Isolation and molecular characterization of single circulating rare cells : developing innovative methods for predictive oncology and non-invasive prenatal diagnosis
299
0
0
Texte intégral
Figure
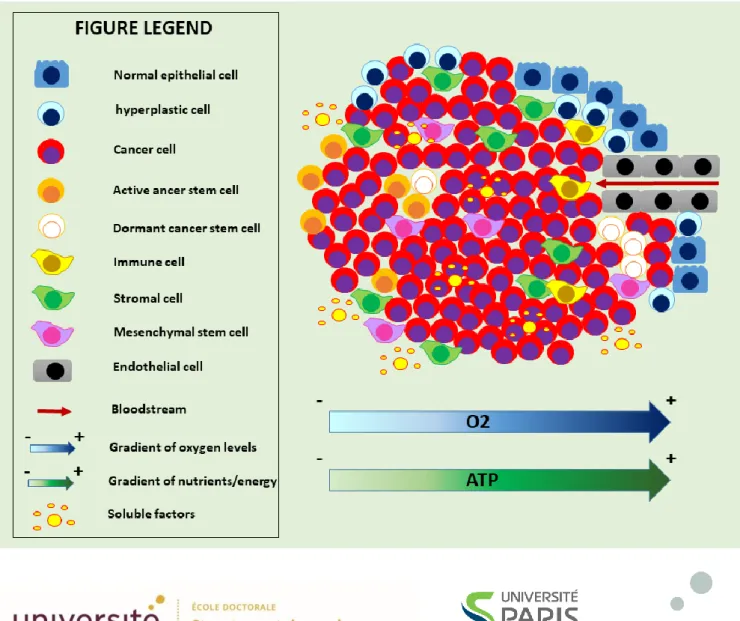
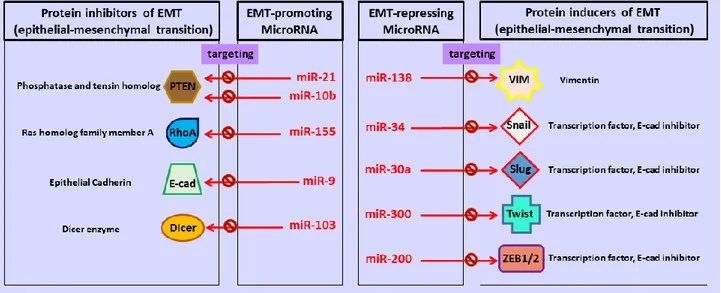

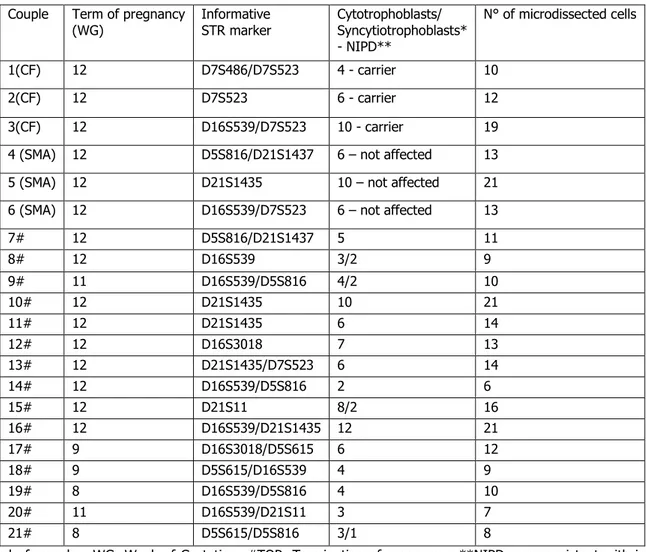
+7
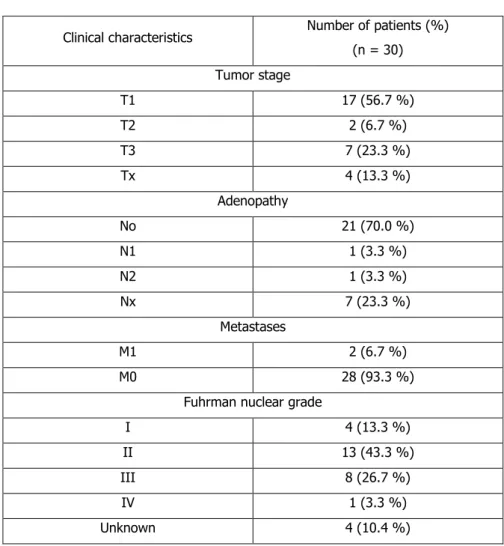
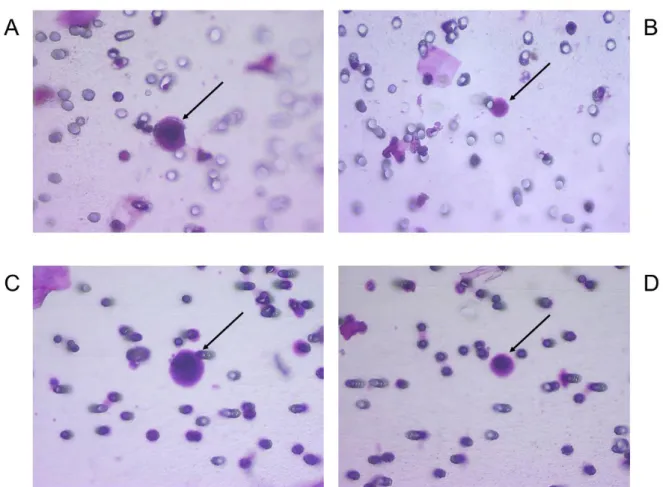
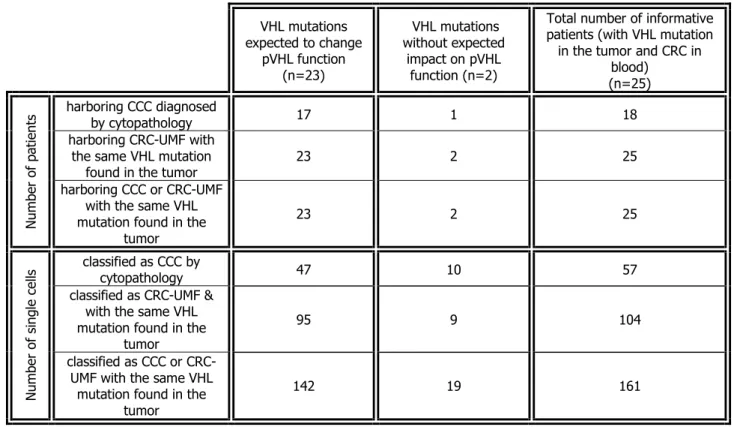
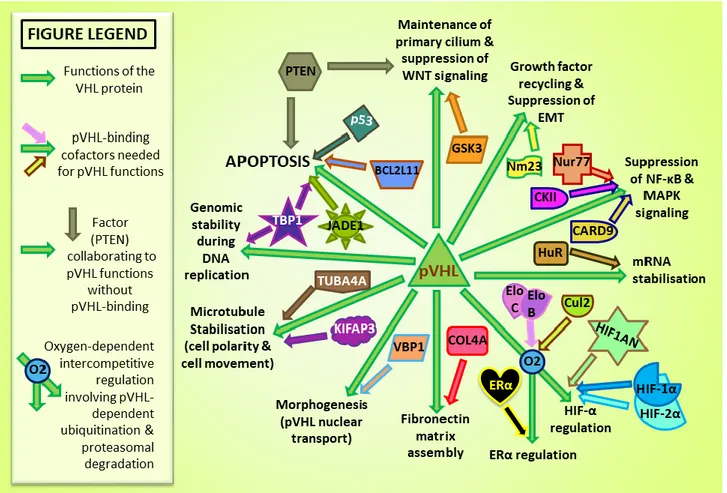

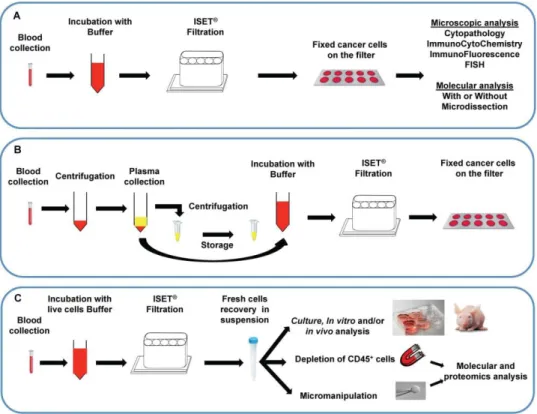
Documents relatifs