary for birds to detect surrounding colours (Maddocks et al. 2002, Hart and Hunt 2007). In fact, it was shown that the stress level in captive birds with UV-deficient conditions was increased; the birds exhibited more hanging on the cage and pecking at the cage types of behaviour which show that they are trying to escape. In light of these observations, it is possible to say that birds can either be attracted or repelled by light.
1.5.2 Attraction to Light
1.5.2.1 Food source
Lights can be signs of abundant food sources for many bird species. In fact, many researchers have found that birds will be lured in proximity to anthropogenic lights (for example streetlamps) to forage. Robertson et al. (2010) discovered that some birds took advantage of an illuminated structure to hunt insects. The aquatic insects preyed on by the birds are polarotactic, which means that they are attracted to tall glass buildings because they see them as water surfaces. This is due to the polarizing effect glass surfaces have by reflecting sunlight and skylight. Birds have been found to take advantage of this behaviour and feed on the insects that swarmed around these surfaces. Although this isn’t an example of the attraction of birds towards a typical light source, it shows that birds can benefit from lighted structures for foraging purposes. In another observation by van Tets et al. (1969), lights on airport runways were the cause for multiple bird strikes. In fact, these lights attracted birds due to the accumulation of insects around them. These examples demonstrate how birds can be attracted to light sources due to the abundance of resource these lights offer.
1.5.2.2 Nesting sites
Light sources can also give birds another type of cue as nesting. Research has found that birds can decide to nest close to light for the same reason stated above; light offers a food source to insectivorous birds. Building a nest in these areas is therefore favorable to a bird’s survival. Rao and Koli (2017) reported that the presence of road lights encouraged birds from 14 different species to nest close to roads, again due to the fact that streetlamps attract insects. De Molenaar et al. (2000) also observed that black-tailed godwits more often chose to place their nests closer to artificial light near roadways. The exact reasons for this behaviour are unknown although it might again be related to food availability.
1.5.2.3 Protection against predators
Birds can also be attracted to light to protect themselves against predators. In fact, Gorenzel and Salmon (1995) observed that crows living in urban environments roost in areas with high illumination. They argued that this behavior was favorable to crows because the high visibility increases their chance to easily detect the approach of humans or predators (like owls) and they can move between roosting sites without difficulty if they perceive a risk of predation.
1.5.2.4 Orientation
One of the most commonly known reasons why birds can be attracted to light is that nocturnal birds need light to orient themselves during migration. Often, the light they will use is celestial light (Rowan 1925, Cochran et al. 2004). However, when weather conditions are bad, birds will use artificial light sources to find their way (Avery et al. 1976, Ogden 1996, Longcore and Rich 2004). This phenomenon was observed to happen frequently on boats with bright lights (Merkel and Johansen 2011). In another study, birds were highly attracted to lighthouses (Jones and Francis 2003).
1.5.3 Repulsion from Light
Although birds have multiple beneficial reasons for being attracted to light, they can also be repulsed by it. In fact, an experiment entailed placing LED lights inside nest boxes and some birds were found to not enter when the light was turned on (Raap et al. 2017). Moreover, in terms of nocturnal migration, birds seem to avoid lighted structures when the sky is clear and the weather is calm (Avery et al. 1976). In another experiment, birds were also found to sleep away from a light source when the intensity was high (Yorzinski et al. 2015). The exact reasons for this behaviour are unknown.
1.6 How studies were performed
The research works studied in this review used observations and experimentations to determine how light affected birds. These manipulations were either done in a laboratory or in the wild.
1.6.1 Experiments in the laboratory
Laboratory experiments were used mostly in the oldest articles (before 2000). The oldest ones consisted of captured wild birds that were either exposed or not to artificial light and used to determine the difference between two bird population samples. In most of these studies, the birds were killed to determine the size of specific organs or the levels of certain compounds in the birds’ organisms. The most recent articles used the following methodology: the scientists captured birds and exposed them to different artificial light intensities in the lab. Different behaviours were then recorded without killing or hurting the animals.
1.6.2 Experiments in the wild
The later experiments consisted mostly of manipulations in the wild. Often natural or artificial nest boxes already present in an area were used. The researchers inserted artificial light inside these boxes and measured different bird behaviour in reaction to this light. These experiments also solely consisted of observations of birds in relation to certain impacts.
1.6.3 Ethical standards
In the oldest articles, there were no restrictions or constraints in relation to the death of test subjects. In the newest research, a section of the article is reserved to ethics. The details are specific to each research. Often, the experiments were conducted according to certain protocols depending on the region the experiment took place. Most of the times, the animals were not harmed in the procedure of the research.
1.7 The consequences of artificial light on birds
Since natural light has so many different important roles for birds, it is possible to think that artificial light can have multiple effects on bird behaviour. Gathering information across multiple articles helped demonstrate how artificial light can be harmful to birds. Six major aspects of bird behaviour were found to be most impacted by light pollution: reproduction, sleep, migration, activity, communication and problem-solving skills. These will either be impacted negatively or positively. These six facets were examined and their implications for the ecosystem and the bird community were explored.
1.7.1 Reproduction
1.7.1.1 First discovery of the consequence
Rowan (1925) suggested that light is important in bird migration patterns. Moreover, he first discovered that light could have negative consequences on the physiology of dark-eyed juncos (Junco hyemalis). He trapped multiple specimens of this species and placed them into two open-air aviaries. One of the shelters served as a control while the other was the experiment. Every night, the artificial lights (two 50-watt lights) were kept on after sunset for 5 minutes and this time was lengthened to an additional 5 minutes every subsequent night. At every two weeks interval, one bird from the control group and one from the light exposure group were killed and examined. By looking at the size of their testes, Rowan was able to determine that the birds under artificial light had a change in their developmental organs, probably due to light exposure. Although he couldn’t explain the specific way in which the change was taking place, he was convinced that artificial light can induce important changes in the physiology of birds (Rowan 1925). This experiment initiated a multitude of subsequent studies on the
impact of light exposure on birds. Other future studies further tested Rowan’s discovery to determine how the changes were taking place.
1.7.1.2 Modification of the timing of reproduction
Light exposure can modify two different components of reproduction: the timing of reproductive behaviour and the sex apparatus itself. Without killing and examining the reproductive organs of birds, it is possible to determine that artificial light causes changes in the timing of reproduction.
A particular study was done on European blackbirds (Turdus merula). Birds of this species were captured from two different environments: a natural area with multiple bushes and trees, and an urban area with a high population density and few trees. They were tagged with a backpack with a radio-transmitter (to determine the state of activity of the bird) and a light logger (to record the intensity of light exposition) and released. They were caught again later to extract the data from the backpack. The researchers found that their results were consistent with the hypothesis that artificial light can induce variation in the onset of reproductive behaviour of birds (Dominoni and Partecke 2015). A separate experiment conducted on the same bird species (European blackbirds) demonstrated the same effect. The authors started this experiment knowing that urban birds usually breed earlier than rural birds and they wanted to determine why. They collected 30 urban and 30 rural nestlings and placed them in individual cages in a laboratory. For two years, their reproductive behaviour was studied. After this period, males were placed either alone or with a female in separate outdoor rooms. The difference in gonadal development between urban and rural birds are explained in detail in the next section. However, one main conclusion of the experiment was that exposition to a female alone cannot explain the earlier onset of reproductive maturity. In fact, the authors suggested that this difference was most likely due to light pollution (Dominoni et al. 2015). Using the same species of birds, scientists discovered that gonadal development and testosterone production happened up to a month earlier in exposed birds. This result was discovered by experimentally exposing rural and urban birds to either light or no light conditions. The researchers also found that a small light intensity is enough for an effect to be observed (Dominoni et al. 2013).
An experiment on blue tits (Cyanistes caeruleus) showed the same pattern as the two above. The researchers recorded the breeding pattern of birds in nests from areas with artificial light and without (only natural light). It was found that female birds in the lighted area laid their eggs 1.5 days earlier than females in the natural area. This effect was also influenced by the intensity of the light source; if the nest was closer to the streetlamp, the female laid her eggs earlier and if the nest was further away from the streetlamp, the female laid her egg later (Kempenaers et al. 2010).
1.7.1.3 Modification of the sex apparatus
As mentioned earlier in the article by Rowan (1925), light pollution can alter the development of the sexual organs of birds. Rowan continued his experimentation with another article published in 1937 on the effect of light on gonadal development. The same bird species was used: dark-eyed junco (Junco hyemalis). The manipulations are similar: male birds were captured and kept in either control cages or experimental cages (under an electric bulb). The light in the experimental cages was turned on every night and turned off a little later every subsequent night to gradually increase exposure to light. Every week, birds were killed and their testes were examined. The experiment took place for 8 weeks. As expected, the testes of the animals exposed to light were larger than those of the control birds. Moreover, the testes of birds increased in accordance with light exposure: the longer the light exposure, the larger the testes became. In another part of his experiment, Rowan recovered wild birds from an urban environment (London) and compared the size of their testes with birds from a rural area (Bedfordshire). Again, for birds collected at approximately the same date, the size of the testes was larger in urban birds than in rural birds (Rowan 1937).
Dominoni et al. (2015) also demonstrated an effect of light on testes size, as stated above. The researchers measured the size of birds’ testes four times during the experiment by incision. It was discovered that males housing with a female had functional sexual organs 3.5 days earlier than males without a female, but the authors explain that this difference is not significant. Moreover, there was no difference in the size of testes between rural and urban birds. These conclusions are unsatisfying in terms of the effect of social cues on bird reproductive behaviour. Again, the authors suggest that the main difference that is usually seen in the wild in testes size is most probably caused by artificial light exposure (Dominoni et al. 2015).
A study by Yoshimura et al. (2003) demonstrated a possible mechanism by which birds’ sexual organs are affected by lighting. The medial basal hypothalamus (MBH) is a part of the brain which is responsible for the response to photoperiodic time exposure. Simulating this area increased the growth of the testis in male birds. The authors exposed male Japanese quail to either 1 hour of light exposure or no light. They then extracted their MBH. A gene for a specific enzyme was highly expressed in birds exposed to light: Dio2. This shows that this enzyme is probably responsible for the physical response of birds to light exposure.
1.7.2 Sleep pattern
1.7.2.1 Modification of the timing and the duration of sleep
Light pollution has been known to affect the sleep pattern of birds. The two main ways in which it happens is through the modification of the timing of sleep onset and offset and the modification of the total time spent
sleeping. In fact, birds are seen to fall asleep later and wake up earlier, which in turn reduces the amount of sleep the birds get.
Sun et al. (2017) made these observations in two birds: the great tits (Parus major) and the blue tits (Cyanistes caeruleus). They did so by installing LED lights inside wild nest boxes in a semi-rural area. Some nests were lighted and others served as control (without a light). The nests were kept dark the first night and then the light was turned on. After filming the movements of the wild birds, the researchers found that great tits were highly affected by light pollution; they entered their nest later at night and exited their nest earlier in the morning. Moreover, they fell asleep later and woke up earlier. The sleep percentage (the amount of time spent sleeping on the total amount of time spent in the nest) of great tits was reduced by 3% and their total sleep time (the number of minutes spent sleeping per night) was reduced by an hour. Blue tits were not affected as much; their latency (the time spent in the nest box before falling asleep at night) was lengthened, although not as strongly as in great tits (Sun et al. 2017). Raap et al. (2017) observed wild great tits in the same way that Sun et al. (2017) did. They placed LED lights inside nest boxes and determine the effect of this light on sleep. Birds were found to wake up earlier and leave their nest box earlier than in dark conditions. Moreover, they fell asleep 15 minutes later. Morning and evening latency were increased. The length of sleep the birds had was reduced by approximately 40 minutes due to light exposure. A novel discovery from this study was that light intensity doesn’t affect the changes in sleep behaviour. A higher light intensity delayed sleep onset whereas a lower intensity didn’t. However, that is the only difference in light intensity observed (Raap et al. 2017)., The same pattern was observed by Yorzinski et al. (2015) in adult peahens. The authors observed that the birds had a reduced time spent sleeping due to the increased vigilance they had to do in high luminated areas. They did so by measuring the level of head movements with accelerometers attached to the birds’ heads. Captured peahens were staying in either a control cage (no light) or an experimental cage (with light) for seven nights. The birds showed more head movements (used as a proxy for vigilance in this experiment) when the artificial light was on compared to when it was off. These birds were also the ones spending less time sleeping. When artificial light was low or there was no artificial light, birds were less vigilant because being vigilant in these conditions is useless since vision is decreased (it’s dark) (Yorzinski et al. 2015).
1.7.3 Migration
Many nocturnal bird species use natural light to orient themselves during their migration. As expected, artificial light sources can replace this light and the migration pattern of birds will be disrupted.