Capillary assemblies in a rotating magnetic field
Texte intégral
Figure





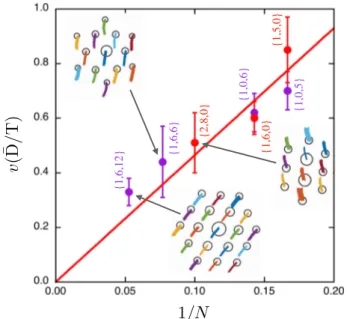
Documents relatifs
From our measured dust polarization angles, we infer a magnetic field orientation projected onto the plane of the sky (POS) that is remarkably ordered over the full extent of the
If the magnetic mode amplitudes are reduced, being for instance reduced to the fourth of the case just con- sidered in Figure 4, this differential heating is strongly attenuated as
In the absence of resonances, that will be specified below, the dramatic improvement on magnetic confinement induced by a low magnetic shear condition will be recalled in Section II
These results are extended in order to take into account strong- coupling effects and we show how measurements under a magnetic field may provide information
The main properties of this inner magnetospheric region are briefly recalled. The dynamical response of the plasma s phere to enhancements of geomagnetic activity
- In a nematic phase and, in particular, near a N -+ S, transition, the determination of the y1 twist viscosity coefficient in order to analyse the conditions
Theoretical and experimental studies of the spin-over mode, as well as a more general short-wavelength Lagrangian approach, demonstrate that the linear growth rate of the
However, it can be observed that, for an intermediate range of anisotropy—curves for K eff = 20 and 22 kJ/m 3 are good examples—the τ Z (φ) function is not symmetrical with respect
