LES DYSFONCTIONS METABOLIQUES ET IMMUNUAIRES CHEZ LES VACHES LAITIERES PERIPARTURffiNTES
Par
Marie-Claude Loiselle
memoire presente a la faculte des sciences en vue de l'obtention du grade de maitre es science (M.Sc.)
FACULTE DES SCIENCES UNIVERSITE DE SHERBROOKE
1*1
Library and Archives Canada Published Heritage Branch 395 Wellington Street OttawaONK1A0N4 Canada Bibliotheque et Archives Canada Direction du Patrimoine de I'edition 395, rue Wellington OttawaONK1A0N4 CanadaYour file Votre reference ISBN: 978-0-494-53179-2 Our file Notre reference ISBN: 978-0-494-53179-2
NOTICE: AVIS:
The author has granted a
non-exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
telecommunication or on the Internet, loan, distribute and sell theses
worldwide, for commercial or non-commercial purposes, in microform, paper, electronic and/or any other formats.
L'auteur a accorde une licence non exclusive permettant a la Bibliotheque et Archives Canada de reproduire, publier, archiver, sauvegarder, conserver, transmettre au public par telecommunication ou par I'lnternet, prefer, distribuer et vendre des theses partout dans le monde, a des fins commerciales ou autres, sur support microforme, papier, electronique et/ou autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur conserve la propriete du droit d'auteur et des droits moraux qui protege cette these. Ni la these ni des extraits substantiels de celle-ci ne doivent etre imprimes ou autrement
reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
Conformement a la loi canadienne sur la protection de la vie privee, quelques formulaires secondaires ont ete enleves de cette these.
While these forms may be included in the document page count, their removal does not represent any loss of content from the thesis.
Bien que ces formulaires aient inclus dans la pagination, il n'y aura aucun contenu manquant.
1*1
Lel5mai2009
lejury a accepte le memoire de Mme Marie-Claude Loiselle dans sa version finale.
Membres dujury M. Brian Talbot Directeur Departement de biologie M. Pierre Lacasse Codirecteur
Centre de rercherche et developpement sur le bovin laitier et le pore
M. Martin Lessard Membre
Centre de recherche et developpement sur le bovin laitier et le pore
M. Viktor Steimle President-rapporteur Departement de biologie
SOMMAIRE
La periode de transition entre la gestation et lactation chez la vache laitiere est marquee par des changements metaboliques, hormonaux et immunitaires ayant un impact sur l'incidence des infections et des maladies metaboliques. Le but de ce projet etait d'induire une production de lait reduite en debut de lactation afin d'attenuer le bilan energetique negatif et les problemes metaboliques tout en augmentant les fonctions immunitaires. Pour realiser cet objectif, 22 vaches multipares ont ete traites 1 fois par jour ou 2 fois par jour durant la premiere semaine post-partum. Par la suite, les animaux ont ete soumis a deux traites par jour, ce qui correspond a une regie classique d'un troupeau laitier. Des prelevements sanguins et des isolements de leucocytes ont ete realises durant la periode experimentale couvrant les 5 semaines avant et 5 semaines apres la parturition. Le traitement a permis de reduire la production de lait de 30% durant la semaine suivant le velage, par contre apres le retour a deux traites par jour la production laitiere demeure inferieure de 3.5 Kg de lait par jour. Le projet a permis de mettre en evidence que la parturition induit une augmentation des acides gras non esterifies (AGNE), du P-hydroxybutyrate (BHB), de l'uree et de la bilirubine. L'augmentation des AGNE et BHB A sont significativement plus hautes chez les vaches traites 2 fois par jour. Durant cette meme periode, les concentrations de glucose et de leptine diminuent, mais demeurent superieure chez les vaches traites 1 fois par jour. La difference entre les deux groupes persiste au-dela de la periode de traitement soit jusqu'au jour 14
post-partum pour le BHB A et jusqu'au jour 24 post-post-partum pour le glucose et les AGNE. Ces
parametres demontrent bien que la reduction de la frequence de traite induit un moins grand deficit energetique et une moins grande mobilisation des reserves de gras et des glucides mettant en evidence les bienfaits de notre traitement. Les fonctions immunitaires evaluees ne demontrent pas clairement le phenomene d'immunodepression retrouve chez les vaches laitieres en debut de lactation. Done, en l'absence d'une defaillance immunitaire marquee, il est impossible de conclure quant a l'apport de la frequence de traite sur ces parametres. En conclusion, la diminution de la frequence de traite en debut de lactation limite les desordres
metaboliques, mais induit une reduction de la production laitiere meme apres le retour a une frequence normale de 2 traites par jour.
REMERCIEMENTS
Tout d'abord, je tiens a remercier le Dr Pierre Lacasse, le Dr Brian Talbot, ainsi que le Dr Louis Delbecchi pour m'avoir permis de realiser ce projet de maitrise dans un environnement dynamique et enrichissant.
Je tiens a remercier tout specialement le Dr Celine Ster et Lisette St-James pour leur patience et pour toute l'energie qu'elles ont consacree a la realisation de ce projet. Un gros merci a toute l'equipe du laboratoire qui a su creer une belle ambiance de travaille durant mon projet: Frederic Beaudoin, Guillaume Tremblay, Jasmin Brochu et Philippe Bernier-Dodier. Un merci particulier a Audrey Ballan et Etienne Quintal pour le soutien technique durant la phase experimentale. J'aimerai remercier Steve Methot pour les analyses statistiques ainsi que tout le personnel de l'etable pour leur collaboration dans le projet.
Je tiens aussi a remercier tous les membres de mon comite de maitrise pour leur soutien et leurs conseils.
Merci aux divers organismes pour le soutien financier et pour avoir rendu possible cette etude : la federation des producteurs laitiers du Canada, Agriculture et Agro-alimentaire Canada, le Conseil de recherches en sciences naturelles et en genie du Canada ainsi que l'Universite de Sherbrooke.
Je tiens a temoigner toute ma reconnaissance envers mes parents, ma famille et mes amis pour leur support moral et leurs encouragements.
TABLES DES MATIERES
SOMMAIRE ii REMERCIEMENTS iv
TABLES DES MATIERES v LISTE DES ABREVIATIONS vii LISTE DES TABLEAUX viii LISTE DES FIGURES ix 1. INTRODUCTION 1
1.1 Evolution de la production laitiere 1 1.2 Mise en place de la lactation 3
1.3 Le bilan energetique 4 1.4 Changements metaboliques/hormonaux et l'immunodepression 5
1.4.1 Changements metaboliques 7 1.4.1.1 Calcium 7 1.4.1.2 Phosphore 10 1.4.1.3 Uree 11 1.4.1.4 Bilirubine 13 1.4.1.5 Glucose 14 1.4.1.6 Les acides gras 16
1.4.2 Changements hormonaux 19
1.4.2.1 Les estrogenes 19 1.4.2.2. Stanniocalcine (STC) 21
1.4.2.3 Leptine 23
1.5 Dysfonctions immunitaires et phenomene d'immunodepression 26
1.5.1 Immunite innee 26 1.5.2 Immunite acquise 29
1.5.3 Cytokines 31
1.6 Les causes de l'immunodepression 33
1.6.1 Deficit energetique 33 1.6.2 Changements hormonaux 34 1.6.3 Les metabolites et vitamines 35
1.7 Reduction de l'immunodepression 36
1.8 La traite prepartum 37 1.9 L'influence de la frequence de traite 37
1.10 Hypotheses 39 1.11 Objectifs 40
1.11.1 Objectifs specifiques 40
2-ARTICLE 41 2.1 Reference de l'article 41
2.2 Contribution de l'auteur 41 2.3 Originalite du travail et apport a la science 42
2.4 Article 43 2.4.1 INTERPRETIVE SUMMARY 43
2.4.2 ABSTRACT 45 2.4.3 INTRODUCTION 47 2.4.4 MATERIALS AND METHODS 50
2.4.5 RESULTS 59 2.4.6 DISCUSSION 73 2.4.7 ACKNOWLEDGEMENTS 79 2.4.8 REFERENCES 80 3 DISCUSSION 87 4 CONCLUSION 90 5 BIBLIOGRAPHIE 91
LISTE DES ABREVIATIONS AGNE: ARNm: BEN: BHB: El: E2: ELAM: ERp: ERcc: GH: HUVEC: ICAM: IgA, E, M, D, G, Gi IFN-y: IL-2, 4, 8, 10 MCP-1: NK: PBMC: PTH: PMN: SEM: STC: TAG: TNF-cc: VLDL:
Acides gras non esterifies Acide ribonucleique messager Balance energetique negative P-hydroxybutyrate
Estrone 17P-estradiol
Molecule d'adhesion endotheliale-leucocyte Recepteur beta de l'estrogene
Recepteur alpha de l'estrogene 'Growth hormone'
'human umbilical vein endothelial cells' Molecule d'adhesion inter cellulaire Immunoglobuline A, E, D, M, G, Gi Interferon gamma
Interleukine 2, 4, 8, 10
'Monocyte chemotactic protein-1' 'Natural killer'
Cellules mononucleates du sang peripherique Parathormone
Polynucleaires neutrophiles Erreur standard de la moyenne Stanniocalcine
Triacylglycerol
Facteur de necrose tumorale alpha 'Very low density lipoprotein'
LISTE DES TABLEAUX
1. Resume des differents changements metaboliques et hormonaux survenant durant la
LISTE DES FIGURES
Production moyenne de lait en Kg (toutes les races)- releves officiels
1960-2001 2
Nombre de vaches laitieres en fonction du nombre de kilolitres de lait produit de
1976-2001 au Canada 3
Courbe typique de la lactation d'une vache laitiere 5
Concentration de calcium plasmatique chez des vaches intactes et des vaches
mastectomisees durant la periode periparturiente 9
Concentration de Cortisol plasmatique chez les vaches periparturientes ayant
developpe des symptomes d'hypocalcemie et de fievre du lait 10
Estimation de la demande globale en glucose du corps compare a la quantite de
glucose ingere durant la periode peripartum 15
Concentration plasmatique des acides gras non esterifiees chez des vaches intactes et
mastectomisees 18
Concentration plasmatique des estrogenes et de la progesterone chez des vaches en fin
de gestation 21
Les changements de la concentration plasmatique de la leptine durant la periode
Moyenne de l'expression de la L-selectine sur les neutrophiles actives avec l'activateur de plaquettes chez des vaches intactes et des vaches mastectomisees
durant la periode entourant la parturition 28
Moyenne de l'activite de la myeloperoxidase chez des vaches intactes et des vaches
mastectomisees durant la periode entourant la parturition 29
Moyenne des pourcentages des lymphocytes totaux durant la periode
periparturiante 31
Activite transcriptionnelle de TNF-alpha, DL-8 et IL-10 en debut, milieu et fin de
1. INTRODUCTION
1.1 Evolution de la production laitiere
La production laitiere bovine a debute en Amerique du Nord dans les annees 1600 avec l'arrivee des premiers colons. Jusqu'en 1850, la production laitiere est locale et destinee a la consommation de lait et de beurre. Avec 1'industrialisation, les installations laitieres grossissent de facon importante et l'emergence de produits transformes tels que les fromages et le lait condense font leur entree sur le marche. Des 1905, le gouvernement instaure un programme canadien d'aptitudes des bovins laitiers pour evaluer sur de longues periodes des troupeaux entiers et de permettre l'obtention d'une lignee avec un rendement eleve en lait.
Depuis les annees 60, une augmentation de plus de 100% de la production de lait par vache est observee grace aux ameliorations genetiques, a 1'amelioration de la gestion des troupeaux ainsi qu'a l'acquisition de nouvelles connaissances sur 1'alimentation et la sante des animaux (Agriculture Agroalimentaire Canada, 2002) (figure 1.1). Cette augmentation des rendements laitiers a permis de diminuer le nombre de vaches en lactation tout en gardant une quantite de lait produite egale ou superieure (Statistique Canada) (figure 1.2).
Le rendement laitier des vaches laitieres au Canada a presque double depuis les annees 60 grace a diverses avancees agronomiques et genetiques. La selection des animaux a ete effectuee principalement en fonction de la productivite laitiere. Les rendements laitiers quotidiens de plus en plus eleves amenent certains problemes de sante chez les vaches. Des maladies infectieuses ainsi que des troubles metaboliques sont souvent associes avec de hauts rendements laitiers. Ces problemes engendrent des pertes economiques importantes pour les
pouvant affecter le systeme musculaire et nerveux, possede un taux de mortalite de 4 %. La fievre du lait engendre des pertes de 275 $ incluant une perte de production de 130 kg de lait par animal atteint (Frappier, 2001).
10000 9000 8000 7000 H 6000 H 5000 4000 400 1960 1965 1970 1975 1980 1985 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 ^ ^ M i l k / L a i t -o>Fat/Gras =£~Protein/Protelne
Figure 1.1: Production de lait moyenne (toutes les races)- releves officiels 1960-2001
(Figure provenant d'Agriculture Agroalimentaire Canada - Conseil Canadien du controle laitier).
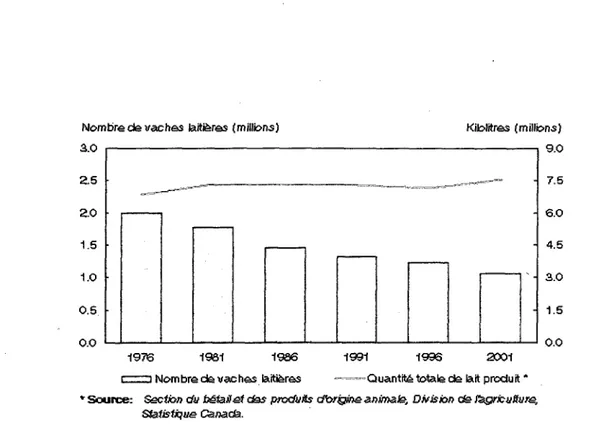
Nombre de vach&s laitieres (millions) Kilolitres (millions) 3.0 2.5 2.0 1.5 1.0 0.5 0.0 1976 1961 1936 1991 1996 2001 C==Z3 Nombre d& vacheslaiti&res ™=—-Quantity totalede lait produit *
*" Source: Sscfen du b§isil&t das produits dtor&ir>& animate, Dffisktn ds fegricuttur&, Sfatisfiqas Canada.
Figure 1.2 : Nombre de vaches laitieres en fonction du nombre de kilolitres de lait produit de
1976-2001 au Canada (Statistique Canada).
1.2 Mise en place de la lactation
La lactogenese correspond a une serie d'evenements cellulaires dans la glande mammaire qui donne lieu une production de lait abondante au moment de la parturition. Avant la mise bas, un processus de differentiation cellulaire et plusieurs changements enzymatiques permettent la synthese des constituants du lait dans la glande mammaire (Lactation and the mammary gland, 2002). Cependant, la production de lait en quantite importante ne debutera qu'a la parturition initiee par plusieurs changements endocriniens. La secretion de la progesterone diminue a cause de la regression du corps jaune. Cette diminution entraine la liberation massive de la prolactine qui stimule la synthese des composantes du lait. De plus, 1'augmentation des glucocorticoi'des et des estrogenes durant la periode qui entoure la parturition joue aussi un role considerable dans la lactogenese (Biology of lactation, 2000).
Dans la regie normale des troupeaux laitiers au Canada, des la parturition, la vache est traite deux fois par jour. La production de lait augmente jusqu'a un pic de lactation se situant entre 2 et 8 semaines post-partum et diminue graduellement en meme temps que le nombre de cellules epitheliales mammaires pour finalement atteindre une periode de tarissement environ 305 jours apres le velage. La periode la plus critique pour la sante des vaches se situe de la parturition jusqu'au pic de lactation, car durant cette periode la vache passe d'une periode de tarissement a une periode de production de lait maximale (Grummer et ah, 1995). De plus, les vaches subissent plusieurs changements hormonaux et metaboliques lies a la mise en place de la lactation qui ont un impact sur leur sante (Drackley et ah, 1999). En prime, on assiste a une grande mobilisation de l'energie, car celle fournie par l'apport alimentaire est generalement insuffisante pour combler celle necessaire pour la production de lait ce qui entraine un deficit energetique (Accorsi et ah, 2005).
1.3 Le bilan energetique
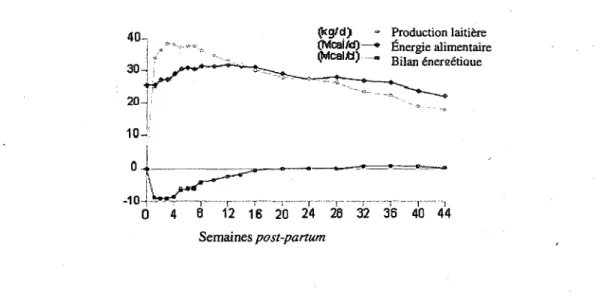
Le bilan energetique represente l'energie nette consommee via la nourriture moins l'energie requise pour les fonctions vitales et les activites quotidiennes. Un bilan energetique negatif (BEN) aura pour effet d'augmenter la mobilisation des reserves adipeuses, proteiques et glucidiques afin de combler le deficit. Ce phenomene est observe chez la vache laitiere en debut de lactation. L'apport alimentaire est incapable de combler la demande en energie necessaire pour la production de lait (Goff et Horst, 1997). De plus, une diminution de l'appetit des vaches laitieres en debut de lactation accentue le BEN. Le bilan energetique retrouve son equilibre en moyenne vers la 16e semaine de lactation, cependant, ce phenomene est tres variable d'un animal a l'autre (DeLaval, 1980) (figure 1.3). Les problemes de sante sont les plus frequents et les plus nombreux a survenir chez les vaches laitieres au meme moment ou le deficit energetique est a son plus haut c'est-a-dire durant les premieres semaines de lactation (Drackley, 1999).
Pour contrecairer les problemes de santes relies au deficit energetique, la variation des rations alimentaires a souvent ete etudiee pour ameliorer la sant€ des vaches periparturientes. Les vaches en debut de lactation, nourries avec une diete riche en glucides, demontrent une moins grande mobilisation des reserves d'acides gras comparativement aux vaches nourries avec une ration riche en lipides (Van Knegsel et al, 2007). De plus, celles nourries avec une ration riche en lipides ont des desordres metabolites plus importants. Mais, aucune alimentation particuliere n'a permis d'enrayer completement le deficit energetique en debut de lactation.
(bg/d) - Production laitifere S5?ca!SJ—* fiiergie alimentaire (Mcalfl).-, Bilan6nersetiQue - 1 0 - J - ^ — - T - r- T - i -r 0 4 8 12 16 20 24 28 32 36 40 44 Semaines post-partum
Figure 1.3 : Courbe typique de la lactation d'une vache laitiere (Tiree de DeLaval, 1980).
1.4 Changements metaboliques/hormonaux et Fimmunodepression
Plusieurs changements metaboliques et hormonaux caracterisent la periode entourant la parturition (tableau 1). Ces changements sont surtout lies a la mise en place de la lactation et a la reorganisation des reserves energetiques afm de combler le bilan negatif cause par la production de lait. En plus, cette periode est aussi marquee par une diminution des fonctions immunitaires qualifiees d'immunodepression. Plusieurs etudes semblent montrer une diminution du recrutement et de la defense des neutrophiles (Monfardini et ah, 2002 ; Hoeben
qu'une modification dans l'activite des lymphocytes (Lacetera et al, 2005 ; Nonnecke et al, 2003 ; Kehrli et al, 1989). Les causes de cette immunodepression ne sont pas clairement identifiees. Plusieurs etudes tentent de demontrer l'impact des changements metaboliques et hormonaux sur ce phenomene. II est done primordial de bien connaitre les changements metaboliques et hormonaux durant la periode periparturiante afin de mieux comprendre 1' immunodepression post-partum.
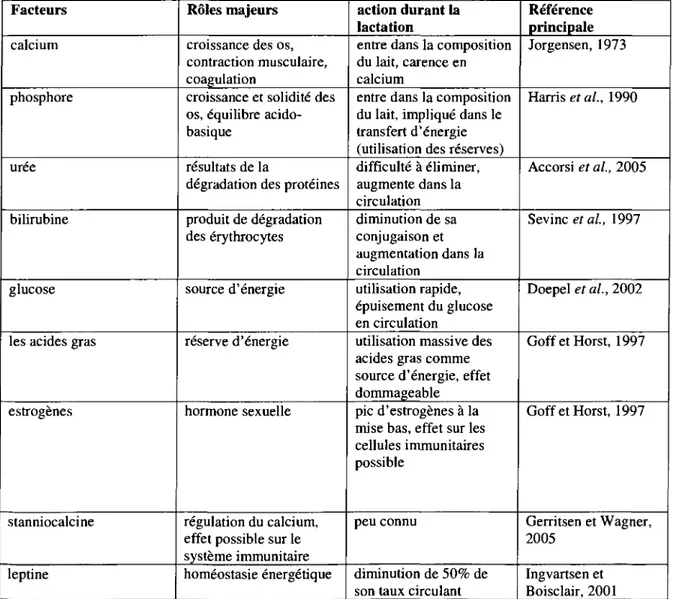
Tableau 1. Resume des differents changements metaboliques et hormonaux survenant durant
la periode peripartum chez les vaches laitieres. Facteurs calcium phosphore uree bilirubine glucose
les acides gras
estrogenes
stanniocalcine
leptine
Roles majeurs croissance des os, contraction musculaire, coagulation
croissance et solidite des os, equilibre acido-basique
resultats de la
degradation des proteines produit de degradation des erythrocytes source d'energie reserve d'energie hormone sexuelle regulation du calcium, effet possible sur le systeme immunitaire homeostasie energetique
action durant la lactation
entre dans la composition du lait, carence en calcium
entre dans la composition du lait, implique dans le transfert d'energie (utilisation des reserves) difficulte a eliminer, augmente dans la circulation diminution de sa conjugaison et augmentation dans la circulation utilisation rapide, epuisement du glucose en circulation
utilisation massive des acides gras comme source d'energie, effet dommageable
pic d'estrogenes a la mise bas, effet sur les cellules immunitaires possible
peu connu
diminution de 50% de son taux circulant
Reference principale Jorgensen, 1973 Harrises al, 1990 Accorsi et al, 2005 Sevinc etal, 1997 Doepel et al., 2002 GoffetHorst, 1997 GoffetHorst, 1997 Gerritsen et Wagner, 2005 Ingvartsen et Boisclair, 2001
1.4.1 Changements metaboliques
Durant la periode entourant la mise bas, les vaches laitieres subissent plusieurs changements particulierement au niveau des metabolites. Ces changements peuvent provoquer de graves problemes de sante chez le bovin laitier pouvant mener jusqu'a la mort de 1'animal. Les desordres metaboliques entrainent des pertes economiques importantes notamment au niveau des traitements et du rendement laitier (Frappier, 2001). Les vaches ayant des problemes de sante quelconque durant la periode peripartum produisent 7.2 Kg de lait en moins par jour durant les 20 premiers jours de lactation comparativement aux vaches en sante (Drackley, 1999). Au niveau des metabolites sanguins, plusieurs changements sont observes : une baisse du calcium, de phosphore, du glucose ainsi qu'une augmentation de la bilirubine, de l'uree, des AGNE (acides gras non esterifies) et des BHB (beta-hydroxybutyrate) (tableau 1).
1.4.1.1 Calcium
Le calcium est un mineral tres abondant dans le corps. Plus de 98 % du calcium de l'organisme se trouve dans les os et les dents (Tortora et ah, 1999). Les roles du calcium dans le metabolisme sont nombreux. Le calcium est implique dans la contraction musculaire en se liant a la troponine (muscle squelettique ou cardiaque) ou encore a la calmoduline (muscle lisse) (Hartshorne et Siemankowski, 1981). Le transfert d'information a travers les synapses chimiques lors d'influx nerveux necessite aussi la presence de calcium. De plus, le calcium est necessaire durant les etapes de la coagulation sanguine. Le calcium agit en tant que second messager dans la signalisation cellulaire notamment chez les lymphocytes (Grafton et Thwaite, 2001). La presence d'une concentration minimale de calcium intracellulaire est necessaire pour 1'activation des lymphocytes. Les ions calcium sont soumis a un equilibre precis permettant d'avoir une concentration constante et peu variable dans le sang. Le calcium dans l'organisme est regule par deux hormones soit la parathormone (PTH) et la
calcitonine permettant une homeostasie du calcium (Tortora et al, 1999). La PTH permet la liberation d'ions calcium a partir de la matrice osseuse et augmente la reabsorption calcique tandis que la calcitonine inhibe la resorption osseuse ainsi que la reabsorption d'ions calcium (Tortora et al, 1999).
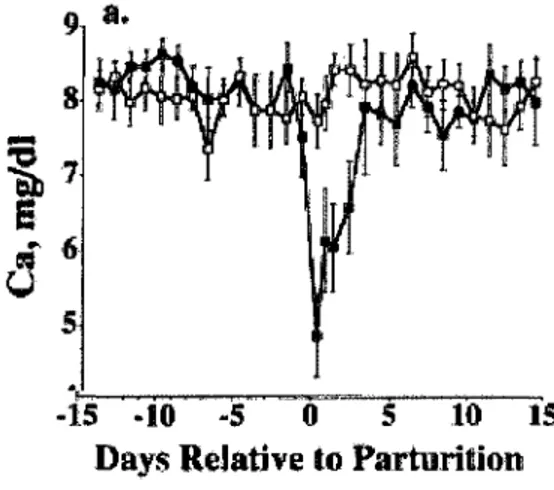
La production de lait est la principale raison de la chute du calcium sanguin chez la vache periparturiente. Chez les vaches mastectomisees, la baisse du calcium est moindre que chez les vaches ayant leur glande mammaire (figure 1.4) (Goff et al, 2002). En debut de lactation, la demande en calcium augmente considerablement pour la production du colostrum et du lait. Malgre la grande resorption du calcium a partir des os, les mecanismes de regulation du calcium deviennent rapidement insuffisants pour repondre a la forte demande. De plus, la reponse des os face a la PTH semble diminuer chez les vaches ayant le syndrome de la fievre du lait (hypocalcemie) ce qui contribue a accroitre la chute du calcium sanguin. Chez les vaches en calcemie faible (inferieur a 8 mg/dL), certaines repercussions graves sur les systemes musculaires et nerveux peuvent etre constatees. En cas d'hypocalcemie severe, soit un taux de calcium inferieur a 5 mg/dL, une paresie post-partum peut survenir chez 1'animal. Generalement, les vaches ayant des fievres du lait sont a risque de presenter d'autres desordres metaboliques. Les vaches plus agees et ayant eu un nombre eleve de lactation sont plus susceptibles de developper des problemes d'hypocalcemie durant la periode peripartum (Jorgensen, 1973).
Le pic d'estrogene avant le velage et les hormones relachees lors du stress a la parturition telles que les glucocorticoides et l'epinephrine contribuent egalement a reduire le taux de calcium circulant (Jorgensen, 1973). Des etudes demontrent que le niveau de Cortisol sanguin est plus eleve chez les animaux souffrant de fievre du lait (figure 1.5) (Goff et Horst, 1997). Le Cortisol est une hormone qui provoque une baisse du systeme immunitaire pouvant possiblement amplifier le phenomene d'immunodepression present chez la vache en debut de
lactation (Goff et Horst, 1997). De plus, en tant que glucocorticoi'de, elle diminue le taux de calcium circulant. En debut de lactation, l'exces de Cortisol combine avec la diminution du tonus musculaire au niveau de l'uterus et des sphincters des trayons (cause par le declin de calcium) augmentent les risques d'incidence de retention placentaire et de mammites (Goff et Horst, 1997).
L'hypocalcemie diminue la secretion d'insuline ce qui decroit la quantite de glucose pouvant etre capte par les tissus (Littledike et ah, 1970). Suite au velage, ce phenomene entraine une diminution de glucose disponible ce qui accentue la mobilisation des lipides pouvant causer des dommages importants particulierement au niveau du foie.
V , , . . . — ^ — _ .
-15 - 1 0 - 5 0 5 10 IS
Days Relative to Parturition
Figure 1.4 : Concentration de calcium plasmatique chez des vaches intactes (n=8, noir) et des
©
M l
et e
•2 1 0 1 2
Day of Calving
Figure 1.5: Concentration de Cortisol plasmatique chez les vaches periparturientes ayant
developpe des symptomes d'hypocalcemie et de fievre du lait (cercle noir). Vaches ayant developpe une hypocalcemie, mais sans fievre du lait clinique (cercle blanc). Vaches n'ayant pas eu d'hypocalcemie ou de fievre du lait (triangle blanc) (Goff et Horst, 1997).
1.4.1.2 Phosphore
Le phosphore tout comme le calcium est regule par la PTH et la calcitonine (Tortora et al, 1999). Le phosphore agit a plusieurs niveaux dans l'organisme. Tout d'abord, le phosphore agit en synergie avec le calcium dans la croissance et la solidite des os et des dents (Tortora et
ah, 1999). II est necessaire au metabolisme energetique ainsi que dans l'equilibre
acido-basique de l'organisme. Le phosphore est implique dans plusieurs reactions enzymatiques. L'absorption du phosphore implique la vitamine D et s'effectue dans le petit intestin (Barlet et
Le taux de phosphore dans 1'alimentation des vaches laitieres recommande par le National Research Council est de 0.26 a 0.40 % de matiere seche (National Research Council, 2001). Durant la lactation, les besoins en phosphore augmentent, car il est grandement implique dans la composition du lait. Plus la production de lait est importante, plus le besoin en phosphore augmente. Le lait contient environ 0.095% de phosphore (Harris et ah, 1990). Ce pourcentage de phosphore est constant dans le lait, peu importe la quantite contenue dans 1'alimentation. Les cas severes de carence en phosphore sont caracterises par une augmentation de la fragilite des os. De plus, une carence alimentaire en phosphore peut perturber la production de lait ainsi que diminuer l'appetit des animaux. Le phosphore est aussi tres important dans diverses reactions biochimiques notamment en ce qui concerne le transfer! d'energie. II est implique dans l'utilisation des lipides et des glucides. En debut de lactation, une carence en phosphore peut done diminuer l'utilisation des reserves lipidiques et glucidiques et ainsi accentuer le deficit energetique.
1.4.1.3 Uree
La majorite des proteines alimentaires et les sources d'azotes non proteiques sont fermentees par les bacteries dans le rumen pour produire des chatnes carbonees et de l'ammoniaque. Par la suite, l'ammoniaque est utilisee par les bacteries pour former des acides amines et etre incorpore dans les proteines microbiennes (Leng et Nolan, 1984). Ces proteines microbiennes deviennent la source principale de proteines pour les ruminants et seront absorbees par l'intestin grele. Cependant, le surplus d'ammoniaque provenant de la fermentation ruminale diffuse a travers la paroi du rumen pour se retrouver dans la circulation sanguine. L'ammoniaque qui est toxique pour 1'animal est transformee en uree dans le foie (Leng et Nolan, 1984). Une partie de l'uree produite par le foie retourne dans le rumen via la salive. L'uree non recyclee sera eliminee par voie urinaire.
Tout ce processus necessite une quantite importante d'energie pouvant causer des deficits si la quantite de proteines et d'azote alimentaire n'est pas controlee (Tamminga, 2006). Une quantite trop importante de proteines dans 1'alimentation amene un degagement d'ammoniaque plus grand ce qui necessite plus d'energie pour l'eliminer sous forme d'uree. Cependant, une quantite trop petite de proteines dans 1'alimentation entraine une faible quantite d'ammoniaque dans le rumen disponible pour les microorganismes. Ce phenomene resulte en une diminution de la synthese proteique des microorganismes.
En debut de lactation, les vaches se retrouvent en deficit energetique et peuvent eprouver certaines difficultes et un manque d'energie pour evacuer l'ammoniaque ce qui conduit a une augmentation de l'uree dans le sang. Generalement chez la vache laitiere, on note une diminution de l'uree plasmatique juste avant velage suivi d'une augmentation apres la parturition. Une hypothese est que les estrogenes et la progesterone presents durant la gestation auraient un effet suppresseur sur le cycle ureique causant ainsi des difficultes pour evacuer le surplus d'uree (Accorsi et ah, 2005).
Normalement chez la vache laitiere, on observe la presence d'uree dans le sang et dans le lait a des concentrations peu dommageables pour la sante (10 a 16 mg/dL) (Calberry, 2003). Par contre en debut de lactation, 1'augmentation d'uree dans la circulation peut produire des effets nefastes pour les vaches. Tout d'abord, on note une correlation positive de l'uree et de l'ammoniaque sur l'incidence de la mammite bovine (Duval, 1995). Une etude a aussi permis de demontrer une correlation positive entre l'uree et le compte des cellules somatiques chez les vaches Holstein-Friesian (Tietze et Majewski, 1997). Un compte eleve en cellule somatique indique souvent une infection de la glande mammaire (Leslie, 1985). De plus, un taux d'uree superieur a 15,4 mg/dL compromet les chances de fertilite chez la vache laitiere (Rajala-Schultz et ah, 2000 ; Ferguson et ah, 1993). La diminution de la fertilite est causee par la diminution du pH uterin durant la phase luteale (Elrod et Butler, 1993). En debut de
lactation, 1'augmentation de l'uree semble entrainer un declin de la fertilite et aggraver le deficit energetique pouvant causer des problemes majeurs de sante et accentuer le phenomene d'immunodepression (Tamminga, 2006 ; Rajala-Schultz etal, 2000 ; Ferguson et al, 1993).
1.4.1.4 Bilirubine
La bilirubine est un pigment jaune qui est le principal produit de la degradation de l'hemoglobine (heme) des erythrocytes (Tortora et al, 1999). La bilirubine est formee par deux enzymes : l'heme oxygenase et la biliverdine reductase. La bilirubine existe sous deux formes : la bilirubine libre (non conjuguee) et la bilirubine conjuguee (Stocker et al, 1987). La bilirubine libre est produite dans la rate et la moelle osseuse lors de la destruction des erythrocytes arrives au terme de leur existence. La bilirubine libre est tres peu soluble dans l'eau et se lie a l'albumine pour etre acheminee vers le foie (Stocker et al, 1987). Dans le foie, la bilirubine est conjuguee avec l'acide glucuronique par la glucuronyl transferase pour devenir soluble. La bilirubine conjuguee est ensuite excretee dans la bile. La bile est stockee dans la vesicule biliaire qui deverse son contenu dans l'intestin. La bilirubine est alors excretee dans l'intestin ou grace a la fermentation par la flore intestinale elle se retrouvera sous forme d'urobilinogene et eliminee dans les selles. Une faible proportion de l'uribilinogene est reabsorbe et sera excrete dans les urines sous forme d'urobiline.
Chez la vache laitiere, 1'evaluation de la bilirubine est utilisee pour estimer les fonctions hepatiques. La bilirubine et les acides biliaires sont parmi les indicateurs les plus utiles pour deceler une defaillance hepatique (Braun et al, 1986). Une augmentation de la bilirubine totale et non conjuguee est souvent presente lors d'une acetonemic chez la vache (Sevinc et
al, 1997). La forte accumulation de gras dans le foie entrame un ralentissement de la
L'augmentation de la production laitiere entraine un accroissement du deficit energetique et accentue la mobilisation des graisses causant des dommages hepatiques ainsi qu'une reduction de la conjugaison de la bilirubine.
1.4.1.5 Glucose
Le glucose est le principal combustible de l'organisme et agit comme source d'energie premiere dans l'organisme notamment au niveau des muscles et du cerveau. La glycemie dans l'organisme est soumise a une regulation tres controlee. Le glucose est regule par quatre hormones principales : l'insuline (produite dans le pancreas), le glucagon (produite dans le pancreas), rhormone de croissance (GH) et 1'adrenaline (produite dans la medullosurrenale) (Annison et ah, 1968). Le glucagon, la GH et 1'adrenaline sont hyperglycemiants tandis que l'insuline est hypoglycemiante. Le foie est un organe vital implique dans la glycemie de l'organisme. Celui-ci controle la glycemie par trois voies metaboliques : la glycogenogenese (stockage du glucose : sous forme de glycogene), la glycogenolyse (liberation de glucose-1-phosphate) et la neoglucogenese (production de glucose a partir d'acide propionique). Chez les ruminants, le glucose est derive presque exclusivement de la neoglucogenese (Young, 1997). La majorite des glucides contenus dans 1'alimentation sont utilises par les microorganismes colonisant le rumen. Le propionate provenant de la fermentation ruminale chez les vaches est le substrat majeur employe par le foie pour la neoglucogenese (Young, 1997). Les acides amines, le lactate et le glycerol apportent une faible contribution a la synthese du glucose chez les ruminants.
Chez la vache laitiere periparturiente, la demande en glucose croit rapidement suite au velage et pendant les premieres semaines de lactation pour faire face a la grande demande d'energie necessaire a la production laitiere et a la grande quantite de lactose qui entre dans la composition du lait (figure 1.6) (Drackley et al., 2000). Pour chaque kilogramme de lait
produit, une vache depense 72 g de glucose. En consequence, le glucose sanguin de la vache laitiere periparturiente diminue drastiquement apres le velage (Drackley et al, 2000 ; Accorsi
et al, 2005). Cette diminution de glucose est certainement reliee a la diminution de l'apport
alimentaire qui reduit la quantite de propionate disponible pour la neoglucogenese (Doepel et
al, 2002). Ainsi, une demande accrue et une diminution de la disponibilite du glucose sont
responsables du faible taux sanguin de glucose chez la vache periparturiente. Cependant, il est courant de voir un pic de glucose une journee avant ou apres velage possiblement cause par la liberation de glucocorticoi'des ayant un effet catabolique sur les proteines et un effet hyperglycemias, notamment en stimulant la conversion des acides amines en glucose dans le foie (Doepel et al, 2002).
2800
• I I 4 1 « - I I 4 1 4< 4 4 4 4 4 I * t r I I I I I • • i f 11 11
Day relative to parturition
Figure 1.6 : Estimation de la demande globale en glucose compare a la quantite de glucose
ingeree durant la periode peripartum. Les vaches ont ete nourries ad libitum avec une alimentation a grain moyen durant la periode seche (Drackley et al., 2000).
1.4.1.6 Les acides gras
La mobilisation des acides gras est accentuee chez la vache laitiere en debut de lactation. Premierement, parce que l'apport alimentaire diminue autour du velage du a une diminution de l'appetit possiblement causee par la pression du foetus sur le rumen et par les changements hormonaux. Le taux de glucose sanguin diminue beaucoup durant cette periode (Goff et al, 2002). De plus, la mise en place de la lactation necessite egalement une quantite importante d'energie. Tous ces evenements aboutissent une mobilisation des reserves lipidiques afin d'obtenir une source d'energie additionnelle.
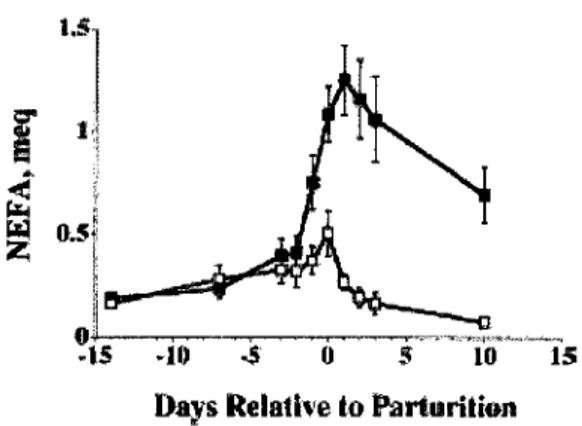
La mobilisation des acides gras se traduit par une serie de reactions biochimiques declenchees par l'activation du systeme de l'adenylate cyclase au niveau des membranes des adipocytes. Le declenchement du systeme amene a l'activation d'une lipase intracellulaire qui catalyse la lipolyse des triglycerides en liberant du glycerol et des acides gras non esterifies (AGNE) dans la circulation. La liberation des AGNE est nettement moins marquee chez les vaches mastectomisees ce qui demontre bien la demande energetique elevee necessaire pour la synthese du lait (figure 1.7) (Goff et al, 2002).
Les acides gras libres dans la circulation peuvent exercer un retrocontrole sur le taux de lipolyse en inhibant l'activite de l'adenylate cyclase. Les acides gras liberes lors de la lipolyse sont captes par differents tissus, mais la majorite est captee par les tissus hepatiques. Lorsqu'ils sont dans le foie, les acides gras libres sont esterifies et incorpores dans les VLDL sous forme de TAG (triacylglycerol). Lorsque la mobilisation des acides gras a partir des tissus adipeux est plus elevee que la capacite du foie a secreter des VLDL, il y a accumulation de TAG dans le foie. Cet etat est connu sous le nom du 'syndrome du foie gras' (Ametaj, 2005). En plus, lorsque les acides gras libres entrants dans le foie sont en exces, en particulier lors d'une grande mobilisation des reserves suite a la mise en place de la lactation, celui-ci
n'arrive pas a tous les esterifier alors une partie est oxydee. L'oxydation des acides gras depend de la capacite du cycle de krebs. L'oxaloacetate semble etre le facteur limitant de ce cycle (Goff et Horst, 1997). Une fois la capacite maximale du cycle de krebs atteinte, les autres acides gras non esterifies ou non oxydes sont utilises pour la formation de corps cetoniques (acetoacetate, |3-hydroxybutyrate). Les corps cetoniques peuvent etre utilises comme sources d'energie par l'animal lorsqu'il y a une carence en glucose, comme en debut de lactation. Chez la vache laitiere, 1'augmentation du BHBA dans le systeme circulatoire est observee environ une semaine suite a 1'augmentation des AGNE dans le plasma ce qui confirme que 1'apparition des corps cetoniques est bien une consequence de la mobilisation des acides gras (Busato et al., 2002).
Des concentrations faibles a moderees d'AGNE ont des effets sur les leucocytes polymorphonuclear (PMN) et sur les cellules mononucleates du sang peripherique (PBMC)
in vitro. Une diminution de la capacite des PMN a produire des radicaux libres lors de la
flambee oxydative est observee en presence d'AGNE (Scalia et al, 2005). Par ailleurs, une etude sur les PBMC bovins in vitro a permis de demontrer que l'ajout d'AGNE inhibe la production d'IgM et d'IFN-Y(Lacetera et al, 2003). Cependant, l'incubation des PMN bovins en presence de AGNE a des concentrations de 0 a 2 mmol/L n'influence pas leur phagocytose (Scalia et al, 2005).
Lorsque les corps cetoniques augmentent dans le lait et tout le systeme sanguin, ils ont pour effet de diminuer l'appetit et la mobilisation des lipides et d'aggraver le deficit energetique. De plus, certaines etudes constatent que les fonctions leucocytaires telles que le chimiotactisme, la phagocytose sont diminuees en presence de corps cetoniques (Suriyasathaporn et al, 2000). Par consequent, les vaches avec des taux eleves en corps cetoniques sont plus susceptibles de developper une mammite a coliforme que les vaches saines (Janosi et al, 2003; Kremer et al, 1993). Ainsi, 1'augmentation des AGNE puis des
BHBA en debut de lactation suite a la mobilisation des gras pourrait etre impliquee dans l'immunodepression des vaches laitieres.
L'accumulation des TAG non secretes sous forme de VLDL dans le foie mene aussi a des problemes metaboliques et plusieurs symptomes semblables a ceux provoques par l'accumulation des corps cetoniques. En plus, l'exces de TAG dans le foie diminue la capacite de synthese de l'uree ce qui provoque une accumulation d'ammoniaque. L'ammoniaque diminue la capacite des hepatocytes a synthetiser du glucose a partir du propionate ce qui accentue le deficit energetique (Drackley, 1999). Done l'excedent des acides gras mobilises entraine des repercussions nefastes sur la sante des animaux ainsi que sur le systeme immunitaire.
1.5-,
Days Relative to Patiurition
Figure 1.7 : Concentration des AGNE plasmatiques chez des vaches intactes (n=8, noir) et
1.4.2 Changements hormonaux
La periode de la parturition se distingue par des variations hormonales importantes. En effet, au moment de la parturition il y a une diminution importante de la progesterone et une augmentation marquee de l'estrogene et des glucocorticoi'des. D'autres hormones non sexuelles varient aussi apres la parturition telle que la leptine et la stanniocalcine. Toutes ces variations d'hormones influencent l'organisme et en particulier le systeme immunitaire. H est done bien important de comprendre ces variations afin d'evaluer les roles de celles-ci dans le phenomene d'immunodepression present chez la vache en debut de lactation.
1.4.2.1 Les estrogenes
Durant les premiers mois de gestation, le taux d'estrogene (estrone El et 17P-estradiol E2) est relativement bas. Vers la mi-gestation, celui-ci monte jusqu'a 300 pg/mL soit environ 10 fois plus que lors du cycle de l'estrogene (Goff et Horst, 1997). La semaine avant le velage, l'estrogene s'eleve jusqu'a une concentration de 2000 pg/mL. Juste avant la mise bas sa concentration (en particulier l'estrone) peut s'amplifier drastiquement jusqu'a des valeurs de 4000-6000 pg/mL (Goff et Horst, 1997). La production d'estrogenes par le placenta est initiee par 1'augmentation de Cortisol secrete par le foetus durant les quelques semaines avant la parturition. Tandis que les estrogenes augmentent, la progesterone chute precipitamment lors de la parturition causee par la regression du corps jaune induit par les prostaglandines qui augmentent durant cette raeme periode (figure 1.8).
Delbecchi et al. (2005) suggere que l'injection de la 17P-estradiol (E2) chez les vaches lactantes tend a augmenter l'incidence de mammites. L'E2 aurait done un role important dans
le systeme immunitaire chez la vache particulierement au niveau de la defense de la glande mammaire. L'E2 influence negativement les fonctions des PMN telles que le chimiotactisme, la phagocytose et la flambee oxydative (Lamote et al, 2004 ; Lamote et al, 2006).
Le mecanisme par lequel l'E2 exerce son effet immunosuppresseur n'est pas connu. Cependant, Winters et al. suggere que les PMNs bovins expriment des recepteurs a estrogene (2003) de type (3 et possiblement des recepteurs a. L'expression des ARNm et proteines correspondants a ces deux recepteurs ont ete retrouves chez les PMN bovins (Lamote et al, 2006). Cependant, aucune difference dans l'expression des recepteurs n'a ete observee en comparant des PMN de fin de gestation (concentration elevee d'estrogene) avec des PMN de debut de lactation (faible concentration d'estrogene).
La meme equipe a aussi observe une diminution de l'expression de CD47 sur les neutrophiles bovins en presence d'E2 (Lamote et al., 2005). La proteine CD47 a ete recemment etudiee comme etant un element important dans la migration des PMN humains a travers les epitheliums intestinaux, mais aucune donnee n'a ete reporte concernant les PMN bovins (Reaves et al, 2004). La diminution de l'expression de CD47 en fin de gestation en presence d'une concentration elevee de E2 pourrait entrainer une diminution dans la capacite des neutrophiles a defendre l'organisme.
Par contre, les travaux de Winters et al. (2003) n'ont demontre aucune influence directe d'E2 sur la flambee oxydative de PMN bovins in vitro. Par ailleurs, l'injection d'E2 augmente l'expression de l'hormone stanniocalcine (STC) dans le plasma et le lait (Delbecchi et al, 2005). La STC aurait peut-etre un effet sur le systeme immunitaire des bovins. Pour l'instant, seules des etudes chez les cellules humaines et murines ont permis d'observer un role potentiel de la STC sur les reponses immunitaires et inflammatoires (Kanellis et al, 2004).
230 240 250 260 27ft im 299
Days After Conception
Figure 1.8 : Concentration plasmatique des estrogenes (cercle) et de la progesterone (triangle)
chez des vaches en fin de gestation. (Goff et Horst, 1997)
1.4.2.2. Stanniocalcine (STC)
La stanniocalcine est une hormone decouverte chez les poissons au 18e siecle, reconnue pour son role dans la regulation du calcium. La STC est liberee dans le sang en reponse a une augmentation de calcium. Chez les poissons, elle agit sur les cellules epitheliales afin de reduire l'acquisition de calcium (Gerritsen et Wagner, 2005). La decouverte de la stanniocalcine chez les mammiferes s'est effectuee beaucoup plus tard (Gerritsen et Wagner, 2005). Par contre, la comprehension de ses mecanismes et de ses roles de la stanniocalcine chez les mammiferes reste incomplete.
Le transcrit de la STC de 247 residus est exprime dans plusieurs tissus chez les mammiferes : rein, muscle du cceur, poumon, thymus, pancreas, thyroi'de, rate et les ovaires. De plus, chez les mammiferes, la production de la STC est purement locale et de type paracrine (Gerritsen et Wagner, 2005).
Un des roles connus de la STC chez les mammiferes est son action dans la stimulation de la mineralisation des os. La surexpression de la STC augmente 1'acquisition de phosphates par les osteoblastes. La STC joue possiblement un role au niveau du cceur et du myocarde. En effet, l'ajout de STC a des cardiomyocytes de rats en culture a permis d'observer une diminution du rythme cardiaque ainsi qu'une diminution du pic de calcium associe a chaque contraction (Gerritsen et Wagner, 2005).
La STC pourrait etre impliquee dans les phenomenes immunitaires et inflammatoires. La STC aurait un role dans le chimiotactisme des macrophages (Kanellis et al, 2004). Le changement de calcium intracellulaire est associe avec 1'activation de diverses kinases et phosphatases qui entrainent differents phenomenes tels que la degranulation et le remodelage de l'actine necessaire pour l'adhesion et la mobilite des macrophages. L'ajout de stanniocalcine in vitro (a partir de 10 ng/mL) semblerait diminuer le taux de calcium et ainsi reduire la capacite chimiotactique des macrophages (Kanellis et al, 2004).
Un effet inhibiteur de la STC sur la migration des macrophages et des lymphocytes T humains au travers d'une membrane endotheliale a ete decrit. Par contre, aucun effet n'est observe chez les neutrophiles et les lymphocytes B (Chakraborty et al, 2007). Probablement que cette distinction entre les cellules est le reflet d'une difference dans l'expression des recepteurs pour la STC.
Chez la vache laitiere, peu d'etudes ont ete effectuees concernant la stanniocalcine. Une etude chez les bovins n'a demontre aucune difference dans la concentration de la stanniocalcine sanguine entre les differents stades de lactation (Miller et ah, 2006). Cependant, chez les vaches multipares, la concentration de la stanniocalcine dans le lait augmente au cours de la lactation (Miller et ah, 2006). Chez les rongeurs, la stanniocalcine augmente durant la gestation et diminue suite a la mise bas (Deol et ah, 2000). H serait done important de verifier les variations de stanniocalcine chez la vache laitiere particulierement en debut de lactation pour verifier si cette hormone aurait un role potentiel dans le phenomene d'immunodepression qui caracterise cette periode.
1.4.2.3 Leptine
La leptine est une hormone qui est secretee par les tissus adipeux et participe a la regulation de l'homeostasie energetique du corps. Le gene de la leptine (ob) est aussi exprime dans d'autres tissus tels que le placenta, les tissus foetaux, la glande mammaire, les muscles et bien d'autres (Chillard et ah, 2001). Elle est secretee de facon pulsatile chez les ruminants, mais sans effet diurne marque (Ingvartsen et Boisclair, 2001). Son role le plus connu est le controle de la prise alimentaire en se fixant a l'hypothalamus au niveau des domaines Ob-Rb sur les recepteurs a leptine. On observe que la sous-nutrition a pour effet de diminuer le taux de leptine circulant afin de creer un signal de faim et ainsi d'augmenter la prise alimentaire. Chez les ruminants, on observe que l'injection de leptine provoque des effets de satiete (Ingvartsen et Boisclair, 2001). De plus, sa concentration systemique est en etroite correlation avec la taille des adipocytes bovines et fait done de la leptine un bon indicateur de l'adiposite animale (Chillard et ah, 1999). En plus de son implication dans la prise alimentaire, la leptine joue un role tres important au niveau du systeme immunitaire. L'action de la leptine sur les cellules immunitaires est enclenchee par des recepteurs Ob-Rb. L'expression de ce recepteur est retrouvee sur les lymphocytes (CD4+, CD8+), les cellules CD34+, les cellules sanguines
mononucleates, les PMNs et les cellules souches hematopoi'etiques (Invargesten et Boisclair, 2001). L'expression du recepteur de la leptine sur ces cellules a ete confirmee autant chez l'humain que chez les ruminants. Peu d'etudes quant a son action immunitaire chez le bovin ont ete effectuees. Cependant, il a ete repertorie que les souris deficientes en leptine eprouvent des desordres immunitaires importants. Notamment, on observe une diminution des lymphocytes circulants, une anomalie dans l'immunite cellulaire des lymphocytes T (Fantuzzi et Faggioni, 2000) ainsi qu'une diminution de la proportion des macrophages phagocyte actif. Chez les humains, on observe une concentration plasmatique de leptine superieure chez les gens qui survivent a des septicemics ce qui suggere que la leptine joue un role important dans la defense du systeme.
Diverses etudes in vitro ont aussi demontre le role important de la leptine sur les parametres immunitaires. Tout d'abord, les neutrophiles murins ont leur capacite de phagocyter
Klebsiella pneumonia attenuee en absence de leptine. Cet effet est renverse par l'ajout de
leptine exogene (Moore et ah, 2003). En plus, elle regule la synthese de plusieurs cytokines. La presence de leptine augmente la concentration d'IL-2 et EFN-yet reduis la concentration de IL-4. Cette regulation se repercute sur le rapport Thl-Th2. De plus, la leptine favorise in vitro la production d'une chimiokine par les cellules endotheliales (HUVEC) agissant sur les monocytes (MCP-1). En prime, la leptine possede un effet synergique in vitro avec le facteur de cellules souches pour stimuler la proliferation des progeniteurs hematopoi'etiques (Invargesten et Boisclair, 2001).
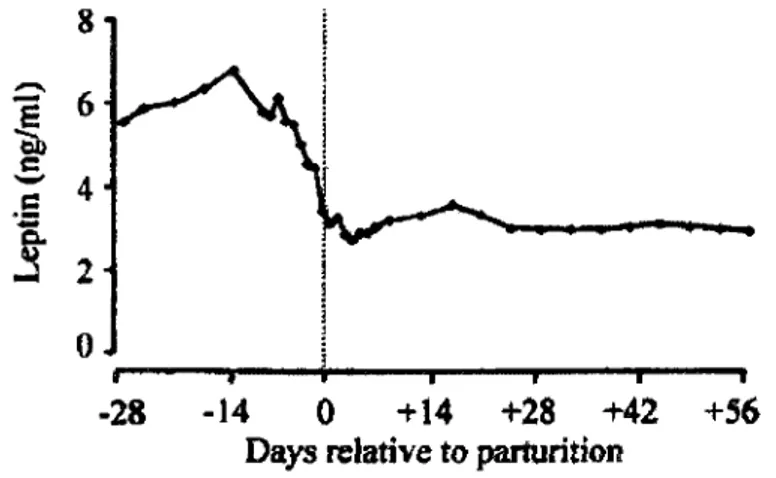
Chez la vache laitiere, le taux de leptine augmente generalement au cours de la gestation. Le placenta semblerait etre a l'origine de cette augmentation. Par contre, quelques jours avant le velage, le taux de leptine se met a diminuer (figure 1.9) (Block et ah, 2001 ; Accorsi et ah, 2005). La diminution post-partum est de l'ordre de 50% par rapport au taux pre velage (Block et ah, 2001). Selon cette meme etude, ce declin serait essentiellement du a la
diminution du taux de synthese de leptine causee par l'hypotrophie des adipocytes (Block et
al, 2001). Le bilan energetique negatif est la cause principale de la diminution des reserves
adipeuses apres la parturition causant ainsi un abaissement de la production de la leptine. La baisse de la leptine durant cette periode conduit a divers problemes de fertilite. Suite a la parturition, chez les vaches, une correlation importante entre la leptine et la secretion pulsatile de l'hormone lutei'nisante (LH) semblerait expliquer le delai avant le retour des premieres ovulations post-partum (Kadokawa et al., 2006).
Apres la parturition, la vache laitiere souffre d'une immunodepression temporaire et d'un deficit energetique important. La cause de cette immunodepression n'est pas entierement connue a ce jour. Une etude a permis de constater que chez des souris souffrant d'immunodepression causee par une sous-alimentation, une administration de leptine renverse le phenomene (Lord et al., 1998). Des etudes plus approfondies chez les bovins laitiers permettraient de verifier le role possible de la chute de la leptine dans 1'immunodepression des vaches en debut de lactation.
|
1
! i 1 I " " " « i ' I I ' I I -28 -14 0 +14 +28 +42 +56 Days relative to parturitionFigure 1.9 : Les changements de la concentration plasmatique de la leptine durant la periode periparturiente chez la vache laitiere multipare. (n=8) (Block et al, 2001)
•2
6-e
s
1.5 Dysfonctions immunitaires et phenomene d'immunodepression
Chez la vache laitiere en debut de lactation, on observe un phenomene d'immunodepression caracterise par une augmentation de l'incidence des infections (Kehrli et ah, 1999 ; Mallard et
al., 1998). Un grand nombre de cas de mammites surviennent durant les premiers 70 jours de
lactation (Kehrli et Harp, 2001). Le phenomene d'immunodepression se caracterise par diverses dysfonctions qui affecteraient les deux grandes categories du systeme immunitaire : l'immunite innee (non specifique) et l'immunite acquise (specifique)
1.5.1 Immunite innee
L'immunite innee est la premiere ligne de defense contre les pathogenes. Les neutrophiles, les macrophages, les cellules natural killer (NK), le complement (voie alterne) et certains autres facteurs solubles font partie de l'immunite innee.
Les leucocytes polymorphonucleaires (PMN) sont reconnus comme etant l'element cle du systeme immunitaire inne et ils agissent comme premiere ligne de defense en cas d'invasion par un pathogene. Les neutrophiles sont presents dans le sang sauf en cas d'infection ou ils sont recrutes sur le site infectieux. Ce recrutement s'effectue par la reconnaissance des PMN sanguins sur l'endothelium via l'expression de molecules d'adhesions tant sur les neutrophiles (L-selectine, CD11, CD18) que sur l'endothelium (ICAM-1, ICAM-2, ELAM-1). Ensuite, les neutrophiles quittent la circulation sanguine par diapedese. Tout ce processus de recrutement s'effectue grace aux agents chimioattractants tels que les fragments du complement C5 et les interleukines 1,2 et 8 attirent les PMN vers le site infectieux. Les PMN utilisent leur capacite de phagocytose pour 1'elimination du pathogene. L'opsonisation par les immunoglobulines ou
par les proteines du complement augmente grandement 1'efficacite de la phagocytose. Une fois rendu dans le phagosome, le pathogene est elimine soit par digestion par les enzymes lysosomales ou par l'attaque des radicaux libres.
A la parturition, les mecanismes de recrutement des PMN et de destruction du pathogene semblent compromis (Newbould, 1976 ; Guirdy et al, 1976). L'expression de la L-selectine diminue contribuant ainsi a reduire 1'efficacite du recrutement des PMN aux sites infectieux (figure 1.10) (Monfardini et al, 2002 ; Kimura et al, 1999). Apres la parturition, le chimiotactisme chez les neutrophiles semble etre diminue en particulier chez les vaches ayant une retention placentaire. Apres la mise bas, le placenta doit etre reconnu par le systeme immunitaire afin d'etre expulse. La retention placentaire pourrait done etre une consequence d'une dysfonction immunitaire au niveau du recrutement des leucocytes chez les vaches laitieres les plus affectees par l'immunodepression post-partum (Kimura et al., 2002 b). En plus des moyens de recrutement, les moyens d'elimination des pathogenes sont aussi alteres. La capacite des neutrophiles a phagocyter des bacteries {Staphylococcus aureus) est diminuee de 11 % jusqu'a 2 semaines apres la parturition (Kehrli et al., 1989 b). De plus, la flambee oxydative des neutrophiles caracterisee par la liberation de plusieurs radicaux oxygenes est aussi affectee. Les neutrophiles du lait et du sang ont une activite de flambee oxydative beaucoup plus faible en periode periparturiante. On note une diminution de la flambee oxydative une semaine avant la mise bas. Le retour a une activite normale s'effectue generalement 10 jours a 2 semaines apres le velage (Hoeben et al, 2000 ; Moreira Da Silva et
al, 1998 ; Kehrli et al, 1989 b). Une autre facon d'evaluer la flambee oxydative est la mesure
de 1'activite de la myeloperoxidase. L'activite de cette enzyme est diminuee environ 20 jours avant la parturition. Cette diminution perdure souvent jusqu'a 3 semaines apres le velage (figure 1.11) (Kimura et al., 1999). Le systeme immunitaire inne est done grandement affecte par l'immunodepression caracteristique des vaches laitieres en debut de lactation notamment au niveau des neutrophiles dont le recrutement et la capacite d'elimination des pathogenes sont affaiblis.
100 90 ] 80 o C/3 60 5<H 40
i-M
-28 -21 -14 -7 0 7 14Days Relative lo Parturition
28
Figure 1.10 : Moyenne de l'expression de la L-selectine sur les neutrophiles actives par
l'activateur de plaquettes chez des vaches intactes (n=8, noir) et des vaches mastectomisees (n=10, blanc), durant la periode entourant la parturition. L'expression de la L-selectine est mesuree en fonction de l'intensite de fluorescence d'un anticorps fluorescent anti-L-selectine lie a la surface des neutrophiles (Kimura et al., 1999).
)50n
2 5 - I — ' — • , . . . , . — . • . . . . • . . - . . .T. • . , . . , . . . — , - , - . - . . . - — • i — i — • — • — . . . .r. . . ....,•.. 1
-2S -21 -14 -? 0 7 14 *l 2ft
Days Relative to Parturition
Figure 1.11: Moyenne de l'activite de la myeloperoxidase chez des vaches intactes (noir) et des vaches mastectomisees (blanc), durant la periode entourant la parturition (Kimura et al,
1999).
1.5.2 Immunite acquise
L'immunite adaptative specifique survient apres l'arrivee des facteurs et des cellules de l'immunite innee. Ce type de reponse permet la reconnaissance de determinants antigeniques. L'immunite permet une reponse selective ainsi qu'une memoire immunologique permettant une meilleure elimination du pathogene lors d'une deuxieme rencontre. Les lymphocytes, les cellules presentatrices d'antigene ainsi que les anticorps et le complement (voie classique) sont les elements cle de la reponse immunitaire acquise.
Le systeme immunitaire adaptatif comprend les lymphocytes. Ceux-ci sont capables de reconnaitre differents antigenes grace a des recepteurs specifiques a leur surface. Deux groupes de lymphocytes sont particulierement affectes par l'immunodepression en debut de lactation soit les lymphocytes T et B. Les lymphocytes T sont subdivises en plusieurs types : les T cytotoxiques, les T auxiliaires et les T regulateurs. Les cellules T cytotoxiques (CD8) reconnaissent les antigenes presentes via le complexe majeur d'histocompatibilite de classe I et eliminent les cellules etrangeres ou infectees par un pathogene intracellulaire. Les lymphocytes auxiliaires (CD4) secretent des cytokines en reponse a une stimulation avec un complexe antigene et complexe majeur d'histocompatibilite de classe II permettant de diriger ainsi la reponse vers le type humoral ou cellulaire. Quant a eux, les lymphocytes regulateurs sont specialises dans la regulation des cellules immunitaires et dans le controle des reactions auto-immunes. Suite a une stimulation par des cytokines, les lymphocytes B eux peuvent produire des anticorps specifiques diriges contre les pathogenes. II existe plusieurs types d'immunoglobulines secretees par les lymphocytes B : IgD, IgM, IgG, IgA, IgE.
Les lymphocytes semblent voir leur activite modifiee durant la periode entourant la parturition. Tout d'abord, la capacite de proliferation des lymphocytes est grandement diminuee la semaine suivant la parturition (Lacetera et al, 2005 ; Kehrli et al, 1989, a). En plus, le nombre de lymphocytes sanguins decline durant cette meme periode. Selon une etude realisee par Kimura et al. en 2002 a, le nombre de lymphocytes (CDS, CD4, CD3, Ty8) diminue 27 jours avant la parturition et dans certains cas reste sous la normale jusqu'a 3 semaines apres le velage (figure 1.12). Par contre, aucune difference n'est observee dans le nombre de lymphocytes B circulants (Kimura et al, 2002 a). Bien que le nombre de lymphocytes B durant la periode peripartum ne semble pas varier, leur capacite a produire des anticorps est affectee. Une diminution des IgM produits par les lymphocytes B lors d'une stimulation in vitro est observee une semaine avant jusqu'a une semaine apres la parturition (Nonnecke et al, 2003). Le systeme immunitaire acquis est done affecte par l'immunodepression caracteristique des vaches laitieres suite a la parturition.
?27 -18 -9 0 9 18 27 ?27 -18 -9 0 9 18 27 Days Relative to Parturition Days Relative to Parturition
Figure 1.12 : Moyenne des pourcentages des lymphocytes totaux sanguins (a), Thelper (b),
T-cytotoxique (c), T gamma-delta (d) dans le sang de vaches intactes (n=8, noir) et mastectomisees (n=10, blanc) durant la periode periparturiante (Kimura et ah, 2002 a)
1.5.3 Cytokines
En plus d'une modulation de l'activite des neutrophiles et des lymphocytes, la periode entourant la parturition semble etre caracterisee par des fluctuations au niveau de diverses cytokines dans le sang et le lait. Tout d'abord, l'activite transcriptionnelle de l'interleukine-10 (IL-10) dans les cellules epitheliales de la glande mammaire est beaucoup plus importante en
debut de lactation qu'a la mi-lactation ou encore qu'a la fin de la lactation. De plus, son activite predomine par rapport a celle d'autres cytokines telles que le TNF-a et l'IL-8 (figure 1.13) (Britti et al, 2005). Ceci est probablement du au fait qu'en debut de lactation une majorite de T-auxiliaire de type Th2 predomine dans le lait ce qui augmenterait la secretion de lTL-lO (Shafer-weaver et al, 1999). Cette cytokine est reconnue pour etre immunosuppressive en affectant les cellules de l'immunite (Weiss et al, 2002 ; Sweeney et
al, 1998; Yoshihara et al, 2007). Elle est aussi reconnue pour supprimer la production de
1'IFN-Y ainsi que pour diminuer le recrutement et l'activation des neutrophiles (Shafer-weaver
et al, 1999). L'augmentation de l'IL-10 dans le lait en debut de lactation pourrait etre l'un
des elements impliques le phenomene d'immunodepression et contribuer a l'incidence des infections intra mammaires.
La cytokine pro-inflammatoire TNF-a augmente a la parturition possiblement suite a une augmentation importante du Cortisol. La production de cette cytokine est influencee par la parite des vaches (Lessard et al, 2004). Les cellules des vaches multipares produisent moins de TNF-a en presence de LPS que celles des vaches primipares (Lessard et al, 2004). En plus, en presence de serum autologue, les PBMCs des vaches periparturientes produisent plus de TNF-a qu'en presence de serum bovin foetal (Lessard et al, 2004). Ceci indique que les substances presentes dans le serum apres la parturition exercent une influence sur la production de cette cytokine. De plus, une concentration trop elevee du TNF-a semble etre associe avec le syndrome du foie gras observe chez les vaches en debut de lactation. L'augmentation du TNF-a dans l'organisme diminue l'appetit et accentue le deficit energetique ce qui a pour effet d'augmenter la mobilisation des acides gras (Ametaj, 2005). A la parturition, l'augmentation de la cytokine TNF-a plasmatique observee (Rontved et al, 2005) entraine differents problemes de sante pour les vaches laitieres.
Early-lactation Mid-laclsitioit Late-lactation
Figure 1.13 : Activite transcriptionnelle de TNF-alpha, IL-8 et EL-10 en debut, milieu et fin
de lactation dans les cellules epitheliales de glande mammaire de vaches (Britti et ah, 2005).
1.6 Les causes de l'immunodepression
Les causes de 1'immunodepression sont multiples et ne sont pas encore pleinement connues.
1.6.1 Deficit energetique
L'incidence de mammites est particulierement elevee durant la periode post-partum au meme moment ou la vache est en deficit energetique. Aucune indication jusqu'a present n'a permis de relier directement le deficit energetique avec l'immunodepression survenant apres la parturition. Par contre, on peut suspecter grandement un lien entre les deux phenomenes.
de l'organisme afin de supporter un etat physiologique ou une fonction prioritaire. Par exemple, la vache laitiere en debut de lactation ne mange pas suffisamment pour supporter les besoins energetiques necessaires a la production de lait. Durant cette periode, la production de lait est prioritaire par rapport aux autres fonctions de 1'organisme. On peut alors supposer que la defense immunitaire de l'organisme fait parti des fonctions non prioritaires durant cette periode. D'ailleurs, chez les vaches mastectomisees, on observe une absence de deficit energetique et le phenomene d'immunodepression est presque inexistant (Kimura et al, 1999; Kimura et al, 2002 a,b; Nonnecke et al, 2003). De plus, la balance energetique negative entraine plusieurs repercussions sur l'utilisation des reserves d'energie pouvant elle-meme avoir des effets sur le systeme immunitaire. Le deficit energetique en debut de lactation contribue surement a la reduction de l'efficacite des defenses immunitaires de 1'animal.
1.6.2 Changements hormonaux
La quantite d'estrogenes et de Cortisol dans le sang augmente a la fin de la gestation. Ces hormones sembleraient influencer negativement le systeme immunitaire. Certaines etudes ne montrent pas d'effet des estrogenes sur les cellules immunitaires, mais la majorite semble dire que les estrogenes ont un effet suppressif important sur l'immunite (voir la section sur les changements hormonaux). De plus, les glucocorticoi'des tels que le Cortisol sont reconnus pour leur effet immunosuppressif (Griffin, 1989). La hausse des estrogenes et du Cortisol en fin de gestation pourrait bien etre une des causes de l'immunodepression des vaches laitieres. Cependant, aucune etude ne permet pour 1'instant de determiner 1'importance in vivo reelle de ces hormones dans l'immunodepression en periode peripartum.
Une autre hormone possiblement impliquee dans le phenomene d'immunodepression est la leptine. La leptine diminue suite a la parturition et elle possede un role important dans le
maintien du systeme immunitaire. Une chute de la leptine en debut de lactation pourrait entrainer une deterioration du systeme immunitaire. Les changements hormonaux durant la periode peripartum pourraient §tre une des causes dans l'immunodepression en debut de lactation.
1.6.3 Les metabolites et vitamines
Durant la periode de la parturition et celle du debut de la lactation, les vaches subissent plusieurs changements metaboliques. On observe plusieurs changements des metabolites resultants de la mise en place de la lactation. La hausse notable de certains metabolites tels que les AGNE et le BHB pourrait aussi etre impliquee dans la diminution des capacites du systeme immunitaire.
En debut de lactation, une majeure partie des vitamines est sequestree pour entrer dans la composition du colostrum. Une diminution des vitamines A et E (38% et 47%) ont ete constatees chez les vaches laitieres en debut de lactation (Goff et Stabel, 1990 ; Goff et ah, 2002). De plus, des supplements en vitamines A et E durant cette periode sembleraient ameliorer les reponses immunitaires (Daniel et ah, 1991; Politis et al., 2004). Une autre cause non negligeable de l'immunodepression pourrait etre la diminution des vitamines dans l'organisme.
Les causes de l'immunodepression des vaches laitieres en debut de lactation semblent etre multiples et reliees entre elles. Par exemple, un deficit energetique cause par une grande production de lait entraine une mobilisation des reserves lipidiques ce qui contribue a augmenter les concentrations circulantes de AGNE et de BHBA et a diminuer la
concentration sanguine de leptine. Done, le phenomene de depression du systeme immunitaire durant cette periode est probablement relie a un ensemble de facteurs dont la somme entraine des problemes de sante tout en augmentant l'incidence de diverses infections.
1.7 Reduction de 1'immunodepression
Pour contrer ce phenomene d'immunodepression, quelques etudes sur la nutrition en periode
peripartum ont ete effectuees. Premierement, une etude basee sur l'augmentation de la valeur
energetique de rations durant les 8 dernieres semaines de gestation n'a pas eu d'impact important sur le systeme immunitaire autant avant qu'apres la parturition. (Meglia et al, 2005). Une autre etude effectuee pour ameliorer le systeme immunitaire des vaches en lactation a permis de demontrer une absence d'effet de 1'alimentation supplements en glutamine sur le phenomene d'immunodepression (Doepel et al, 2006). De plus, un lien entre la prise d'une grande quantite d'antioxydant et les fonctions des neutrophiles a ete examine dans une etude ou les vaches recevaient de la vitamine C, un antioxydant hydrosoluble chez les mammiferes, avant et apres la parturition. L'acide ascorbique est reconnu pour augmenter lors de 1'activation des neutrophiles pour prevenir les dommages causes par les radicaux oxygenes. Cependant, l'ajout de la vitamine C alimentaire n'a eu aucun effet sur la capacite de phagocytose des neutrophiles des vaches laitieres periparturientes (Weiss et Hogan, 2007). La vitamine E quant a elle, lorsqu'administree oralement 4 semaines avant la parturition jusqu'a 12 semaines apres et apporte un effet positif sur les neutrophiles. En effet, la semaine avant et apres la parturition la production de superoxydes est superieure chez les neutrophiles stimules et isoles des vaches supplementees en vitamine E (Politis et al, 2004). Cependant, les taux de production de superoxydes demeurent tout de meme relativement plus faibles que ceux obtenus a la 4e semaine de lactation. Jusqu'a present, les moyens pour reduire 1'immunodepression par l'alimentation ou les supplements alimentaires ne sont pas tres efficaces.