PAR
MICHEL FOURNIER
THESE SOUMISE A LA FACULTE DES ETUDES GRADUEES ET DE LA RECHERCHE EN VUE DE L'OBTENTION
DU GRADE DE Ph.D.
DEPARTEMENT DE MEDECINE EXPERIMENTALE UNIVERSITE McGILL
SOMMAIRE.
Deux conditions sont essentielles pour le declenchement de la leucemie lymphofde des souris AKR: le virus polytropique et un micro-environnement thymique favorable.
Nous demontrons ici que des injections d'une fraction thymique insoluble (ITF)
a
de·s souris prlHeucemiques acceH!rent le declenchement de la leucemie alors que celles d'une fraction thymique soluble (STF) la retardent. Ceci peut etre explique par la modulation par ITF et STF des ce11ules sensibles au virus ou de la reponse immunitaire. Cette derniere hypothese devient mains attrayante devant 1'inactivite de l'ITF et du STFa
modifier 1'evolution de la leucemie et du melanome 816.D'autre part, l'augmentation avec l'age des cellules sensibles au STF, indicatrice d'une perte graduelle de STF, couplee
a
l'absence de STF chez les souris leucemiques, sont suggestives d'une anomalie de la differenciation des thymocytes resultant en un embouteillage de cellules sensibles au virus. La reconstitution de la leucemie par 1 'ITF chez des souris thymectomisees appuie cette hypothese.ABSTRACT
Spontaneous lymphoid leukemia occurs in AKR mice at about six months. Two conditions must be met for leukemia to be triggered: the polytropic virus and the appropriate thymic microenvironment.
In the present work, we show that in preleukemic mice, injections of insoluble thymic fraction (ITF) accelerated the onset of leukemia whereas injections of soluble thymic fraction (STF) delayed it. This is interpreted as reflecting either the modulation of the virus-sensi-tive cells or the in~une functions of T-cells. The inactivity of ITF and STF on peripheral immune response against leukemic cells or 816 melanoma renders the latter hypothesis less attractive.
On the other hand, the age related loss of STF activity resulting in the gradual increase of virus sensitive cells combined with the reconstitution of leukemia sensitivity in thymectomized mice by ITF suggest a breakdown in the T-cell differentiation process leading to the accumulation of virus sensitive cells and therefore to leukemia.
iv
A Pauline et
a
tous ceux que j 'a·imeTABLE DES MATIERES
TITRE • • • . • • . • • • • • • • • • • . • • • • • • • . • . • . • • . • • • • • . • • • • • • • • • • • • . • • • i
SOMMAI RE . . . • • . • • . . • • . • • • • . • • • • . • • . . • . • • • • . . • . . • • • . . . • • • . . • . • i i
ABSTRACT. . • • • • • • • . • • . • . . . • . • • • • . • . . • . . • • • . • • • • • • • • • • • • . • • • . . i i i
DEDICACE... i V
TABLE DES MATIERES... v
LISTE DES TABLEAUX... ix
LISTE DES FIGURES... X LISTE DES ABREVIATIONS... xi i INTRODUCTION... 1
REVUE BIBLIOGRAPHIQUE... 4
CHAPITRE 1 ROLES POSSIBLES DU THYMUS DANS LE PROCESSUS DE CARCINO-GENESE. . . . • . • . . • . . . • . • . . • • . . . . • • • • • • • . . • • • . • • . • • . • • • . . . 5
CHAPITRE 2 ROLE DU THYMUS DANS LA DIFFERENCIATION DES LYMPHOCYTES T 11 2.0 Introduction . . . • . . . 11 2.1 Structure du thymus. . . • . . . • • • • . . • . . . • . • . 11 2.1.1 Histologie... 11 2 . 1 . 1 . 1 Cortex . . . 12 2.1.1.2 Jonction cortico-medullaire... 15 2.1.1.3 Medulla... 16 2.1.2 Vascularisation... 18
2.2 Differenciation des lymphocytes thymiques... 19
2.3 Role du thymus dans la diff~renciation des lympho-cytes T. . . 25
2.3.1 Introduction... 25
2.3.3 Microenvironnemer.t thymique ...•.•..•... 2.3.4 STF et ITF ... . 2.4 Conclusion ...•.•...••.•..••...•... CHAPITRE 3 30 34 37
LES SOURIS AKR... 41
3.0 Introduction... 41
3. 1 Oeve 1 oppement de 1 a souche AKR ...•.•.... ·. . . . • • . . . . 42
3.2 La leucemie AKR... 43
3.3 Etat immunologique des souris AKR... 44
3 . 4 Vi ru s de Gross . . . .. . . . 4 7 3.5 Periode de latence... 49
3 • 6 Con c 1 us ion . . . • • . . . • . . . 52
CHAPITRE 4 THYMUS ET LEUCEMIE AKR... 54
4.0 Introduction ... . 54
4.1 Effet des thymectomies et des greffes de thymus sur la leuc~mie ... . 54
4.2 Roles possibles du thymus dans le processus de leuc~mogenese.. . . 56
4.3 Thymus et facteurs thymiques chez AKR... 61
4.4 Conclusion... 68
MATERIEL ET METHODES... 70
Animaux... 71
Souri s . . . .. . . . 71
Lapins... 72
Preparation des fractions thymiques soluble (STF) et insoluble (ITF) ... ... -.... 72
Surnageant de culture de cellules epitheliales thymi-ques . . . .. . . • . . . . • . . . .. . . . 73
Serun1 anti-STF. . . . • . . . .. . . . .. . . . .. . . . .. . . . 7 4 Serum anti-ITF... 75
Technique d'imrnunofluorescence indirecte... 76
Thy~ ctomi e . . . 7 7 Test de cytotoxicite differentielle... 77
Induction de la reaction GvH... 79
Mel a name Bl6.. . . 80
Determination des criteres de leucemie... 81
Technique de palpation d'organe... 81
Mort par leucemie... 82
Statistiques... 82
TRAVAIL EXPERIMENTAL... 84
1. Introduction... 85
2. Effet de l'ITF et du STF sur le declenchement de la leucemie des souris AKR... 85
3. Action peripherique du STF et de l'ITF... 90
3.1 Introduction... 90
3.2 Effet du STF et de 1 'ITF sur l'evolution de la leucemie... 91
3.3 Transfert de cellules leucemiques
a
des rece-veurs pretraitesa
1 'ITF et au STF... 943.4 Transfert de cellules de souris pretraitees au STF
a
des souris preleucemiques... 983.5 Effet des fractions thymiques sur 1 'evolution de la tumeur 816... .• . .. .. 103
3.5 .1 Introduction... 103
3.5.2 Effet des fractions thymiques sur l'evolution de la tumeur 816, cas d • une DL 1 00 . . . 1 04 3.5.3 Effet des fractions thymiques sur l'evolution de la tumeur 816, cas d'une DL 50... 109 3.5.4 Effet des fractions thymiques sur
l'e-volution de la tumeur B16; cas des souris thymectomis~es ...•...•...
3 • 6 Con c 1 us i on . . . • • . . . ...•...
4. Action centrale du STF et de l'ITF ...••...•.... 4.1 Etat du STF et de 1 'ITF endogene ... . 4.2 Etat fonctionnel du STF dans la periode
pre-leucemique ... . 4.2.1 Mode le GvH •...•..••.••..•.••••.. 4.2.2 Marqueurs Thy-1 .•..•..•••.•••••••••.... 113 117 118 118 119 120 124
e·
4.3 Reconstitution de l'activite leucemogenique
chez des souris AKR thymectomisees... 129
5. Resume des resultats... 133
DISCUSSION GENERALE... 135
I. Effet de l'ITF et du STF sur la leucemie des souris AKR. . . • . . . . • • • . • . • . . • • . • • • . . • • . . • . • . . • • • . . . • . • • . 1 36 II. HypothAse de l1action peripherique del 'ITF et du STF... . . 145
Ill. Hypothese de l'action centrale de l'ITF et du STF.. 150
IV. Modele propose pour expliquer le processus de leuce-mogenese . .. ·. . . • . • . . . • . . . .. . . . • . . . 155
V. Importance de ces recherches pour 1 'etude de la leucemie humaine... 158
CONCLUSIONS GENERALES. . . . • • . . . 160
REMERCIEMENTS... 162
COMMUNICATIONS ET PUBLICATIONS... 164 BIB L I OGRAPH I E . • . . . . • . . • • . . . • . . • • . . . • . . • . . • . . . • . • . . . • . . • . . . 16 8 ANNEXE I CONDENSED ENGLISH VERSION 11ROLE OF THYMIC ~1ICRO
ENVIRONMENT IN THE LELIKEMOGENESIS OF AKR MICE" .. ANNEXE II EFFECT OF DIFFERENT COMPONENTS OF THE THYMIC
STROMA ON THE ONSET OF AKR LEUKEMIA ... . ANNEXE Ill: ROLE OF TWO THYMIC MICROENVIRONMENTAL FRACTIONS
IN THE MODULATION OF THE GRAFT-VERSUS-HOST
REACT I ON . ..••••..••..••.•••••••••••••••••••••..• ANNEXE IV AGE-DEPENDENT ACCUMULATION OF THYMIC HORMONE
SENSITIVE CELLS ...•.. ANNEXE V QUANTITATIVE CHANGES IN AKR THYMOCYTE SUBSETS
PRECEDING THE ONSET OF LEUKEMIA ..••...• ANNEXE VI TRIGGERING OF LEUKEMIA IN THYMECTOMIZED AKR MICE
BY AN INSOLUBLE THYMIC FRACTION ....••••...•.. ANNEXE VII: T CELL MARKERS AND RECEPTORS: POTENTIAL USE IN
IMt4UNE MODULATION ... .
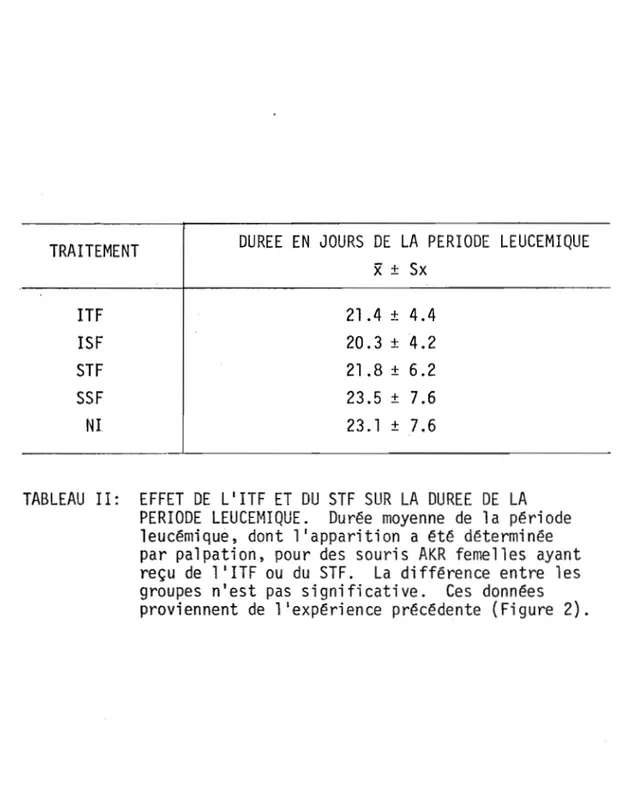
TABLEAU I TABLEAU DES DIFFERENTES PREPARATIONS 28 DE FACTEURS THYMIQUES.
TABLEAU II EFFET DE L1
ITF ET DU STF SUR LA DUREE 93 DE LA PERIODE LEUCEMIQUE
TABLEAU
III
TRANSFERT DE CELLULE$ LEUCEMIQUES DE 95 THYMUSTABLEAU IV TRANSFERT DE CELLULES LEUCEMIQUES DE 96
-
RATETABLEAU V TRANSFERT DE CELLULES LEUCEMIQUES DE 97
X
LISTE DES FIGURES
FIGURE 1 SCHEMA DE LA DIFFERENCIATION DES LYMPHOCYTES T 38 FIGURE 2 EFFET DE L'INJECTION D'ITF ET DE STF SUR LA 87
SURVIE DE SOURIS AKR FEMELLES.
FIGURE 3 EFFET DE L'INJECTION D'ITF SUR LA SURVIE DE 89 SOURIS AKR MALES.
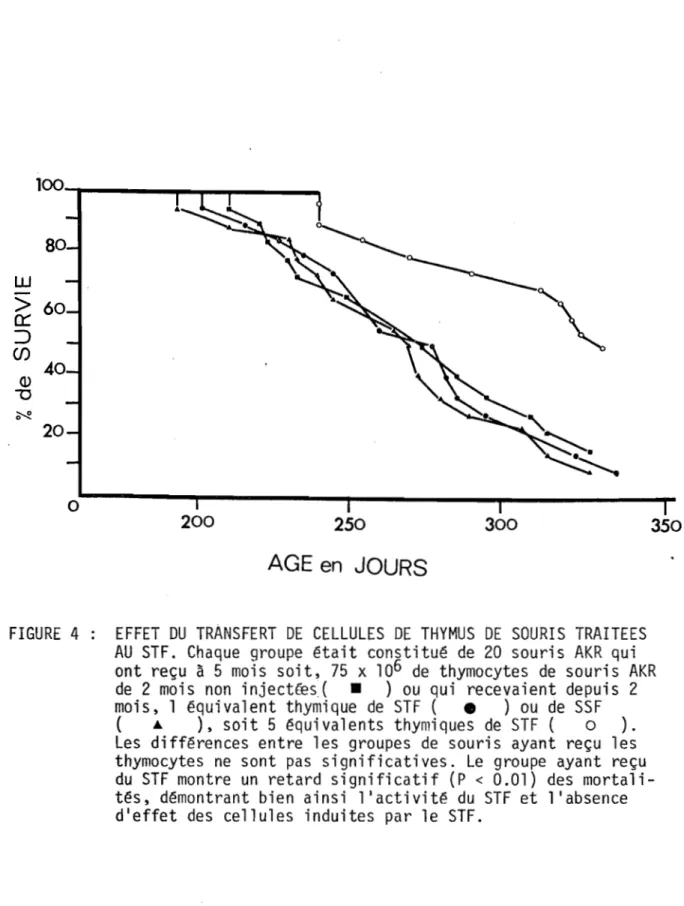
FIGURE 4 EFFET DU TRANSFERT DE CELLULE$ DE THYMUS DE 100 SOUR IS TRAITEES AU STF.
FIGURE 5 EFFET DU TRANSFERT DE CELLULE$ DE RATE DE 101 SOURIS TRAITEES AU STF.
FIGURE 6 EFFET DU TRANSFERT DE CELLULE$ DE GANGLIONS 102 DE SOURIS TRAITEES AU STF.
FIGURE 7 EFFET DES FRACTIONS THYMIQUES SUR LE POURCEN- 106 TAGE DE PRISE DE LA TUMEUR Bl6, CAS D'UNE DL 100. FIGURE 8 EFFET DES FRACTIONS THYMIQUES SUR LA MORTALITE 107
PAR LA TUMEUR B16, CAS D'UNE DL 100.
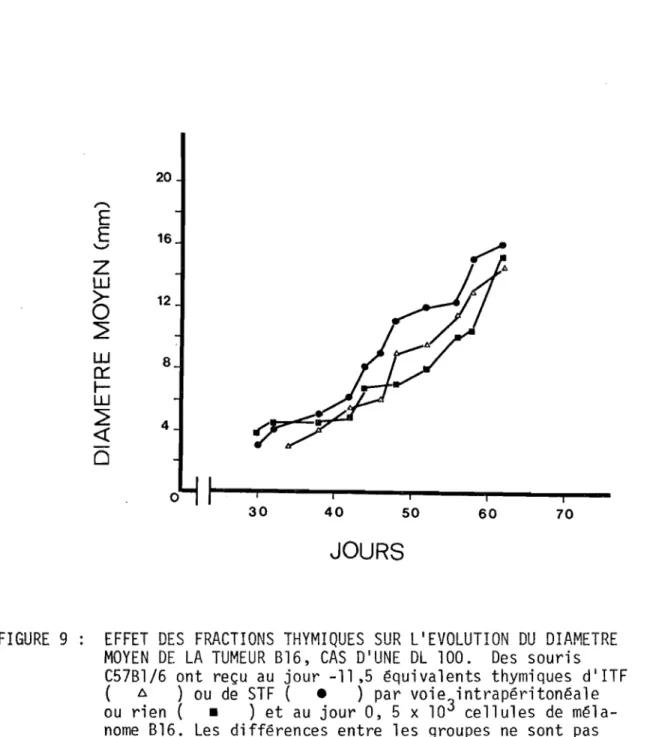
FIGURE 9 EFFET DES FRACTIONS THYMIQUES SUR L'EVOLUTION 108 DU DIAMETRE MOYEN DE LA TUMEUR Bl6, CAS D'UNE
DL 100.
FIGURE 10 EFFET DES FRACTIONS THYMIQUES SUR LE POURCENTAGE 110 DE PRISE DE LA TUMEUR 816, CAS D'UNE DL 50.
FIGURE 11 EFFET DES FRACTIONS THYMIQUES SUR LA MORTALITE 111 PAR LA TUMEUR B16, CAS D'UNE DL 50.
FIGURE 12 EFFET DES FRACTIONS THYMIQUES SUR l'EVOLUTION 112 DU DIAMETRE MOYEN DE LA TUMEUR 816, CAS D'UNE
DL 50.
FIGURE 13 EFFET DES FRACTIONS THYMIQUES SUR LE POURCENTAGE 114 DE PRISE DE LA TUMEUR 816, CAS DES SOURIS
THY-MECTOMISEES.
FIGURE 14 EFFET DES FRACTIONS THYMIQUES SUR LA MORTALITE 115 PAR LA TUMEUR 816, CAS DES SOURIS THYMECTOMISEES. FIGURE 15 EFFET DES FRACTIONS THYMIQUES SUR L'EVOLUTION DU 116
DIAMETRE MOYEN DE LA TUMEUR 816, CAS DES SOURIS THYMECTOMISEES.
FIGURE 16 AUGMENTATION DU POTENTfEL INDUCTEUR DE GvH DE 122 CELLULE$ DE SOURIS AKR ET BALB/c;
FIGURE 17 AUGMENTATION AVEC L'AGE DES CELLULES A F-Thy 1 126 DANS LE THYMUS DE SOURIS AKR ET BAL8/c.
FIGURE 18 AUGMENTATION AVEC L'AGE DES CELLULES A F-Thy 1 128 DANS LA RATE DE SOURIS AKR ET 8ALB/c.
FIGURE 19 EFFET DE L'ITF ET DU STF SUR LA SURVIE DE SOURIS 131 AKR THYMECTOMISEES.
Con A CRE DL 50 DL lOO f-Thy-1 F-Thy-1 GvH ISF
ITF
LSF PBS PHA PM STF SSF TES Thy-1 N.B.:LISTE DES ABREVIATIONS
Concanavaline A
Cellule reticuloepitheliale Dose letale 50
Dose letale lOO
xii
Cellules
a
faible concentration en antig~ne Thy-1 (Low-Thy 1) Cellulesa
forte concentration en antig~ne Thy-1 {High-Thy 1) Reaction du greffon contre l 1h6te (Graft-versus-Host) Fraction insoluble de rate (Insoluble Spleen Fraction) Fraction insoluble de thymus (Insoluble Thymic Fraction) Hormone thymique du groupe de Metcalf (Lymphocytosis Stimulating Factor)
Tampon phosphate salin (Phosphate Buffer Saline) Phytohemagglutinine
Poids moleculaire
Fraction soluble de thymus {Soluble Thymic Fraction) Fraction soluble de rate (Soluble Spleen Fraction) Surna9eant de culture de cellules epitheliales thymi-ques {Thymic Epithelial Supernatant)
Antig~ne theta, alloantig~ne des lymphocytes T de la souris
Nous avons prefere conserver les abreviations anglaises car elles sont mieux connues et ce sont celles retrouvees dans
INTRODUCTION
Parmi les approches envisagees dans la lutte contre le cancer, l'etude du processus et des circonstances entratnant la transforma-tion des cellules normales en cellules neoplasiques est certainement une voie non-negligeable. En effet, la connaissance de ces pheno-menes nous permettrait d'interferer avec 1 'apparition de la neoplasie ou du mains d'intervenir dans ses stades precoces. Un des types de cancer pour lequel de telles etudes pourraient etre importantes est le cas des leucemies lymphofdes, A cause des pauvres succes obtenus en therapie. En effet~ dans ces leucemies, les cellules neoplasiques apparaissent en premier dans le thymus et envahissent rapidement, par la circulation, les autres organes, ce qui rend les interventions therapeutiques complexes. Dans le present travail, nous examinerons
2.
le role du thymus dans le processus de leucemogenese afin de permettre l'in-tervention· .. dans les leucemies, directement au niveau du thymus,
c'est-a-dire, avant l'expansion extra-thymique de la neoplasie.
Pour ce faire, nous reverrons brievement les roles possibles du thymus dans le developpement des cancers. Par la suite, nous examinerons la structure du thymus en limitant toutefois notre description aux
elements pertinents a la comprehension du role du thymus et de ses composantes (hormone et microenvironnement) dans la differenciation des lymphocytes T. Nous etudierons ensuite la pertinence du choix de la leucemie des souris
AKR
comme modele de la leucemie humaine et nous-
(d~gagerons les indications. exp~frime·ntales~qui permettertt de reiier.le processus de leucP.mog~n~se A une anomalie de la diff~renciation des lymphocytes T.
c•est ce dernier point qui sera explor~ par les diverses exp~
riences d~crites dans cette th~se. Ces derni~res nous permettront de determiner le role de deux elements du thymus dans la leuc~mog~n~se
CHAPITRE 1
ROLES POSSIBLES DU THYMUS DANS LE PROCESSUS DE CARCINOGENESE
Ce chapitre ne se veut pas une etude exhaustive de tous les types de cancer dans lesquels le thymus semble implique mais plutot une analyse globale des roles possibles du thymus dans le cancer.
Chez la souris et le rat, un des types de neoplasie dans lequel l'importance du thymus peut etre plus facilement mis en evidence est le cas des leucemies lymphofdes. En effet, dans la plupart des souches de souris, c'est dans le thymus qu'apparaissent les premiers lympho-cytes leucemiques et c'est aussi 1 'organe le plus affecte par la mala-die. C'est notamment le cas des souches AKR et C58 qui montrent une haute incidence de leucemies spontanees (Furth et Boon, 1945). Mais c'est aussi le cas chez d'autres souches de souris dans lesquelles la faible occurence peut etre augmentee par l'emploi de radiations {Kaplan, 1947), de carcinogenes chimiques tels que le methylcholanthrene ou
l'urethane (Law et Miller, 1950), d'oestrogenes (Gardner et coll ., 1944) ou par surrenalectomie ou gonadectomie (Law, 1947).
De tous ces cas nous pouvons degager un point commun qui fait vraiment ressortir l'importance du thymus dans le processus de la
0
{McEndy et call., 1944; Furth et Boon, 1945; Law et Miller, 1950; Kaplan, 1950, 1954; Metcalf, 1962) et cela peu importe la souche de souris ou la nature de l'agent oncogene. De plus, chez ces animaux nous notons un retablissement de leur susceptibilite
a
la leucemie lymphofde qu'elle soit spontanee {Law et Miller, 1950; Miller, 1960a, b) ou induite par des virus {Levinthal et call., 1959; Miller, l960a, b; tevinthal et Buffet, 1961), par des carcinog~nes (Law et Miller, 1950} ou par radiations {Kaplan et coll., 1953; Kaplan, 1961), lors-qu'un thymus leur est greffe.Dans d'autres types de cancer tels que les fibrosarcomes induits
~
par des carcinogenes chimiques (Prehn, 1964) ou des tumeurs mammaires (Martinez, 1964) nous notons aussi une reduction de l'incidence suitea
la thymectomie bien que ce ne soit pas dans le thymus que les pre-mieres cellules neoplasiques apparaissent.Inversement, dans beaucoup de cancer, l'ablation du thymus s'ac-compagne d'une augmentation de l'incidence des neoplasies. C'est le cas notamment des polyomes ~ virus (Ting et Law, 1965) des tumeurs induites soit par des virus (Kirschstein et coll., 1964), soit par des agents chimiques (Miller et coll., 1963; Grant et Miller, 1965) ou physiques (Defendi et Roosa, 1964).
Parmi les diverses possibilit~sinvoqu~es pour expliquer le role du thymus dans le d~veloppement de ces types de cancer, trois hypoth~ses
retiennent parti cul ierement 1' attention. La thymus pourrait influencer
le processus n~oplasique:
1) en affectant la r~ponse immunitaire de 1 'hote, particuliere-ment au moparticuliere-ment de l'~mergence de la tumeur;
2) par la production de facteurs ayant une action trophique sur les cellules tumorales;
3) en fournissant un environnement propice pour le d~clenchement
et l'expansion de la tumeur.
1) la premiere hypothese, celle d'une influence thymique sur la r~
ponse immunitaire dirig~e contre les cellules tumorales, implique que ces dernieres doivent posseder des antigenes qui les signa-lent au systeme immunitaire. Sur la base de 1 'antig~nicite des cellules tumorales, nous pouvons retrouver deux types d'~volution
tumorale:
a) les cellules tumorales ~mergentes sont d~ficientes en antigenes n~oplasiques, c'est
a
dire en antigenes sp~cifiques aux cellules tumorales, la tumeur pourra alors se
d~velopper
a
l'abri du systeme immunitaire;b) les cellules tumorales presentent des antigenes neoplasi-ques, la croissance de la tumeur pourra alors etre
0
Comme dans la majorite des mod~les animaux dans lesquels les neoplasies sont induites par des virus ou par des agents chi-miques et physiques, les cellules tumorales presentent une for-te concentration d'antigenes n~oplasiques, nous concentrerons principalement notre attention sur le deuxieme type d'evolution tumorale (Prehn et Main, 1957; Klein et coll., 1960; Old et coll., 1962; Slettenmark et Klein, 1962; Klein et coll ., 1962; Globerson et Feldman, 1964). Considerant d'une part que la thy-mectomie neonatale ou adulte s'accompagne generalement d'une baisse de la reactivite du systeme immunitaire (Miller et coll., 1964; ling et Law, 1965; Miller, 1965) et d'autre part que cette
meme
thymectomie est suivie dans beaucoup de modeles animaux d'une augmentation de la susceptibilitea
plusieurs types de cancer (Metcalf, 1966),a
la lumiere de la forte antigenicite des cellules tumorales dans ces mod~les, nous pensons facilement relier la hausse de la susceptibilite aux neoplasiesa
la baisse de la reponse immunitaire chez ces animaux thymectomises.2) Par contre, dans les cas ou la thymectomie se traduit par une baisse de la susceptibilite aux neoplasies, 1 'effet des thymec-tomies sur le developpement des neoplasies pourrait etre expli-que par l'enlevement de facteurs d'origine thymiexpli-que ayant une action trophique sur les cellules tumorales. Ce concept d'une influence trophique du thymus remonte aux premiers travaux sur
le thymus on Rowntree et collaborateurs (1935) ont rapport~ que des injections r~p~t~es d'extraits thymiques provoquaient des hyperplasies thymiques. Au meilleur de nos connaissances, ces travaux n'ont jamais ~t~ confirmes. En effet, aucun auteur n'a signale d'induction de neoplasies du thymus subsequentes
a
des injections d'extraits thymiques oua
des greffes multiples de thymus. Certains auteurs ont signale une augmentation de la taille du thymus suitea
des injections d'extraits thymiques(Zollinger et Potworowski, 1977; Zollinger et coll., 1977), mais cette hausse du volume thymique n'etait aucunement reliee
a
l'ap-parition d'une tumeur.Szent-Gyorgyi et collaborateurs (1962} ont sugg~re que le thymus pourrait contenir deux facteurs: un facteur inhibiteur (-retine) et un facteur promoteur (promine) ayant une activite sur la croissance de cellules tumorales carces ·deux··extraits ·solubles de thymus avaient des effets oppos~s sur la croissance tumorale.
Toutefois, ces conclusions demandent encore
a
@tre confirmees.Done, en absence de support exp~rimental, ce concept d'une action trophique du thymus sur les cellules tumorales prend une allure purement sp~culative jusqu'a ce que de meilleures preu-ves soient produites.
0
3) Le fait que le thymus fournisse un environnement ideal pour le declenchement et 1 'expansion des tumeurs pourrait expliquer l'effet antitumoral de la thymectomie. Toutefois cette possi-bilite ne pourrait s'appliquer qu'aux cas on la tumeur debute dans le thymus comme le cas de la plupart des leucemies lymphof-des. Comme cette possibilite sera discutee en detail plus loin . {p. 54 ) , nous ne l'aborderons pas dans ce chapitre.
Done, en resume, le thymus est implique dans la determination de la susceptibilite
a
plusieurs types de cancer, tel que demontre par les experiences de thymectomie et de greffes thymiques. L'augmentation de l'incidence de tumeurs subsequentea
la thymectomie pourrait etre relieea
l'effet immunosuppresseur de ce traitement. Pour ce qui en est de l'effet antitumoral de la thymectomie, les travaux sont nette-ment mains convainquants. Dans cette these, nous tenterons de deter-miner quel est le ou les elements du thymus qui pourraient etre respon-sables de l'induction de tumeurs en concentrant nos recherchesa
l'etu-de l'etu-de la leucemie lymphoblastique aig6e l'etu-de la souris. Mais avant d'en-trer plusa
fond dans ce domaine, nous allons revoir 1 'etat des con-naissances sur la structure et la fonction du thymus.CHAPITRE 2
ROLE DU THYMUS DANS LA DIFFERENCIATION DES LVMPHOCYTES T
2.0 INTRODUCTION
Du chapitre precedent, il semble que le thymus soit implique dans bon nombre de cancers soit en permettant la differenciation des cellu-les immunologiquement competentes contre cellu-les cellucellu-les cancereuses, soit en tant que site de la transformation oncogene. Il devient done essen-tiel de faire un bref rappel de la structure du thymus et de son role dans la differenciation des lymphocytes T. Nous ne ferons toutefois pas une etude exhaustive de l'histologie thymique, nous limiterons notre examen aux elements qui seront necessaires
a
la comprehension desme-canismes de differenciation des lymphocytes T et de la leucemogenese des souris AKR
2. 1 STRUCTURE DU THYf~US
2.1.1 Histologie
Toutes les experiences decrites dans cette these ayant ete effectuees chez la souris, nous examinerons essentiellement la structure du thymus de souris. Cet organe lymphoepithelial, situe derriere le manubrium sternal, est compose de deux lobes distincts. Chaque lobe est entoure d'une enveloppe de tissu conjonctif, la
capsule. Cette capsule se prolonge sous forme de travees inter-lobulaires, vers l'interieur del 'organe, delimitant ainsi les lobules. Le nombre et l'agencement des cellules d'un lobe thymi-que permettent de distinguer une zone externe plus basophile, le cortex, d'une zone interne plus eosinophile, la medulla.
C'est en examinant les differents types de cellules pre-sents dans un lobule thj~ique que nous pouvons mieux saisir la distinction entre le cortex et la medulla.
2.1 . 1 • 1 Cortex
~ Le cortex contient, immediatement sous le tissu
conjonctif capsulaire, une couche de cellules, dites sous-capsulaires, qui entourent le lobule. Certaines de ces cellules ressemblent
a
des fibroblastes, et d'autres, aux cellules reticulees du cortex.Nous trouvons
a
la peripherie du cortex, sous la couche de cellules capsulaires, une zone dite sous-capsulaire de cellules lymphoblastiques. Ces cellules, ayant les caracteristiques de gros lymphocytes, ont un cycle cellulaire rapide (Metcalf et Wiadrowski, 1966; Clark, 1968;Fabrikant et Foster, 1969; Michalke et call., 1969). Ces observations, ainsi que le contact etroitentre ces cellules et les cellules reticulees, ont amene certains auteurs ~ attribuer un role mitogenique aux cellu-les reticulees (Hoshino~ 1963; Brumby et Metcalf, 1967; Mandel, 1969, 1970). 11 semblerait toutefois que l'on ne puisse relier cette activite mitogenique
a
une quelconque stimulation antigenique ~ cause de la structure particulie-re des vaisseaux sanguins dans cette particulie-region qui empechent toute entree d'antigenes dans le thymus (Raviola et Karnovsky, 1972). Sous cette couche de cellules blastiques, c'est-a-dire dans le cortex profond, nous trouvons deux types pre-dominants de cellules: les cellules reticulees et les lym-phocytes.Les cellules reticulees ou reticuloepitheliales, presentent, lorsque co1orees
a
1 'hematoxyline/eosine, un gros noyau ovale pale ainsi qu'un reseau de chromatine lache. Leur abondant cytoplasme n'est pas mis en evidence avecles colorants usuels (Good, 1964; Sainte-Marie et Leblond, l964a, 1965). Ces cellules se relient entre elles par de nombreux prolongements cytoplasmiques ~ la jonction des-quels nous trouvons des desmosomes. De nombreux tonofi1a-ments traversent ces cellules au niveau des desmosomes
(Clark, 1963, 1973). C'est
a
l'interieur des mailles de ce reticulum, qui forme un treillis cellulairea
travers tout0
le cortex, que se trouvent les lymphocytes.
Les lymphocytes du thymus montrent une assez grande h~t~rog~neite de tailles. D'apr~s leur diam~tre,
les auteurs les classifient en trois groupes: les grands, les moyens et les petits lymphocytes. Les grands lympho-cytes ont un diametre sur frottis variant entre 18 et 23 ~.
Ils ont un noyau pale contenant 2 ou 3 nucleoles fonces et un cytoplasme bleu pale
a
l'hematoxyline/eosine. Les petits lymphocytes dont la taille varie entre 7 et 13 ~' sur frottis, ont un noyau dense et tres fonce contenant 1 ou 2 petits nucleoles. Leur cytoplasme bleu pale est extremement r~duit et difficilea
voir. Entre ces deux formes extremes, nous pouvons distinguer environ 6 formes intermediaires avec des cellules variant entre 13 et 18 ~de di ametre.
Les lymphocytes sont distribues de fa~on regulie-re; les gros lymphocytes sont en p~ripherie et nous notons une diminution de la taille des lymphocytes plus on se rap-proche de la medulla (Sainte-Marie et Leblond, 1964a, 1964b, 1965). Le nombre de mitoses des lymphocytes corticaux est plus eleve en peripherie qu'au centre (Dustin, 1913, 1920; Sainte-Marie, l964a). C'est grace
a
cette doubletion qu'a ete etabli ce premier modele de differenciation des lymphocytes thymiques discute en detail plus bas (section 2.2).
2.1.1.2 'Jonction cortico-medullaire
A la jonction cortico-medullaire, nous trouvons des cellules reticulees. Le cytoplasme de ces cellules est plein de granulations de diverses tailles qui, lorsqu'exa-minees au microscope
a
fluorescence, emettent spontanementune lumiere jaune. Ces cellules autofluorescentes sont aussi nommees "lipid-laden foamy cells" (Bergmann, 1943) car leurs granulations se colorent en noir au Soudan (colorant des corps gras) ou encore "PAS positive reticulum cell11
(Sainte-Marie et Leblond, 1964b) car elles sont positives
a
la co-loration APS. L'ultrastructure de ces cellules est sembla-blea
celle des cellules reticulees cystiques, decrites en details plus bas (Mandel, 1968, 1968a). Nous n'attribuons pas de fonction precise aux cellules autofluorescentes. C'est egalement dans cette region ·que se trouve la majorite des macrophages thymiques (Hoefsmithet Gerver, 1975;0
2 .1.1.3 Mt!!dull a
La medulla presente une situation inverse au cortex en ce qui a trait aux proportions des difft!!rentes populations cellulaires; par unite de surface, elle con-tient beaucoup plus de cellules reticulees et beaucoup moins de lymphocytes que le cortex.
Les cellules reticulees de la medulla ont des structures variees (Hammar, 1921; Downey, 1948). Mandel les a classifiees en quatre groupes d'apres leurs carac-teristiques ultrastructurales: les non-differenciees, les squarneus,es, les villeuses et les cystiques (Mandel, 1968).
Les cellules reticulees non-differenciees, pre-sentes dans toute la medulla, ont un noyau ovale et inden-te, une quantite de cytoplasme variable, de nombreux
poly-ribosomes et un reticulum endoplasmique granu1aire ou
agranul ai re.
Les cellules reticulees cystiques ont un reticu-lum endoplasmique principalement agranulaire, de nombreux ribosomes libres, un appareil de Golgi peu developpe et de petites mitochondries. Ces cellules tirent leur nom de la presence en grand nombre de vacuoles ("cysts111
) dans
le cytoplasme. Les auteurs sont generalement d'accord pour attribuer
a
ces cellules la fonction de secretion de l'hormone thymique (Mandel, 1963; Clark, 1963; Hoshino,1963; Clark, 1968; Mandi et Glant, 1973; Teodorczyk et coll., 1975).
Les deux autres types de cellules reticulees se retrouvent dans les corpuscules de Hassall. Ces structu-res caracteristiques du thymus sont des amas concentriques de cellules reticulees squameuses (Clark, 1973). A la pe-ripherie des corpuscules de Hassall, nous trouvons des cellules reticulees villeuses. Ces cellules caracterisees par des prolongements cytoplasmiques en forme de villosite, sont reliees entre elles et entourent le corpuscule.
Les lymphocytes de la medulla sont surtout de
pe-tits et moyens lymphocytes. Ils possedent un noyau de
forme irreguliere. Sainte-Marie et Leblond attribuent ce phenomene
a
la mobilite de ces cellules dans la medulla(Sainte-Marie et Leblond, 1964a, 1964b). Nous crayons qu'une partie des cellules quitterait l'organe par diapedese
a
travers l'endothelium des veinules post-capillaires pour aller peupler les organes peripheriques (Clark, 1963; Sainte-Marie et Leblond, 1964a, 1964b, 1965; Goldstein et call.,1968).
2.1.2 Vascularisation
L'apport sanguin du thymus est assure par une artere penetrant chaque lobe par son hile (Smith et coll ., 1952). Celle-ci se ramifie et ses branches atteignent tous les lobules thymi-ques en cheminant soit
a
travers la medulla, soit dans les travees ·interlobulaires. A partir de ces branches, dans le tissu conjonc-tif interlobulaire et capsulaire, prennent naissance de fines arterioles qui deviennent des capillaires penetrant dans les lo-bules perpendiculairement au cortex pour rejoindre la medulla. A la jonction corticomedullaire, ces capillaires se transforment en veinules post-capillaires qui s•unissent pour traverser la medulla et les travees interlobulaires et devenir la veine hilai-re qui quitte l'organe (Smith et coll., 1952; Defendi, 1964; Ham, 1965; Raviola et Karnovsky, 1972).Les veinules post-capillaires du thymus sont caracterisees par leurs cellules endotheliales et la presence de gaines peri-vasculaires. Les cellules endotheliales, qui bordent ces vaisseaux, sont legerement plus grosses que celles des autres vaisseaux et leur noyau bombe vers la lumiere. Ces cellules paraissent aussi plus nombreuses par unite de surface comparativement
a
d'autres veinules. Dans la medulla, des gaines, generalement au nombre dedeux ou trois, ceinturent les veinules post-capillaires.
L'anti-g~nicit~ specifique de ces doubles membranes (Potworowski, 1977)
coupl~e
a
leur absence chez les rudiments thymiques de la souris nude {Raviola et Karnovsky, 1972) pourrait indiquer leur importan-ce dans les tous premiers stages de la diff~renciation des lym-phocytes thymiques. A l'int~rieur des espaces dits "p~rivasculaires", c'est-a-dire, entre la paroi interne de la gaine et la paroi externe de la veinule, se trouve une couche de petits lym-phocytes de forme irr~guliere laissant supposer une activit~ de
diap~dese et de migration de ces lymphocytes. Dans les canaux ex-ternes de ce complexe, des granulocytes, des mastocytes et des plasmocytes sont parfois presents (Sainte-Marie et Leblond, 1958, 1962, l964a, 1964b, 1965).
2.2 DIFFERENCIATION DES LYMPHOCYTES THYMIQUES
11 est maintenant generalement accept~ que les lymphocytes T se differentient dans le thymus (Bastenie, 1932; Kindred, 1940; Andreasen et Ottesen, 1945; Bierring, 1960; Schooley et Kelly, 1961). Par contre, l'origine de leurs precurseurs, leur mode de differenciation en lympho-cytes et la destinee des lympholympho-cytes thymiques sont plus discut~s.
Examinons d'abord l 'origine embryonnaire et adulte des precurseurs.
L'origine embryonnaire des lymphocytes thymiques est encore dis-cutee. Un premier groupe d'auteurs considere que les lymphocytes
proviennent de la differenciation de cellules reticu1oepithe1ia1es de l'ebauche thymique (Prenant, 1894; Dustin, 1913~ de Winiwarter, 1924; Dustin et Gregoire, 193la, 193lb; Choi, 1931; Bastenie, 1932; Kindred, 1940), alors que d'autres attribuent ce role de precurseurs aux cellu-les mesenchymateuses (Hammar, 1905; Maximow, 1909; Maximow et Bloom, 1935; Bloom, 1938; Norris, 1938}. Ces deux hypotheses reposent sur l'examen de preparations histologiques et l'observation de cellules ayant des caracteristiques intermediaires entre le presume precurseur
(reticule ou mesenchymateux) et les lymphocytes.
Par 1 'etude in vitro de la differenciation des 1ymphocytes T, on
~ a essaye de concilier les deux hypotheses. Des resultats obtenus par cultures de cellules mesenchymateuses et epitheliales, "pures11 et melan-gees, suggerent que les lymphocytes peuvent provenir de 1•un ou l'autre de ces types de cellules (Auerbach, 1960; Ball et Auerbach, 1960;
Clark, 1963; Auerbach, 1964a, 1964b; Ackerman et Knouff, 1965). Toute-fois cette conclusion peut etre mise en doute de par la purete reelle de 1eur preparation ce11ulaire.
Des elements plus concrets de reponse ant ete apportes par les experiences de Le Douarin (1975}, realisees chez le poulet ayant rec;u des cellules souches de cailles. Ces experiences ant pu demontrer le peuplement des thymus de poulets par les cellules mesenchymateuses de cailles. De plus, les experiences de Pykeet Papiernik {1979) et
ce11es de Jotereau et collaborateurs (1980}, qui dans un systeme de migration in vitro ont demontre lBxistence d'une substance produite par les cellules epitheliales attirant les cellules mesenchymateuses, appuient tres fortement les resultats de Le Douarin.
Chez la souris adulte, 1'origine des precurseurs des lymphocytes, c'est-a-dire les cellules blastiques de la zone sous-capsulaire, est moins controversee; des experiences utilisant des marqueurs chromoso-miques chez des animaux en parabiose (Harris et coll., 1964) ou encore des transferts de cellules antigeniquement identifiables (Gengozian et call., 1957; Ford et co11., 1966; Order et Waksman, 1969; Takada et Takada, 1973; Hiesche et Revesz, 1974; Kadish et Basch, 1976; Four-nier et call., 1979) indiquent que la moelle osseuse en est probable-ment la source principale.
La question de la voie d'entree des precurseurs dans le thymus est encore entiere; plusieurs possibilites ont ete envisagees pour l'elucider. Les precurseurs pourraient entrer soit par le tissu con-jonctif capsulaire et interlobulaire (Everett et Tyler, 1969; Sainte-Marie, 1971), soit par diapedese
a
travers 1es capil1aires de la re-gion corticale sous-capsulaire (Clark, 1973) ou encore quitter la circulation au niveau des vaisseaux medul1aires (Brumby et Metcalf, 1967; Pereira et Clermont, 1971}. Toutefois, se1on la derniere possi-bilite, les cellules auraienta
se deplacer contre le mouvement centri-pete de differenciation des thymocytes pour atteindre la zonesous-capsulaire du cortex (Sainte-Marie, 1973), ce qui rend nettement moins plausible 1 'entree des precurseurs au niveau de la medulla.
Il semble toutefois que des cellules de moelle osseuse migrent vers le thymus pour donner naissance aux grands et moyens lymphocytes dans la zone sous-capsulaire du cortex. Ces cellules etant absentes · du cortex pro fond et de 1 a medulla, nous pouvons no us demander si
elles degenerent in situ ou si elles se transforment en petits lympho-cytes par mitoses reductives. Kindred (1955) a calcule que le taux de mitoses de ces lymphocytes est de 7.0% et leur taux de pycnose, 2.7% dans un thymus de rat age de 80 jours. Sur la base du temps relatif d'une mitose par rapport
a
une pycnose (Regaud et Lacassagne, 1927), Sainte-Marie (1964b) a estime que la production des gros et moyens lymphocytes etait de vingt fois superieurea
leur destruction par pycnose; ce phenomene ne peut done pasa
lui seul expliquer la dispa-rition de ces cellules. Certains auteurs ont suggere que ces cellules se transformeraient en petits lymphocytes par 7 "mitoses reductives11{Dustin, 1913, 1920; Dustin et Gregoire, 1931). Des experiences d'auto-radiographie, en plus de confirmer la justesse de cette hypothese, montrent que les petits lymphocytes nouvellement formes, passent dans la medulla (Bryant et Kelly, 1953; Yoffey et coll., 1959; Cronkite et coll., 1959; Everett et coll., 1960), dans laquelle la majorite d'entre eux seraient detruits {Matsuyama et coll., 1966; Poste et
01 son, 1973} •
Le fait que le sang veineux du thymus soit plus riche en lympho-cytes que le sang arteriel~ suggere que le thymus fournit des lympho-cytes
a
la circulation (Sainte-Marie et leblond, 1964a,b). Une partie des lymphocytes medullaires quittent done le thymus, probablement par diapedesea
travers l'endothelium des veinules post-capillaires (Sainte-Marie et Leblond, 1964a, l964b, 1965).En resume, un schema de la differenciation des lymphocytes T commence
a
se preciser: les grands lymphocytes abondants en peripherie du cortex thymique, subissent une serie de 7 mitoses nucleoreductives. Ce processus de maturation lymphocytaire est accompagne par une reduc-tion de la quantite de cytoplasme. Le volume cytoplasmique diminuant plus rapidement que le volume nucleaire, le resultat net est une augmen-tation du rapport nucleo-cellulaire. Cette chaine de maturation est orientee dans le thymus de fa~on centripete, c'est-a-dire, que les grands lymphocytes en peripherie d'un lobule donnent naissance a une serie de petits lymphocytes pres de la m~dulla. Une des consequences directes de cette distribution est la genese d'une pression qui pousse les pe-tits lymphocytes dans la medulla. Les lymphocytes acquierent alors une grande mobilite, ce qui leur permettra de quitter le thymus, via la circulation, par diapedese au travers des parois des veinules post-capillaires (Sainte-Marie et Leblond, 1964a, 1964b, 1965).Au niveau de la membrane cellulaire, ce processus de maturation se manifeste par des modifications antigeniques (Boyse et call., 1968;
0
Order et Waksman, 1969; Sch1es·inger, 1972). Ce1les-ci consistent en une diminution dans la concentration des antigenes Thy-1 et une aug-mentation des antigenes H-2. Accompagnant aussi cette maturation, nous pouvon·s signaler une baisse de la sensibilite
a
la cortisone(Blomgren et Andersson, 1970; Claman, 1972). Clark (1973) a montre une augmentation de la reponse
a
la PHA des lymphocytes T de fa~onconcommitante
a
leur maturation.Le modele de la differenciation des lyrnphocytes thymiques enonce precedemment a ete elabore grace
a
des techniques d'histologie quanti-tative, en particulier l'ana1yse du nombre de ce1lu1es et de mitoses de chaque type cellu1aire d'une lignee {Marie, 1962; Sainte-Marie et Leblond, 1964b). Ce modele, base sur des etudes histo1ogiques ne tient toutefois pas compte de la grande heterogeneite fonctionne1le des lymphocytes T (Shortman, 1968; Shortman et coll., 1972; Shortman et call., 1975; Shortman et coll., 1976). Trois hypotheses sont ac-tuellement envisagees pour expliquer cette heterogeneite des lymphocy-tes. Elle representerait des etapes differentes d'une voie unique de differentiation, des divergences de maturationa
l'interieur de cette meme voie ou l'existence d'autres voies de differentiation. Ce pro-b1eme encore largement ouverta
la discussion et depassant un peu 1es buts de cette these, ces hypotheses ne seront pas abordees dans ce travail.2.3 ROLES DU THYMUS DANS LA DIFFERENCIATION DES LYMPHOCYTES T 2.3.1 Introduction
A partir de criteres morphologiques, il a ete etabli de-puis longtemps que le thymus etait un organe producteur de lym-phocytes (KBlliker, 1879; Prenant, 1894; St(jhr, 1906; Maximow, 1909; Salkind, 1915; de Winiwarter, 1924; Choi, 1931; Bastenie, 1932; Gregoire, 1932; Bloom, 1938; Kindred, 1940; Andreasen et Ottesen, 1945; Bierring, 1960; Schooley et Kelly, 1961). Toute-fois, on ne soup~onnait pas l1importance du thymus dans ce
pro-cessus et le caractere unique des lymphocytes thymiques. C1 est Miller (1961) qui, en signalant les anomalies importantes de la reponse immunologique
a
mediation cellulaire consecutivea
une thymectomie neonatale, permit de demontrer le role essentiel du thymus dans la differenciation d'une classe de lymphocytes specia-lises dans 1 'elaboration de la reponse ditea
mediation cellulai-re: les lymphocytes T.Depuis 1961, de tres nombreuses indications experimentales ont contribue
a
confirmer ce point eta
mettre en evidence deux elements du thymus impliques dans le processus de differenciation des lymphocytes T: les facteurs thymiques et le microenvironnement inducteur. Dans cette section, nous reverrons l'etat des connais-sances sur ces deux elements du thymus.2.3.2 facteurs thymigues
Dans cette section nous analyserons brievement les types d'indications experimentales demontrant le role essentiel de facteurs produits par le thymus et impliques dans la differen-ciation des lymphocytes T sans faire une etude detaillee et comparative de chacun de ces facteurs. De te 11 es etudes ont
d'ailleurs deja ete faites {Bach et Carnaud, 1976; Fournier, 1979; Zollinger, 1980).
l'importance des facteurs thymiques dans la differenciation des lymphocytes T a ete demontree par de tres nombreuses experien-ces qui peuvent etre regroupees en quatre differents types:
l} la correction, par traitement avec des facteurs thymiques, de deficiences immunologiques subsequentes a des thymec-tomies neonatales ou adultes (de Somer et coll., 1963; Sudo et coll., 1972} ou par des implantations de thymus en chambre de Millipore (Osoba et Miller, 1963};
2) la stimulation de la differenciation des lymphocytes par injection de facteurs thymiques a des animaux normaux (Bomskov et call., 1940; Roberts et White, 1949; Nakamoto, 1957; Duplan et coll., 1962; luckey et Venugopal, 1975; Mizutani et call., 1975);
3) la transformation in vitro de precurseurs en cellules ayant les caracteristiques antigeniques et fonctionnelles
des lymphocytes T (Bach et coll., 1971; Wilson et Bhaumick, 1973);
4) 1 'inhibition de la diff~renciation des lymphocytes T
par traitement avec des anticorps anti-facteur thymique (Potworowski et coll., 1975).
Cette liste de travaux d~montrant clairement un role actif des facteurs thymiques dans la diff~renciation des lymphocytes
T
pourrait s'allonger, mais nous avons d~cide d'en faire un tableau de fa~ona
faciliter la lecture de cet ouvrage (Tableau I).Plusieurs donn~es exp~rimentales permettent d'associer gene-ralement la production de ces facteurs aux cellules reticul~es
de la medulla. Les premieres indications sont d'ordre histologique; pour plusieurs auteurs, le thymus est considere comme une glande endocrine
a
cause de la pr~sence de cellules ayant lescaracteris-tiques de cellules secretrices: les cellules reticuloepitheliales
{CRE) de la medulla (Clark, 1963; Hoshino, 1963; Weiss, 1963).
Plus r~cemment, Teodorczyk et collaborateurs (1975), par l'emploi d'anticorps diriges contre leur preparation de facteur thymique ont pu demontre par immunofluorescence, la presence de ce facteur dans les cellules reticuloepitheliales de la medulla thy-mique.
AUTEURS BOMSKOV et colt., 1940 ROBERTS et WHITE, 1949
-METCALF, 1956 . NAKAMOTO, 1957 DUPLAN et co11.,1962 DeSOMER et coli., 1963 KLEIII, 1966 GOLOSTEJN et CO 11., 1966 TRAININ et con .• 1969 FLORENTIN et KIGER, 1971,73 SUOO et coll., 1972 WILSON et BHAUMICK, 1973 KOMURO et BOYSE, 1973 GOLOSTEIN, 1974. 1975 LUCKEY et VENUGOPAL, 1975 HIZUTANI et coll., 1975 BACH et CARNAUO, 1976 TABLEAU I-
-SOURCE PREPIIRATION ET FltACTIONNrt-1ENT CHUliQUf NATUP.£ DOSAGE BIOLOGIQUE veau alcool 11 pi de augmentation du phocytcs p~riph~riques non~re de lym-chez
l'hon~ue et le rat veau solvant alcal in .-thana~ froid prot~ine nucl~o- lymphopoi~se chez le rat
-
--sourls solution saline substance lymphopoii!se chez la souris hOPlPlC thermolabile noo!Vea!H1ee et la sourls adulte
et dialyse thymectOPlist'!e
nollln~ LSF
veau acicle acHique, sub;tances lymphopoii!se chez le lapin alcool, Hher ~on-prot(liques
-rat ( vof r Mete a lf) LSF rapport lymphocytes/polynucl!-aires chez la jeune souris veau solution tampon de Hank i ndHemi ntle lymphopoii!se chez la jeune sou-ris thymectomis~ a la
naissan-ce
veau COIII!Ie Goldstein thymosine hyperplasie ganglionnaire veau solution saline, ac~tone, sul fate glycoprot~i thYP~osine, ne lymphopoii!se
rosette d'ammonium, ultra- fraction 5
filtration, l!lectro PM:3200 focalisation
solution saline, THF
veau chromatographie polypeptide GvH in vitro PM:3000
-veau ~than::>l indtltemintle baisse immunologiques d~ plusieurs fonctions veau acide sulfurique, sulfate d'ammoniun sique thermo-prot!!ine ba- lymphopoiese chez la souris
1
thymectomis~e 11 la naissan-chrOPlatographie su labile ce; effet radioprotecteur colonne P~l: 20,000 I
veau sulfate d'ammonium proteine neu- induction de ultracentrifugati~ tre: PM de l'antig~ne Thy-1 94 ,000; prot~i
ne atide:P~l
de 110,000
souris COPlme Goldstein thymosine differentiation des lympho-cytes ~ partir de precurseurs in vitro
veau tampon alllnonium, thymopofl!tine electromyographie de transmis-ultrafiltration et I et 11; pep- sion neuromusculaire
chrOPlatographie tides Pf1:5562 souris
COJllllE! Hetcalf 2 protliines ce lymphopoi~se et au9mentation
boeuf PM 17,000 et de la synth~se d'anticorps che7. 80,000 la souris nouveau·n~
boeuf a c.:! tone 2 prot!ines de lJPlphopoi~se et hypocalcemie PM 68,000 et
57,000
-si! rum ultrafiltration, facteur thymi induction de l'antig~ne Thy-1 de pure chrOPlatographie que ci rculant rosette
peptide PM: 1000
TABLEAU DES DIFFERE~TES PREPARATIONS DE FACTEURS THYMIQUES
L'activite secretrice du thymus a pu etre mise en evidence par la correction de l'effet immunosuppresseur de la thymectomie par une greffe de thymus en chambre millipore (Levey et coll., 1963; Osaba et Miller, 1963; Law et co11., 1964}. Nous pouvons mentionner que les facteurs sont souvent prepares
a
partir du stro-ma epithelial {Gregoire et Duchateau, 1956; Potworowski et call., 1977) ou de surnageant de cultures de cellules epitheliales in vi-tro {Kruisbeek et call., 1978)Bien que maintenant la source des facteurs thymiques soit assez bien connue, il est encore impossible de determiner s'il existe un ou plusieurs facteurs thymiques. En effet, les simi-litudes de poids moleculaires observees entre certaines preparations (fraction 8 de la thymosine: 3,200 et 2,400 daltons, facteur
humoral thymique, THF: 3,000 daltons, le facteur thyrnique soluble, STF: 2,700 daltons) suggereraient une identite de ces facteurs. Dans cette recherche d'une parente biochimique, les differences de poids moleculaires notees pour d'autres preparations pour-raient etre reliees
a
des degres de polymerisation differents. Ce dernier point est appuye par 1 'observation de 1 'augmentation du facteur thymique circulant {1,000 daltons) subsequemmenta
une injection de thymosine (3,000 daltons) suggerant que cette derni~re serait un precurseur metabolique du facteur thy-mique circulant. La grande identite qui existe dans la sequence0
des acides amin~s entre le facteur thymique circulant et le facteur humoral thymique apporte des arguments suppl~mentaires
a
1 'identite des differentes hormones thymiques. Toutefois, le meme type de comparaison de sequences d'acides amines effectuee cette fois-ci entre la thymoproteine II et le facteur thymique circulant souligne l'absence compl~te d'identite entre ces deux preparations (Bach et Carnaud, 1976). Il faut done conclure qu'en pr~sence de donnees encore trap fragmentaires, la question del'identit~ des hormones thymiques demeure toujours non resolue.
Pour les besoins de cette thase, ces differentes prepara-tions seront regroup~es sous le nom "d'horrnone thymique" en spe-cifiant toutefois son origine.
11 semble maintenant clair que 1 'hormone thymique possade un role actif dans la differenciation des lymphocytes T. Toute-fois, l'echec de reconstitution par 1 'hormone de certains modeles de souris thymoprivees comme la souris 11nude" (Stutman, 1974; Pierpaoli et Besedowski, 1975) semble indiquer qu'elle n'est pas le seul element inducteur de la differenciation des lymphocytes T.
2.3.3 Microenvironnement thymigue
Considerant d'une part le role essentiel du thymus dans la differenciation des cellules T et d'autre part l'incapacite de
l'hormone d'assurer toutes les etapes de ce processus, il deve-nait important d'etudier le role du microenvironnement thymique, c'est-a-dire du stroma epithelial depourvu de facteur soluble dans la maturation des lymphocytes T.
L'examen de la litterature ne nous montre pas d'experien-ce realisee dans d'experien-ce but car il semble impossible d'obtenir un microenvironnement non altere par traitement mais epure de toute hormone thymique. La principale difficulte reside dans le fait que l'hormone est justement un produit de secretion des CRE.
Ainsi, dans la litterature, nous ne retrouvons que des arguments indirects exprimes
a
partir de l'echec de reconstitution par les facteurs thymiques de certains modeles,comme la souris athymique 11nude", couplesa
des succes obtenusa
partir degref-fes de thymus. Nous pouvons signaler
a
cet effet les experiences de Stutman suggerant la participation du microenvironnementa
la differenciation des lymphocytes T.Dans ses experiences, Stutman a utilise des souris agees de 45 jours,. thymectomisees
a
la naissance. Aces souris, il greffa soit des thymus normaux, soit des thymus enfermes dans des chambres de millipore excluant ainsi toute possibilite de contact entre le thymus et les precurseurs du receveut. L'efficacite des differentes cellules precurseursa
reconstituer la reponseimmune de l'anima1 a ~t~ etudiee. Stutman a alors montr~ que des cellules hematopoietiques d'embryons de 16 jours ou moins n~cessitaient la presence du thymus libre alors que pour les precurseurs d'embryons plus ag~s, les thymus en chambre de
millipore, c'est-a-dire 1 'hormone thymique seule, etaient suffi-sante
a
la differenciation (Stutman et coll., 1969; Stutman, 1977).De plus, il a ~te d~montre que les souris 11nude" pouvaient etre
rendues immunocompetentes par une greffe de thymus entier (Prit-chard et Micklem, 1972; Prit(Prit-chard et co11., 1973; Wortis, 1974; Stutman, 1974, 1975) alors que des extraits solubles de thymus ou des thymus en chambre de millipore etaient inefficaces (Stutman et Good, 1971, 1973; Stutman, 1974, 1975; Pierpaoli et Besedovsky, 1975).
Toutes ces donnees demontrent que 1'accessibi1ite au micro-environnement thymique semble necessaire pour d~clencher la diffe-renciation des precurseurs 11
precoces" (presents avant le 16e jour de gestation), a1ors que pour les precurseurs 11
tardifs" {presents apres le 16e jour de gestation) le contact avec les elements du microenvironnement n'est plus essentiel, peut-etre parce qu'il a deja eu lieu. Stutman appe1le ces deux types de precurseurs, les cellules prethymique et post-thymique respectivement. Les donnees de Levey et Burleson (1972, 1975) sur la competence immunitaire induite
a
des precurseurs par perfusion in vitro dans un thymussemblent appuyer ce point.
P1usieurs auteurs attribuent aux CRE thymiques la fonc-tion inductrice du microenvironnement thymique (Ford, 1966; Moore et Owen, 1967). Ainsi, des cultures de CRE thymiques se sont
av~r~es efficaces dans 1 •induction de marqueurs de lymphocytes T
a
des pr~curseurs de moelle osseuse (Pyke et Gelfand, 1974;Papiernick et coll., 1975; Sato et coll., 1976; Gershwin et coll., 1978; Willis-Carr et call., 1978). Toutefois, ces travaux peu-vent etre critiqu~s sur la pr~sence possible d1
hormone thymique
s~cr~t~e par les CRE, tel que demontre par plusieurs auteurs {Pyke et call., 1975; Willis-Carr et call., 1978; Kruisbeek et Astaldi, 1979}.
La nature du premier element inducteur de la
diff~rencia-tion, celui qui prec~de 1 •hormone thymique, est encore sujet
a
discussion; les deux possibilites les plus vraisemblables sont, soit une forte concentration en hormones thymiques in situ, soit un element insoluble et probablement membranaire. Les travaux de Potworowski mettant en evidence une antig~nicite sp~cifique des membranes perivasculaires thymiques supportent p1utot la deuxie-me hypoth~se (Potworowski, 1977).2.3.4 STF et ITF
Dans notre laboratoire, nous avons identifi~ deux frac-tions du stroma qui pourraient correspondre respectivement
a
l'element inducteur du microenvironnement discute pr~c~demment et
a
l'hormone thymique.Le premier element est la fraction insoluble du stroma thymique (ITF). Cette fraction membranaire possede une
antigeni-cit~ specifique localisee dans les doubles membranes perivascu-laires de la m~dulla thymique (Potworowski, 1977). lorsqu'in-jecte
a
des souris, 1 'ITF provoque une accumulation de grandes cellules lymphoblastoVdes en peripherie du cortex thymique (Zollinger et Potworowski, 1977; Zollinger et coll ., 1977; Fournier, 1979), qui r~sulte d'une augmentation de la migration de precurseurs venant de la moelle osseuse (Fournier et Potworow-ski, 1979; Fournier, 1979). De plus, l'ITF provoque aussi par injection, une augmentation de la population de cellules de rate sensiblesa
l'hydrocortisone (Zollinger et Potworowski, 1979). Lorsqu'injecte seul, l'ITF est incapable de reconstituer la po-pulation de ~ellules cooperatrices avec 1es lymphocytes B pour la production d'anticorps, tel que d~termine par la technique des PFC (plaque forming cells} de souris thymectomisees, irradiees et ayant re~u de la moelle osseuse (Zollinger et coll ., 1980) oude souris nude (Teodorczyk-Injeyan et Potworowski, 1980). De plus, 1 'ITF permet 1 'induction de 1 'antig~ne Thy-1
a
des cellu-les de moelle osseuse (Paquette et co11., 1981).De son cote, le STF, une molecule d'environ 3,000 daltons de poids moleculaire, est localise presqu'exclusivement dans le cytoplasme des cellules reticuloepitheliales (Potworowski, 1977). Du point de vue de son activite, le STF est comparable aux autres preparations d'hormone thymique decrites dans la litterature et plus particulierement
a
celle de Trainin (Trainin et call., 1975): l'incubation de thymocytes avec du STF les transforme en cellules resistantesa
1 'hydrocortisone (Zollinger et Potworowski, 1979) et diminue la concentration de surface en antig~ne Thy-1 (Paquette et call., 1981). Lorsqu'injecte seul, le STF est incapable d'as-surer la reconstitution en PFC de souris thymectomisees irradiees et ayant re~u de la moelle osseuse (Zollinger et call., 1980) ou de souris nude (Teodorczyk-lnjeyan et Potworowski, 1979). Deplus, le STF permet l'induction de l'antig~ne Thy-1 sur des cellu-les de moelle osseuse {Paquette et call., 1981).
L'action sequentielle de l'ITF et du STF dans l'induction et la perte de sensibilite
a
l'hydrocortisone (Zollinger et Potworow-ski, 1979), 1 'induction de l'antigene Thy-1 et sa diminution de concentration de surface (Paquette et call., 1981) et lareconstitu-tion en PFC de souris thymectomisees (Zollinger et coll ., 1980) et de souris nude (Teodorczyk-Injeyan et Potworowski, 1979) nous permettent de relier 1 'ITF au premier element inducteur de la differenciation des lymphocytes T, tel que postule par Stutman et le STF comme etant 1 'hormone thyrnique.
Un autre systeme dans lequel 1 'effet de 1 'ITF et du STF fait bien ressortir leur role dans la differenciation des lympho-cytes T, est la reaction du greffon contre 1 'hote (GvHR). Il a ete demontre qu'au moins deux populations de lyrnphocytes collabo-rent dans 1 'initiation de la reaction GvH: une premiere peu matu-re ayant un role d'arnplificateur de l 'intensite de la matu-reaction et comp-osee de lyrnphocytes
a
forte concentration en anti gene Thy-1 et une deuxierne plus mature, representant les cellules effectri-ces (Zollinger et Potworowski, 1979; Paquette et coll ., 1981).Il a ete demontre qu'une injection d'ITF
a
des souris pa-rentales augrnentait leur potentiel d'induction de la GvH dans des souris F1, alors qu'une injection de STF le reduisait (Zollinger et Potworowski, 1979; Paquette et coll ., 1981). Par 1 'utilisation de souris parentales thymectomisees, irradiees et reconstituees avec de la moelle osseuse, il a pu etre demontre que les cellules induites par 1 'ITF, seules, ne pouvaient induire de GvH dans un F
1, alors que celles induites par le STF seul provoquaient une GvH
lorsqu'injectees seules
a
des souris F1. Ces dernieres cellules presentent une forte concentration en antigene Thy-1 qui n'est pas sensiblea
l'effet reducteur de 1 'incubation avec le STF, contrairement aux cellules induites par l'ITF (Paquette et coll ., 1981). Tous ces resultats pris dans leur ensemble suggerent tres fortement que la population de cellules amplificatrices de la reaction GvH correspond aux cellules induites par l'ITF alors que celles induites par le STF correspondraient aux cellules effectrices de la reaction.2.4 CONCLUSION
Considerant d'une part ces resultats et d'autre part le modele de Stutman mentionne plus haut, un schema d'ensemble de la differen-ciation des lymphocytes T commence
a
se dessiner (voir Figure 1 ). Les cellules precurseurs prethymiques par contact avec l'ITF subissent un premier stage de differenciation qui les rendra sensiblesa
l'action de 1 'hormone, c'est-a-dire des precurseurs post-thymiques. Ces dernie-res cellules pdernie-resentent une forte concentration en antigene Thy-1, une grande sensibilitea
1 'hydrocortisone et se retrouvent aussi bien en peripherie du cortex thymique que dans la rate. Ces cellules cor-respondraient aux cellules amplificatrices de la reaction GvH. Ces cellules ont aussi comme caracteristique leur sensibilitea
l'hormone thymique, en presence de laquelle elles sont differenciees enlympho-PRECURSEURS (MOELLE OSSEUSE) FIGURE 1
ITF
,_ _ _ _ ...,.. F-Thy 1 HYCROCORTICOSENSIBLE AMPLI FI CAT RICE EN GvH STF F-Thy 1EFFECTRICE EN GvH
STF f-Thy l
HYDROCORTICORESISTANTE B-COOPERATRICE
SCHEMA DE LA DIFFERENCIATION DES LYMPHOCYTES T. Les cellules Thy 1 negatives de la moelle osseuse par contact avec l'ITF sont transformees en cellu-les sensibcellu-les
a
l'hydrocortisone et presentant une forte concentration en antigene Thy 1 (F-Thy 1). Les cellules ont une fonction amplificatrice dans la reaction GvH. Elles peuvent etre differentiees en lymphocytes T matures par le STF. Cette diffe-renciation s'accompagne d'une baisse de concentra-tion en antigene Thy-1 {f-Thy l) et d'une perte de sensibilitea
l'hydrocortisone. Ces dernieres cellu-les cooperent avec cellu-les lymphocytes B pour la produc-tion d'anticorps. A un deuxieme niveau, les cellules de moe 11 e os se use peuvent et re differenti ees en ce 11 u-les T,a
forte concentration en antigene Thy-1 non reductible. Ces dernieres cellules sont 1es cellules effectrices de la reaction GvH.cytes T matures. Cette maturation se traduit par une perte de
sensi-bilit~
a
1 'hydrocortisone, une baisse de concentration d'antigene Thy-1, une perte du potentiel d'amplifier la· reaction GvH et un gain de la fonction de collaboration avec les lymphocytes B pour la pro-duction d'anticorps.Ces resultats suggerent aussi que le STF agirait directement sur les precurseurs prethymiques pour l'induction de l'antigene Thy-1 et de la fonction effectrice dans la reaction GvH.
De la discussion du premier chapitre, il est ressorti que le thymus
~
etait implique dans la determination de la susceptibilitea
la leucemie, soit de par son role dans la reponse immunitaire elaboree par leslymphocytes T et dont il assure la diff~renciation, soit de par son microenvironnement propice
a
l'induction de la leucemie lymphoTde. Dans ce chapitre nous avons decrit deux elements biologiquement actifs du microenvironnement thymique, l'ITF et le STF, responsables de la diffe-renciation des lymphocytes T. A partir de ces deux chapitres, il devient interessant de se questionner sur le role de l'ITF et du STF dans le processus de leucemogenese. Pour repondrea
cette question, nous avons decide d'etudier le role du thymus dans la leucemogenese des souris AKR. Avant d'examiner les diverses indications experimen-tales qui nous permettent de relier un rolea
l'JTF et au STF dans le declenchement de la leucemie des souris AKR, un bref rappel de l'histo-rique de cette souche et des caracteristiques de la leucemie AKR sera40.
CHAPITRE 3
LES SOURIS AKR
3.0 INTRODUCTION
Deja vers les annees 1930, plusieurs auteurs se sont interesses au phenomene de la leucemie chez la souris et ont essaye de develop-per des souches de souris
a
haute incidence leucemique (Little, 1920; MacDowell et Richter, 1935; Strong, 1936, 1942; Furth, 1934, 1935, 1939}.L'interet pour ces leucemies de la souris est assez aise
a
corn-prendre. Nous pouvons facilement faire un parallele entre la leucemie de la souris et certaines leucemies humaines. En effet, Furth (1939) et collaborateurs ont montre 1 'existence d'analogies histologiques entre la leucemie de la souris et la leucemie humaine, tant au point de vuedes tissus leucemiques qu1au niveau de simples frottis sanguins: les cellules neoplasiques de ces deux types de leucemie semblent morpholo-giquement similaires.
Au point de vue de 1 'evolution de la maladie, Gross (1961) a note que chez les deux especes, on retrouve certai ns points cornmuns: hyper-plasie ganglionnaire, certaines tumeurs communes et role central du thymus dans l'ontogenese de la maladie. La seule difference que l'on