HAL Id: hal-02823086
https://hal.inrae.fr/hal-02823086
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Analyse préliminaire de l’impact écologique de la
punaise invasive Leptoglossus occidentalis sur plusieurs
espèces de conifères européens. Première approche de la
structuration génétique des populations.
Sophie Guilbon
To cite this version:
Sophie Guilbon. Analyse préliminaire de l’impact écologique de la punaise invasive Leptoglossus occi-dentalis sur plusieurs espèces de conifères européens. Première approche de la structuration génétique des populations.. [Stage] Autres régions du monde. Université d’Orléans (UO), FRA. 2010, 32 p. �hal-02823086�
Sophie GUILBON
INRA Centre de Recherches Orléans, Unité de Recherche Zoologie Forestière, CS40001 ARDON, 2163 Avenue de la pomme de pin, 45075 Orléans cedex2, France
Co-encadrée par :
Alain ROQUES
Marie-Anne AUGER-ROZENBERG
Résumé La punaise prédatrice des cônes, Leptoglossus occidentalis Heidemann, est un
ravageur potentiellement important des vergers à graines sur la côte ouest de l’Amérique du Nord. Nous avons étudié l’effet de prédation de cette punaise sur des espèces de conifères européens largement répandus en Europe, particulièrement Pinus nigra et Pinus sylvestris. L’activité trophique des couples adultes sur deux semaines a réduit la quantité de graines pleines par cône de 30%. Une typologie des dégâts à 5 catégories de dégâts a été réalisée pour 7 essences de conifères européens et a révélé que la radiographie est une technique fiable et précise pour distinguer les graines ayant subi des dégâts. Aucune différence dans la consommation de graines n’a été observée selon le sexe et l’essence, suggérant que la gamme d’hôte de L. occidentalis peut s’avérer assez vaste en Europe, facilitant la progression et l’installation de l’invasif sur ce territoire. En parallèle, nous avons réalisé une première approche moléculaire à l’aide d’un marqueur mitochondrial pour estimer et comparer la variabilité génétique des populations introduites et celles des populations de la zone d’origine. Nos résultats montrent une forte variabilité génétique dans les deux zones, suggèrent un processus d’introductions multiples mais ne nous permettent pas encore de conclure sur l’origine précise de la zone nord-américaine d’où sont issues les populations invasives.
Mots clés : Cytochrome b, dégâts, espèce invasive, punaise, radiographie
Analyse préliminaire de l’impact écologique de la
punaise invasive Leptoglossus occidentalis sur
plusieurs espèces de conifères européens.
Première approche de la structuration génétique
des populations.
Master 2ème année « Biologie Biochimie
»
Spécialité « Ecosystèmes Terrestres et Action de l’Homme » Université d’Orléans – 2009/2010
Abstract The polyphagous western conifer-seed bug, Leptoglossus occidentalis Heidemann,
is a potentially serious pest of conifer seed orchards throughout western North America. We investigated the effect of feeding on European conifer species well ranged in Europe, particullary Pinus nigra et P. sylvestris. Feeding by a couple of adults on caged cones for a 2-wk period reduced the proportion of full seeds per cone by 30%. A 5 damage categories typology has been realised for 7 Europeans conifer species and has indicated that radiography is a reliable tool to distinguish damaged seeds. Seed consumption according to the sexe and the tree species was tested and none of these factors influence the predation, suggesting that the L. occidentalis’s host range could be quite large-scale in Europe, making the progression and the installation of the pest easier on the territory. We have also realised a first molecular approach thanks to a mitochondrial marker in order to estimate and compare the genetic variability between native and introduced populations. Our results show a hight genetic variability in both areas, suggest multiple introductions event but can’t allow us to conclude concerning the exact nord-american area, where the invasive populations come from.
Keywords : Cytochrome b, damages, invasive species, bug, radiography.
INTRODUCTION
L’Homme a joué depuis des millénaires un rôle majeur dans le transport et l’expansion des espèces invasives, avec deux changements radicaux notables correspondant à la fin du Moyen Age et au début de la Révolution Industrielle. De plus, au cours des dernières décennies, le monde est entré dans une nouvelle phase dans l’ampleur et la diversité des invasions biologiques avec l’ère de la Mondialisation (Hulme, 2009). Les conséquences de cette mondialisation se traduisent par l’augmentation des trafics commerciaux et des transports intercontinentaux, ce qui accentue les déplacements d’organismes en dehors de leur aire d’origine, malgré la présence de barrières naturelles et de distances considérables (Brockerhoff et al. 2006). Ces introductions peuvent mener à l’établissement de nouvelles
espèces invasives, se traduisant par la colonisation de nouvelles aires (Lozier et al. 2009) et pouvant constituer une menace pour la biodiversité locale, le fonctionnement des écosystèmes, l’agriculture ou la santé publique (Vitousek et al. 1997, Vila et al. 2010).
Les invasions biologiques concernant les insectes forestiers se situent parmi les plus préjudiciables pour les écosystèmes envahis. Ces dernières peuvent en effet avoir des effets spectaculaires sur la biodiversité, l’écologie et l’économie des régions affectées (Brockerhoff
et al. 2006 ; Liebhold et al. 1995), particulièrement sur les peuplements forestiers naturels ou les vergers à graines dans le cas d’espèces se développant aux dépens de cônes. En ce sens, l’importance croissante des vergers à graines comme source fiable de graines génétiquement améliorées utilisées dans les reboisements, a résulté en une prise de conscience du besoin de connaissance sur les insectes ravageurs de graines et sur leur impact (Bates et al. 2002b). Si le contrôle traditionnel de ces ravageurs compte en outre beaucoup sur l’utilisation d’insecticides à large spectre, l’impact environnemental négatif de ces derniers nécessite le développement de stratégies de gestion plus raisonnables de ces insectes.
L’introduction et la propagation de Leptoglossus occidentalis (Heidemann, 1910) (Hemiptera : Coreidae) représente un des cas particulièrement récents d’insectes exotiques invasifs. Cette punaise prédatrice des cônes de conifères est désormais considérée comme le principal ravageur des graines de conifères dans les vergers à graines commerciaux, non seulement sur la côte Ouest nord-américaine, de la Colombie-Britannique au Mexique, d’où elle est originaire (Blatt et Borden 1996) mais également sur la côte Est qu’elle a envahi dans la deuxième moitié du XXème siècle et sur le continent européen, dont la colonisation a débuté à la fin du XXème siècle. Malgré tout, l’ampleur de ses dégâts est restée longtemps sous-estimée car, au contraire des insectes se développant à l’intérieur de la graine ou du cône, L. occidentalis consomme le contenu de celles-ci de manière fugace et discrète. La punaise insère en effet son long stylet buccal à travers les écailles du cône, atteignant ainsi les graines, puis en extrait le contenu cellulaire, sans qu’il n’y ait de quelconques signes visuels extérieurs de dégâts du cône et de la graine (Bates et al. 2000b). De récents travaux américains ont par ailleurs permis de montrer que la plupart des graines vides et avortées observées en vergers, dont l’origine était préalablement attribuée à des problèmes de pollinisation ou abiotiques, correspondent en réalité à la prédation effectuée par ces punaises (Bates et al. 2000 ). L’apparente polyphagie de cet insecte en fait un invasif particulièrement redoutable. En Amérique du Nord, les principales plantes hôtes de L. occidentalis incluent
Pseudotsuga menziesii, Pinus strobus, P. resinosa, P. ponderosa, P. contorta mais l’insecte a aussi été trouvé sur Abies, Cedrus, Juniperus et Picea.
Des études antérieures sur l’impact de L. occidentalis ont fourni des estimations sur les dégâts potentiels causés par la punaise, mais ont été confinées et restreintes aux populations de la zone d’origine. Aucune étude d’impact de l’insecte n’a encore été menée sur le continent européen, alors que la gamme d’hôtes, même si elle reste encore mal connue, pourrait s’avérer assez riche. En dix ans, la punaise a étendu son aire de répartition sur toute
l’Europe où elle a déjà été observée sur Pinus sylvestris, P. nigra et P. halepensis ainsi que sur Picea sp, Cedrus sp, Abies sp et Juniperus sp (Villa et al. 2001) et menace les vergers à graines et la régénération naturelle des peuplements de conifères dans les pays envahis.
Si son expansion géographique a historiquement été bien documentée le nombre d’évènements d’introductions reste encore inconnu. Signalée pour la première fois en 1999 dans le Nord de l’Italie, elle a ensuite été repérée en Espagne près de Barcelone (2003), en France au Havre (2006), en Grande Bretagne à Weymouth College (2007), ainsi qu’en Belgique à Ostende (2007) (Dusoulier et al. 2007). Sa présence systématique dans les environs de ports commerciaux est très probablement due aux transports maritimes de bois en provenance des Etats-Unis sans que l’on sache encore si cette distribution correspond à l’expansion rapide de la population initialement introduite en Italie ou bien à plusieurs introductions différentes, ce qui semble cependant être l’hypothèse la plus probable. Sur une période comprise entre 2002 et 2007, l’espèce a envahie au moins 20 pays, parmi lesquels la Suisse, la Slovénie, l’Autriche, l’Allemagne, la République Tchèque (Lis et al. 2008).
D’ordinaire, les phases initiales d’invasion (introduction et établissement) sont associées à un effet fondateur – une perte de variabilité génétique par comparaison à la population source – et ce en raison d’un petit nombre d’individus introduits et d’une population de petite taille durant les premières générations (Dlugosh & Parker 2008). Cependant, des introductions multiples peuvent quant à elles enrichir la variabilité génétique des populations invasives, particulièrement lorsque plusieurs populations sources génétiquement différenciées contribuent à l’invasion (Genton et al. 2005). Des marqueurs
moléculaires de modes d’hérédité différents peuvent se révéler très informatifs pour étudier
l’histoire des invasions et pour évaluer la structure des populations d’espèces invasives. Les marqueurs génétiques peuvent ainsi être utilisés pour mesurer l’ampleur de la diversité
génétique dans les populations invasives, indiquer la perte de variation génétique durant les
bottlenecks de colonisation, ou encore tracer la ou les sources des introductions (Sakai et al. 2001, Ciosi et al. 2008). L’introduction récente et surtout la rapidité de l’expansion de la punaise en Europe expliquent la pénurie actuelle de travaux sur les routes d’invasions de L.
occidentalis. De telles informations constituent cependant des éléments essentiels pour la mise en place d’une stratégie de gestion efficace, en constituant le fondement de la prédiction de l’évolution des espèces. En effet, une fois l’espèce invasive établie et l’expansion engagée, l’éradication et le contrôle deviennent la priorité (Sakai et al. 2001), et c’est justement la connaissance du mode de dispersion ainsi que de la structure de la population qui peut améliorer et/ou dessiner les stratégies de gestion (Grapputo et al. 2005).
L’insecte possédant une progression fulgurante, on peut se demander quelle est l’importance des caractéristiques biologiques des populations envahissantes fondatrices et de la composition génétique de ces même populations dans la dynamique et le processus de développement spatio-temporel de cette punaise.
Mon travail s’insère d’abord dans la problématique de l’estimation du potentiel de
dégâts causés par L. occidentalis en Europe. Afin d’établir et d’ajuster les stratégies de
reforestation des peuplements attaqués dans les zones d’introduction, il apparaît nécessaire d’étudier l’impact possible de cette punaise envahissante sur les conifères européens, dans le but d’apprécier les capacités d’adaptation et le potentiel de la punaise à coloniser l’Europe. Sous conditions contrôlées, des tests comparatifs de la sensibilité de différentes espèces de conifères européens ont donc été mis en place. Les objectifs appliqués de ces tests sont (1) de mettre au point une typologie des dégâts par essence, (2) de tester la fiabilité de la radiographie dans la discrimination des différentes catégories de dégâts des graines, (3) de comparer l’effet de prédation selon le sexe des adultes et d’estimer les éventuelles
préférences alimentaires de la punaise vis-à-vis de différentes essences de conifères
européens puis (4) d’identifier l’impact de prédation sur cône de la punaise. Les résultats obtenus aideront à la mise au point d’une classification spécifique des dégâts sur graines par essence, et à apprécier les capacités d’adaptation de la punaise aux essences européennes. En parallèle, j’ai procédé à une étude préliminaire de la structure génétique des populations de L. occidentalis dans les zones natives et d’invasion. Mon objectif pour cette partie de l’étude est tout d’abord d’évaluer la variabilité génétique de différentes populations issues de la zone d’origine ou de la zone d’introduction, dans le but, à terme, de confirmer ou d’infirmer l’hypothèse selon laquelle la punaise aurait été introduite à plusieurs reprises dans différentes parties de l’Europe à partir d’une ou plusieurs populations d’Amérique du Nord. Cette analyse s’appuie sur l’utilisation d’un marqueur moléculaire mitochondrial correspondant à un fragment codant pour le cytochrome b. En effet, l’ADN du gène mitochondrial est sujet à une forte dérive génétique à cause de son hérédité maternelle et haploïde, et bien qu’une grande partie de sa variabilité puisse être perdue durant un goulot d’étranglement à l’occasion d’un processus d’introduction (bottleneck), il peut se révéler particulièrement informatif dans le cas d’invasions à introductions multiples. Les marqueurs nucléaires sont complémentaires et maintiennent de la variabilité sur une plus longue période (Grapputo et al. 2005) mais n’ont pu être utilisés dans le cadre de cette étude faute de temps.
MATERIEL ET METHODES
Partie 1 - Etude de la sensibilité de conifères européens face à L. occidentalis
Cette première approche a été menée au travers de deux expériences complémentaires. La première a porté sur l’impact de prédation de la punaise sur des graines nues de sept essences de conifère : le pin noir (Pinus nigra Arnold), le pin sylvestre (Pinus sylvestris L.), le sapin de Douglas (Pseudotsuga menziesii (Mirb.) Franco.), le sapin de Nordmann (Abies
nordmanniana (Steven) Spach), le pin d’halep (Pinus halepensis Miller), le pin maritime (Pinus pinaster Aiton) et le mélèze d’Europe (Larix decidua Miller). Cette expérience a été réalisée afin de bien identifier et d’associer les dégâts engendrés par la punaise sur des graines de différentes essences, en raison du problème à estimer de manière précise ces dégâts à partir de cônes dont on peut difficilement maîtriser l’état sanitaire initial des graines (Bates, 1997, Schowalter & Sexton, 1990). La seconde a porté sur l’impact de prédation de la punaise sur des cônes de deux essences de conifère, le pin noir et le pin sylvestre. Ces deux expériences ont été réalisées dans une chambre climatisée à 20°C avec 50% d’hygrométrie et une photopériode 16:8h (J:N).
1 – Dégâts engendrés par L. occidentalis sur graines nues
Dispositif : Pour bien apprécier l’impact de la prédation de la punaise sur la productivité des cônes (notamment parce qu’un avortement de la graine est possible sans une activité trophique de la punaise) et pour pouvoir dresser une typologie spécifique des dégâts sur graines par essence, on a procédé à la mise en relation des punaises avec des graines nues et pleines des sept essences de conifère présentées plus haut. Ces graines ont été obtenues via l’ONF, des producteurs de semences (Vilmorin) ou des gestionnaires de vergers à graines. Nous avons sélectionné les graines pleines par une première radiographie et seules celles-ci ont été présentées individuellement à des punaises adultes des deux sexes. Cette expérience « graine » a été effectuée durant la deuxième période du stage – soit de février à mai 2010– avec des punaises récoltées en 2009 dans la région Centre. Ces punaises ont donc été maintenues en hivernation au centre INRA durant l’hiver, dans un bâtiment faiblement chauffé (10°C). Pour éviter un choc thermique trop important, elles ont été transférées dans une salle à 15°C une semaine avant d’être utilisées pour le test alimentaire dans la chambre climatisée à 20°C. Les graines de chaque essence, préalablement réhydratées pendant 48h, ont
été présentées aux punaises par lot de 10 sur des portoirs en polystyrène dans un beurrier de 12 cm x 12 cm x 6,5 cm, par combinaison de trois essences en mélange (Pinus nigra (PN) /
Pinus sylvestris (PS) / Pseudotsuga menziesii (DG) ou Abies nordmanniana (AB) puis après 15J, les punaises ont été transférées sur un mélange Pinus pinaster (PM) / Pinus halepensis (PH) / Larix decidua (LD). L’expérience a compté 32 répétition pour chacun des deux sexes dans la premier mélange, et le second 13 répétitions pour les femelles, 15 pour les mâles. Radiographie des graines : Toutes les graines ont été radiographiées tous les deux jours de façon à pouvoir suivre les attaques des punaises à une échelle de temps assez fine. Les graines ont ainsi été radiographiées dressées sur leur portoir en polystyrène, afin de les maintenir immobiles et suivre le devenir de chacune d’entre elles au fil des jours. Le même appareil HP que précédemment a été utilisé, suivant les même propriétés (4’30’’ à 15 kV).
Mise au point d’une typologie spécifique des dégâts sur graines par essence A l’instar du travail de recherche effectué par Bates et al. (2000a), une échelle de dégâts par essence a été établie. Les graines ont ainsi été classées d’après l’ampleur des dégâts révélée par radiographie et dissection. Puis, parce que les dégâts sur graines causés par les punaises peuvent apparaître variables sur les radiographies (Bates et al. 2000a), les poids de graines de chaque catégorie pour les essences PN, PS, DG et AB ont été comparés afin de tester l’exactitude et la précision de la radiographie pour classifier les différents niveaux de dégât des graines. 3 à 390 graines (selon la disponibilité) de chaque catégorie ont été pesées par utilisation d’une balance analytique Mettler Toledo DeltaRange AG204.
Cinétique de consommation & Préférences alimentaires Cette approche a été réalisée uniquement sur la première série de graines présentées aux punaises (mélanges PN-PS-DG ou AB), le transfert des mêmes punaises sur un nouveau mélange PM-PH-LD au bout de 15 J ayant peut-être biaisé la consommation de ce dernier. Chaque graine consommée par essence a été comptabilisée au fil des jours afin de pouvoir évaluer le nombre de graines consommées quotidiennement et de pouvoir dresser un profil de consommation sur une période de deux semaines. Les préférences alimentaires ont été traitées d’après le nombre total de graines consommées pour chaque essence au bout des deux semaines et pour chacun des deux sexes. Analyses statistiques Toutes les variables ont été testées pour leur normalité en utilisant le test Shapiro-Wilk et en évaluant la répartition graphique des résidus. Le poids des graines de chaque catégorie de dégât a été analysé et comparé par un test non paramétrique Kruskal Wallis α = 0.05, l’analyse des résidus ne validant pas l’homocédasticité. L’effet du sexe sur la prédation des graines a été analysé d’après les valeurs du nombre de graines consommées par jour par individu, et comparé par un test non paramétrique Mann-Whitney α = 0.05.
Les données pour l’analyse des préférences alimentaires ont été normalisées par une transformation en log (x+0.1) et étudiées par une ANOVA à deux facteurs, les résidus répartis suivant une loi normale et justifiant de l’homocédasticité permettant l’utilisation de ce test paramétrique. Les résultats ont été considérés significatif au niveau α = 0.05.
2 - Impact de prédation de L. occidentalis sur cône
Dispositif : La sensibilité de Pinus nigra et de Pinus sylvestris a été testée en présentant du feuillage et des cônes de chaque essence à des couples de punaises, récoltées sur un même site dans les Hautes-Alpes en octobre 2009. Ce dispositif a été mis en œuvre dans des cages en polyester blanc de dimensions 36 X 56 cm, et d’une épaisseur de maille de 1mm. Deux catégories de test ont été réalisés d’octobre à novembre 2009 : un test monospécifique – en ne présentant qu’une seule essence aux punaises, et un test plurispécifique – en présentant les deux essences en mélange aux punaises, afin de pouvoir déceler une éventuelle préférence alimentaire de ces dernières. Pour l’expérience monospécifique, chaque couple de L.
occidentalis a été encagé avec trois cônes de la même essence, portés par des branches dont l’extrémité était placée dans l’eau : un cône témoin protégé par un petit sachet grillagé et donc non soumis à la prédation par les punaises, et deux cônes soumis à l’activité trophique des punaises. L’expérience plurispécifique s’est vue être constituée suivant le même dispositif : un cône témoin protégé et un cône accessible aux punaises, pour chaque essence. Ces cônes ont été exposés au couple de punaises pendant 15 jours afin de pouvoir déterminer l’impact de l’activité trophique de L. occidentalis sur la productivité des conifères. Au bout de la première quinzaine de jours, les cônes ont été retirés de la cage pour extraction des graines, et remplacés par d’autres selon le même protocole pour multiplier les résultats. L’expérience monospécifique a ainsi compté 25 répétitions pour le pin sylvestre et pour le pin noir et 6 répétitions pour le mélange des deux essences.
Extraction des graines et radiographie : Tous les cônes (témoins et soumis) ont été disséqués à la fin de l’expérience (au bout de 15 jours) afin d’en extraire les graines. Celles-ci ont été collées sur un papier autocollant A4 et radiographiées par un appareil HP Cabinet X-Ray system FAXITRON pendant 4’30’’ à 15 kV.
Analyses statistiques Le traitement des données a été conduit après avoir minutieusement dénombré le nombre total de graines par cône, puis le nombre par cône de graines pleines, vides et consommées par L. occidentalis. En outre, il est généralement reconnu dans les pépinières et les vergers à graines que le nombre de graines pleines par cône est une mesure attestant du potentiel productif d’un arbre (Portlock, 1996). Les dénombrements ont été traités
d’après les valeurs brutes des catégories exprimées ci-dessus, ceci en vue de pouvoir tenir compte de la variabilité inter – et intra – spécifiques relative au nombre de graines par cône, puis ont été converties en pourcentages, afin d’ajuster et d’homogénéiser ces valeurs brutes à comparer entre elles. Les dénombrements des graines des différentes catégories ont ensuite été analysés et comparés par un test Kruskal-Wallis, α = 0.05 pour des comparaisons multiples, ou par un test de Mann-Whitney α = 0.05 pour des comparaisons par paires. Seules les données du nombre de graines pour la première quinzaine de jours ont été analysés, les graphiques de variabilité du nombre de graines consommées sur 15 jours ne montrant pas de résultat homogène entre les deux répétitions pour un même couple de punaises.
Partie 2 : Etude de la variabilité génétique de différentes populations de L. occidentalis
1 – Matériel animal utilisé
L’ADN d’individus issus de populations collectées dans 9 pays européens a été extrait, séquencé et comparé à celui de quelques individus récupérés dans deux pays de la zone d’origine. Cette collecte a été permise par l’accord de collègues nord-américains et euroépens, spécialistes de cette punaise (Annexe D).
2 – Méthode
Extraction de l’ADN génomique : L’extraction de l’ADN génomique a été effectuée à partir d’une patte métathoracique. La partie du corps restante a été conservée dans l’éthanol 95% à -20°C pour permettre de nouvelles analyses et des observations morphologiques ultérieures si besoin. L’ADN génomique a été isolé et purifié en utilisant le Kit « NucleoSpin® Tissu » de Macherey-Nagel, et élué de la colonne par deux élutions de 50µl. L’ADN de 49 individus représentant 16 populations a été extrait . Enfin, la quantité et la pureté de l’ADN extrait ont été évaluées par mesure de densité optique au Nanodrop® (Thermo Scientific).
Amplification et purification de l’ADN : Un fragment du gène Cytochrome b (Cyt b) de 791 pb a été amplifié par PCR en utilisant les amorces CP1 (5’-GATGATGAAATTTTGGATC-3’) et CB2 (5’-ATTACACCTCCTAATTTATTAGGAAT-(5’-GATGATGAAATTTTGGATC-3’). Les amplifications ont été exécutées dans un volume total de 25 µl, avec 20 à 60 ng d’ADN génomique, du tampon 1x fina (Sigma), 1u de Red Taq Genomic DNA polymerase (Sigma) et 2,5 mM de MgCl2, 20mM
de dNTP et 1µM de chaque amorce. L’amplification des fragments de gènes mitochondriaux Cytb est réalisée selon le cycle suivant : un cycle initial de 5min à 94°C permettant une
dénaturation, 40 cycles d’amplification (dénaturation : 94°C 45 sec., hybridation : 46°C 45 sec et élongation : 72°C 1min30) et un cycle final de 5 min à 72°C ajoutant 1 pas d’élongation. Après vérification des amplifications par migration sur gel d’agarose 1% avec du Smartladder (Eurogentec), les produits PCR ont été purifiés sur colonne à l’aide du kit « NucleoSpin Extract II » de Macherey-Nagel.
Séquençage : Le séquençage a été réalisé à l’aide du Kit Abi Prism® Big Dye® Terminator V.3.1 Cycle Sequencing Kits (Applied Biosystems) en utilisant les mêmes amorces que celles utilisées précédemment, et les réactions de séquence ont été précipitées à l’éthanol. Les précipités ont été resuspendus dans 30 µl d’H20 avant d’être analysées par un séquenceur automatique Abi Prism® 3100 Genetic Analyser (Applied Biosystems). 49 individus ont été séquencés pour l’analyse génétique, pour in fine rendre compte de la structure génétique de L.
occidentalis dans ses zones d’origine et d’invasion.
Analyse des données moléculaires : Les séquences obtenues ont été corrigées manuellement et alignées avec Clustal W V1.4, intégré dans le logiciel BIOEDIT V7.0.9.0. Le nombre de sites variables a été calculé avec MEGA V3.1 et les haplotypes déterminés à partir des alignements et à l’aide du logiciel DNASP v5.10. La diversité haplotypique H et la diversité nucléotidique π ont été également été obtenues avec DNASP, uniquement pour les deux grands groupes « zone d’origine » « zone d’introduction », le faible nombre d’échantillons par localité étant assez faible pour en tirer des résultats informatifs. Les relations phylogénétiques entre les différents haplotypes ont été représentées par un réseau d’haplotypes (parcimonie statistique) obtenu avec le logiciel TCS V1.21 (Clément et al. 2000).
RESULTATS
Partie 1 - Etude de la sensibilité de conifères européens face à L. occidentalis
1.1 - Typologie spécifique des dégâts par essence et Validation de la détermination des dégâts par radiographie
La prise alimentaire particulière de L. occidentalis (Annexe A) induit que les graines qu’elle endommage sont quasiment indemnes de symptômes extérieurs visibles et de traces ostensibles de prédation (Bates et al. 2000a); l’orifice de piqûre sur le péricarpe de la graine est l’unique preuve visuelle de dégât de la graine avec la présence d’un reliquat de salive autour du trou de perforation (Fig.1A). Le diamètre de cet orifice a ainsi pu être estimé à
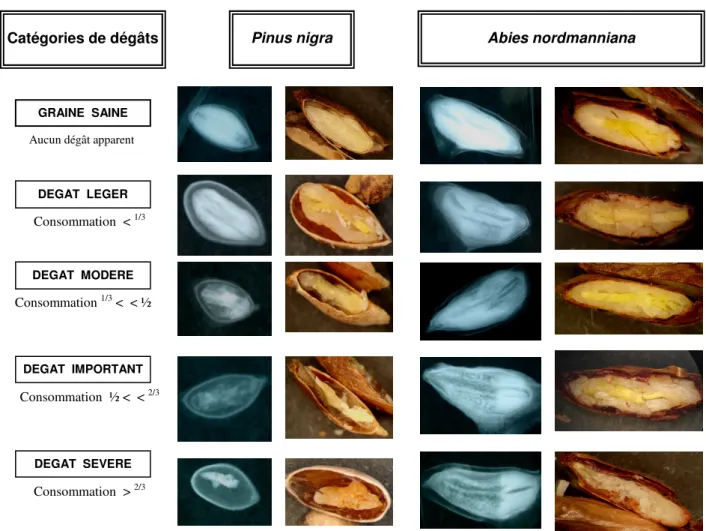
seulement 0, 004 mm. Sur une graine de coloration externe foncée, le dégât apparaît souvent sous la forme d’un minuscule trou blanc (Fig. 1B). Ce fin orifice est causé par la digestion partielle de l’enveloppe de la graine par la salive de l’insecte. Dans certains cas cependant, l’orifice de piqûre se referme après que la punaise ait retiré son stylet (Debarr, 1970). Une typologie des dégâts engendrés par L. occidentalis a été établie pour les sept essences de conifères européens testées. Pour chaque essence étudiée, ces dégâts sont classés en 5 catégories selon l’interprétation radiographique de la graine : graine saine (pas de dégât apparent), dégât léger (moins du 1/3 de la graine consommé), modéré (consommation entre le 1/3 et la moitié de la graine), important (consommation entre la moitié et les 2/3 de la graine), sévère (plus des 2/3 de la graine consommés par la punaise) (Fig.2 et Annexe B).
Figure 1 : Photographies illustrant (A) les reliquats de salive effectués par L. occidentalis sur le péricarpe d’une
graine et (B) le trou de perforation de la succion.
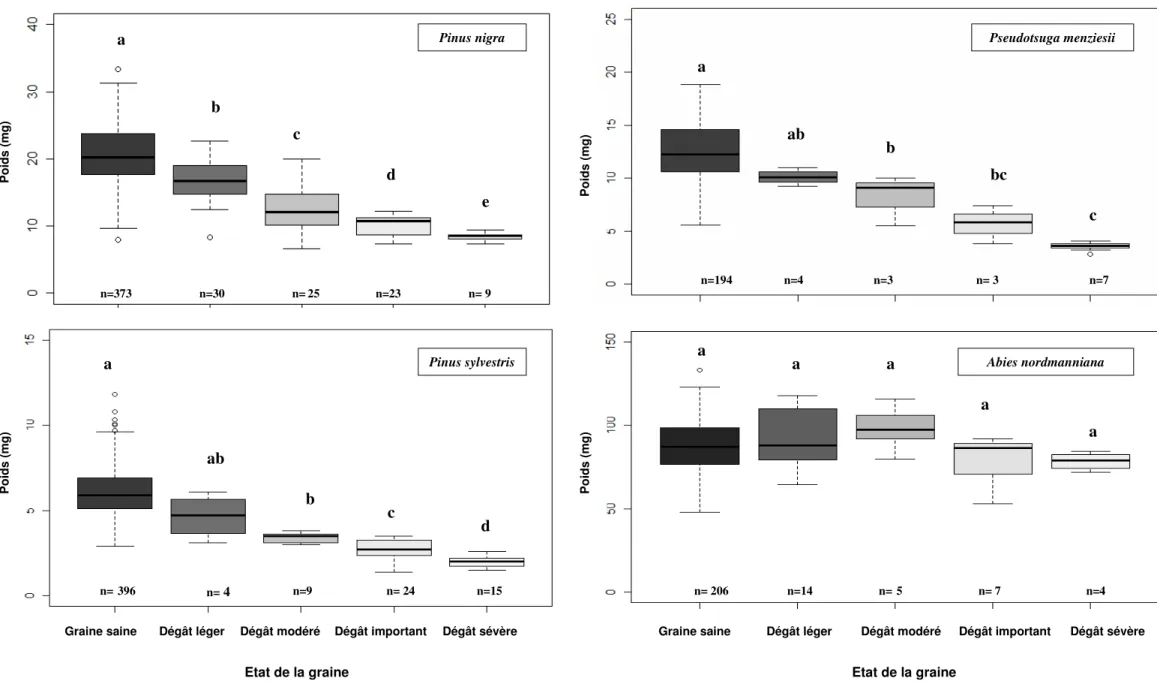
L’état interne des graines endommagées, légèrement, modérément, de façon importante et sévère, supporte les déterminations radiographiques des dégâts ; l’épuisement progressif des tissus du mégagamétophyte et de l’embryon est d’autant plus important que la graine radiographiée est sévèrement endommagée. Les typologies suivent les mêmes tendances et sont relativement semblables pour PN, PS, DG, PH, PM, LD. Elle diffère pour AB dans le sens où l’on ne distingue pas nettement d’épuisement du contenu de la graine suite à la prédation de L. occidentalis. La dégradation du mégagamétophyte apparaît indéniable, l’épuisement de l’embryon, même s’il apparaît plus difficile à mettre en évidence fait très certainement l’objet d’une dégradation par la punaise au vu de nos résultats. Les poids moyens des graines pleines et des graines de chaque catégorie de dégâts se rangent en une distribution linéaire décroissante pour PN, PS et DG (Fig.3), ce qui est cohérent avec l’attribution antérieure des dégâts révélée par radiographie et par dissection. Cette distribution ne suit cependant pas une tendance décroissante pour AB. Les différences de poids entre chacune des catégories de dégâts ne sont pas significatives pour toutes les essences malgré des états différents du gamétophyte selon les dégâts. Elle l’est uniquement et de façon complète pour P.nigra (X2 = 140.42, df=4, p<2.2.10-6).
Figure 2 : Typologie des dégâts pour deux essences de conifères européens, Pinus nigra et Abies nordmanniana. Les graines ont été exposées pleines à des adultes de L. occidentalis pendant 15 jours.
1.2 - Comparaison de l’effet de prédation selon le sexe
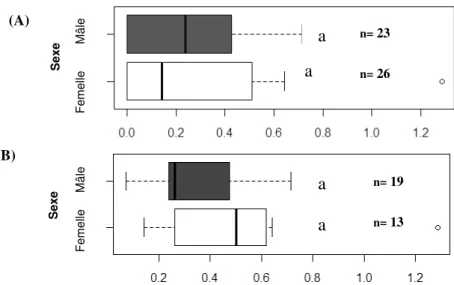
52% des femelles mises en test ont consommé des graines qui leur été présentées, contre 75% chez les mâles. Parce que nous ne savons pas si la non consommation par les individus peut être à relier ou non à un stress dans le conditionnement en boîte, à l’âge ou à l’état physiologique des insectes, nous avons conduit nos analyses statistiques sur la totalité des individus (à la fois les punaises ayant consommé et celles n’ayant pas consommé durant les deux semaines de l’expérience) et sur les insectes consommateurs uniquement. Dans les deux cas, le nombre de graines consommées par insecte et par jour ne diffère pas significativement selon le sexe (test de Mann-Whitney, p=0,7665 pour la totalité des individus p=0,1907 pour les consommateurs uniquement), en s’élevant chez les consommateurs à environ 0,40 graines consommée en moyenne par jour (Fig.4). En outre, il a pu être observé que les femelles ont attaqué un maximum de 4 graines par jour, contre 1 ,7 pour les mâles en moyenne.
Pinus nigra Aucun dégât apparent GRAINE SAINE DEGAT IMPORTANT DEGAT SEVERE Consommation < 1/3 Consommation 1/3 < < ½ Consommation ½ < < 2/3 Consommation > 2/3 Abies nordmanniana Catégories de dégâts DEGAT LEGER DEGAT MODERE
Figure 3. Poids des graines de quatre essences de conifères exposées à L. occidentalis et classés d’après les cinq catégories de dégâts révélées par radiographie. Les nombres
en dessous des boîtes à pattes indiquent le nombre de répétitions. Les barres d’erreur avec une même lettre ne sont pas significativement différentes, test Kruskal-Wallis , P<0.005. n=373 n=30 n=25 n=23 n= 9 n= 4 n=9 n= 24 n=15 n=194 n=4 n=3 n= 3 n=7 n= 206 n=14 n=5 n= 7 n=4 n=396
Graine saine Dégât léger Dégât modéré Dégât important Dégât sévère
Etat de la graine Etat de la graine
Pinus nigra Pinus sylvestris Pseudotsuga menziesii Abies nordmanniana P o id s ( m g ) P o id s ( m g ) P o id s ( m g ) P o id s ( m g )
Graine saine Dégât léger Dégât modéré Dégât important Dégât sévère
a b c d e a ab b c d a ab b bc c a a a a a
Figure 4. Comparaison de la prédation par les adultes mâles et femelles de L. occidentalis sur des graines de
conifère. Le graphique (A) tient compte de tous les individus de l’expérience, le graphique (B) uniquement des adultes consommateurs. Les barres d’erreur avec une même lettre ne sont pas significativement différentes, test de Mann-Whitney, P<0.005.
Les radiographies des graines réalisées tous les deux jours nous ont permis de dresser un profil de consommation des punaises consommatrices selon le sexe sur une période de deux semaines. La figure 5-A nous permet de visualiser une consommation croissante des graines par la punaise à partir du premier jour de consommation, à la fois pour les mâles et les femelles. Les femelles se révèlent être un peu plus voraces que les mâles tout au long de la période de prédation, sans que ces surconsommations par rapport aux mâles ne soient significatives. En outre, cette consommation croissante de graines au fil des jours ne se stabilise pas au cours de la période étudiée. Les profils journaliers de consommation (Fig.
5-B) révèlent quant à eux que les deux sexes ont une cyclicité dans la prise alimentaire, ne
consommant pas des graines quotidiennement. La prédation de graine par les femelles apparaît en outre plus soutenue que les mâles au fil des jours.
1.3 - Susceptibilité des essences testées et préférences alimentaires de L. occidentalis
Le nombre total de graines attaquées par les mâles et les femelle de L. occidentalis au bout des deux semaines d’exposition ont été comparés pour chaque essence dans les deux modalités de test (mélange DG-PN-PS et AB-PN-PS). Les analyses prenant en compte la totalité des individus ou uniquement les consommateurs donnent la même tendance. Pour la première modalité, nous ne mettons pas en évidence d’effet essence (F=2.51, df=2, p=0.09370) mais un effet sexe (F=4.4659, df=1, p=0.04056).
a a a
a
Nombre moyen de graines consommées par jour par individu
S e x e S e x e (A) (B) F e m e lle M â le F e m e lle M â le n= 19 n= 13 n= 26 n= 23
0 1 2 3 4 5 6 Es sence N o m b re d e g ra in e s a tt a q u é e s p a r in d iv id u a u b o u t d e d e u x s e m a in e MALE FEMELLE 0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5 N o m b re d e g ra in e s a tt a q u é e s p a r in d iv id u a u b o u t d e d e u x s e m a in e MALE FEMELLE 0 1 2 3 4 5 6 7 8 9 10
1er J 3ème J 5ème J 7ème J 10ème J 12ème J 14ème J Jours de consommation C u m u l d u n o m b re d e g ra in e s c o n s o m m é e s p a r in d iv id u MALE FEMELLE 0 0,5 1 1,5 2 2,5 3 3,5
1er J 3ème J 5ème J 7ème J 10ème J 12ème J 14ème J Jours de consom m ation
N o m b re d e g ra in e s c o n s o m m é e s p a r jo u r p a r in d iv id u male f emelle
Figure 5. Comparaison de la cinétique de consommation par les adultes mâles et femelles de L. occidentalis sur
des graines de conifère sur une période de 2 semaines. (A) Cumul du nombre de graines consommées et (B) Nombre de graines consommées par jour pendant deux semaines. Les barres d’erreur avec une même lettre ne sont pas significativement différentes, test de Kruskal-Wallis, P<0.005. Les astérisques sous l’axe des x indiquent les jours pour lesquels le nombre de graines consommées quotidiennement diffère significativement selon le sexe, test de Mann-Whitney, P<0.005.
Il n’y a pas d’effet interaction des deux facteurs (F=1.9245, df=2, p=0.15860). Les femelles consomment ainsi significativement davantage de graines que les mâles, mais ces choix alimentaire ne diffèrent pas selon l’essence. Pour la seconde modalité, il n’y a pas d’effet essence, sexe et interaction de ces deux facteurs ; les insectes de chaque sexe consomment égalitairement ces trois essences.
Fig.6. Nombre de graines consommées par individu et par essence sur une période de 2 semaines dans les
mélanges DG-PN-PS (A) et AB-PN-PS (B).
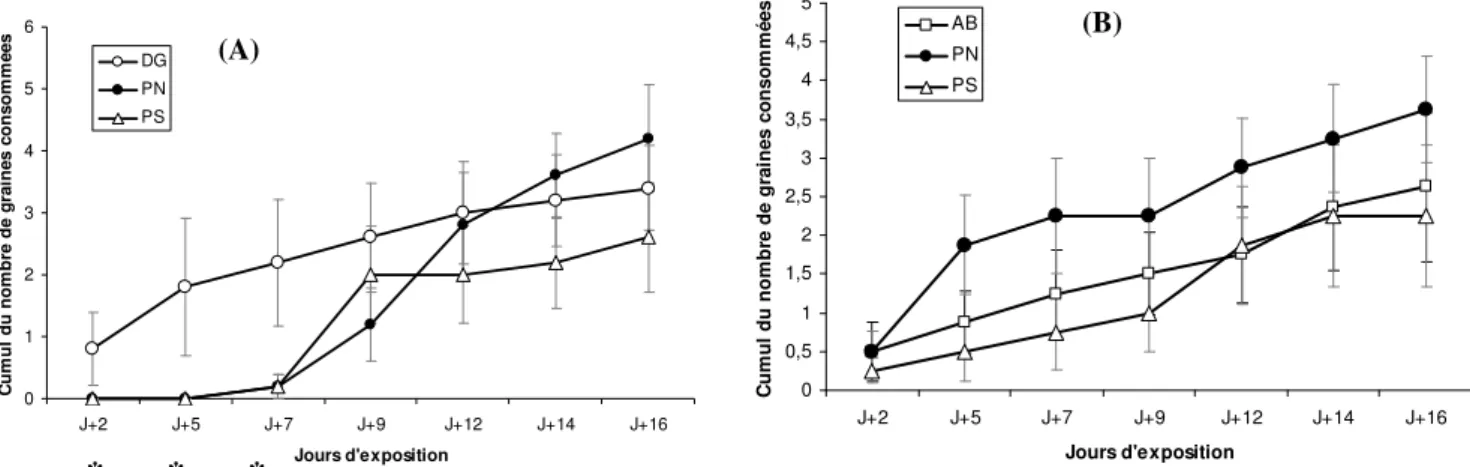
Lorsque nous regardons la façon dont s’orientent les choix de consommation sur une durée de deux semaines parmi les insectes ayant consommé les trois essences en mélange, nous mettons en évidence que pour le mélange DG-PN-PS, une préférence alimentaire significative (p<0,05) se dessine durant la première semaine d’exposition des graines aux punaises, en
Essence
Sapin de Douglas Pin noir Pin sylvestre
(A) (B)
(B) (A)
Sapin de Nordmann Pin noir Pin sylvestre
Essence a ab b bc bc bc bcd bcde cde cde cde de de e * *
0 1 2 3 4 5 6 J+2 J+5 J+7 J+9 J+12 J+14 J+16 Jours d'exposition C u m u l d u n o m b re d e g ra in e s c o n s o m m é e s DG PN PS 0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5 J+2 J+5 J+7 J+9 J+12 J+14 J+16 Jours d'exposition C u m u l d u n o m b re d e g ra in e s c o n s o m m é e s AB PN PS
faveur des graines de sapin de Douglas. Cette dernière constitue la première essence consommée par les punaises, puis la consommation s’oriente progressivement vers la consommation de graines de pin sylvestre et de pin noir, cette dernière essence s’avérant être la plus consommée au terme de l’expérience. Pour le mélange AB-PN-PS, les trois essences sont prédatées de façon équivalente tout au long de la période de consommation, bien que le pin noir termine l’essence préférée à la fin de l’expérience, sans que cette préférence soit significative.
Figure 7. Cinétique de consommation de 3 essences en mélange (P. nigra, P. sylvestris et P. menziensii (A) ou
P. abies (B)). Les astérisques sous l’axe des x identifient les jours pour lesquels la consommation de graine est significativement différente selon l’essence (Test de Mann Whitney, p<0,05).
1.4 - Impact de prédation sur cône de L. occidentalis
Une moyenne +/- SE de 33, 4 ± 0,93 graines ont été extraites par cône de Pinus
sylvestris et 24,6 ± 1,5 graines par cône de Pinus nigra (Annexe C).
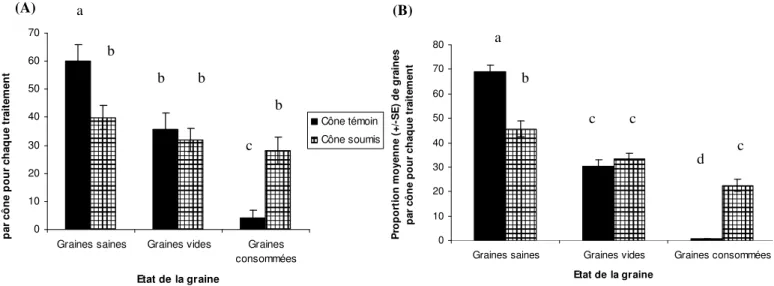
Dans le test monospécifique, pour chaque traitement (cône témoin/cône soumis) ainsi que pour chaque essence, les graines vides ont représenté un peu plus de 30% des graines totales produites (Fig.8). Les proportions relatives de ces graines vides dans les deux essences restent non significativement différentes entre cône témoin et cône soumis (test Mann-Whitney, p=0.6937, pour le pin noir et p=0.5239 pour le pin sylvestre). Pour le pin sylvestre, il a été trouvé un effectif significativement supérieur de graines vides dans les cônes soumis à la prédation des punaises que dans les cônes témoins (test de Mann Whitney, p = 0.02951), mais le nombre total de graines est inférieur dans le dernier cas (Annexe C). Pour un même nombre total de graines dans les cônes des deux traitements chez le pin noir, il n’y a pas de différence significative quant au nombre de graines vides (test de Mann Whitney, p= 0.7582) (Annexe 3). La proportion de graines consommées dans les cônes exposés aux punaises et pour les deux essences en condition monospécifique, atteint 30%, et est significativement plus
(A) (B)
*
0 10 20 30 40 50 60 70
Graines saines Graines vides Graines consommées Etat de la graine P ro p o rt io n m o y e n n e ( + /-S E ) d e g ra in e s p a r c ô n e p o u r c h a q u e t ra it e m e n t Cône témoin Cône soumis 0 10 20 30 40 50 60 70 80
Graines saines Graines vides Graines consommées
Etat de la graine P ro p o rt io n m o y e n n e ( + /-S E ) d e g ra in e s p a r c ô n e p o u r c h a q u e t ra it e m e n t
forte que dans les cônes témoins (test de Mann Whitney, p=3.13.10-5 et p=2.199.10-7 respectivement pour P. sylvestris et P. nigra) (Fig.8 et Annexe C). En outre, la quantité moyenne de graines consommées par L. occidentalis (toujours en monospécifique) est plus importante chez le pin sylvestre que chez le pin noir (7,8 contre 4,7 graines par cône, Annexe
C). On constate avec étonnement une proportion moyenne en graines consommées de 4%
dans les cônes de pin noir témoins protégés (Fig.8). Pour les graines pleines, la prédation de la punaise réduit cette quantité d’un peu plus de 30% chez le pin sylvestre et le pin noir (Fig.8).
Figure 8 : Comparaison de l’effet de prédation de L. occidentalis sur la proportion de graines de Pinus nigra (A) et Pinus sylvestris (B). Les moyennes avec des lettres identiques ne sont pas significativement différentes
(test Kruskal-Wallis ; P < 0,05)
Pour l’expérience plurispécifique, les résultats suivent les mêmes tendances que celles de l’expérience monospécifique ; environ 30% de graines vides par cône et moins de graines pleines dans les cônes soumis aux punaises que dans les cônes témoins bien que cette différence ne soit pas significative (p=1 et p=0.4704 respectivement pour le pin noir et le pin sylvestre) (Fig.9). On constate toujours la présence de graines consommées dans les cônes témoins de pin noir (Fig.9). La quantité de graines consommées ne diffère pas significativement pour le pin noir entre les cônes soumis et les cônes témoins (p=0.0962) mais il y a significativement davantage de graines consommées dans les cônes soumis que dans les cônes témoins pour le pin sylvestre (p=0.01006). La quantité de graines consommées quant à elle ne diffère pas significativement selon l’essence (p=0.9346) mais on observe uniquement une très légère surconsommation de graines de pin sylvestre par rapport au pin noir (4,2 contre 3,5 graines/cône). (A) a (B) b b b c b a b c c c d
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
TEMOIN PN SOUMIS PN TEMOIN PS SOUMIS PS Nature du cône P ro p o rt io n m o y e n n e ( + S E ) d e g ra in e s p a r c ô n e p o u r c h a q u e tr a it e m e n t Graines consommée Graines vides Graines pleines
Figure 9 : Comparaison de l’effet de prédation de L. occidentalis sur la proportion de graines (A) et le nombre
de graines consommées par cône (B) de Pinus nigra et Pinus sylvestris en mélange. Les moyennes avec des lettres identiques ne sont pas significativement différentes (test Kruskal-Wallis ; P < 0,05)
Partie 2 : Etude de la variabilité génétique de différentes populations de L. occidentalis
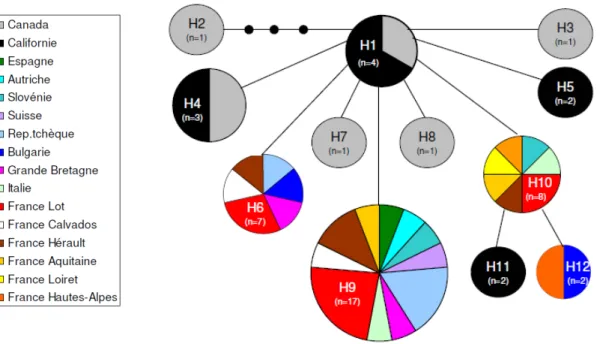
49 séquences ont été obtenues, de 417 pb chacune. Un total de 12 haplotypes a été trouvé parmi les 16 localités et les 49 individus analysés (Fig.10 et Annexe D). Parmi ces 12 haplotypes, 8 sont présents exclusivement dans la zone d’origine et 4 dans la zone d’introduction et on ne retrouve aucun haplotype partageant à la fois des individus de la zone d’origine et de la zone d’introduction (Fig.10 et 11). L’haplotype H9 est le plus commun dans la zone d’introduction (partagé par la moitié des individus) (Annexe D). 4 haplotypes sont représentés par un seul individu (H2, H3, H7, H8), correspondant tous à des insectes issus de la zone d’origine. Notre réseau d’haplotypes montre une structure en étoile à partir de l’haplotype H1, auquel se rattachent les haplotypes restants. La majorité d’entre eux n’en diffère que par un seul pas mutationnel, sauf pour H2 (4 pas mutationnels) puis H11 et H12 (2 pas mutationnels). La diversité génétique des populations de L. occidentalis est plus forte dans les zones natives que dans les zones d’introduction, mais celle-ci reste néanmoins relativement riche dans la zone d’introduction (Fig. 10 et Annexe D). Malgré le faible nombre de localités et d’individus obtenus dans la zone d’origine, on observe une légère structuration génétique dans la zone d’origine avec la présence de 2 haplotypes retrouvés à la fois en Californie et au Canada (H1 et H3) (Fig.11).
PN Témoin PS Témoin PS Soumis
Nature du cône N o m b re m o y e n d e g ra in e s p a r c ô n e ab b a b (A) (B) PN Soumis
Figure 10 : Réseau d’haplotype (H) construit avec TSC sur le fragment de Cyt b de 417pb. Chaque cercle
représente un haplotype. Chaque couleur représente une localité. La taille des cercles et des traits est représentative du nombre d’individus.
Figure 11 : Distribution géographique des 12 haplotypes parmi les 16 localités échantillonnées. A l’inverse de la
figure précédente, chaque camembert représente une localité et chaque couleur un haplotype. La taille des camemberts n’est pas proportionnelle au nombre d’individus récoltés.
DISCUSSION
Partie 1 - Etude de la sensibilité de conifères européens face à L. occidentalis
Il a été montré que le mode de consommation de L. occidentalis induit que ses indices de prédation ne peuvent seulement être visualisés qu’après un examen minutieux de chaque péricarpe de la graine, nécessitant un agrandissement assez considérable de ces derniers (X10 voir X20). Les poids de chacune des catégories de dégâts ne sont totalement et significativement différents les uns de autres que pour P. nigra. Ces résultats confirment les travaux que Bates et al. on réalisé en 2000 sur le Sapin de Douglas (2000a). En revanche, ces différences significatives restent incomplètes pour les autres essences. Cette non significativité dans la distinction des différents dégâts par le poids est très certainement due au manque de répétition des mesures effectuées (Fig. 3). Il apparaît en outre important de discuter de la précision des pesées des graines des différentes essences. La précision de la balance pourrait révéler des écarts différents selon les graines d’essence différente. Cependant, la relation étroite entre (a) les quatre catégories de dégâts attribuées par la radiographie (Fig. 1), (b) le poids correspondant des graines (Fig. 2) ainsi que (c) la morphologie interne des graines de chaque classe de dégâts (Fig.1) indique qu’une analyse radiographique seule s’avère suffisante et particulièrement précise pour catégoriser les dégâts sur graine de P. nigra, P. sylvestris et P. menziesii causés par L. occidentalis. En revanche, l’assignation de la gravité des dégâts par pesée et par radiographie apparaît davantage problématique pour Abies nordmanniana. Quoiqu’il en soit, il apparaît que, dans tous les cas, chacune des trois méthodes énoncées reste insuffisante pour permettre de donner des affirmations quand aux causes des dégâts de graines, particulièrement pour les graines complètement évidées. Ces graines vides peuvent en effet être le résultat d’une prédation par
L. occidentalis tout comme elles peuvent être le résultat d’un avortement de la graine en raison d’explication d’ordre reproductif comme un manque de pollinisation ou de fertilisation (Schowalter & Sexton, 1990, Bates & Borden 2002a). Cette remarque souligne le besoin d’une méthode précise et fiable permettant la discrimination des dégâts causés par la punaise de ceux générés naturellement. A ce titre, un diagnostic biochimique capable de détecter les résidus enzymatiques salivaires laissés par la punaise, pourrait constituer une preuve dans l’attribution des graines évidées par la punaise (Bates et al. 2000a).
Nos tests sous conditions contrôlées ont révélé un effet de prédation de la punaise sur les différentes essences testées. L’effectif de mâles prédateurs s’est révélé être plus important que celui des femelles. Nous ne pouvons pas conclure à un caractère davantage vorace des mâles de par cette observation ; le conditionnement des individus dans les boîtes, ainsi que l’âge ou l’état physiologique des insectes a peut-être conduit à une perturbation dans l’activité trophique de certains insectes. Dans notre étude, parmi les punaises consommatrices, ce sont les femelles qui causent le peu plus de dégâts, bien que cette surconsommation par rapport aux mâle ne soit pas significative. Les études ayant porté sur l’impact de la prédation de L.
occidentalis n’ont pas toutes mis en évidence un effet sexe. Néanmoins, nos résultats, semblant présager une prédation apparemment un peu plus soutenue par les femelles va dans le sens des travaux réalisés par Bates et al. (2002) et par Strong (2006). La consommation un peu plus importante par les femelles pourrait être attribué à leur taille plus imposante ainsi qu’à leurs besoins nutritifs additionnels associés à leur production d’œufs (Bates et al. 2002a).
La prédation effectuée par L. occidentalis sur les cônes de P. nigra et P. sylvestris montre que la punaise s’accommode particulièrement bien de ces deux essences, pouvant considérablement en diminuer le nombre de graines viables par cône (30%).
Les dégâts causés par L. occidentalis ne varient pas fortement selon les différentes essences de conifères. Cette punaise semble s’accommoder de la même manière à différentes espèces de la famille des Pinacea. Nos résultats confirment la nature polyphage de L.
occidentalis, point fortement documenté dans la littérature et indique que la gamme d’hôte connue de cette punaise s’avère assez vaste. Chez les insectes phytophages, les préférences d’hôtes manifestées, notamment chez les femelles, peuvent être corrélées à la performance des larves par rapport à l’essence choisie, et il pourrait y avoir des bénéfices de fitness à se nourrir et pondre sur des hôtes plus ou moins résistants et appétants (Pureswaran & Poland, 2009). Notre punaise ne constitue pas un insecte manifestant un comportement intime avec sa plante hôte et les espèces de la famille Pinacea correspondent à un groupe phylogénétiquement assez hétérogène, présentant des différences d’ordre biologique, physiologique et biochimique, attestant du caractère très polyphage de cette punaise.
Au vu de la présente étude, plusieurs critiques peuvent être soulevées quant à la représentativité des dégâts observés. Premièrement, dans l’expérience cône, l’effet de prédation de la punaise seule n’a pu être révélé précisément, devant la forte proportion de graines vides dans les cônes à la fois témoins et soumis (> 30 % ; Fig. 8). Les travaux de Schowalter et Sexton (1990) indiquent que cette proportion de graines vides dans les cônes
(particulièrement témoins), est à rapporter à un avortement à cause de facteurs non dépendants des insectes. Mesurer l’impact entier de L. occidentalis sur la production de graines de conifères apparaît donc difficile du fait de la présence de graines naturellement avortées, ne pouvant être discernées de celles vidées par la prise alimentaire de la punaise. A ce titre, en n’ayant pas pris en compte les graines vides comme dégât généré par la punaise, nos résultats pourraient donc avoir sous-estimer l’impact actuel et réel de la punaise sur la productivité des cônes. La présence de graines consommées dans les cônes témoins protégés de P. nigra est inattendue. Ce résultat serait à mettre en relation avec un accès possible du stylet de la punaise à travers un filet de protection trop proche du cône ou à une prédation par
L. occidentalis antérieure à l’expérience en pépinière.
De son côté, si l’expérience graine nous a permis de pouvoir identifier les dégâts réels causés par la punaise sur les graines des différentes essences, l’un des points négatifs que cette expérience soulève, est l’attrait et l’accès non représentatif de la graine pour L.
occidentalis. Il apparaît certain que la prédation d’une graine nue n’est pas la même que celle pour une graine enveloppée par un cône. Ce dernier pourrait par exemple freiner l’accès du stylet de la punaise à la graine, ou au contraire attirer la punaise de part les radiations infra-rouges que ces derniers émettent et qui sont reçues par la punaise (Takács et al. 2009). A ce titre, nos résultats pour les essences P. nigra et P. sylvestris, dans les deux catégories de tests – sur cônes et sur graines – ont révélé des « préférences alimentaires » inverses (même si ces choix ne sont pas statistiquement différents). En outre, les individus mis en test pour l’expérience « graines » constituaient des adultes hivernant, et il est possible que la charge de prédation soit différente selon l’âge et la saison (Strong, 2006).
Enfin, les punaises ayant évolué dans un environnement contrôlé, ce dernier n’est pas forcément représentatif des conditions climatiques et écologiques auxquels les populations sauvages sont réellement soumises. Ainsi en conditions naturelles, les adultes ont davantage de phases de vol et peuvent avoir des besoins énergétiques plus importants qu’en condition d’encagement, résultant en un taux de prédation beaucoup plus vorace dans le premier cas. Les variations dans l’état reproductif des femelles peuvent également rentrer en ligne de compte. Au fil des saisons, les signaux environnementaux prédisant l’entrée en diapause hivernale peuvent enfin entraîner une diminution de prédation par les punaises. Pour ces raisons, nos cinétiques de consommation, peuvent être considérés avec une certaine prudence, dans l’attente d’une comparaison en milieu naturel.
Partie 2 : Etude de la variabilité génétique de différentes populations de L. occidentalis
Notre travail représente la première analyse de la structuration génétique des populations nord-américaines et européennes de L. occidentalis. C’est également la première approche moléculaire sur ce ravageur et qui tente de retracer le processus d’invasion sur le territoire européen depuis sa zone d’origine nord-américaine. Néanmoins, l’échantillonnage obtenu pour cette étude ne permet pas encore de confirmer ou d’infirmer l’origine des populations invasives et le travail présenté constitue une approche préliminaire. Le fragment mitochondrial étudié, bien que de taille relativement faible (417pb conservées après alignement en raison de quelques individus pour lesquels l’amplification avait posé problèmes) s’est révélé être un marqueur suffisamment variable pour donner une première estimation de la diversité et de la structure génétique de L. occidentalis.
Les résultats montrent l’existence d’une variabilité génétique importante dans les zones d’origine et d’introduction, avec un nombre d’haplotypes conséquent pour le nombre d’individus analysés. On ne note pas de structuration dans la distribution des haplotypes présents dans la zone d’introduction. Ce résultat peut être rattaché à la taille relativement faible du fragment analysé ainsi qu’au faible nombre d’individus par population (1-8) et de localités échantillonnées. Dans la zone d’origine néanmoins, sur 8 haplotypes, deux sont communs au deux grandes zone échantillonnées (Californie et Colombie-Britannique), ce qui montre une faible structuration mais également l’existence de flux de gènes entre ces deux régions géographiquement éloignées. Ceci est en accord avec les capacités importantes de dispersion de l’insecte (Koerber, 1963).
En outre, chez la plupart des insectes invasifs, la diversité génétique est réduite dans les zones d’introduction par rapport aux zones d’origine (Puillandre et al. 2008). Notre étude montre ainsi une nette diminution du nombre d’haplotypes, des diversités haplotypiques et nucléotidique entre les zones américaines et les pays européens. Néanmoins, cette diversité génétique reste encore assez élevée dans les zones européennes où la punaise a été introduite par comparaison. L’analyse génétique effectuée sur le scolyte Dendroctonus valens avait montré des résultats semblables, laissant suggérer des introductions multiples (Cognato et al. 2005). Ainsi, cette forte diversité retrouvée dans la zone d’introduction témoigne vraisemblablement de processus multiples dans l’introduction de la punaise, car au contraire des introductions uniques, les populations ne semblent pas avoir subi de forts bottlenecks.
Nous n’avons mis en évidence aucun haplotype commun entre les zones d’origine et d’introduction échantillonnées. Ceci ne nous permet pas de conclure à une invasion
européenne depuis l’Ouest de l’Amérique du Nord. Cette observation peut en outre être due au faible nombre d’individus échantillonnés dans cette zone américaine. Une autre hypothèse également pertinente est la possible invasion en Europe d’insectes transportés depuis l’Est de l’Amérique du Nord, zone envahie quelques décennies avant l’Europe, pouvant être davantage probable compte tenu de l’existence d’échanges transatlantiques à partir des grands ports et aéroports de cette zone.
Il apparaît en définitive qu’un échantillonnage complémentaire dans la totalité des zones (et principalement sur toute l’étendue Nord-Sud de la zone d’origine) ainsi que l’utilisation de marqueurs nucléaires sont nécessaires pour conclure sur ce point. Il serait enfin indispensable de coupler à cette analyse génétique, des études sur les aptitudes de vol et les capacités réelles de dispersion naturelle de cette espèce.
REMERCIEMENTS :
Je remercie le docteur A. ROQUES pour m’avoir accueillie dans son laboratoire. Je le remercie également, ainsi que M.-A. AUGER-ROZENBERG pour m’avoir proposé ce sujet passionnant et de m’avoir conseillée pour la rédaction du rapport. Je tiens particulièrement à exprimer ma reconnaissance à A. YART pour m’avoir conseillée, suivie et aidée tout au long de mon stage. J’adresse également mes remerciements à J.-P. RAIMBAULT pour m’avoir aidée dans la réalisation des RX de graines et la mise en place des expériences et merci à B. COURTIAL de m’avoir accompagnée dans mon premier travail de recherche en biologie moléculaire. Enfin, je tiens à remercier C. BASTIEN pour m’avoir accordé du temps pour l’approche statistique malgré son emploi du temps particulièrement chargé. Enfin, merci aussi à tous les membres de l’URZF pour leur accueil, leur générosité, leur gaieté et leur sympathie qui ont fait de ce stage un moment plaisant et enrichissant.
REFERENCES :
Bates, S.L., Borden J.H., Kermode A.R., Bennett R.G. (2000a) Impact of Leptoglossus
occidentalis (Hem.: Coreidae) on Douglas-fir seed production. J. Econ. Ent. 93 1444-1451.
Bates, S.L., Lait, C.G., Borden, J.H., Kermode, A.R. (2002a) Measuring the impact of
Leptoglossus occidentalis (Heteroptera: Coreidae) on seed production in lodgepole pine using an antibody-based assay. J. Econ. Ent. 95 (4): 770-777.
Bates, S.L., Lait C.G., Borden J.H., Kermode A.R., Savoie A., Blatt S.E., Bennett R.G.
(2000b) Impact of feeding by the western conifer seed bug, Leptoglossus occidentalis (Hemiptera: Coreidae), on the major storage reserves of mature Douglas-fir (Pinaceae) seeds.
Can. Entomol. 132: 91-102.
Bates, S.L., Strong, W.B., Borden, J.H. (2002b) Abortion and seed set in lodgepole and western
white pine conelets following feeding by Leptoglossus occidentalis (Heteroptera: Coreidae).
Environ. Entomol. 31: 1023-1029.
Blatt, S.E., Borden, J. (1996) Distribution and impact of Leptoglossus occidentalis (Hemiptera
Coreidae) in seed orchards in British Columbia. Can. Entomol. 128: 1065-1076.
Brockerhoff, E.G., Liebhold, A.M., Jactel, H. (2006) The ecology of forest insect invasions
and advances in their management. Can. J. For. Res. 36: 263-268.
Ciosi, M., Miller, N., Kim, K., Giordano, R., Estoup, A., Guillemaud, T. (2008) Invasion of
Europe by the western corn rootworm, Diabrotica virgifera: multiple transatlantic introductions with various reductions of genetic diversity. Mol. Ecol. 17: 3614-3627.
Clement, M., Posada, D., Crandall, K.A. (2000) TCS: a computer program to estimate gene
genealogies. Mol. Ecol. 9: 1657-1659.
Cognato, A.I., Sun, J.H., Anducho-Reyest, M.A., Owen, D.R. (2005) Genetic variation and
origin of red turpentine beetle (Dendroctonus valens Le Conte) introduced to the People’s Republic of China. Agricultural and Forest Entomology 7: 87-94.
DeBarr, G., L. (1970) Characteristics and radiographic detection of seed bug damage to slash
pine seed. The Florida Entomologist. 53: 109-117.
Dlugosh, K.M., Parker, I.M. (2008)Founding events in species invasions: genetic variation, adaptative evolution, and the role of multiple introductions. Mol. Ecol. 17: 431-449.
Dusoulier F., Lupoli R., Aberlenc H.P., Streito J.C. (2007) L'invasion orientale de
Leptoglossus occidentalis en France: bilan de son extension biogéographique en 2007.
L'Entomologiste 63: 303-308.
Genton, B.J., Shykoff, J.A., Giraud, T. (2005) High genetic diversity in French invasive
populations of common ragweed, Ambrosia artemisiifolia, as a result of multiple sources of introduction. 14: 4275-4285.
Grapputo, A., Bonan, S., Lindström, L., Lyytinen, A., Mappes, J. (2005) The voyage of an