L
a
tomate
est, après
la
pomme de
terre,
le premier légume
consom-mé dans
le
monde
. Elle est adaptée
à
des
conditions
de culture très
variées
et
dest
inée
à
la consommation en
frais ou
à
la t
r
ansformat
i
on in
dustrielle.
La
sé
lect
i
on
s'est attachée,
depuis les
années
40,
à
créer
des var
iétés adaptées
spécifiquement à
diverses
conditions, en
amé
l
iorant
simultanément
la
q
u
alité des
fruits
et
le
niveau de résistance
aux
pat
h
ogènes [
l
]. Pour
cela,
les
ressources
génétiques
ont
été
largement exploitées
et
le recours
aux espèces sauvages
appa-rentées
à
la tomate
cultivée a été
fré-quent, notamment pour
rechercher
des
gènes
de résistance
aux
pathogènes. Les
croisements encre
tomates cult
ivées et
sauvages sont
relativement
aisés.
Il
faut
él
iminer, dans
les
descendances,
les
caractéristiques
défavorables parfois liées
au gène
d
'
intérêt.
Cette étape
peut
s'avé-rer longue
et
diffici
le.
Les marqueurs
moléculaires,
en
permettant d'identifier
les régions c
h
romosomiques porteuses de
gènes cibles
puis de
suivre
leur
introduc-tion dans des
variétés él
ites,
sont venus
apporter
un regain d
'
intérêt pour
ces
res-sources.
Grâce
aux avancées
récente
s
de
M. Causse, C. Caranta, V. Saliba-Colom-bani, A. Moretti, R. Damidaux, P. Roussel-le: INRA, Station de génétique et amélio-ration des fruits et légumes, BP 94, 84143 Montfavet cedex, France.
Tirés
à
part: M. CausseRessources génétiques
Valorisation des ressources
génétiques de la tomate
par l'utilisation
de marqueurs moléculaires
Math
il
d
e
Caus
s
e
,
Ca
r
ole Ca
ra
n
ta,
Vera Sal
i
ba
-
Colomban
i
,
Andr
é
More
tt
i, René Dam
i
dau
x,
Patr
i
c
k
Rousselle
la biologie moléculaire,
la cartographie
chromosomique
de nombreux gènes
majeurs
et
de
gènes
à
effets
quantitatifs
ai~si
que leur clonage
ont
pu
être
entre-pns.
Ressources
génétiques
disponibles
pour l'amélioration
de la tomate
Espèces sauvages
apparentées
à
la tomate
La tomate
appartient au genre
Lycopersi-con
de la famille des So
lanacées.
Ce
genre comprend neuf
espèces
[2], toutes
originaires de la région
andine
du Pérou
et
de l'Équateur,
à
l'exception
de
L.
cheesmanii
que l'on trouve dans l'archipel
des Galapagos
(tableau
1)
.
Ces espèces se
dist
i
nguent par
la couleur
des fruits
à
mat
u
rité, le nombre de feuilles
entre
les
bo
u
quets floraux
et
le
mode de
repro-duction. D'autres
critères
floraux
com-p
lètent
la classification. La caille du fruit
ne dépasse jamais
2-3 cm chez
les
espèces sauvages
(photo
1).
La tomate
cultivée
Lycopersicon
esculen-tum
a été
introduite
en Europe
au
XVTesiècle,
depuis le Mexique.
Elle
a été
domestiquée
à
partir de la forme
L.
escu-Cahiers Aqricultures
2
000
;
9: 1
9
7-2
10
lentum
var.
cerasiforme,
puis a migré de
son centre
d'origine andin vers le nord.
Les
croisements entre
l
a tomate cultivée
et
les formes
sauvages
sont plus ou
moins
facil
es.
L.
esculentum
doit
être
uti-lisée
comme
parent femel
l
e dans cous
les
cas et
le recours
à
la po
ll
in
i
sation par un
mé
lange
de po
llen
(pollen de la tomate
et
po
ll
en
du parent
sauvage) est
fré-quent. Le
sauvetage
d'embryons
in vitro
est
nécessaire
avec
L.
chilense
et
L.
per-uvianum.
Les hybrides obten
us
sont
fré-quemment
aucoscéri
les
et doivent
être
rétrocroisés
ou intercroisés pour obtenir
les générations suivantes.
Quatre
espèces
du
genre
Solanum,
regroupées dans la
«série
juglandifolia
»(S
.
lycopersicoides,
S.
juglandifolium,
S.
ochranthum
et
S. sitiens),
présentent de
fortes analogies morp
h
o
l
ogiq
ues
et
chro-mosomiques
avec
les
espèces du genre
Lycopersicon
(espèces
diploïdes
avec
un
nombre de
chromosomes
de n
=
12).
Des hybr
i
des F
Isont aisément
obtenus
par
croisements sexués
avec
S.
lycopersi-coides,
espèce
intéressante pour ses
carac-téristiques de tolérance au froid,
ses
résis-tances au
virus
de
l
a mosaïque du
concombre (CMV),
à
Clavibacter
michi-ganensis
et
à
Botrytis
cinerea
.
Cependant,
la
stéril
ité
et
des prob
lèmes
d'incompati-bi
l
ité
entravent
l
e process
u
s de
rétrocroi-sem en
c. Des descendan
t
s dip
l
oïdes,
aneuploïdes et
po
l
yploïdes ont
été
obte-nus
en
fécondant des hybrides
intergéné-riques
sesquidiploïdes (deux
génomes de
L.
esculentum
et
un
génome
de
S.
lycoper-sicoides)
avec
du po
ll
en
provenant de
l
ignées dérivées de
L.
pennellii
[3]. Il a
Tableau 1
Caractéristiques des neuf espèces sauvages du genre
lycopersicon
Espèce Couleur Nombre Compatibilité Croisement
du fruit de feuilles et mode de avec L.
esculentum
à maturité entre reproduction (1)bouquets
L.
esculentum Rouge 3 AC,S Facile,var. cerasiforme si
L.
esc. femelleL.
pimpinellifolium Rouge 3 AC, SF Facile,si
L.
esc. femelleL.
cheesmanii Orange 3 AC,S Facile,si
L.
esc. femelleL.
hirsutum Vert 3 Al, FO Facile, siL.
esc. femelle; germination difficile et stérilité F1L.
parviflorum Vert 2 AC,S Facile, siL.
esc. femelleL.
chmielewskii Vert 2 AC, F Facile,si
L.
esc. femelleL.
chilense Vert 2 Al,0
Embryon F1 in vitro; F1 autostérilesL.
peruvianum Vert 2 Al,0
Embryon F1 in vitro; F1 autostérilesL.
pennellii Vert 2 Al,0
Facile,si
L.
esc. femelle1) AC: autocompatible; Al: auto-incompatible; S: autogame; 0: allogame; F: facultatif.
Characteristics of the nine wild Lycopersicon species
Répartition géographique
Écotypes très variables; de l'Équateur au Pérou
Vallées côtières du Pérou
Archipel des Galàpagos
Grande aire de répartition, de 500 à 3 300 m d'altitude, en Équateur et au Pérou
Centre du Pérou, à mi-altitude
Centre du Pérou, à mi-altitude
Zones sèches ou temporairement sèches,
le long de la côte du Pérou et au nord du Chili
Zones sèches ou temporairement sèches, le long de la côte du Pérou et au nord du Chili Zones sèches de la partie centrale du Pérou (à mi-hauteur de la façade ouest
de la cordillère des Andes)
Photo 1. Fruits mûrs de différentes espèces du genre Lycopersicon : de haut en bas et de gauche
à
droite: L. peruvianum, L. hirsutum,L. esculentum, L. pimpinellifolium, L. chilense, L. cheesmanii (photo A. Moretti).
Photo 1. Mature fruit of Lycopersicon species: L.
peruvianum, L. hirsutum, L. esculentum, L. pimpi
-nellifolium, L. chilense, L. cheesmanii.
lignées d'addition monosomiques
pos-sibles, cette collection étant
la
première
chez
le genre
Lycopersicon.
Diversité moléculaire au sein
du genre
Lycopersicon
Si
la
mmate
cultivée
présente
une assez
grande
diversité morphologique, sa
diversité moléculaire,
étudiée notamment
par RFLP,
est
limitée
[
4].
Les espèces
sauvages
représentent un
véritable
réser-voir
de va
riabilité, avec
tOutefois
de
s
potentiels
variables
suivant
les
espèces.
La figure
1 montr
e ainsi
que
les
espèces
autogames
présentent
une
diversité
très
inférieure
à
cell
e
des
espèces
allogames.
Miller
et Tanks
ley
[4]
observent
,
par
exemple,
plus de
variabilité
moléculaire
au sein
d'une
accession
de
L.
peruvia-num
que dans
l'ensemble
des
33
acces-sions
de
L.
esculentum et
de
L.
pimpin-nellifolium
qu'ils
ont étudiées.
Enfin,
si
les variétés
modernes
sont
homogènes,
les
cultivars anciens
présentent
une
varia-bilité interne non
négligeab
le.
Collections publiques
de ressources génétiques
de tomate
Charles
Rick, de
l'Université
de Davis
en
Californie, a été
un
pionnier de
la
géné-tique de
la
tomate.
Il
a
prospecté,
à
par-tir de
1948,
dans
la zone
d'origine
de
la
tomate
et constitué
une
importante
col-lection de
ressources génétiques. Cette
coll
ection, conservée au
CM
Rick
Tomato Genetics Resource Center,
ou
TGRC, est
riche
de plus de
3
000
acces-sions,
dont un millier de
lots
d'espèces
sauvages,
877
mutants monogéniques et
un millier de
ressources
diverses. Une
publication
annu
elle (Tomato Genetics
Cooperative
Report,
http:
//genome.cor-nell.
ed
u/ so
lgenes/ abou tso
lgenes.html)
dresse régulièrement
l'inventaire
de
cette
collection et
des travaux récents sur
la
génétique
de
la tomate
.
La
collection
du
ministère de
!'Agriculture
des
États-Un
is
(USDA)
multiplie des
variétés
tradition-nelles du monde
entie
r
et vient comp
lé-ter
la collection
du
TGRC.
La
coll
ection
de
l
'
Asian Vegetable
Research
and
Deve-lopment Center (A VRDC)
à
Taïwan est
la
plus riche
en
nombre de lots. La
col-lection russe de l
'
institut
Vavilov est
éga-lement très riche
en espèces sauvages.
En Europe,
plusieurs
collections
publiques
sont
maintenues,
notamment
L. peruvianum +
L. chilense
L. pennellii
aux
Pays-Bas
(Wagen
ingen), en
Alle-magne
(Gacersleben) et en
Hongrie.
Cette liste
n'est pas
exhaustive. En
Fran-ce,
!'
INRA maintient
à
Montfavet
une
collection d'environ
1
000
gé
notypes.
L'INRA
et
le GEVES se sont
associés
à
huit
sociétés
de
sélection, en
liaison
avec
le
Bureau des
ressources génétiques,
pour
multiplier
et
décr
ire c
h
aq
ue a
nné
e
1 OO
génotypes
de
cette co
ll
ection
.
Chaque partenaire
assure
la
multiplica-tion de dix
génotypes,
tandis que
!'INRA
assure
le stockage
des
graines,
leur
distri-bution,
la
description des
lors et
la
ges-tion du
catalogue.
Rôle essentiel
de quelques gènes
.
ma Jeurs
La tomate
est
l'une
des
espèces végétales
chez
qui
le
nombre de
caractères
mono-géniques exploités
dans
les variétés
culti-vées est
le plus important. Plusieurs de
ces caractè
res résu
lte
nt
de mutations
apparues spontanément
dans des
culti-vars ou
découvertes dans
les espèces
apparentées
à
la romace cultivée.
Les
tableaux
2
et
3
présentent
les
principaux
gènes
majeurs utilisés dans les
variétés
de
romate ou
potentiellement intéressants.
Ils
concernent
les résistances aux
patho-gènes,
l'architecture
de
la
plante
,
la
cou-leur,
la composition ou
la durée de
conservation
des frui
es.
Cahiers Agricultures
2000
;
9
:
197-
210
L. pimpinellifolium
L. hirsutum
Figure 1. Pour
centa-ge de fragments de
restriction révélés par
RFLP, uniquement
trouvés dans chaque
taxon et non
retrou-vés dans les cultivars modernes. L.
escu-/entum représente
des accessions de L.
escu/entum var. cera-siforme et trois cu
lti-v ars traditionnels
p respectés au Pérou,
en Equateur et au
Mexique (d'après Mil-ler et Tanksley [4]).
Figure 1. Percentage
of restriction fragment length polymorphisms found in each taxon but not in modern L. esculentum cultivars.
Pour la construction
d'hybrides résistants
aux maladies
et parasites animaux
Parmi
les
quelque
200
agents
pathogènes
qui
attaq
uent
les
différents types de
culture
de tomate
à
travers
le
monde,
environ
la
moitié
sont
des para.sites
d'ori-gine
tellurique tels
que
les champignons
des
genres Fusarium, Verticillium,
Pyreno-chaeta,
les
bactéries du
genre Ralstonia
ou
encore
les
nématodes
à
galles
Meloidogy-ne.
Pour
lutter
contre ces
nombreux
pathogènes, des
sources
de résistances
génétiques ont été
recherchées
et
exploi-tées
à
partir des
années
40.
Toutes ces
résistances sont
issues des
espèces
sau-vages et
la
plupart
sont
monogéniques
dominances (tableau
2).
Certaines
pro-viennent d'espèces difficilement
compa-tibles
avec
la
tomate
et ont
donc
deman-dé de
longs efforts
pour
être
introduites
dans
des
lign
ées
utilisables par
les
sélec-tionneurs.
Il
s'agir
notamment des
résis-tances au
virus
de
la
mosaïque du
tabac
(TMV) et aux
nématodes, issues de
l'espèce
L.
peruvianum.
L'introduction
des caractères de
résistance
peut
se
limiter
au stade
de
l'hybride
interspécifique. Par
exemple,
les hybrides
Mogéor et
Beaufort
sont
issus
de
croisements entre
une
toma-te
porteuse des
gènes Ve,
!,
1-2,
Fr,
Tm-2
2
et Mi et
un
génotype
de
L.
hirsutum
possédant
une
résistance dominante
au
Pyrenochaeta
et à Didyme/la lycopersici.
Ces
hybrides
sont
utilisés comme
Tableau 2
Caractéristiques des résistances aux principaux pathogènes de la tomate
Agents Pathogènes Gènes Cloné Chromosome Origine Niveau Contourné
Virus
Tobacco Mosaic Virus (TMV)/ Tm-1 2 L. hirsutum Partiel
Virus de la mosaïque du tabac Tm-2, Tm-22 9 L. peruvianum Très élevé
Potato Virus Y (PVY)/ 2 gènes En cours L. hirsutum Très élevé
Virus Y de la pomme de terre
Tobacco Etch potyVirus (TEV) 1 gène En cours L. hirsutum Très élevé
Cucumber Mosaic Virus (CMV)/ Polygénique Plusieurs espèces Partiel
Virus de la mosaïque du concombre
Alfa/fa Mosaic Virus (AMV)/ Am En cours L. hirsutum Très élevé
Virus de la mosaïque de la luzerne
Tomato Spotted Wilt Virus (TSWV)/ Sw-5
*
9 L. peruvianum Très élevé*
Maladie bronzée de la tomate
Tomato Yellow Leaf Cur/ Virus (TYLCV)/ Ty-1+0TL 6 L. chilense; Partiel Virus de l'enroulement foliaire Polygénique + autres plusieurs autres sp.
Champignons
Cladosporium fulvum Cf-4
*
1 L. hirsutum Très élevé*
Cf-9, Cf-2 1, 6 L. pimpinellifolium
*
Cf-5 6 L. pimpinellifolium
*
+ autres gènes
Verticillium daliae Ve 9 L. pimpinellifolium Très élevé
*
Stemphylium spp.
Sm
11 L. pimpinellifolium Très élevéPyrenochaeta lycopersici/ py-1 3 L. peruvianum Partiel
Maladie des racines liégeuses
Leveillula taurica/Oïdium interne Lv 12 L. chilense Élevé
*
Oidium lycopersicum/Oïdium externe 0/-1 + QTL 6 L. hirsutum Partiel'
Polygénique + autres
Phytophthora infestans/Mildiou Ph-1, Ph-2, Ph-3 7, 9, 10 L. pimpinellifolium Partiel + autres
Fusarium oxysporum f. sp. /ycopersici/ /, 1-2
*
11 L. pimpinellifolium Très élevé*
Fusariose vasculaire 1-1 7 L. pennel/ii
Fusarium oxysporum f. sp. radicis lycopersici/ Fr/ 9 L. peruvianum Très élevé Fusariose du collet et des racines
Botrytis cinerea Polygénique L. hirsutum Élevé
Bactéries
Pseudomonas tomato Pto
*
5
L. pimpinellifolium Très élevéC/avibacter michiganensis/ polygénique Localisation Plusieurs espèces Partiel
Chancre bactérien de plusieurs OTL
Xanthomonas campestris pv. vesicatoria/ Bs3-2 L. escu/entum Élevé
Gale bactérienne + autres gènes L. pimpinellifolium
polygénique
Ralstonia solanacearum/ Polygénique Localisation L. pimpinellifolium Partiel
Flétrissement bactérien de plusieurs QTL et L. esculentum
dont chrom. 6 var cerasiforme
Parasites et ravageurs
Nématode Meloidogyne spp. Mi, Mi-3
*
6, 12 L. peruvianum Très élevé*
+ autres gènes
Nématode Globodera rostochiensis Hero 4 L. pimpinellifolium Très élevé
Puceron Macrosiphum euphorbiae Meu-1
*
6 L. peruvianum ÉlevéMouche Liriomyza spp. Polygénique L. hirsutum Partiel
L. cheesmanii
Mouche Trialeurodes vaporariorum Polygénique Localisation L. hirsutum Partiel Insectes en général de plusieurs QTL f. glabratum
Par convention, les gènes récessifs sont notés en minuscules, contrairement aux gènes dominants qui commencent par une majuscule.
Characteristics of resistance
to
the main tomato pathogens
Tableau 3
Les principaux gènes majeurs utilisés pour l'amélioration de la qualité du fruit et du mode de croissance
Gène Phénotype
ale (alcobaca) Maturation inhibée (semi-dominant)
at (apricot) Fruits oranges
B
(Beta) Fruits jaunesDel (Delta) Fruits oranges
hp (high pigment) Teneur en lycopène accrue
j (jointless) Absence de zone d'abcission du pédoncule
j-2 (jointless) Absence de zone d'abcission du pédoncule
nor (non ripening) Maturation inhibée (semi-dominant)
ag
e
(old gold-crimson) Teneur en lycopène accruer (yellow flesh) Fruits jaunes
rin (ripening inhibitor) Maturation inhibée (semi-dominant)
sp (self pruning) Croissance déterminée
t (tangerine) Fruits oranges
u (uniform ripening) Absence de collet vert
V
Épiderme incolore OrigineL.
esculentumL.
escu/entumL.
hirsutumL.
hirsutumL.
esculentumL.
esculentumL.
cheesmaniiL.
esculentumL.
esculentumL.
escu/entumL.
esculentumL.
esculentumL.
esculentumL.
esculentumL.
esculentum Cloné*
*
*
*
*
*
*
*
Chromosome10
56
12 1211
1110
6
35
6
10
10
Par convention, les gènes récessifs sont notés en minuscules, contrairement aux gènes dominants qui commencent par une majuscule.
Major genes used in breeding to enhance fruit quality and plant architecture in tomato
Ac
tuellement, l
a
sél
ect
i
o
n p
o
ur l
a
r
és
i
s
t
an-ce aux
m
ala
di
es e
t p
a
r
as
it
es a
nim
a
u
x
d
é
b
o
u
c
h
e s
ur d
es
h
y
br
i
d
es
F
1po
rtan
t d
es
gè
n
es
dominant
s
p
e
rm
e
ttant
d
e c
ontr
ô
l
e
r
d
e se
pt
à
neuf p
a
th
ogè
n
es (T
MV
, TSWV,
Cladosporium fù
l
vum, Vertici
LL
ium daliae,
Stemphy
l
ium
s
pp
.,
Fusarium o.
f
sp.
Lyco-persici
e
t
Fusarium o.
f
sp.
radicis-Lycoper-sici, Pseudomonas tomato
e
t n
é
m
a
tod
es à
g
all
es).
D
es
lign
ées
qu
as
i
i
s
og
é
niqu
e
s dan
s
l
e ry
p
e
Mon
ey
m
a
k
e
r
s
ont di
s
ponibl
es
po
u
r
l
es gè
n
es
Ve,
L
I
-2
,
Fr/,
py-
1
,
Mi,
Sm
,
P
to, Tm
-1
, Tm-2, Tm-:22
,
e
t
Sw
-
5
[
5
]
.
Le
ni
vea
u d
e ces
r
és
i
s
t
ances est gé
n
é-ral
e
m
e
nt tr
ès é
l
evé,
l
a
plup
art
d
e ces gè
n
es
é
tant impliqu
és
dan
s
d
es
rel
a
ti
o
n
s avec
l
e
p
a
tho
gè
n
e
d
e type gè
n
e
à
gè
n
e (
d
éte
rmi-n
ées
p
ar
l'
int
e
r
ac
ti
o
n
e
ntr
e
un
gè
n
e
d
e
r
és
i
s
t
ance c
h
ez
l
a
pl
ante e
t un
gè
n
e
d'avi
-r
ul
ence c
h
ez
l
e
p
a
th
ogè
n
e).
Qu
e
l
e
ur u
ti-li
sa
ti
o
n
so
ie
in
te
n
s
i
ve o
u non
,
plu
s
i
e
ur
s
d
e
ces
g
è
n
es s
ont
co
n
to
urn
és
p
a
r d
es races
a
d
a
pt
ées
du p
a
th
ogè
n
e. C'es
t n
o
t
am
m
e
nt
l
e cas
d
es gè
n
es
Mi,
L
Ve, Sw
-
5 (photo 2)
et
d
e ce
rt
ains gè
n
es
d
e
l
a sé
ri
e
Cf
D
e ce
fa
i
r,
l
es st
r
a
t
ég
i
es
d
e sé
l
ec
ti
o
n
évo
lu
e
nt
act
u
e
ll
e
m
e
nt
ve
r
s
l
e
cumul d
e
plu
s
i
e
u
rs
gè
n
es c
oncr
ô
lanc
d
es
m
éc
ani
s
m
es
d
e
r
és
i
s-tance
di
s
tinct
s
p
o
ur lut
ter co
ntr
e
un
se
ul
Photo 2. Symptômes de Tomato Spotted
Wilt Virus sur fruits (photo A. Moretti).
Photo 2. Tomato Spotted Witt Virus symptoms on fruit.
p
athogène
. P
ar exe
mpl
e,
l
e
ch
a
mpi
g
n
o
n
Cladosporium fo.
L
vum
es
t
ca
p
a
bl
e
d
e s
ur
-mont
e
r t
rès ra
pid
e
m
e
nt l
es
nou
vea
ux
gè
n
es
d
e
r
és
i
s
tan
ce
qui
lui
so
nt
o
pp
osés
(
huit
races o
nt
é
t
é
id
ent
ifi
ées e
n
France).
L
es
p
rogra
mm
es de
sél
ec
ti
o
n
ac
tu
e
l
s
v
i
se
nt d
o
n
c
à assoc
i
er
d
eux gè
nes,
p
a
r
exe
mpl
e
Cf6,
n
o
n
e
n
core s
urm
o
nté,
e
t
Cf 9,
qui n'est
s
urm
o
nt
é
qu
e
d
a
n
s
d
e
r
ares zo
n
es
d
e c
ultur
e
.
Cahiers Agricultures 2000
;
9
:
197-210
Pour améliorer la qualité
des fruits
C
ont
ra
ir
e
m
e
nt
a
ux
gè
n
es
d
e
r
es
1
s
c
a
.n
ce
qui pro
v
i
e
nn
e
nt tou
s
d'es
p
èces sa
u
vages,
d
e
n
o
mbr
e
u
ses
mut
a
ti
o
n
s a
pp
ar
u
es c
h
ez
l
a
tom
at
e c
ulti
vée s
ont utili
sées
p
o
ur
l
'
a
m
é
li
ora
ti
o
n d
es ca
r
ac
t
é
ri
s
t
i
qu
es
d
es
fruit
s
(tab
l
eau 3).
Couleur du fruit
L
a co
ul
e
ur du
fr
ui
t
d
é
pen
d
d
e
l
a
t
e
n
e
ur
tot
ale e
n p
ig
m
e
nt
s,
n
o
t
a
mm
e
nt
e
n
caro-t
è
n
e et
lyco
p
è
n
e.
Plu
s
i
e
ur
s
mut
a
ti
o
n
s
mo.n
ogé
niqu
es ag
i
sse
nt
s
ur
l
e
ur ten
e
ur
et
pa
r co
n
sé
qu
e
nt
s
u
r
l
a c
oul
e
ur du
fr
uit [6].
Ce
rt
a
in
es
bl
o
qu
e
nt l
a sy
nth
èse
du
l
yco
p
è
n
e :
l
es
fruit
s
p
ortan
t l
es gè
n
es
r, t, at
o
u
B
s
ont
ja
unes
o
u
oran
g
és. A
u
co
nt
rai
r
e,
l
es fr
ui
ts
p
ortant
l
es gè
n
es
ot
,
dg
o
u
hp
pr
ése
nt
e
nt un
ro
u
ge
plu
s
int
e
n
-se
.
Ce
rt
a
in
es
mut
a
tion
s
d
e
l
a co
ul
e
ur
du
fr
ui
t o
nt
ét
é
id
e
ntifi
ées c
h
ez
l
es es
p
èces
sa
uv
ages,
notamm
e
nt
c
h
ez
L.
hirsutum
.
L'ab
se
n
ce
d
e co
ll
e
t
(
p
ar
tie
du
fr
uit
sit
uée
a
utour d
e
l'
a
tt
ac
h
e
p
é
d
o
n
c
ul
a
ire)
vert
fon
cé ava
nt m
a
tu
r
it
é est
due
a
u
gè
n
e
r
écess
i
f u,
déso
rm
a
i
s
l
a
r
ge
m
e
nt u
t
ili
sé
dan
s
l
es var
iét
és
de
m
a
r
c
hé d
e fra
i
s
[l]
.
Teneur en sucre des fruits
D
es va
ri
a
nc
s
int
é
r
essa
nt
s
d
e gè
n
es
de
s
tru
c
ture du m
é
t
a
b
o
li
s
m
e ca
rbon
é
ont
é
t
é i
dencifi
és, c
h
ez
L.
ehmie
l
ewskii
p
o
ur
l
'
ac
ti
v
it
é
in
ve
rt
ase
[
7
]
e
t
c
h
ez
L.
hirsu
-tum
pou
r
l'A
G
P
ase
[
8],
p
e
rm
e
tt
a
nt
d
'o
bt
e
n
ir
de
s co
mp
osit
i
o
n
s o
ri
g
in
ales e
n
s
u
c
r
es.
L
e gè
n
e suer
d
'
in
ve
rt
ase
,
mut
é
c
h
ez
L.
ehmielewskii
,
co
ndui
t à des
fruit
s
acc
umul
a
nt du
sacc
h
arose et
n
o
n du
fru
crose et d
u
g
l
u
cose. C
h
ez
L.
hirsutum,
un
al
l
è
l
e
d
e
l
'
AGPase, bea
u-co
up plu
s e
ffi
cace
qu
e
l
'
al
l
è
l
e
d
e
l
'
es
p
èce
culti
vée, a é
t
é
d
é
t
ec
t
é et co
nduit
à
un
e
a
u
g
m
e
nt
a
tion d
e
l
a
t
e
n
e
ur fin
a
l
e e
n
s
u
c
r
e
du fruit.
Mutants de maturation :
rin, nor, ale
Plu
s
i
e
ur
s gè
n
es ayan
t u
ne act
i
on s
ur l
a
m
a
tur
a
tion du
fr
uit
so
nt
co
nnu
s. Le
plu
s
utili
sé e
n
sé
l
ec
ti
o
n
est
l
e gè
n
e rin
(ripe-ning inhibitor).
D
écri
t d
e
pui
s
1
968
,
il
p
rov
i
e
nt d
'
un
e m
u
tat
i
o
n
s
p
o
n
tanée
a
pp
a
ru
e
d
ans
du m
a
t
ér
i
el e
n
sé
l
ec
ti
on
d
e
L.
eseu
l
entum.
D
e
u
x a
u
t
r
es
mu
ta
ti
o
n
s
d
e
m
a
tur
a
tion int
éresse
nt l
es sé
l
ec
ti
o
n-n
e
ur
s
:
nor (non ripening)
,
qui pro
v
i
e
nt
d
'
un
e
v
a
ri
é
t
é ca
n
a
di
e
nn
e
d
éco
u
ve
r
te c
h
ez
un immi
g
rant it
al
i
e
n qui
l
a
d
es
tin
a
it
à
l
a
co
n
so
mm
a
ti
o
n
e
n hi
ver, e
t
ale (alcobaea),
qui
a é
t
é o
b
servée
p
o
ur l
a
pr
em
i
è
r
e fo
i
s
a
u P
o
rtu
g
al
. C
h
ez
l
es
pl
anres
pr
ése
nr
an
t
l
es m
u r
at
i
ons rin, nor o
u
a
l
e
à
l
'
é
t
a
t
h
omozygo
t
e, to
u
s
l
es process
u
s
d
e
l
a
matu
r
a
t
io
n
so
nt bl
oq
u
és e
t l
es fr
ui r
s
r
es-tent ja
un
es à ora
n
ges cla
ir
s (photo 3). Les
fr
uit
s
p
e
u
ve
nt
se c
on
se
r
ve
r
à
r
e
m p
é
r
a
tur
e
a
mbi
a
nt
e
p
l
u
s
i
e
ur
s
m
o
i
s sa
n
s é
v
o
l
u
e
r.
E
n
r
eva
n
c
h
e, à
l
'
é
t
a
t h
é
t
é
ro
zygo
t
e,
l
es
fruit
s
r
o
u
g
i
sse
nt
e
t mCtri
sse
nt
,
ma
i
s
plu
s
l
e
nt
e
-me
nt
, e
t d
o
n
c se co
n
serve
nt mi
eux
qu
e
l
es fr
uit
s
n
e
p
o
rt
an
t p
as
l
a
mut
a
ti
o
n
[9].
Architecture
Photo 3. Fruits de
lignées isogéniques sauf pour les gènes de maturation. De gauche à droite :
lignée normale, mutations ale, nor, rin (photo J. Phi -louze). Photo 3. Fruits of isogenic lines that are closely-related, except in terms of maturation genes. From left ta right, normal fruit, ale, nor and rin mutations.

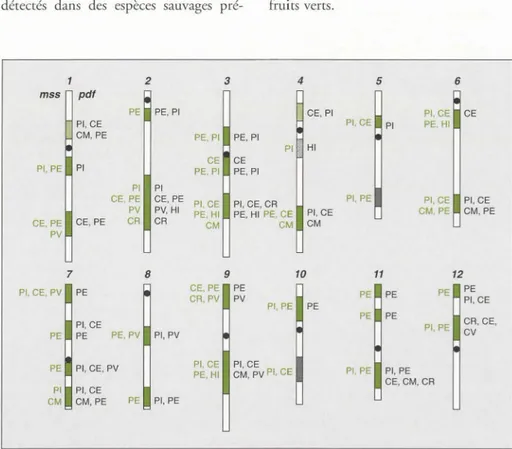
![Figure 2 . Lo ca lisation chromosomique des gènes majeurs (en noir) e t QTL (en vert) de résis- résis-tance aux maladies c he z la tomate [ 18]](https://thumb-eu.123doks.com/thumbv2/123doknet/2313309.27321/7.871.41.830.54.477/figure-lisation-chromosomique-genes-majeurs-resis-resis-maladies.webp)