T
T
H
H
È
È
S
S
E
E
En vue de l'obtention du
D
D
O
O
C
C
T
T
O
O
R
R
A
A
T
T
D
D
E
E
L
L
’
’
U
U
N
N
I
I
V
V
E
E
R
R
S
S
I
I
T
T
É
É
D
D
E
E
T
T
O
O
U
U
L
L
O
O
U
U
S
S
E
E
Délivré par l'Université Toulouse III - Paul SabatierDiscipline ou spécialité : Microbiologie
JURY
Dr. Stéphane BERTAGNOLI, Maître de Conférence à l'ENVT ... Directeur de thèse Dr. Christelle CAMUS-BOUCLAINVILLE, Maître de Conférence à l'ENVT ...Co-directeur de thèse Dr. Robert DRILLIEN, Directeur de Recherche à l'INSERM ... Rapporteur Dr. Christophe PASQUIER, Professeur à l'UPS ... Examinateur Dr. Ginette DAMBRINE, Directeur de Recherche à l'INRA ... Examinateur Dr. Philippe ERBS, Chercheur à Transgène ... Examinateur
Ecole doctorale : Biologie-Santé-Biotechnologies Unité de recherche : UMR 1225 INRA/ENVT Directeur(s) de Thèse : Dr. Stéphane BERTAGNOLI
Dr. Christelle CAMUS-BOUCLAINVILLE Rapporteurs : Dr. Max BERGOIN, Professeur
Dr. Robert DRILLIEN, Directeur de Recherche
Présentée et soutenue par Sophie BLANIE
Le 9 février 2009
Titre : Les protéines à motif Ankyrine du virus myxomateux : Caractérisation et implication dans le pouvoir pathogène.
Messieurs les Professeurs Robert Drillien et Max Bergoin pour avoir accepté de juger et de commenter mon travail ;
Les membres du jury qui ont consacré de leur temps à lire et écouter mon travail ;
Monsieur le Docteur Stéphane Bertagnoli et Madame le Docteur Christelle Camus-Bouclainville pour les nombreuses discussions qui ont permis de mener à bien ce projet ;
Jacqueline Gelfi pour son aide précieuse ;
Tous les « locataires » du laboratoire, qu’ils soient permanents ou seulement de passage.
Cette thèse a été financée par le Ministère de l’Enseignement supérieur et de la Recherche.
Un très grand merci
A Nicoco pour m’avoir fait découvrir le Myxo ;
Aux résidents du « poulailler », en particulier Béa et Magalou pour leur soutien et Jérémy pour avoir été un stagiaire plein d’entrain et de bonne humeur.
A toute ma famille,
A mes parents et à ma sœur Sandrine,
A Gérald,
AVANT-PROPOS
Le virus de la myxomatose (MYXV) circule certainement depuis très longtemps sur le continent américain, à l’état endémique, dans les populations de lapins du genre Sylvilagus qui sont des porteurs sains. C’est suite à l'introduction du lapin Européen (Oryctolagus cuniculus) en Amérique du sud que le pouvoir pathogène du MYXV est révélé. Il est alors identifié par Sanarelli à la fin du XIXe siècle.
Dès lors, ce virus est le sujet d’une histoire riche, faisant l'objet de considérations et d'utilisations aussi variées que contraires : tour à tour arme biologique et outil de vaccination, voici un résumé de son histoire.
Dans les années 50, sa première utilisation fut en tant qu'arme biologique en Australie pour lutter contre le lapin Européen. Ce dernier, importé par les colons, et en l’absence de prédateurs, s’est reproduit de manière dramatique. Il ravage les cultures et devient rapidement une menace pour la faune et la flore indigène. Pour lutter contre ce fléau, les autorités décident d’utiliser le virus de la myxomatose. C’est à ce jour le premier et seul exemple d'introduction volontaire d'un agent viral dans la nature, dans le but d'éradiquer une espèce animale nuisible.
L'introduction illégale du virus de la myxomatose en France découle quant à elle de l’initiative privée d’un médecin voulant éradiquer les lapins de son jardin. Il en résulte une implantation fulgurante et durable du MYXV dans toute l’Europe.
Suite à cela, et en réponse à la demande des éleveurs et chasseurs, pour qui la myxomatose représente un fléau, les études sur ce virus ont été orientées vers la recherche d'un vaccin. Des souches vaccinales homologues ont été ainsi développées, permettant une vaccination contre la myxomatose. Des souches recombinées ont aussi été utilisées quelques années plus tard comme vecteur vaccinal contre la maladie hémorragique virale du lapin.
Des études ont ensuite montré que le virus de la myxomatose pouvait être utilisé comme vecteur vaccinal chez des espèces non-hôte, comme c'est le cas pour la calicivirose féline (McCabe et al., 2002). De nouveaux travaux sont en cours sur l'utilisation du virus myxomateux en tant que vecteur pour la vaccination des ruminants contre la fièvre catarrhale ovine.
Plus récemment, le virus de la myxomatose s’est révélé être un bon candidat pour la virothérapie anti-cancéreuse chez l’homme (Lun et al., 2005).
Afin d’optimiser le virus de la myxomatose, notamment en tant que vecteur vaccinal et virus oncolytique, il est nécessaire de mieux connaître ses facteurs de pathogénicité et leur implication dans les grandes voies de signalisation.
Notons aussi que l’étude de gènes viraux (notamment MV-LAP et les scrapines) permet parfois d’associer une fonction à des gènes humains, et eucaryotes en général, et de découvrir des mécanismes cellulaires physiologiques.
Mon projet s’intéresse à trois gènes du virus myxomateux: M148R, M149R et M150R, qui codent des protéines à répétitions ankyrine.
La partie bibliographique de ce document présente les caractéristiques générales communes aux poxvirus ainsi que les différents mécanismes mis en œuvre par ces virus pour échapper à la réponse immunitaire de l’hôte infecté, pour finalement se focaliser sur les facteurs de pathogénicité présentant une structure commune, les répétitions ankyrine, sujet de notre étude.
La partie expérimentale présente l’ensemble de nos travaux de recherche réalisés au laboratoire. La première partie s’intéresse aux études concernant la caractérisation de M148R et M149R, tout en faisant le parallèle avec les données connues sur M150R (MNF). La deuxième partie présente la continuation des études menées sur MNF.
Table des Matières
TABLE DES ABREVIATIONS ... TABLE DES ILLUSTRATIONS ...
ÉTUDE BIBLIOGRAPHIQUE ... 1
I. CARACTERES GENERAUX DES POXVIRUS ET DU VIRUS MYXOMATEUX... 1
A. Classification... 1
B. Structure des poxvirus ... 4
1. Structure des virions ... 4
2. Organisation du génome... 8
C. Cycle viral ... 11
1. Entrée du virion dans la cellule ... 11
2. Expression et régulation des gènes viraux... 14
3. Expression des gènes précoces... 18
4. Réplication du génome viral... 18
5. Expression des gènes intermédiaires et tardifs ... 19
6. Assemblage et maturation des virions ... 20
II. LE VIRUS DE LA MYXOMATOSE... 20
A. Pathogénie et réponse immunitaire... 20
1. Pathogénie ... 20
2. Réponse immunitaire... 24
B. Facteurs de pathogénicité ... 25
1. Facteurs anti-apoptotiques... 25
2. Inhibition de l’Interféron ... 31
3. Inhibition de l’activation des leucocytes ... 34
4. Autres facteurs... 36
III. LES PROTEINES A MOTIF ANKYRINE CHEZ LES POXVIRUS... 40
A. Structure ... 40
1. Originalité des protéines poxvirales à motif ANK ... 40
2. Le motif ankyrine ... 43
3. Le motif F-box et la poly-ubiquitination ... 45
ETUDE EXPERIMENTALE ... 56
I. M148R ET M149R ... 56
A. Introduction ... 56
B. Analyses de séquences de M148R et M149R... 59
C. Expression au cours du cycle viral... 61
D. Localisation cellulaire... 63
E. M148R et M149R n’affectent pas la réplication virale dans les cellules permissives ... 65
F. M148R et M149R sont des facteurs de pathogénicité chez le lapin Européen67 1. Essais cliniques... 67
2. Analyses histopathologiques ... 67
G. M148R et M149R interagissent avec la culline 1 (CUL1) ... 70
H. Discussion... 72
I. Article ... 74
II. MNF ... 75
A. Introduction ... 75
B. Construction et validation du virus recombiné ... 76
C. Recherche des partenaires de MNF ... 80
D. Localisation de MNF au cours du cycle viral ... 86
E. Discussion... 94
DISCUSSION GENERALE ... 97
MATERIEL ET METHODES ... 105
Table des Abréviations
Ac Anticorps
ADN Acide DésoxyriboNucléique
ADNc ADN complémentaire
Ag Antigène
ANK Ankyrine
ARN Acide ribonucléique
ARNm ARN messager
ATP Adénosine triphosphate
BGMK Baby Green Monkey Kidney
BRdU Bromodeoxyuridine
CPA Cellule Présentatrice d’Antigène
CRL Cullin-RING ubiquitin Ligases
DMEM Dulbecco’s Minimum Essential Medium
dNTP désoxynucléoside triphosphate
ECL ElectroChimioLuminescence
EDTA Ethylenediaminetetraacetic acid
EEA1 Early Endosome Antigen1
EEV Extracellula Enveloped Virion, forme extracellulaire enveloppée
GFP Green Fluorescent Protein
IκB Inhibiteur du facteur NFκB
IL Interleukine
IEV Virion Intracellulaire Enveloppé
IFN Interféron
IMV Virion Intracellulaire Mature
IP Immunoprécipitation
MNF Myxoma Nuclear Factor
m.o.i Multiplicity Of Infection, multiplicité d’infection
NFκB Nuclear Factor Kappa B
NK Natural Killer
OAS OligoAdenylate Synthetase
OptiMEM Optimised Minimum Essential Medium
ORF Open Reading Frame, cadre de lecture ouvert
PAGE Polyacrylamide gel electrophoresis
PBS Phosphate-Buffered Saline
PCR Polymerase Chain Réaction, réaction de polymérisation en chaîne
PFA Paraformaldéhyde
PFU Unité Formant Plage
p.i. post-infection
PS Pénicilline /Streptomycine
Rbx RING box protein
RL5 Lignée cellulaire de lymphocytes de lapin
RS Récepteur Substrat
RT Reverse Transcription
RK13 Rabbit Kidney 13
SCF Skp1-Cullin-F-box
SDS Sodium Dodecyl Sulfate
Skp1 S-phase Kinase-associated Protein 1
SMART Simple Modular Architecture Research Tool
SVF Sérum de Veau Fœtal
TNFα Tumor Necrosis Factor α
TNFR Récepteur du TNF
Table des Illustrations
Liste des Figures
Figure 1 : Représentation schématique d’un orthopoxvirus EEV ou CEV ...5
Figure 2 : Observation du MYXV en microscopie électronique à transmission...6
Figure 3 : Représentation schématique du génome viral de Poxviridae ...9
Figure 4 : Représentation schématique de l’organisation génomique du MYXV...10
Figure 5 : Cycle de réplication des Poxviridae ...12
Figure 6 : Cinétique d’expression des gènes, fonctions et localisations associées ...17
Figure 7 : Forme classique de myxomatose induite par la souche T1...23
Figure 8 : Mécanismes d’échappement du MYXV à l’apoptose ...30
Figure 9 : Répartition des protéines à motif ANK ...41
Figure 10 : Structure de protéines à répétitions ANK en haute résolution...44
Figure 11 : Les différentes valences d’ubquitination ...47
Figure 12 : Organisation fonctionnelle d’un complexe CRL ...49
Figure 13 : Diversité des substrats ubiquitinés par le SCF...52
Figure 14 : Modèle du mécanisme d’action des protéines à motif ANK poxvirales ...55
Figure 15 : Localisation de M148R, M149R et MNF sur le génome du MYXV ...57
Figure 16 : Protéines à motif ANK du MYXV ...58
Figure 17 : Analyse des motifs protéiques de M148R et M149R. ...60
Figure 18 : Cinétique d’expression de M148R, M149R et MNF...62
Figure 19 : Localisation cellulaire de M148R et M149R...64
Figure 20 : Courbes de croissance des virus ...66
Figure 21 : Observation histologique du myxome primaire...69
Figure 22 : M148R et M149R interagissent avec CUL1...71
Figure 23 : Carte du virus recombiné T1-GFPMNF ...78
Figure 24 : Expression de la fusion GFP-MNF...79
Figure 25 : Résultat des CoIP sur gel SDS PAGE ...81
Figure 26 : Résultat des CoIP sur extraits solubles sur gel SDS PAGE ...83
Figure 28 : Localisation de MNF 1 heure p.i. ...88
Figure 29 : Localisation de MNF 2 heures p.i...89
Figure 30 : Localisation de MNF 3 heures p.i...90
Figure 31 : Localisation de MNF 5 heures p.i...92
Figure 32 : Localisation de MNF 14 heures p.i...93
Figure 33 : Carte des virus recombinés T1∆M148R, T1∆M149R et T1∆M148R∆M149R. ...107
Liste des Tableaux
Tableau I : Classification des Chordopoxvirinae...2Tableau II : Les membres du genre Leporipoxvirus et leurs pathologies associées ...3
Tableau III : Séquences consensus des différents promoteurs es gènes de la vaccine..15
Tableau IV : Facteurs de pathogénicité avérés du MYXV ...38
Tableau V : Facteurs de pathogénicité supposés du MYXV ...39
Tableau VI : Incidence du motif F-box au sein des protéines poxvirales à motif ANK 42 Tableau VII : Protéines et motifs majoritaires entrant dans la composition des CRL ..51
Étude bibliographique
I.
Caractères généraux des poxvirus et du virus
myxomateux
Le virus de la myxomatose appartient à la famille des Poxviridae. Il possède de ce fait les caractéristiques communes aux membres de cette famille originale de virus à ADN.
A.
Classification
C’est en 1957 que Fenner et Burnet établirent les bases de la classification des poxvirus (Fenner & Burnet, 1957). La famille des Poxviridae est divisée en deux sous-familles dont les membres infectent les vertébrés pour l'une (Chordopoxvirinae) et les invertébrés pour l'autre (Entomopoxvirinae).
La sous-famille des Chordopoxvirinae est elle-même subdivisée en 8 genres (tableau I) définis sur la base de protections croisées chez l’animal, de neutralisations croisées sur cultures cellulaires et d’hybridations croisées des génomes (Van Regenmortel, 2000).
Le virus prototype des Poxviridae est le virus de la vaccine (VACV), chef de file du genre Orthopoxvirus. Historiquement, le VACV est à l’origine du terme de vaccination. En effet, il a longtemps été utilisé comme vaccin contre la variole (maladie humaine causée par un autre Orthopoxvirus), jusqu'à son éradication en 1980 (WHO, 1980).
Le virus de la myxomatose (MYXV) appartient quant à lui au genre Leporipoxvirus. Les membres du genre Leporipoxvirus et leurs hôtes respectifs sont décrits dans le tableau II.
Genre
Espèce type
Principales autres espèces
Orthopoxvirus Virus de la vaccine
Mousepox (ectromélie) Cowpox virus
Virus de la variole du chameau Virus de la variole du singe Virus de la variole humaine
Raccoonpox virus Taterapox virus
Volepox virus
Parapoxvirus Orf virus
Virus de la stomatite papuleuse des bovins Pseudocowpox virus
Squirrel parapoxvirus
Avipoxvirus Virus de la variole aviaire
Virus de la variole du canari Virus de la variole du dindon Virus de la variole du pigeon + virus spécifiques de plusieurs espèces
d’oiseaux
Capripoxvirus Virus de la clavelée Virus de la variole caprine Lumpy skin disease virus
Leporipoxvirus Virus de la myxomatose
Virus du fibrome de Shope Virus du fibrome malin du lapin
Virus du fibrome du lièvre Virus du fibrome de l’écureuil
Suipoxvirus Virus de la variole porcine
Molluscipoxvirus Virus du Molluscum contagiosum
Yatapoxvirus Tanapox virus de
l’homme Virus Yaba de la tumeur du singe ND Crocodilepox virus
Tableau I : Classification des Chordopoxvirinae. D’après Van Regenmortel et al.,
Virus
Pathologie chez
l’hôte naturel
Pathologie chez Oryctolagus
cuniculus
Virus de lamyxomatose
Sylvilagus sp : myxome bénin localisé
Myxomatose généralisée et immunodépression
Virus du fibrome de Shope
Sylvilagus floridanus : fibrome bénin localisé
Fibrome bénin localisé, généralisation chez les nouveaux-nés
Virus du fibrome malin du lapin ND Nodules dermiques, immunodépression généralisée Virus du fibrome de l’écureuil Sciurus sp : fibromes
multiples Non sensible
Virus du fibrome du lièvre
Lepus sp : fibromes
cutanés localisés Fibromes bénins localisés
Tableau II : Les membres du genre Leporipoxvirus et leurs pathologies associées.
B.
Structure des poxvirus
1. Structure des virions
Les Poxviridae font partie des plus gros et des plus complexes virus animaux connus. Ils ont une forme ovoïde de 200 à 400 nm de long (Murphy, 1999). Les études de coupes fines de virions en microscopie électronique ont permis de définir quatre structures distinctes : le core, les corps latéraux, la membrane, et un nombre variable d’enveloppes (Figure 1).
Le core est une coque interne épaisse abritant le génome viral, constitué d'une molécule unique d'ADN linéaire double brin, et des protéines virales, nécessaires aux premières étapes du cycle de réplication.
Les corps latéraux sont des structures lenticulaires situées dans les concavités du core. Pour certains auteurs, il ne s’agirait que d’artéfacts de microscopie électronique (Dubochet et al., 1994), bien que d’autres réfutent cette théorie (Griffiths et al., 2001b). Des analyses en cryomicroscopie ont montré qu’ils avaient une réalité physique (Cyrklaff et al., 2005).
La membrane est une bicouche lipoprotéique classique de 5 nm d’épaisseur entourant le core. Sa composition est différente de celle de la cellule hôte. Certains auteurs prônent qu’elle serait synthétisée de novo (Dales & Siminovitch, 1961). Cette hypothèse est controversée par des auteurs montrant une fusion entre des membranes cellulaires et la membrane des IMV (Husain et al., 2006; Sodeik et al., 1993).
Figure 1 : Représentation schématique d’un orthopoxvirus EEV ou CEV. D’après
Van Regenmortel et al., 2000. ND = non déterminé.
IMV IMVIMV IMV (Surface) (Surface) (Surface) (Surface) EEV EEV EEV EEV ((((CoupeCoupeCoupeCoupe))))
Figure 2 : Observation du MYXV en microscopie électronique à transmission. (J.L.
Duteyrat, communication personnelle) A : MYXV intracellulaire en microscopie. B : MYXV purifié sur gradient de saccharose et coloré à l’acide phosphotungstique.
100 nm
A
B
Le nombre et la nature des enveloppes varient au cours du cycle viral et permettent ainsi de distinguer quatre formes de virions : les IMV (Virions Intracellulaires Matures), les IEV (Virions Intracellulaires Enveloppés), les EEV (Virions Extracellulaires Enveloppés) et les CEV (Virions Enveloppés associés à la Cellule).
Les IMV sont formés dans le cytoplasme. Certains auteurs suggèrent qu’ils présenteraient deux membranes formées de deux bicouches lipidiques accolées (Griffiths et al., 2001a; Griffiths et al., 2001b; Sodeik et al., 1993). Cependant, on leur attribue classiquement une seule membrane et pas d’enveloppe, ce qui a été confirmé par des observations en microscopie électronique (Carter et al., 2005; Hollinshead et al., 1999). Les IMV représentent la forme la plus abondante de la progénie virale et sont libérés lors de la lyse de la cellule hôte. Cette forme est la plus résistante et serait adaptée au passage du virus d’un hôte à l’autre.
Une petite partie des IMV acquiert une double enveloppe supplémentaire dérivée de l’appareil de Golgi (Hiller & Weber, 1985; Schmelz et al., 1994) ou des endosomes (Tooze et al., 1993) et modifiée par l’insertion d’au moins 7 protéines virales (Condit et al., 2006; Smith et al., 2002). Ils constituent les IEV. Ils migrent à la surface cellulaire grâce aux microtubules du cytosquelette et fusionnent avec la membrane plasmique de la cellule hôte (Hollinshead et al., 1999). Les IEV perdent ainsi leur enveloppe la plus externe et permettent la libération d’EEV par exocytose. Une partie des EEV est retenue à la surface cellulaire, formant les CEV.
EEV et CEV contiennent une membrane de plus que les IMV et une de moins que les IEV. Ils ne représentent qu’une faible proportion de la progénie virale (environ 1 %). Leur enveloppe supplémentaire leur confère des propriétés particulières en terme de reconnaissance des cellules cibles et de protection contre les effecteurs du système immunitaire. Si naturellement l’infection n’induit que peu d’anticorps neutralisants dirigés contre les EEV, une sérothérapie utilisant ces Ac permet de protéger l’animal (Law & Smith, 2001). De même, les EEV sont résistants au complément (Vanderplasschen et al., 1998). Les CEV assurent l’infection des cellules de proche en
proche alors que les EEV permettent une dissémination virale sur de plus longues distances.
2. Organisation du génome
Le génome des poxvirus est formé d’une molécule d’ADN bicaténaire, géneralement riche en A-T (60 à 70%) (sauf parapoxvirus), aux extrémités liées de façon covalente par des boucles terminales en épingle à cheveux (Figure 3) (Van Regenmortel, 2000). La taille de l'ADN varie de 130 à 375 kpb dans l’ensemble de la famille. La région centrale comporte des gènes hautement conservés, codant les enzymes et les protéines structurales essentielles au cycle viral. Cette région est bordée de séquences terminales inversées répétées (TIR) de 10 à 15 kpb. A la jonction de ces régions et au niveau des séquences terminales, soit environ 25 kpb, on trouve des gènes non essentiels. Ils codent principalement des facteurs de pathogénicité (Moss, 2001).
La rareté des séquences non codantes et l’absence d’introns permettent aux génomes poxviraux de coder 150 à 200 gènes.
Le MYXV quant à lui possède un génome de 162 kpb codant 171 gènes (Cameron et al., 1999).
Les gènes sont nommés en fonction de leur position sur le génome, par ordre croissant de gauche à droite et selon qu’ils sont transcrits respectivement vers la droite (suffixe R) ou bien vers la gauche (suffixe L) (Figure 4).
La plupart des génomes de poxvirus ont été séquencés. Les séquences complètes sont disponibles sur le site internet suivant : http://www.poxvirus.org.
Figure 3 : Représentation schématique du génome viral de Poxiviridae (modèle : VACV). TIR : séquences Terminales Inversées Répétitées (Fenner et al., 1989). La région centrale comporte des gènes hautement conservés nécessaires à la replication virale. Au niveau des TIR et dans les régions à proximité se trouvent essentiellement des gènes de pathogénicité.
TIR ≈≈≈≈15kpb TIR ≈≈≈≈15 kpb Séquences uniques ≈≈≈≈130 kpb
Figure 4 : Représentation schématique de l’organisation génomique du MYXV. Les groupes de gènes sont représentés par des rectangles. Ils sont dessinés sur ou sous la ligne bleue selon qu’ils sont transcrits respectivement vers la droite (suffixe R) ou vers la gauche (suffixe L). Séquences uniques (M009L, …M156R) TIR TIR TIR TIR gauche (M000.5L, …M008.1L) […] TIR droite (M008.1R, …M000.5R)
C.
Cycle viral
Le cycle réplicatif des poxvirus a été étudié en détail et à l’échelle moléculaire en prenant le VACV pour modèle. Ce cycle a la particularité de se dérouler intégralement dans le cytoplasme de la cellule hôte. Jusqu’à présent, aucun exemple d’intégration d’ADN poxviral dans le génome de la cellule hôte n’a été montré. De même, il n’existe pas de phase de latence au cours du cycle réplicatif poxviral. Le cycle réplicatif se déroule en 12 à 24 h pour le VACV (Moss, 2001) et environ 16 h pour le MYXV (Duteyrat et al., 2006). La figure 5 illustre les différentes étapes de ce cycle.
1. Entrée du virion dans la cellule
L’existence de plusieurs formes infectieuses complique fortement les études menées sur le mode d’entrée des virions dans la cellule hôte. Bien que plusieurs mécanismes de pénétration des poxvirus dans les cellules cibles aient été proposés, à ce jour, aucune hypothèse n’a été fermement démontrée. La question fait toujours débat et des publications récentes en proposent un état des lieux (Condit et al., 2006; Roberts & Smith, 2008).
Selon certains auteurs, les IMV perdraient leur enveloppe avant d’entrer dans la cellule, et seul le core traverserait ainsi la membrane plasmique (Sodeik & Krijnse-Locker, 2002).
Cependant, l’hypothèse la plus généralement acceptée est que la pénétration des IMV se fait par fusion membranaire. Des images de microscopie électronique démontrent d’ailleurs la continuité des membranes virale et cellulaire (Carter et al., 2005). Enfin, certains auteurs suggèrent que les IMV seraient endocytés, puis le core libéré par fusion membranaire dans le cytoplasme (Townsley et al., 2006). Ces deux derniers mécanismes seraient utilisés, la préférence pour une voie ou l’autre serait déterminée par la souche virale et le type cellulaire (Townsley et al., 2006).
Figure 5 : Cycle de réplication des Poxviridae – modèle VACV
1 : Attachement à la cellule. 2 : Pénétration dans la cellule et libération du core dans le cytoplasme. 3 : Transcription des gènes précoces. 4 : Déshabillage du core. 5 : Réplication de l’ADN et formation de concatémères. 6 : Transcription des gènes intermédiaires. 7 : Transcription des gènes tardifs. 8 : Début de l’assemblage avec les premiers fragments de membranes. 9 : Résolution des concatémères et incorporation de l’ADN. 10 : maturation et formation des IMV. 11 : Acquisition d’une double enveloppe (IEV). 12 : Migration vers la surface cellulaire viale cytosquelette. 13 : Fusion avec la membrane plasmique cellulaire, formation des CEV et libération des EEV.
Facteurs de transcription tardifs ARNm intermédiaires ARN polymérase Facteurs de transcription intermédiaires N Nooyyaauu C Cyyttooppllaassmmee M Meemmbbrraanneeppllaassmmiiqquuee ARNm précoces ARNm tardifs ADN polymérase ARN polymérase Facteurs de transcription intermédiaires Enzymes de coiffage
Polymérase PolyA Facteurs de croissance Facteurs de pathogénicité ADN Core Enveloppe Enzymes tardives Protéines structurales Facteurs de transcription précoces 1 2 3 4 5 6 7 8 9 10 12 13 11 E EEEVV IIMMVV I IMMVV I IEEVV E EEEVV
Les mécanismes d’entrée dans la cellule des EEV sont encore plus controversés. Selon certains auteurs, les EEV pénètreraient par endocytose et seraient pris en charge par des vésicules intracellulaires (Ichihashi, 1996). Le pH acide des vésicules induirait la rupture de l’enveloppe externe et le core serait libéré par fusion de la membrane des IMV avec celle des vésicules (Townsley et al., 2006). Récemment, un autre mécanisme a été proposé, sur les bases d’observations en microscopie électronique. La membrane des EEV se romprait au contact de la membrane cellulaire, et resterait à l’extérieur de la cellule (Law et al., 2006). La particule IMV ainsi libérée pénètrerait par fusion membranaire, comme décrit ci-dessus.
À ce jour, les récepteurs cellulaires empruntés par le VACV n’ont pas été identifiés. De même, il n’a pas été démontré que les poxvirus présentent un ligand majeur nécessaire à leur attachement à la cellule via un récepteur.
L’entrée des IMV nécessite un complexe de 9 protéines virales pour la fusion membranaire (A16, A21, A28, F9, G3, G9, H2, J5 et L5) (Moss, 2006; Senkevich et al., 2005). Cependant, ces mêmes protéines ne semblent pas être impliquées dans l’attachement à la cellule. Il est vraisemblable que des interactions peu spécifiques interviennent dans les contacts précoces entre les IMV et la cellule cible. Ainsi, les protéines H3, A27 et D8 du VACV se fixent aux glycosaminoglycanes présents à la surface des cellules (Chung et al., 1998; Hsiao et al., 1999; Lin et al., 2000) et A26 interagit avec la laminine, protéine de la matrice extracellulaire (Chiu et al., 2007). On dispose de peu de renseignements concernant les interactions des EEV avec la cellule. La rupture de l’enveloppe des EEV au contact de la cellule impliquerait des polyanions tels que les héparines cellulaires ainsi que les protéines virales B5 et A34 (Law et al., 2006).
Notons ici que plusieurs virus peuvent infecter une même cellule, chaque core internalisé donnant lieu à un site de réplication virale distinct (Katsafanas & Moss, 2007).
Concernant l’entrée du VACV dans la cellule, de récentes images en microscopie électronique semblent confirmer le mécanisme d’entrée des EEV par rupture préalable de l’enveloppe et entrée des IMV (Law et al., 2006). Des images d’endocytose et macropinocytose ont également été rapportées (Mercer & Helenius, 2008).
2. Expression et régulation des gènes viraux
Le fait que le cycle de réplication virale des Poxviridae se déroule entièrement dans le cytoplasme de la cellule infectée implique qu’ils soient relativement autonomes pour les processus de réplication et de transcription qui ont habituellement lieu dans le noyau de la cellule infectée. De ce fait, les poxvirus codent leurs propres enzymes et principaux facteurs de transcription et disposent d’une machinerie pour la réplication de leur génome.
On distingue trois types de promoteurs et quatre classes de gènes exprimés de manière séquentielle au cours du cycle viral : les gènes précoces, transcrits avant la réplication du génome viral, les gènes intermédiaires et gènes tardifs, transcrits après. Ces différentes classes sont définies par un promoteur distinct (Tableau III) et parfois un terminateur. Les gènes possédant des éléments de promoteur précoce et tardif forment la quatrième classe de gènes, nommés précoce/tardif (Broyles, 2003).
Récemment, une cinquième classe de gènes a été identifiée (Assarsson et al., 2008). Il s’agit des gènes précoces immédiats, exprimés dès les premières minutes de l’infection. Ils se caractérisent par un très fort taux d’expression, qui culmine à 2 heures post-infection avant de décliner. Ces mêmes auteurs remettent en cause l’existence de la classe des gènes intermédiaires, ces derniers étant répartis, selon leur analyse, au sein des classes précoces, précoces/tardifs et tardifs ou non classifiés, du fait d’un profil d’expression atypique.
Promoteurs
Séquences consensus
Précoces (A/T)8G(A/T)8N12(G/A)
Intermédiaires (A/T)8N12TAAA(T/A)GG
Tardifs (A/T)6N9TAAAT
Tableau III : Séquences consensus des différents promoteurs des gènes de la vaccine. D’après Broyles, 2003. Rouge souligné : site d’initiation de la transcription.
L’expression des gènes au cours du cycle viral est étroitement associée à leur fonction, les gènes précoces codant principalement les protéines nécessaires à la réplication virale et à l’échappement au système immunitaire tandis que les gènes tardifs codent les protéines structurales entrant dans la composition du core ou des membranes (Figure 6A et 6B). Enfin, il semble que les gènes des différentes classes soient groupés sur le génome (Figure 6C) (Assarsson et al., 2008).
La succession d’événements au cours du cycle réplicatif contrôle l’expression des gènes : la transcription des gènes intermédiaires nécessite l’intervention de protéines exprimées précocement, celle des gènes tardifs faisant appel aux protéines intermédiaires et celle des gènes précoces aux protéines tardives qui sont encapsidées et disponibles pour le cycle suivant.
Les mécanismes de transcription semblent conservés au sein de la famille. Une même ARN polymérase à 9 sous-unités est utilisée pour la transcription des différents types de gènes.
Figure 6 : Cinétique d’expression des gènes, fonctions et localisations associées (Assarsson et al., 2008). A : Fonctions associées aux différentes classes de gènes (jaune : répication, rouge : pathogénicité, vert : transcription, bleu : structure du virion). B : Classes de gènes associées aux différentes fonctions (jaune : précoce-immédiat, rouge : précoce, vert : précoce-tardif, bleu : tardif). C : Les gènes des différentes classes sont regroupés sur le génome, le brin du haut représente les gènes transcrit vers la droite et celui du bas, les gènes transcrits vers la gauche.
3. Expression des gènes précoces
Dès l’entrée du virus dans la cellule, et avant même sa décapsidation, les premiers ARNm sont transcrits. Ils sont synthétisés dans le core viral encore intègre, grâce à la présence de l’ARN polymérase, codée par le génome viral et encapsidée dans le virion. Ces ARNm viraux sont modifiés grâce à une enzyme de coiffage des ARNm, une poly(A) polymérase et une 2’-O-methyltransferase, elles aussi codées par le génome viral et encapsidées dans le virion. Les transcrits s’échappent ensuite par des pores à la surface du core, et sont traduits via les ribosomes cellulaires. La transcription des gènes précoces ne semble ainsi faire appel à aucune protéine cellulaire (Broyles, 2003).
Les promoteurs précoces ont une taille de 30 pb environ (voir tableau III). Ils possèdent un résidu guanine presque toujours placé à - 21 ou - 22 en amont du site d’initiation de la transcription, et flanqué par des régions variables riches en AT. Les séquences régulatrices sont assez conservées entre les genres. La transcription s’initie en général 15 à 30 bases en aval du promoteur.
Le signal de terminaison de la transcription des gènes précoces est situé juste en aval de l’ORF et est constitué de la séquence TTTTTNT.
Les protéines impliquées dans la réplication de l’ADN, la biosynthèse des nucléotides, la transcription des gènes intermédiaires mais aussi certains mécanismes d’échappement aux défenses de l’hôte sont exprimés précocement. La synthèse des protéines précoces semble nécessaire à la décapsidation et à la libération du génome viral.
4. Réplication du génome viral
Après désintégration du core, l’ADN viral devient totalement accessible aux enzymes. Sa réplication se déroule de façon autonome dans le cytoplasme de la cellule infectée, à l’aide d’enzymes d’origine essentiellement virale. L’ADN viral nu s’associe aux
membranes du réticulum endoplasmique (RE) qui l’enveloppent pour former un compartiment dans lequel la réplication s’initie (Mallardo et al., 2002).
Au début du cycle, la boucle de chaque télomère de l’ADN viral est coupée par une endonucléase spécifique. Les télomères poxviraux sont, par analogie aux chromosomes eucaryotes, des séquences d’ADN non codantes situées aux extrémités de la molécule linéaire d’ADN viral. C’est dans les télomères qu’ont lieu l’initiation et la terminaison de la réplication (DeLange & McFadden, 1987; DeLange & McFadden, 1990). Cependant, il ne semble pas exister d’origine de réplication spécifique.
5. Expression des gènes intermédiaires et tardifs
La transcription de ces gènes nécessite la synthèse de novo de l’ARN polymérase virale et participation de protéines de la cellule hôte.
Les promoteurs des gènes intermédiaires et tardifs, présentés dans le tableau III, se composent d’une séquence de type TAAA(T/A) et TAAAT(G/A), respectivement, au site d’initiation de la transcription, et d’une séquence plus variable mais riche en AT en amont.
De nombreux gènes intermédiaires de la vaccine ont ainsi longtemps été confondus avec des gènes tardifs. Les ARNm intermédiaires possèdent une séquence polyA en 3' d'environ 30 nucléotides, non codées par le génome. Peu de gènes intermédiaires ont été étudiés. Pour la majorité, ils sont impliqués dans la régulation de la transcription des gènes tardifs.
Les deux catégories de gènes se distinguent cependant par la distance qui sépare la région variable en AT du site initiateur, plus courte dans les gènes tardifs.
L’extrémité 3’ des gènes intermédiaires et tardifs est très variable et aucune séquence signalant la fin de la transcription n’a été identifiée.
Les gènes intermédiaires et tardifs codent respectivement les facteurs de la transcription des gènes tardifs et précoces, ainsi que des protéines participant à la morphogenèse et l’assemblage des virions.
6. Assemblage et maturation des virions
Les virions s’assemblent dans des régions circonscrites du cytoplasme, identifiables en microscopie électronique. Ces sites, appelés virosomes, apparaissent dès 4 heures post-infection. Des membranes virales commencent par s’y accumuler. Leur origine est très discutée. Elles seraient synthétisées de novo (Dales & Siminovitch, 1961) ou dériveraient des membranes du compartiment intermédiaire entre le réticulum endoplasmique et l'appareil de Golgi (Sodeik et al., 1993) ou encore résulteraient de la fusion de petites vésicules issues d’organites. Après incorporation du génome, des éléments de la nucléocapside et maturation, les IMV sont formés. Certains s’entourent d’une double enveloppe supplémentaire dérivée de l’appareil de Golgi ou d’endosomes (Tooze et al., 1993) et se dirigent vers la surface cellulaire. Ce déplacement fait appel à la polymérisation, l’utilisation des microtubules et à la destruction du centrosome (Smith et al., 2002). Ils fusionnent avec la membrane plasmique et perdent leur enveloppe la plus externe pour libérer des EEV et CEV. Les CEV restent accolés à la membrane cellulaire puis sont expulsés par une microvillosité actinique. D’autres projections, formant des ponts cytoplasmiques, identifiées par microscopie électronique, sont aussi susceptibles de véhiculer les CEV vers les cellules voisines (Duteyrat et al., 2006).
II.
Le virus de la myxomatose
A.
Pathogénie et réponse immunitaire
La myxomatose est une maladie hautement létale du lapin Européen. La transmission du virus se fait principalement par le biais d’arthropodes piqueurs, vecteurs passifs.
Le MYXV infecte donc directement les cellules du derme, majoritairement les cellules dendritiques exprimant le CMH-II. Ces cellules représentent le site primaire de réplication du MYXV, avant migration vers le nœud lymphatique drainant, où il est détectable dès 24 heures post-inoculation. A ce niveau, le MYXV se réplique fortement au sein des lymphocytes T (LT) du paracortex lymphatique, entraînant une réduction massive du nombre de LT. Le virus est ensuite disséminé, via les lymphocytes infectés vers les autres tissus, notamment les poumons, la rate, les testicules et la peau.
L’évolution de la maladie dépend à la fois de la souche virale mais aussi de la résistance naturelle de l’hôte. Ainsi, cinq grades d’infectiosité peuvent être distingués (le grade I étant le plus élevé) selon le temps de survie, le taux de mortalité, l’aspect du myxome primaire, l’intensité et le temps de génération de la maladie, l’apparition ou non de signe nerveux, la teneur en virus et la virulence des lésions.
L’inoculation de la souche sauvage de MYXV T1 (Toulouse 1) de grade I, utilisée au laboratoire conduit à l’évolution de la maladie sous sa forme classique.
Les premiers signes cliniques apparaissent autour du 4e jour post-infection et se traduisent par une lésion cutanée érythémateuse au site d’inoculation évoluant sur plusieurs jours : le myxome primaire.
Dans les jours qui suivent, des myxomes secondaires apparaissent sur la face (oreilles, paupières et museau), puis au niveau des pattes et des régions dorsolombaire et anogénitale. L’apparition de ces myxomes signe la diffusion du virus par la voie lymphatique.
Parallèlement, l’infection par le MYXV entraîne une immunodépression qui se traduit par le développement de surinfections bactériennes de la sphère respiratoire et une
blépharoconjonctivite. On observe ainsi des écoulements muqueux puis purulents de la sphère oculo-nasale.
Les lapins infectés meurent généralement en une quinzaine de jours sous l’effet conjugué de dégâts tissulaires et des suites des infections bactériennes entraînant une détresse respiratoire et parfois des énucléations oculaires.
Figure 7 : Forme classique de myxomatose induite par la souche T1. On note au huitième jour post-infection de nombreux myxomes secondaires sur les oreilles et la face.(source UMR 1225, IHVV)
2. Réponse immunitaire.
La mise en place de la réponse immunitaire contre le MYXV commence dès l’infection des cellules dendritiques du derme et des LT (au niveau du nœud lymphatique) par le virus. Les lapins produisent ainsi des IgM (à durée de vie courte) et IgG persistantes contre le MYXV. Cependant, les vaccins à virus inactivés ne protègent pas les lapins de la myxomatose alors même que des anticorps sont produits.
Il semble donc que la réponse à médiation humorale ne soit pas suffisante dans le contrôle de l’infection par le MYXV, bien qu’elle joue un rôle non négligeable pour prévenir une réinfection.
La réponse contre le MYXV semble être principalement de type cellulaire. Outre que le MYXV infecte les cellules clés de la réponse cellulaire, les cellules dendritiques et les LT, il code de nombreux facteurs de pathogénicité qui détournent et interfèrent avec la mise en place de la réponse à médiation cellulaire. Ainsi, le MYXV possède la capacité de déréguler les molécules du CMH-I de surface des cellules infectées, altérant la présentation des antigènes viraux aux LT CD8+, acteurs de la réponse cellulaire. Il possède aussi des facteurs aux propriétés anti-apoptotiques, anti-inflammatoires, des inhibiteurs d’interféron, de chemokines et de cytokines. L’action cumulée de tous ces facteurs empêche la mise en place d’une réponse immunitaire à médiation cellulaire efficace. Enfin, il semble que des LT infectés par le MYXV soient, in vitro, incapables de répondre à un stimulus mitogène, empêchant leur prolifération, et ce dès quatre jours post-infection (Stanford et al., 2007).
Tous ces éléments indiquent que la réponse à médiation cellulaire joue un rôle prépondérant dans le contrôle immunitaire de l’infection par le MYXV.
B.
Facteurs de pathogénicité
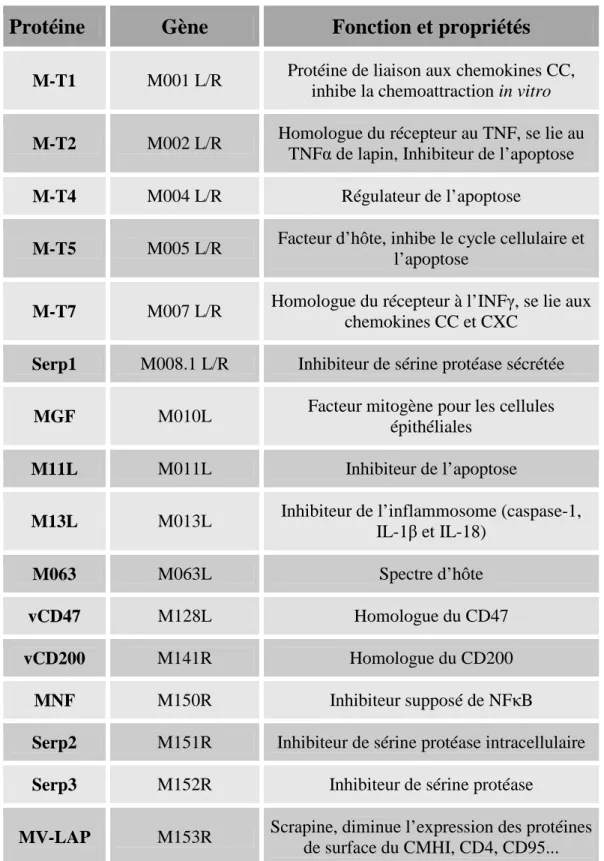
Contrairement aux virus ARN, pour qui la variation génétique joue un rôle prépondérant dans l’évasion aux réponses de l’hôte, les poxvirus codent de nombreux facteurs destinés à leurrer, inhiber ou détourner au profit du virus les voies de signalisation impliquées lors de la mise en place de la réponse immunitaire. Les facteurs de pathogénicité avérés sont listés dans le tableau IV.
Parmi ces facteurs de pathogénicité, nombreux sont des homologues de protéines cellulaires inactifs ou entraînant une réaction différente.
Les virorécepteurs et les virokines sont respectivement des pseudo-récepteurs mimant des récepteurs cellulaires et des pseudo-ligands. Les virorécepteurs sont généralement sécrétés ou exposés à la membrane cellulaire. Leur fonction est de lier les ligands cellulaires impliqués dans les défenses anti-virales et d’inhiber leur action par inhibition de compétition. Les virokines quant à elles, sont sécrétées et vont se lier à des récepteurs cellulaires, inhibant ou détournant les cascades de signalisation cellulaires qui découlent de l’interaction récepteur/ligand.
D’autres facteurs, pour lesquels l’homologie est moins flagrante, inhibent ou détournent les grandes voies de signalisation au sein même de la cellule. L’étude de certains facteurs a même permis d’identifier des mécanismes cellulaires physiologiques.
1. Facteurs anti-apoptotiques
a) L’apoptose
L’apoptose est un mécanisme de défense cellulaire qui consiste en la mort programmée des cellules âgées, présentant un dysfonctionnement, ou infectées. Elle peut être induite par deux mécanismes principaux : le premier, déclenché par des signaux intracellulaires (voie intrinsèque) et le second via des signaux externes (voie extrinsèque).
La voie intrinsèque ou mitochondriale
Cette voie fait intervenir des protéines anti-apoptotiques (Bcl-2 et Bcl-xl) et des protéines pro-apoptotiques de type BH3 (Bad, Bak, Bax et Bid) (Cory et al., 2003). Le déclenchement de l’apoptose repose sur l’équilibre entre ces deux types de facteurs.
Les protéines pro-apoptotiques sont des senseurs présents dans le cytoplasme. En réponse à un signal de stress, elles vont se transloquer à la surface des mitochondries où sont situées les protéines apoptotiques. L’interaction entre les protéines pro et anti-apoptotiques induit la formation de pores dans la membrane mitochondriale, entraînant le relargage du cytochrome c dans le cytoplasme où il va déclencher une cascade d’activation des caspases, effectrices de l’apoptose (Bras et al., 2005).
La voie extrinsèque ou des récepteurs de mort
La voie extrinsèque comprend deux modèles faisant intervenir les récepteurs au TNF (Tumor Necrosis Factor) (TNFR) : l’un est médié par le TNF et l’autre par l’interaction Fas-Fas ligand.
Le TNF est produit par les macrophages activés. En se liant au TNFR1, il initie l’activation de la caspase 8 (Ashkenazi & Dixit, 1999). La liaison du TNF peut aussi déclencher de manière indirecte l’activation de facteurs de transcription comme NFκB, impliqués dans la mise en place de la réponse inflammatoire.
Le récepteur Fas, ou CD95, se lie au récepteur FasL, induisant la formation du DISC (Death-Inducing Signaling Complex) avec les caspases 8 et 10. Il en découle une cascade d’activation de caspases conduisant à la mort cellulaire (Ashkenazi & Dixit, 1999).
b) Inhibition de l’apoptose par le MYXV
Les principaux facteurs intervenant dans l’inibition de l’apoptose sont représentés sur la figure 8.
M-T2 (virorécepteur)
M-T2 fut le premier virorécepteur identifié, et ce par homologie de séquence avec le domaine de liaison N-terminal des récepteurs cellulaires au TNF. M-T2 est sécrété sous deux formes : monomérique ou dimérique, ces deux formes se liant au TNF avec une affinité similaire à celle du récepteur cellulaire (Schreiber et al., 1996).
L’infection de lymphocytes de lapins (RL5) avec un MYXV délété de M-T2 se caractérise par une apoptose accrue des RL5, en comparaison avec une infection par la souche sauvage. Il agirait donc comme un pseudo-récepteur soluble, séquestrant le TNF avant qu’il n’atteigne les récepteurs cellulaires.
Cependant, l’ajout de MT-2 exogène dans le milieu de culture ne suffit pas à empêcher cette apoptose massive (Macen et al., 1996). De plus, des formes tronquées de M-T2, non sécrétées et incapables de se fixer au TNF, conservent leurs propriétés anti-apoptotiques. Il semblerait donc que M-T2 inhibe plutôt l’apoptose au niveau intracellulaire (McFadden et al., 1997; Schreiber et al., 1997).
M-T4
M-T4 est un facteur de virulence, localisé au niveau du réticulum endoplasmique (RE) des cellules infectées. L’infection de RL5 avec un MYXV délété de M-T4 se caractérise par une apoptose accrue (Barry et al., 1997). Son action anti-apoptotique empêcherait le clivage de BAP31, protéine du RE, associée aux protéines Bcl2 / BxL et la procaspase 8 (Hnatiuk et al., 1999).
De plus, l’interaction de M-T4 avec BAP31 semble inhiber le transport des molécules du CMH-I du RE vers la membrane cellulaire (Zuniga, 2002).
L’inoculation de lapins avec ce virus délété montre une atténuation des symptômes qui s’accompagne d’une forte réaction inflammatoire (Hnatiuk et al., 1999). M-T4 aurait donc une double fonction : anti-apoptotique et anti-inflammatoire.
M-T5
M-T5 est une protéine à motif ankyrine, ne possédant pas d’homologie avec des protéines cellulaires. Elle appartient à la super famille des protéines impliquées dans le spectre d’hôte, fonction tout d’abord associée à ce motif. L’infection de RL5 avec un MYXV délété de M-T5 est abortive. Cette infection est associée à une apoptose importante des RL5 et une rapide inhibition de la synthèse des protéines de l’hôte ainsi que des protéines virales. Cependant, si un tel mutant ne peut se répliquer sur lymphocytes de lapin, il conserve ses capacités réplicatives sur fibroblastes de lapin (Mossman et al., 1996a).
L’inoculation du MYXV délété de M-T5 à des lapins montre que le virus mutant ne diffuse pas à partir du site d’inoculation, entraîne une réponse inflammatoire rapide et efficace et ne permet pas la mise en place de la réponse cellulaire (Mossman et al., 1996a).
M-T5 joue aussi un rôle dans la régulation du cycle cellulaire. Par son interaction avec la culline 1, M-T5 entraîne l’ubiquitination de p27 et sa dégradation par le protéasome, permettant ainsi la progression du cycle cellulaire du stade G0/G1 au stade G2/M (Johnston et al., 2005b).
Enfin, M-T5 interagit et active Akt, facteur intervenant dans la régulation de l’apoptose et de la prolifération cellulaire (Wang et al., 2006).
M-T5 est donc, par son implication dans plusieurs processus visant à favoriser notamment la réplication virale, une protéine essentielle à la pathogénie du MYXV. Nous étudierons plus en détails ses mécanismes d’action dans la partie consacrée aux protéines à motif ANK.
M11L
M11L ne possède pas motif structural défini, à l’exception d’une séquence de 25 aa formant un domaine transmembranaire similaire à ceux présents dans les protéines anti-apoptotiques de la famille Bcl-2 (Everett et al., 2000). M11L se localise au niveau de la membrane externe des mitochondries où elle interagit avec le récepteur aux benzodiazépines, pour contrer l’apoptose cellulaire (Everett et al., 2002).
Une étude plus récente sur M11L démontre que, indépendamment de son interaction avec le récepteur aux benzodiazépines, la protéine forme également un complexe avec
la protéine Bak, facteur de la famille Bcl-2, pour inhiber le changement conformationnel et l’activation de la protéine pro-apoptotique Bax, et prévenir ainsi le relargage du cytochrome c (Su et al., 2006). Il semble donc que M11L soit impliqué dans plusieurs fonctions et mécanismes d’actions distincts, tous destinés à contrer les mécanismes apoptotiques déclenchés suite à une infection par le MYXV.
Serp2
Serp2 appartient à la superfamille des serpines (SERine Protéases INhibitor) (Petit et al., 1996). Un mutant délété de Serp2 se réplique de manière comparable au virus sauvage dans les RL5 et les fibroblastes de lapins. Cependant, une rapide apoptose au niveau du nœud lymphatique drainant le site d’inoculation est observée chez les lapins infectés avec le virus knockout pour Serp2, ceci indiquant que Serp2 a une réelle action anti-apoptotique in vivo. En outre les études histologiques montrent que Serp2 possède aussi un rôle anti-inflammatoire (Messud-Petit et al., 1998).
Figure 8 : Mécanisme d’échappement du MYXV à l’apoptose. Les protéines virales sont indiquées en bleu sur la figure.La protéine M-T2 sous sa forme sécrétée inhibe la fixation du TNFα à son récepteur le TNFαR. Sous sa forme réticulaire, elle empêcherait la translocation du TNFα-R à la membrane cellulaire. M-T4 inhibe l’activité de la protéine BAP31, protéine réticulaire associée aux protéines Bcl2/BxL et à la caspase-8. M-T5 inhibe l’apoptose en agissant sur le cycle cellulaire (p27). La protéine M11L est capable de se lier aux protéines mitochondriales Bak et Bax, inhibant leur activité pro-apoptotique. Serp2 interfère avec la voie des caspases.
Noyau Cytoplasme Membrane plasmique Mitochondrie Réticulum Endoplasmique Procaspase 8 ou 10 MYXV TNFα M-T2 M M--TT55 Récepteur au TNFα Serp2 M11L M-T4 Caspase 8 ou 10 Caspase 3 Procaspase 3 Dégradation de l’ADN Mort cellulaire Caspase 9 Procaspase 9 M-T2 Bax Bax Bax BH3 p27 Bcl2 BAP 31 BAP 31
2. Inhibition de l’Interféron
Les IFN (interférons) ont été nommés ainsi en relation avec leur capacité à interférer avec l’infection virale des cellules. C’est le mécanisme anti-viral par excellence. Le rôle des IFN de type I (α et β) et de type II (γ) dans la défense des cellules contre les virus a été mis en exergue du fait que de très nombreux virus ont développé des stratégies anti-IFN.
a) Synthèse des interférons
Les IFN de type I sont synthétisés par la plupart des cellules de l’organisme, l’ IFNα
étant principalement produit par les leucocytes et l’ IFNβ par les fibroblastes. L’infection virale conduit généralement à la présence d’ARN bicaténaire, considéré comme le principal inducteur de la synthèse d’IFN de type I. Il permet l’activation de la PKR (ARNdb dépendante) qui elle même active la kinase d’IκB (IKK). La phosphorylation d’IκB entraîne son ubiquitination puis sa dégradation via le protéasome, libérant ainsi le facteur de transcription NFκB (Maran et al., 1994). Ce dernier, transloqué dans le noyau, induit la synthèse d’IFN de type I ainsi que de nombreuse cytokines.
L’IFNγ, composant de la réponse cellulaire adaptative, est sécrété quant à lui par les lymphocytes T CD4 et CD8, suite à la présentation antigénique par les CPA (Cellules Présentatrices d’Antigènes) (Young, 1996). Les mécanismes de synthèse de l’IFNγ sont encore mal compris. Cependant, il est avéré que les interleukines IL-12 et IL-18 jouent un rôle déterminant dans la stimulation de la production de l’IFNγ dans une voie indépendante de la présentation des antigènes. De même, les cellules NK (Natural Killer) peuvent produire de l’IFNγ sans présentation d’antigènes (Singh et al., 2000).
b) Mode d’action des interférons
Les IFN de type I et II possèdent des récepteurs distincts, IFNAR et IFNGR respectivement. La fixation de l’IFN à son récepteur spécifique active une cascade de signalisation par l’intermédiaire du système Jak/STAT permettant ainsi la transcription d’un ensemble de gènes impliqués dans la résistance antivirale. Ces protéines empêchent la synthèse des protéines virales en bloquant la traduction.
On trouve ainsi l’oligoadenylate synthétase (OAS) qui polymérise l’ATP en 2’-5’ (les acides nucléiques étant normalement liés en 3’-5’). Cela active la RNase L, une endoribonucléase qui va dégrader tous les ARN, y compris les ARNm viraux (Rebouillat & Hovanessian, 1999).
On retrouve aussi la PKR (sérine-thréonine kinase), activée par l’IFN-α et l’IFN-β
(Clemens & Elia, 1997). Cette enzyme phosphoryle le facteur d’initiation de synthèse protéique eIF-2α, inhibant la traduction et contribuant ainsi à l’inhibition de la réplication virale (Clemens et al., 1993). L’activation de la PKR régule aussi d’autres facteurs de transcription tels que l’IRF-3 impliqué dans l’induction de l’apoptose induite par l’infection et c-Jun (Chu et al., 1999). Enfin, la PKR contrôle différents processus cellulaires comme la croissance cellulaire, la différenciation, l’apoptose, la réponse au stress mais aussi l’activité antivirale et anti-tumorale (Clemens & Elia, 1997).
c) Inhibiteurs poxviraux de l’interféron M-T7
M-T7 est la plus abondante des protéines virales sécrétées par les cellules infectées par le MYXV. Elle possède une forte homologie avec les récepteurs humains et murins de l’IFNγ (Upton et al., 1992). M-T7 est donc capable de lier l’IFNγ et de ce fait d’en inhiber l’action anti-virale. De plus, il a été montré que M-T7 est capable d’interagir avec des membres des familles de chemokines C, CC et CXC (Lalani & McFadden, 1997).
L’inoculation de MYXV délété pour M-T7 à des lapins se traduit par une augmentation du nombre d’infiltrats leucocytaires au niveau des lésions, ainsi qu'un grand nombre de
lymphocytes activés au niveau des nœuds lymphatiques et de la rate (Mossman et al., 1996b). M-T7 semble inhiber les connections entre CPA des tissus infectés et les lymphocytes des organes lymphoïdes secondaires.
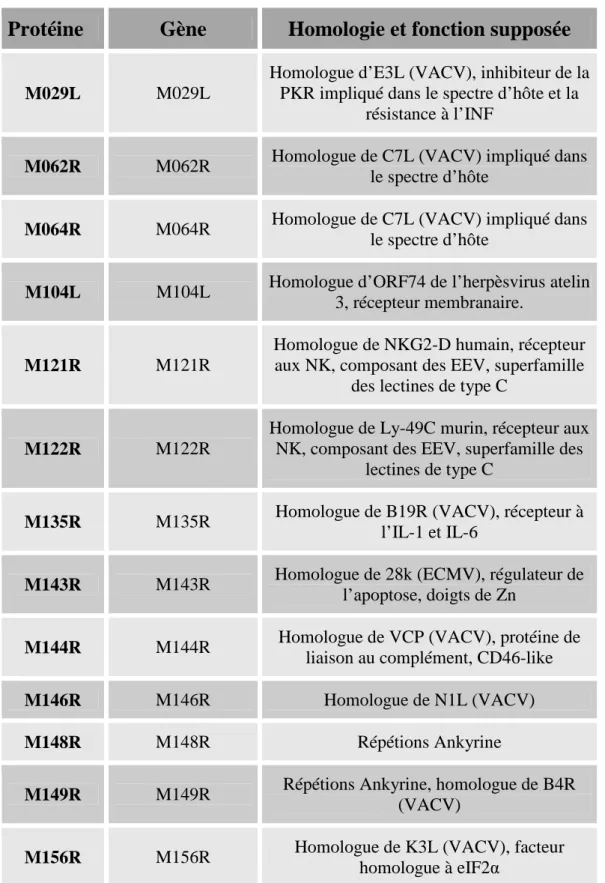
M029L et M156R
M029L et M156R sont respectivement les homologues d’E3L (VACV) et de K3L (VACV), facteurs impliqués dans la résistance à l’IFN.
E3L possède un domaine de liaison à l’ARNdb, essentiel pour la résistance à l’IFN et au large spectre d’hôte du VACV. E3L est aussi un inhibiteur de la PKR in vitro et in vivo, en interagissant directement avec la PKR (Davies et al., 1993). Enfin, E3L inhibe aussi le système OAS précédemment décrit (Rivas et al., 1998).
K3L est un homologue structural du facteur de transcription eIF2. En tant que pseudo-substrat de phosphorylation de la PKR, elle empêche la phosphorylation d’eIF2, inhibant la réponse anti-virale médiée par cette voie (Davies et al., 1993).
M029L et M156R, du fait de leur homologie avec E3L et K3L respectivement, sont suspectés de jouer un rôle dans la réponse anti-IFN, bien que n’ayant pas fait l’objet d’études approfondies.
3. Inhibition de l’activation des leucocytes
M-T1
M-T1 est une glycoprotéine sécrétée qui se lie aux chemokines CC, inhibant ainsi leurs propriétés chemoattractantes in vitro (Lalani & McFadden, 1997). Cela se traduit in vivo
par une augmentation de l’afflux de monocytes et de macrophages au site d’inoculation par un MYXV délété de M-T1 (Lalani et al., 1999). Cependant, la délétion de M-T1 n’a pas d’impact significatif sur l’évolution de la maladie.
M13L
M13L possède un domaine Pyrine caractéristique d’une superfamille de régulateurs de l’apoptose et de l’inflammation. M13L interagit avec ASC-1, composant de l’inflammosome, modulant l’activité de la caspase-1 et la maturation des interleukines IL-1β et IL-18 (Johnston et al., 2005a). La virulence réduite d’une souche de MYXV délétée de M13L est attribuée à son incapacité à infecter les lymphocytes (Johnston et al., 2005a).
M141R
M141R est une protéine associée aux membranes, homologue des protéines de la famille du CD200 cellulaire (OX-2), responsable de la régulation de l’activité des cellules myéloïdes (Cameron et al., 2005a). M141R est essentielle pour le développement de la myxomatose. D’un point de vue histologique, l’infection de lapins par un MYXV délété de M141R, se caractérise par un important recrutement et une forte activation des monocytes/macrophages ainsi que par des lymphocytes exprimant de forts taux d’oxyde nitrique au niveau des organes lymphoïdes infectés. De même, M141R réduit le taux d’activation des lymphocytes T dans les nœuds lymphatiques et inhibe la capacité des cellules T circulantes à répondre à la présentation antigénique (Cameron et al., 2005a). De plus, lors de l’infection de PBMC de lapin activées par le MYXV délété de M141R, une augmentation du taux d’IFNγ est détectée. Toutes ces données montrent que M141R est essentielle à la pathogénicité du MYXV en inhibant la capacité des macrophages à présenter les antigènes viraux aux lymphocytes T.
M128L
M128L est une protéine associée aux membranes, homologue au CD47 cellulaire (IAP, Integrin Associated Protein) connu pour sa régulation de la réponse immunitaire, notamment par son implication dans l’activation cellulaire, la modulation de l’adhésion, la mobilité et la phagocytose des leucocytes. D’un point de vue histologique, l’infection de lapins par un MYXV délété de M128L, se caractérise par une augmentation du nombre de cellules monocytes et macrophages activés au site d’infection et dans les tissus lymphoïdes (Cameron et al., 2005b).
MNF
MNF est une protéine à motif ANK. Elle possède 9 répétitions ANK, dont la huitième lui sert de signal d’import nucléaire. En transfection, dans des cellules activées au TNFα, elle colocalise avec NFκB, facteur clé de la réponse inflammatoire. Ce résultat concorde avec les analyses histologiques menées sur des lapins inoculés avec une souche de MYXV délétée pour MNF, montrant une forte réaction inflammatoire au site d’inoculation. MNF est donc un facteur anti-inflammatoire qui inhiberait la voie NFκB dépendante (Camus-Bouclainville et al., 2004).
Nous étudierons plus en détails ses mécanismes d’action dans la partie consacrée aux protéines à motif ANK.
MV-LAP
MV-LAP appartient à la famille des scrapines (Surface Cell Receptor Abductor Protein). Elle possède un domaine LAP (Leukemia Associated Protein) auquel est associée une fonction E3 ligase d’ubiquitine. Elle permet ainsi la dérégulation de molécules de surfaces telles que le CMH-I, CD95 (Guerin et al., 2002) et CD4 (Mansouri et al., 2003) en les ubiquitinant, entraînant ainsi leur dégradation par le protéasome (Collin et al., 2005). Les molécules du CMH-I permettent la présentation de peptides dérivés de protéines cytosoliques à la surface des cellules infectées (peptides du soi et issus d’antigènes viraux). Le peptide couplé au MHC-I en interagissant avec le récepteur TCR (T cell receptor) des LT CD8, induit la multiplication clonale des LT CD8 spécifiques de l’antigène.
Serp-1
Serp-1 est une serpine sécrétée. Elle se fixe et inhibe de nombreux facteurs tels que la plasmine, l’urokinase ou l’activateur de plasminogène. L’inoculation de MYXV délété de Serp-1 à des lapins se traduit par une augmentation des infiltrats de cellules mononucléées et une réponse inflammatoire plus prononcée au site d’inoculation (Lun
et al., 2005).
Serp-3
Serp-3 est la troisième serpine codée par le MYXV. L’inoculation de MYXV délété de Serp-3 à des lapins se traduit par une augmentation de la réponse inflammatoire au site d’inoculation et une absence de myxomes secondaires au niveau des nœuds lymphatiques, conduisant à une forte atténuation du pouvoir pathogène (Guerin et al., 2001).
4. Autres facteurs
MGF
MGF (Myxoma Growth Factor) est une glycorotéine sécrétée qui présente des similarités avec les membres de la famille des facteurs de croissance épithéliaux (EGF ; Epithelial Growth Factor). Le MGF est capable de réguler la prolifération et la différentiation cellulaire et de stimuler la mitose des cellules adjacentes. In vivo, le MGF est un facteur de virulence du MYXV en lui-même puisque la maladie est significativement atténuée lorsque des lapins sont inoculés avec un virus n’exprimant plus ce facteur de croissance (Opgenorth et al., 1992).
M063R
M063R est un facteur influençant le spectre d’hôte. Ainsi, un virus délété de M063R est incapable de se répliquer sur de nombreuses cellules de lapin (lignées cellulaires et cellules primaires), les gènes tardifs n’étant pas exprimés (Barrett et al., 2006). L’inoculation d’un tel virus à des lapins se traduit par une absence de signes cliniques (Barrett et al., 2006). Elle confère cependant une bonne protection contre la réinfection par une souche sauvage. Le mécanisme d’action de M063R reste inconnu même si des