Université de Montréal
Développement d'un modèle murin de la maladie de
Parkinson par augmentation compensatoire de
l'arborisation axonale dopaminergique-nigrostriée
par
William Tanguay
Département de neurosciences Faculté de médecine
Mémoire présenté à la Faculté des études supérieures en vue de l’obtention du grade de Maître ès Sciences (M. Sc.)
en Neurosciences
Décembre 2016
Résumé
Les neurones dopaminergiques de la substance noire (SNc) sont les plus vulnérables à la dégénérescence dans la maladie de Parkinson et ses modèles animaux. Suite à des travaux antérieurs et à des résultats préliminaires du laboratoire Trudeau, notre hypothèse actuelle suggère que la très grande taille de l'arborisation axonale des neurones de la SNc soit un facteur clé à l'origine de leur vulnérabilité, puisque cet état devrait être associé à un taux élevé de phosphorylation oxydative et de production de radicaux libres. En accord avec cette hypothèse, les autres populations dopaminergiques, dotées d'arborisations de moindre taille, résistent mieux aux lésions expérimentales et à la maladie chez l'humain. L'objectif du présent projet était de développer un modèle murin dans lequel les neurones de la SNc présentent une taille d'arborisation axonale plus grande, se rapprochant davantage de celle observée chez l'humain et en reproduisant la vulnérabilité, ce qui pourrait représenter une percée importante dans l'identification de nouvelles approches thérapeutiques. Basée sur le bourgeonnement axonal compensatoire des neurones dopaminergiques suite à des lésions partielles, la méthode utilisée fut l'injection unilatérale intranigrale de la toxine 6-hydroxydopamine (6-OHDA) à quelques jours de vie (P5), en visant l'élimination de 50% des neurones de la SNc. Un immunomarquage contre la tyrosine hydroxylase (TH), enzyme de synthèse de la dopamine, ainsi qu'une quantification du signal TH dans le striatum et des comptes neuronaux stéréologiques ont permis de quantifier la lésion partielle et de mettre en évidence la présence d'une croissance axonale compensatoire des neurones dopaminergiques survivants, à 10 et 90 jours post-lésion, suggérant une compensation précoce. Afin de mettre en évidence l'origine du bourgeonnement axonal, nous avons injecté un vecteur viral de type AAV encodant une protéine fluorescente (EYFP) dans la SNc ou la VTA des animaux adultes. Nos résultats confirment la présence de neurones nigrostriés à plus grande arborisation suivant une lésion unilatérale précoce à la 6-OHDA, dont la vulnérabilité accrue pourra être évaluée dans des expériences à venir par des protocoles lésionnels au MPTP, une toxine permettant de modéliser la maladie de Parkinson chez la souris. Mots-clés : Maladie de Parkinson, Dopamine, Modèle animal, Vulnérabilité, Arborisation axonale, 6-hydroxydopamine, Substance noire, Aire tegmentaire ventrale,
Abstract
Dopaminergic neurons of the substantia nigra (SNc) are amongst the most vulnerable to neurodegeneration in Parkinson's disease and its animal models. According to previous work and preliminary results in our laboratory, our present hypothesis postulates that the large axonal arborisation size of SNc neurons is a key driving factor in their vulnerability, since this characteristic is associated with increased oxidative phosphorylation levels and free radicals production. In agreement with this hypothesis, other dopaminergic populations with smaller axonal arbors better resist to experimental lesions and to the disease process in humans. The current project aims to develop a mouse model in which SNc neurons present an axonal arborisation of increased size, closer to what is encountered in humans, thus reproducing their vulnerability, which could represent an important breakthrough in the identification of new therapeutic approaches. Based on compensatory axonal sprouting of dopaminergic neurons following partial lesions, the method used was the unilateral intranigral injection of the toxin 6-hydroxydopamine (6-OHDA) at an early age (P5), to induce the loss of approximately 50% of SNc neurons. Immunostaining against tyrosine hydroxylase (TH), an enzyme required for the synthesis of dopamine, TH signal quantification in the striatum and stereological counting of neurons allowed for the quantification of the partial lesion and demonstrated compensatory axonal sprouting at 10 and 90 days post-lesion, with our results suggesting an early compensation. To better characterize the origin of axonal sprouting, we injected an AAV viral vector encoding a fluorescent protein (EYFP) in either the SNc or the VTA of adult animals. Our results confirm the presence of nigrostriatal neurons with increased arborisation sizes following early unilateral lesion using 6-OHDA, whose increased vulnerability will be evaluated in future experiments through lesion protocols using MPTP, a toxin used to model Parkinson's disease in mice.
Keywords : Parkinson's disease, Dopamine, Animal Model, Vulnerability, Axonal Arborisation, 6-Hydroxydopamine, Substantia Nigra, Ventral Tegmental Area, Compensatory Sprouting
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières... iv
Liste des tableaux et graphiques ... vii
Liste des figures ... x
Liste des sigles et abréviations ... xi
Remerciements ... xiv
Avant-propos... xv
Introduction ... 1
Le système dopaminergique ... 3
Aspects neuroanatomiques ... 4
Les neurones dopaminergiques de la SNc et la VTA ... 4
Le striatum ... 11
Aspects moléculaires ... 15
La DA et ses voies de synthèse et de dégradation ... 15
La terminaison dopaminergique ... 15
Les récepteurs de la dopamine ... 16
La maladie de Parkinson ... 18
Symptomatologie de la maladie de Parkinson ... 18
Symptômes cardinaux ... 19
Autres symptômes moteurs ... 20
Symptômes sensitifs, végétatifs et cognitifs ... 20
Évolution clinique ... 21
Pathophysiologie de la MP ... 26
Aspects neuropathologiques ... 26
Fonctionnement et dysfonctionnement des voies directe et indirecte ... 28
Compréhension actuelle de la pathogénèse de la neurodégénérescence dopaminergique dans la maladie de Parkinson ... 31
Modèles animaux actuels de la maladie de Parkinson ... 35
Modèles neurotoxiques ... 35
Modèles génétiques ... 38
Limites des modèles actuels et perspectives futures ... 39
Modèle d’accroissement compensatoire de l’arborisation axonale par lésion partielle précoce de la substance noire ... 41
La taille de l'arborisation axonale: possible facteur de vulnérabilité neuronale dans la maladie de Parkinson ... 42
Réinnervation et compensation dopaminergique ... 44
Hypothèse de recherche ... 46
Principes expérimentaux ... 49
Matériel et méthodes ... 63
Résultats ... 80
Identification des coordonnées stéréotaxiques d’injection ... 80
Identification de la dose de 6-OHDA permettant une lésion partielle de la SNc chez la souris P5 ... 81
Influence du génotype sur le taux de survie des neurones TH suivant une injection de 0,5μl de solution de 6-OHDA à concentration de 0,25 μg/ml ... 86
Lésion Partielle de la SNc – Groupe P15... 87
Lésion Partielle de la SNc – Groupe P90... 99
Expression d'EYFP dans les groupes transfectés dans la SNc ... 111
Expression d'EYFP dans les groupes transfectés dans la VTA ... 119
Discussion ... 131
Dose de 6-OHDA intranigrale requise pour une lésion partielle des neurones DA de la SNc chez le souriceau P15 ... 131
La vulnérabilité des neurones DA du mésencéphale pourrait varier en fonction du génotype
et des niveaux de DAT qui y sont associés ... 133
Les neurones DA du mésencéphale semblent compenser presque complètement la perte de neurones de la SNc en rétablissant la quantité d'arborisation axonale TH+ dans le striatum post-lésionnel à un temps précoce ... 135
La compensation observée dans le dSTR semble provenir à la fois de la VTA et de la SNc, d'après les résultats des premières transfections virales ... 138
Le volume de l'arborisation axonale moyenne des neurones DA de la SNc est plus grand que celui des neurones DA de la VTA ... 141
Les neurones DA survivants dans la SNc augmentent la taille de leur arborisation axonale après une perte partielle de leur population ... 141
Relation entre l'importance de la lésion nigrale dopaminergique et l'augmentation de la taille de l'arborisation axonale des neurones dopaminergiques mésencéphaliques ... 143
Considérations méthodologiques ... 145
Améliorations possibles au modèle actuel ... 151
Perspectives futures ... 158
Conclusion ... 165
Bibliographie... i Annexe ... I
Liste des tableaux et graphiques
Tableau I. Nombre de neurones DA dans la SNc et la VTA de différents mammifères ... 43 Tableau II. Coordonnées stéréotaxiques des injections de 6-OHDA chez l'animal P15 ... 65 Tableau III. Coordonnées stéréotaxiques des injections d'AAV-EYFP chez l'animal P60 .... 67 Tableau IV. Résultats des décomptes neuronaux préliminaires ... 85 Graphique 1. Lésion de la SNc et effet sur le signal TH striatal – Groupe P15 WT ... 90 Graphique 2. Survie des neurones TH+ dans la SNc et la VTA et volume de signal TH dans le dSTR et le vSTR – Groupe P15 WT ... 91 Graphique 3. Volume moyen de l'arborisation axonale d'un neurone de la SNc et de la VTA et changement par rapport au côté contralésionnel – Groupe P15 WT ... 93 Graphique 4. Lésion de la SNc et effet sur le signal TH striatal – Groupe P15 DAT-Cre+/- .. 94
Graphique 5. Survie des neurones TH+ dans la SNc et la VTA et volume de signal TH dans le dSTR et le vSTR – Groupe P15 DAT-Cre+/- ... 95
Graphique 6. Volume moyen de l'arborisation axonale d'un neurone de la SNc et de la VTA et changement par rapport au côté contralésionnel – Groupe P15 DAT-Cre+/- ... 97
Graphique 7. Augmentation de la taille de l'arborisation axonale des neurones TH+ de la SNc et de la VTA en fonction de l'importance de la perte de neurones TH+ dans la SNc – Groupe P15 WT ... 98 Graphique 8. Augmentation de la taille de l'arborisation axonale des neurones TH+ de la SNc et de la VTA en fonction de l'importance de la perte de neurones TH+ dans la SNc – Groupe P15 DAT-Cre+/- ... 98
Graphique 9. Lésion de la SNc et effet sur le signal TH striatal – Groupe P90 WT ... 102 Graphique 10. Survie des neurones TH+ dans la SNc et la VTA et volume de signal TH dans le dSTR et le vSTR – Groupe P90 WT ... 103 Graphique 11. Volume moyen de l'arborisation axonale d'un neurone de la SNc et de la VTA et changement par rapport au côté contralésionnel – Groupe P90 WT ... 104 Graphique 12. Lésion de la SNc et effet sur le signal TH striatal – Groupe P90 DAT-Cre
Graphique 13. Lésion de la SNc et effet sur le signal TH striatal – Groupe P90 DAT-Cre
+/-(sous-groupe ayant reçu une transfection d'AAV-EYFP ciblée sur la VTA) ... 105 Graphique 14. Survie des neurones TH+ dans la SNc et la VTA et volume de signal TH dans le dSTR et le vSTR – Groupe P90 DAT-Cre+/- ... 107
Graphique 15. Volume moyen de l'arborisation axonale d'un neurone de la SNc et de la VTA et changement par rapport au côté contralésionnel – Groupe P90 DAT-Cre+/- ... 108
Graphique 16. Augmentation de la taille de l'arborisation axonale des neurones TH+ de la SNc et de la VTA en fonction de l'importance de la perte de neurones TH+ dans la SNc – Groupe P90 WT ... 109 Graphique 17. Augmentation de la taille de l'arborisation axonale des neurones TH+ de la SNc et de la VTA en fonction de l'importance de la perte de neurones TH+ dans la SNc – Groupe P90 DAT-Cre+/- ... 110
Graphique 18. Transfection de la SNc et effet sur le signal EYFP striatal – Groupe P90 DAT-Cre+/- (sous-groupe ayant reçu une transfection d'AAV-EYFP ciblée sur la SNc) ... 114
Graphique 19. Taux de transfection des neurones TH+ par l'AAV2-EYFP – Groupe P90 DAT-Cre+/- avec injections ciblées sur la SNc ... 115
Graphique 20. Estimations du volume moyen de l'arborisation axonale des neurones de la SNc – Sous-groupe P90 DAT-Cre+/- avec injections ciblées sur la SNc ... 116
Graphique 21. Estimations de l'augmentation moyenne du volume moyen de l'arborisation axonale d'un neurone de la SNc ipsilésionnelle – Sous-groupe P90 DAT-Cre+/- avec injections
ciblées sur la SNc ... 118 Graphique 22. Transfection de la VTA et effet sur le signal EYFP striatal – Groupe P90 DAT-Cre+/- (sous-groupe ayant reçu une transfection d'AAV-EYFP ciblée sur la VTA) ... 122
Graphique 23. Taux de transfection des neurones TH+ par l'AAV2-EYFP – Groupe P90 DAT-Cre+/- avec injections ciblées sur la VTA ... 123
Graphique 24. Estimations du volume moyen de l'arborisation axonale des neurones de la VTA – Sous-groupe P90 DAT-Cre+/- avec injections ciblées sur la VTA ... 124
Graphique 25. Estimations de l'augmentation moyenne du volume moyen de l'arborisation axonale d'un neurone de la VTA ipsilésionnelle – Sous-groupe P90 DAT-Cre+/- avec injections
Graphique 26. Relation entre la taille de la lésion dans la SNc et l'augmentation de la taille de l'arborisation axonale des neurones dopaminergiques survivants dans la SNc – Sous-groupe P90 DAT-Cre+/- avec injections ciblées sur la SNc ... 128
Graphique 27. Relation entre la taille de la lésion dans la SNc et l'augmentation de la taille de l'arborisation axonale des neurones dopaminergiques survivants dans la VTA – Sous-groupe P90 DAT-Cre+/- avec injections ciblées sur la VTA ... 128
Liste des figures
Figure 1. Anatomie des groupements dopaminergiques A9 et A10 chez l'humain et la souris . 5
Figure 2. Afférences de la SNc et de la VTA chez la souris ... 7
Figure 3. Voies efférentes dopaminergiques de la SNc et de la VTA ... 10
Figure 4. Anatomie du striatum humain et murin ... 12
Figure 5. Caractérisations neuropathologiques des synucléinopathies ... 27
Figure 6. Fonctionnement normal et pathologique des voies directe et indirecte du mouvement et schématisation de la neurotransmission striatale correspondante ... 29
Figure 7. Paradigme actuel de la pathophysiologie de la maladie de Parkinson ... 34
Figure 8. Chronologie des processus compensatoires suite à une lésion de la SNc ... 45
Figure 9. Illustration du fonctionnement du système Cre-lox ... 50
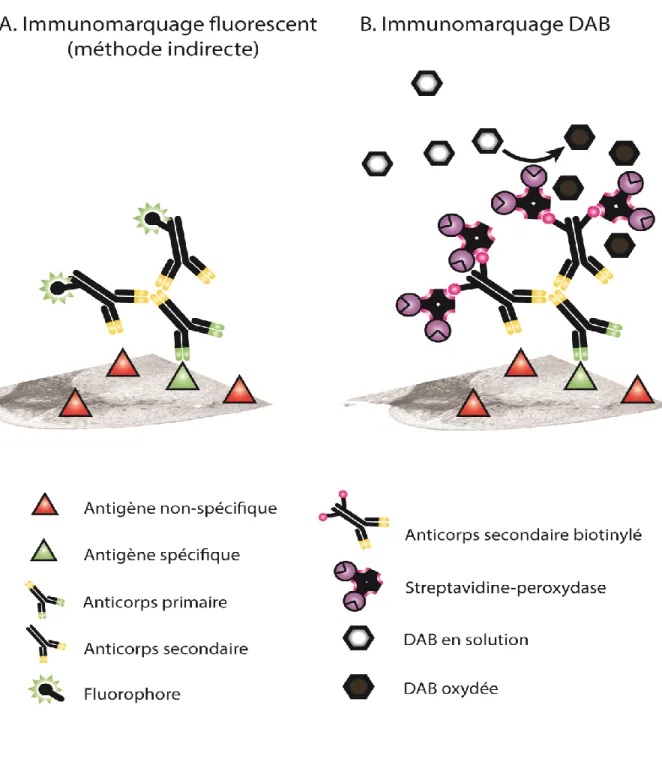
Figure 10. Immunomarquage fluorescent et immunomarquage à la DAB ... 56
Figure 11. Microscopie à épifluorescence et imagerie confocale à balayage laser ... 58
Figure 12. Fraction d'échantillonnage de la structure ... 60
Figure 13. Fraction d'échantillonnage de l'aire et échantillonnage aléatoire systématique ... 60
Figure 14. Fraction d'échantillonnage de l'épaisseur du tissu ... 61
Figure 15. Échantillonnage striatal pour quantification en microscopie confocale ... 74
Figure 16. Recherche des coordonnées d'injection stéréotaxique pour la lésion intranigrale chez le souriceau à 5 jours de vie ... 81
Figure 17. Identification de la dose de 6-OHDA permettant une lésion nigrale partielle ... 83
Figure 18. Décomptes neuronaux préliminaires ... 84
Figure 19. Exemples de lésions nigrales obtenues chez la souris P15 DAT-Cre+/- et WT ... 86
Figure 20. Effet de l'injection unilatérale de 6-OHDA dans la SNc gauche dans le groupe d'animaux P15 (WT et DAT-Cre+/-) ... 88
Figure 21. Effet de l'injection unilatérale de 6-OHDA dans la SNc gauche dans le groupe d'animaux P90 (WT et DAT-Cre+/-) ... 100
Liste des sigles et abréviations
AAV Virus adéno-associéARNm Acide ribonucléique messager ADN Acide désoxyribonucléique BO Bulbe olfactif
BSA Bovine serum albumin (albumine de sérum bovin)
CE Coefficient d'erreur
DA Dopamine
DAB 3,3’-Diaminobenzidine
DAT Transporteur membranaire de la dopamine DCX Doublecortin
EYFP Enhanced yellow fluorescent protein
GAP-43 Growth associated protein 43
GDNF Facteur neurotrophique dérivé de la glie GFP Green fluorescent protein
GP Globus pallidus
GPe Globus pallidus externe
GPi Globus pallidus interne
IL-1 Interleukine 1 LC Locus coeruleus
L-DOPA L-3,4-dihydroxyphenylalanine LPS Lipopolysaccharide
LTD Long-term depression (dépression synaptique à long terme)
LTP Long-term potentiation (potentialisation synaptique à long terme)
MAO Monoamine oxydase
MDV Mitochondria-derived vesicle (vésicule dérivée des mitochondries)
MFB Faisceau télencéphalique médian MOR Récepteur -opioïde
MPP+ 1-methyl-4-phenylpyridinium
NA Noradrénaline
NET Transporteur membranaire de la norépinephrine P(nombre) Jour postnatal (nombre)
PBP Noyau parabrachial pigmenté PCR Polymerase chain reaction
ROS Reactive oxygen species (dérivés réactifs de l'oxygène)
RRF Retrorubral field
SN Substance noire
SNc Substance noire, pars compacta
SNcd Substance noire, pars compacta, niveau dorsal SNcv Substance noire, pars compacta, niveau ventral SNl Substance noire, pars lateralis
SNr Substance noire, pars reticulata STN Noyau sous-thalamique
SYN1 Synapsine 1
TH Tyrosine hydroxylase
VGLUT2 Transporteur vésiculaire du glutamate (type 2) VMAT Transporteur vésiculaire des monoamines VTA Aire tegmentaire ventrale
WT Wild Type (Type sauvage)
5-HT 5-hydroxytryptamine, sérotonine 6-OHDA 6-hydroxydopamine
Remerciements
J’aimerais remercier Louis-Éric Trudeau pour son ouverture, sa disponibilité constante et son apport critique qui me fut d’une grande aide au long de mon cheminement. J’aimerais également remercier Marie-Josée Bourque, qui m’a appris à remettre en question mes approches et à toujours viser un questionnement plus rigoureux. J’aimerais enfin remercier Charles Ducrot, Nicolas Giguère, Consiglia Pacelli, Rafael Koerich Varaschin et Aurore Voisin pour leurs conseils, leur soutien à ma formation durant ces deux années passées au laboratoire, et surtout pour les souvenirs qu’ils me laissent. Si le café n’était pas toujours de qualité, les débats d’idées que nous avons eus en le buvant chaque jour l’étaient immanquablement, et ont façonné mon raisonnement scientifique bien plus que je ne l’aurais espéré.
Avant-propos
« Even when the work of a genius is subjected to critical analysis and no errors are found, it is important to realize that everything he has discovered in a particular field is almost nothing in comparison with what remains to be discovered. »
Santiago Ramón y Cajal Traduit de Reglas y Consejos sobre Investigación
Cientifica: Los tónicos de la voluntad, 1916
S'adressant aux jeunes chercheurs de son époque, Cajal trouvait les mots justes pour évoquer non seulement l'histoire, mais aussi le futur des neurosciences: plusieurs génies se sont succédé avant et après lui sans que cette pensée ne perde de sa justesse. Au fil des dernières années, les neurosciences ont connu une série de révolutions, ouvrant la voie à de larges horizons de recherches fondamentales et cliniques. La neuroimmunologie, la neuropharmacologie, la neurooncologie, la neuroréabilitation, les interfaces neuro-computationnels, les maladies psychiatriques et les maladies neurodéveloppementales sont autant de champs appelés à connaître des avancements menant, à terme, à des bénéfices importants au niveau de la santé des populations. Un défi qui demeure pourtant l’un des plus criants sera de mieux comprendre les maladies neurodégénératives pour mieux y faire face, car la population humaine en sera de plus en plus atteinte. Parmi ces maladies, la maladie de Parkinson et la maladie d’Alzheimer sont sans doute les plus connues, et sont à l’heure actuelle et depuis bon nombre d’années déjà le sujet d’efforts et d’investissements massifs pour en comprendre les causes et y trouver des traitements. Pourtant, la médecine reste encore aujourd’hui relativement impuissante face à leur progression, malgré les ressources consacrées à en élucider l'étiologie. L'ambition d'un jour pouvoir en traiter la cause est le tónico de la voluntad qui assure la poursuite des recherches.
Introduction
La maladie de Parkinson est la seconde maladie neurodégénérative en importance après la maladie d’Alzheimer, et sa prévalence devrait fortement augmenter dans les prochaines décennies en parallèle au vieillissement de la population (Kowal, Dall, Chakrabarti, Storm, & Jain, 2013). Bien que plusieurs mutations aient à ce jour été identifiés comme prédisposant directement à la maladie, environ 90% des cas sont considérés sporadiques (Lau & Breteler, 2006), et leur étiologie exacte est encore mal comprise. De plus, la maladie est détectée tardivement, puisque les symptômes moteurs, les plus manifestes, n’apparaissent souvent qu’après la disparition de 70 à 80% des neurones de la substance noire (SN) (Greenbaum et al., 2013). Bien que plusieurs traitements pharmacologiques et chirurgicaux existent pour diminuer ou retarder les symptômes, aucun ne permet de renverser ou d’arrêter cette mort neuronale. Une meilleure compréhension de la physiopathologie de la maladie de Parkinson est cruciale pour espérer d’une part dépister la maladie plus tôt, et d’autre part pour développer des traitements permettant d’en freiner l’évolution avant que les symptômes n’apparaissent.
De nombreux modèles animaux, tant chez les primates que chez le rat et la souris, ont permis d’élucider certains aspects de la maladie en se basant sur une lésion de la voie nigrostriée en utilisant diverses neurotoxines, la roténone, le paraquat, le 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) et la 6-hydroxydopamine (6-OHDA) étant les plus fréquemment utilisées (Prou & Przedborski, 2005). Les études lésionnelles, en plus de modéliser l’étiologie de certains symptômes, permettent de mieux comprendre le rôle des mécanismes de compensation dans la maladie chez l’homme. En effet, plusieurs changements surviennent tant chez le parkinsonien que chez l’animal subissant une lésion des neurones de la SN, dont l’induction d’une hypersensibilité striatale médiée par la surexpression des récepteurs postsynaptiques D2, la diminution de l’expression du transporteur de la dopamine (DAT) chez les neurones dopaminergiques, l’augmentation de la synthèse de dopamine (DA) par les neurones survivants, et le changement de phénotype d’une sous-population de neurones GABA
voie de synthèse de la DA (Greenbaum et al., 2013; Huot, Lévesque, & Parent, 2007). Par ailleurs, d’un point de vue structurel, les neurones dopaminergiques de la SN, qui projettent de façon diffuse à travers le striatum, vont, s’ils sont réduits en nombre, augmenter de façon compensatoire le nombre et la grosseur de leurs vésicules synaptiques, ainsi que multiplier les embranchements et collatéralisations de leur arborisation axonale (Finkelstein et al., 2000; Parent and Parent, 2006). Ce dernier mécanisme comporte des implications non-négligeables quant au métabolisme de ces neurones, dont l’équilibre est déjà très précaire, puisqu’une augmentation de l’arborisation axonale et du nombre de ses terminaisons augmente exponentiellement le coût bioénergétique du maintien de la machinerie cellulaire ainsi que du contrôle des taux de calcium, de DA et de radicaux libres cytoplasmiques (Bolam & Pissadaki, 2012; Pissadaki & Bolam, 2013; Surmeier, 2007). La sensibilité des neurones DA de la
substantia nigra pars compacta (SNc) à la dysfonction mitochondriale, couplée au
bourgeonnement compensatoire des neurones survivants, mène à l’hypothèse que cette compensation pourrait paradoxalement perpétuer la mort de cette population neuronale (Bolam & Pissadaki, 2012; M Parent & Parent, 2006). Par ailleurs, l’arborisation axonale des neurones de la SNc, en plus d’être beaucoup plus ramifiée et étendue que celles de la plupart des autres types de neurones, est environ dix fois plus volumineuse et effectue dix fois plus de synapses chez l’homme que chez le rat (Pissadaki & Bolam, 2013), ce qui pourrait expliquer en partie pourquoi la maladie est exclusive à l’humain.
L’objectif du présent projet était de contribuer au développement d’un modèle murin dans lequel l’arborisation axonale des neurones DA de la SNc est fortement augmentée par rapport à l’animal sauvage, et d’étudier ensuite si cette caractéristique augmente la vulnérabilité de ces neurones in vivo. Pour ce faire, la capacité de croissance compensatoire des neurones de la SNc suivant une lésion précoce à la 6-OHDA a été mise à profit. Dans un premier temps, les aspects principaux du système dopaminergique, de la maladie de Parkinson et de ses modèles seront abordés. Dans un second temps seront présentés l'hypothèse de recherche, les principes expérimentaux et la méthode utilisés pour développer le modèle, ainsi que les principaux résultats obtenus quant à sa caractérisation. Dans un troisième temps, ces résultats seront commentés et des perspectives futures seront discutées par rapport à la suite du développement de ce modèle.
Le système dopaminergique
En 1957, Arvid Carlsson et son équipe démontraient que la dopamine, jusqu'alors considérée comme un simple précurseur de la noradrénaline, pouvait agir en tant que neurotransmetteur à part entière dans le système nerveux central (Björklund & Dunnett, 2007). Cette découverte ouvrit la voie à une série de découvertes, dévoilant tour à tour l'existence de groupes de neurones dopaminergiques formant des réseaux de connections distinctes, leur rôle dans les fonctions physiologiques du mouvement et de la récompense, et enfin leur implication dans les processus pathologiques tels que la maladie de Parkinson. Depuis, la littérature sur le système dopaminergique, ses fonctions et les pathologies qui y sont liées est un des domaines les plus prolifiques en neurosciences, ayant contribué grandement à la compréhension des mécanismes qui sous-tendent des fonctions aussi diverses que l'initiation du mouvement, la motivation ou la dépendance. Nous présenterons dans cette section l'anatomie du système mésencéphalique dopaminergique ainsi que les aspects d'anatomie microscopique et neurochimiques relatifs aux neurones qui le composent.
Les études histochimiques de la répartition des neurones produisant la DA chez l'humain, le primate et le rongeur classifient ces neurones en groupes étiquetés A8 à A16, les neurones noradrénergiques et sérotoninergiques composant par ailleurs les groupes A1 à A7. Les groupes A8, A9 et A10 regroupent l'ensemble des corps cellulaires dopaminergiques du mésencéphale (Zaborszky & Vadasz, 2001). Parmi ceux-ci, nous nous attarderons aux groupes A9 et A10, soit les neurones dopaminergiques de la substantia nigra et de l'aire tegmentaire ventrale, qui sont les deux groupes les plus importants en nombre du système dopaminergique et ceux impliqués directement dans la pathogénèse de la maladie de Parkinson. Le groupe A8, ou champ rétrorubral (RRF), semble être impliqué dans des fonctions similaires au groupe A10 (Waraczynski & Perkins, 2000) et présente des projections à l'hippocampe (Gasbarri et al., 1996), mais demeure relativement peu étudié. Au niveau du diencéphale, les neurones dopaminergiques sont répartis entre les groupes A11 à A15, situés dans certains noyaux de
bulbe olfactif. Enfin, un groupe dit télencéphalique désigne des populations disparates de neurones à phénotype dopaminergique, réparties à travers diverses structures du télencéphale (substantia innominata, bande diagonale de Broca, tubercule olfactif, aire prépiriforme, striatum, claustrum, couches corticales profondes du lobe frontal, capsule externe, capsule extrême et matière blanche frontale) (Benavides-Piccione & Defelipe, 2007).
Aspects neuroanatomiques
Nous présenterons ici la structure neuroanatomique macroscopique de la SNc et de la VTA, structures où se retrouvent les soma des groupes A9 et A10 respectivement, en les comparant chez l'homme et chez la souris. Les afférences et efférences des neurones DA de la SNc et de la VTA ainsi que les trois principales voies dopaminergiques mésencéphaliques seront ensuite décrites telles que retrouvées chez la souris. L'anatomie macroscopique et microscopique du striatum, structure innervée par les neurones dopaminergiques de la SNc et de la VTA, sera ensuite présentée, en comparant encore une fois la macroanatomie humaine et murine.
Les neurones dopaminergiques de la SNc et la VTA
Anatomie macroscopique
Chez l'humain comme chez le rongeur, la subdivision des structures dopaminergiques du mésencéphale est globalement préservée. Toutefois, les frontières exactes des groupes A8, A9 et A10, ainsi que leur subdivision en sous-noyaux, est plus difficile à définir (Franklin & Paxinos, 2008; Paxinos, Halliday, Watson, Koutcherov, & Wang, 2006; Zaborszky & Vadasz, 2001). La figure 1 présente la subdivision macroscopique de la SNc et de la VTA chez l'humain (homo sapiens) et chez la souris (mus musculus).
Figure 1. Anatomie des groupements dopaminergiques A9 et A10 chez l'humain et la souris
(Figure 1) Planches neuroanatomiques comparant la localisation des groupements cellulaires dopaminergiques A8, A9 et A10 chez l'humain et chez la souris (les coupes ne sont pas à l'échelle). A. Coupe coronale du cerveau humain illustrant l'emplacement de la SNc et de la VTA. B. Coupe axiale d'un hémi-mésencéphale humain illustrant la répartition des neurones dopaminergiques entre les groupements A8 (RRF, en bleu), A9 (SNc, en jaune) et A10 (VTA, en rouge). C.Vue inférieure d'une coupe axiale du mésencéphale humain (lobe frontal vers le bas de l'image), sur laquelle les structures dopaminergiques sont visibles en bleu-gris grâce aux dépôts de neuromélanine. D. Coupe coronale du cerveau murin illustrant l'emplacement de la SNc et de la VTA. E. Reconstruction tridimentionnelle illustrant la position relative des groupements A8 (bleu), A9 (jaune) et A10 (rouge) à l'intérieur du mésencéphale murin. Figure créée à partir d'images tirées de (German & Manaye, 1993; Mendez et al., 2005; R. Williams, 2000; T. H. Williams, Gluhbegovic, & Jew, 1997).
En outre, des immunomarquages contres certaines protéines ont permis de préciser une ségrégation histochimique de ces neurones en fonction de certaines caractéristiques. Par exemple, chez l'humain, les neurones dopaminergiques exprimant la calbindin, une protéine liant le calcium intracellulaire, sont surtout situés en médiodorsal des groupes A8, A9 et A10, et ne sont pas retrouvés dans la SNcv (SNc pars ventralis), (Mendez et al., 2005). Par ailleurs, de façon intéressante, l'expression de cette protéine semble corrélée à une vulnérabilité réduite dans la maladie de Parkinson et ses modèles animaux (Dopeso-Reyes et al., 2014). À l'inverse, les neurones dopaminergiques exprimant la protéine Girk2 (canal potassique à rectification entrante activé par les protéines G, type 2) sont surtout retrouvés dans la SNcv (Reyes et al., 2012). Enfin, bien que des sous-groupes à l'intérieur de la SNc et de la VTA aient plus récemment été identifiés en fonction de leurs projections respectives, de caractéristiques protéomiques ou électrophysiologiques précises (Roeper, 2013), les subdivisions neuroanatomiques classiques, basées sur l'immunoréactivité contre la TH demeurent encore la norme dans la littérature actuelle.
Afférences des neurones dopaminergiques de la SNc et de la VTA
Les neurones dopaminergiques de la SNc et de la VTA recoivent au niveau de leur arborisation somatodendritique une innervation riche et extrêmement diversifiée. Ces afférences proviennent de structures aux fonctions très diversifiées, ce qui souligne l'importance de la SNc et de la VTA comme points de convergence et d'intégration de signal. Watabe-Uchida et al. ont récemment fourni une description très détaillée de ces afférences (Watabe-Uchida, Zhu, Ogawa, Vamanrao, & Uchida, 2012) en utilisant une nouvelle méthode de tracage basée sur une transfection virale rétrograde sélective, corroborant essentiellement les résultats obtenus par le passé avec les traceurs rétrogrades traditionnels, mais présentant l'avantage de ne pas marquer les afférences des neurones non-dopaminergiques, tel qu'illustré à la figure 2.
Le striatum et le pallidum sont les structures contenant le plus grand nombre de neurones afférents tant vers la VTA que la SNc. Dans le striatum, il existe une innervation préférentielle
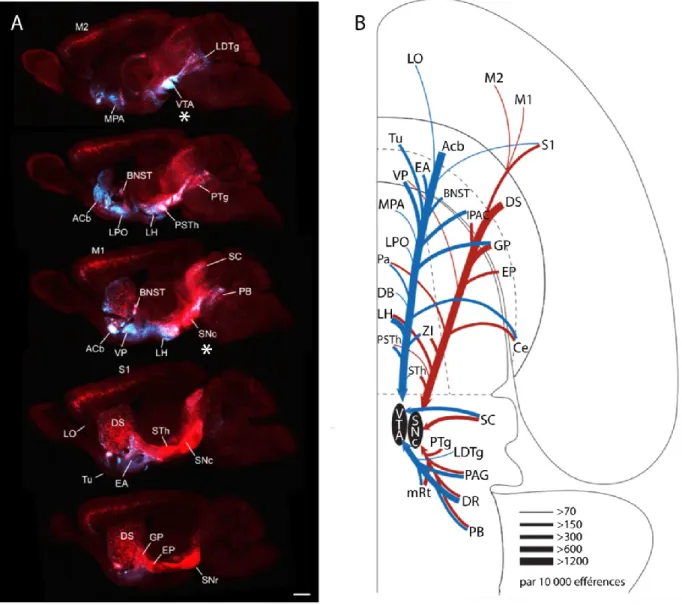
Figure 2. Afférences de la SNc et de la VTA chez la souris
(Figure 2) Illustration des afférences de la SNc et de la VTA. A. Coupes parasagittales d'un cerveau de souris illustrant la distribution des afférences monosynaptiques de la VTA (en bleu) et de la SNc (en rouge), tel que révélée par l'utilisation d'un virus de la rage modifié (SADΔG-GFP(EnvA) encodant le gène de la GFP et ne transfectant de façon transsynaptique que les neurones innervant la VTA ou la SNc (Watabe-Uchida et al., 2012). B. Schéma horizontal des afférences les plus importantes vers la VTA (en bleu) et la SNc (en rouge). LO: cortex latéral orbitaire, M1: cortex moteur primaire, M2: cortex moteur secondaire, S1: cortex somatosensoriel primaire, Tu: tubercule olfactif, Acb: noyau accumbens, DS: striatum dorsal, Ce: noyau central de l'amygdale, VP: pallidum ventral, EA: amygdale étendue, BNST: noyau de la strie terminale, IPAC: noyau interstitiel de la branche postérieure de la commissure
diagonale de Broca, LH: aire latérale hypothalamique, Zi: zona incerta, PSTh: noyau para-sous-thalamique, STh: noyau sous-para-sous-thalamique, SC: colliculus supérieur, PTg: noyau tegmentaire pédonculopontin, LDTg: noyau tegmentaire latérodorsal, PAG: substance grise périacqueducale, mRt: noyau de la commissure postérieure, DR: noyau dorsal du raphé, PB: noyau parabrachial. Figure adaptée de (Watabe-Uchida et al., 2012).
de la SNc par la partie dorsale du striatum, et une innervation préférentielle de la VTA par le noyau accumbens et le tubercule olfactif. Au niveau du pallidum, les structures davantage médiales (pallidum ventral, amygdale étendue), projettent surtout vers la VTA alors que les structures plus latérales (globus pallidus (GP), noyau endopédonculaire) innervent la SNc de façon prédominante. Au niveau de l'hypothalamus, la LH semble avoir une innervation préférentielle de la VTA alors que le STh semble innerver de façon préférentielle la SNc, et une ségrégation spatiale semble par ailleurs gouverner l'innervation hypothalamique globale de la SNc et de la VTA, avec un gradient dorsolatéral en faveur de la SNc. Enfin, au niveau de l'amygdale, seul le noyau central semble fournir des afférences, tout comme le noyau de la strie terminale (BNST), et ce tant vers la SNc que la VTA (Watabe-Uchida et al., 2012; Yetnikoff, Lavezzi, Reichard, & Zahm, 2014).
Au niveau du cortex, ces auteurs rapportent des afférences diffuses et provenant surtout dans l'hémicortex rostral, comprenant les cortex moteur, somatosensoriel, médial préfrontal et orbitofrontal. Pour la SNc, l'afférence corticale la plus dense provient de M1 et de M2, et la plus importante de S1. La VTA reçoit moins d'afférences corticales que la SNc, et celles-ci proviennent surtout du cortex orbitofrontal latéral (LO). Enfin, très peu d'afférences semblent probenir de l'hémicortex caudal (cortex pariétal, visuel, auditif, entorhinal).
Au niveau du mésencéphale et du rhombencéphale, le raphé dorsal (DR) semble contenir la population neuronale la plus dense innervant les deux structures (avec une préférence pour la VTA), le colliculus supérieur (SC), la substance grise périaqueducale (PAG), le noyau parabrachial et certains noyaux cérébelleux fournissent aussi des afférences à la SNc et la VTA. Par ailleurs, la SNc est innervée par le noyau pédonculotegmentaire (PTg) et le noyau
cunéiforme (CnF), alors que la VTA est innervée par le noyau tegmentaire latérodorsal. Contrairement à ce qui avait été avancé antérieurement, les afférences du septum et de l'habenula médiale vers la SNc ou la VTA n'en innervent que les neurones non-dopaminergiques (Watabe-Uchida et al., 2012; Yetnikoff et al., 2014).
Efférences dopaminergiques de la SNc et de la VTA
Les neurones dopaminergiques de la SNc et de la VTA envoient des prolongements axonaux dans plusieurs structures mésencéphaliques, diencéphaliques et télencéphaliques, traditionnellement regroupées en trois voies principales empruntant dans leurs parcours un tractus commun, le faisceau télencéphalique médian (MFB). La figure 3 illustre la répartition des neurones DA mésencéphaliques de chacune des voies et leurs aires de projection.
La voie mésostriée est composée de neurones du groupe A9 qui projettent au striatum dorsal (dSTR), préférentiellement dans son compartiment striosomal, ainsi que de neurones du groupe A10 qui projettent dans le striatum ventral (vSTR) (figure 3A). C'est cette voie, surtout dans son aspect dorsolatéral, qui est impliquée dans la planification et l'initiation motrices, et qui est atteinte dans la maladie de Parkinson (Bjorklund & Dunnett, 2007; Ikemoto, 2007; Yetnikoff et al., 2014).
La voie mésolimbique désigne les projections dopaminergiques innervant le vSTR (noyau accumbens), le septum et l'amygdale, provenant surtout de la partie latérale de la VTA ainsi que de la RRF, mais aussi de la SNcl (pars lateralis) (figure 3B). L'innervation dopaminergique de ces structures affecte notamment la perception subjective de plaisir, la motivation pour les stimuli de récompense, et l'apprentissage moteur relié à ces stimuli, et est la voie neuronale principale à travers laquelle sont médiés les phénomènes de dépendance (Bjorklund & Dunnett, 2007; Ikemoto, 2007; Yetnikoff et al., 2014).
Figure 3. Voies efférentes dopaminergiques de la SNc et de la VTA
(Figure 3) Illustration des voies efférentes dopaminergiques de la SNc et de la VTA. A. La voie mésostriée comprend les projections provenant de la SNc vers le dSTR (jaune) et de la VTA vers le noyau accumbens (vert). B. La voie mésolimbique comprend les projections de la VTA vers le tubercule olfactif et le septum, et les projections de la VTA et de la SNcl vers l'amygdale. C. La voie mésocorticale comprend les projections de la VTA vers le cortex préfrontal et le cortex périrhinal et les projections de la VTA et de la SNc vers le cortex périrhinal. Toutes les voies dopaminergiques illustrées empruntent un tractus commun, le MFB (non-illustré). Figure adaptée de (Bjorklund & Dunnett, 2007).
La voie mésocorticale est composée de projections provenant de neurones répartis de façon diffuse dans les aires A8, A9 et A10, et est beaucoup plus développée chez le primate que chez le rongeur. En effet, chez le rongeur, l'innervation dopaminergique se restreint surtout aux cortex frontal, cingulaire et entorhinal (figure 3C), alors que chez le primate, ce territoire s'étend à tout le manteau cortical (Bjorklund & Dunnett, 2007). Elle est impliquée dans le fonctionnement cognitif normal, la motivation et la réponse émotionnelle, et son dysfonctionnement est mis en cause dans l'aboulie, la psychose et la schizophrénie, entre autres.
Il est à noter, toutefois, que cette taxonomie des projections dopaminergiques de la SNc et de la VTA masque le chevauchement réel qui existe au niveau de l'innervation respective de leurs structures cibles, bien qu'il existe un gradient préférentiel d'innervation des structures latérales par la SNc et médiales par la VTA. Ces trois voies existent bien de facon distincte, mais proviennent de neurones qui ne sont pas ségrégués de façon absolue selon les frontières des structures mésencéphaliques. Par ailleurs, en séparant les neurones des aires A9 et A10 en niveau dorsal (exprimant la calbindin) et ventral (exprimant Girk2), on remarque que les neurones du niveau dorsal innervent les aires limbiques et corticales, le vSTR et le compartiment matriciel du dSTR, alors que les neurones du niveau ventral innervent presque exclusivement le compartiment striosomal du striatum (Damier, Hirsch, Agid, & Graybiel, 1999a; Dopeso-Reyes et al., 2014; Reyes et al., 2012). Enfin, la présence d'une certaine innervation dopaminergique du GPi, du GPe, du noyau sous-thalamique et de la SNr a été mise en évidence (Bjorklund & Dunnett, 2007). Parent et al. ont illustré de façon probante que de telles innervations pouvaient être réalisées par des neurones possédant des terminaisons en passant dans le GP ou les noyaux thalamiques, en chemin vers leur champ d'innervation terminal au niveau du dSTR, et mis en lumière la diversité des patrons d'innervation retrouvés chez les neurones dopaminergiques du mésencéphale (Prensa & Parent, 2001).
Le striatum
Anatomie macroscopique
Le striatum, structure innervée par les prolongements axonaux des neurones dopaminergiques de la voie mésostriée, est au cœur de la compréhension des symptômes de la maladie de Parkinson. Il importe de mentionner que ses composantes possèdent une nomenclature et une anatomie différente chez chez le rongeur et chez le primate. En effet, chez le rat et la souris, la littérature divise le striatum en parties dorsale (dSTR) et ventrale (vSTR), cette dernière structure contenant le noyau accumbens (nAc) segmenté en cœur (nAc core) et en coquille (nAc shell) et le tubercule olfactif (Tu). Chez le primate, le dSTR est composé de
retrouve ventromédialement à la tête (partie la plus antérieure) du noyau caudé. La figure 4 compare l'anatomie macroscopique de ces deux structures chez l'humain et chez la souris.
Figure 4. Anatomie du striatum humain et murin
(Figure 4) Planches neuroanatomiques comparant la localisation et la composition du striatum chez l'humain et la souris (les coupes ne sont pas à l'échelle). A. Coupe coronale du cerveau humain illustrant l'emplacement du dSTR, composé du noyau caudé (en rouge) et du putamen (en jaune), et du vSTR (noyau accumbens, en vert). B, C. Emplacement et structure tridimentionnelle du noyau caudé, du putamen, du noyau accumbens et du globus pallidus (en bleu, composé des globus pallidus interne et externe). D. Coupe coronale du cerveau murin illustrant l'emplacement du dSTR (en jaune) et du vSTR (en vert). E. Reconstruction tridimentionnelle illustrant la position et la structure du dSTR et du vSTR chez la souris. Figure créée à partir d'images tirées de (Hjornevik et al., 2007; Mallet, Moine, Ballion, Le Moine, & Gonon, 2006; Voelker, 2004; T. H. Williams et al., 1997).
Anatomie microscopique, afférences et efférences striatales
Le striatum contient une population neuronale hétérogène, composée de neurones moyens épineux (medium spiny neurons (MSN), également appelés spiny projection neurons), d'interneurones cholinergiques, et d'interneurones GABAergiques. Les MSN sont des neurones GABAergiques qui représentent environ 95% des neurones du striatum humain, et sont divisés en deux sous-populations, l'une exprimant le récepteur D1 et faisant partie de la voie de projection striatonigrale (dite directe), et l'autre exprimant le récepteur D2 et faisant partie de la voie de projection striatopallidale (dite indirecte). Bien qu'il existe des évidences d'une co-expression des récepteurs D1 et D2 chez un sous-groupe restreint de MSN durant le développement embryonnaire, la ségrégation des deux populations s'établit rapidement et est maintenue chez l'animal adulte (Biezonski, Trifilieff, Meszaros, Javitch, & Kellendonk, 2015; Calabresi, Picconi, Tozzi, Ghiglieri, & Di Filippo, 2014; Thibault, Loustalot, Fortin, Bourque, & Trudeau, 2013). Les propriétés de ces neurones, dont les projections axonales constituent l'efférence striatale principale, seront détaillées plus loin.
En outre, les interneurones cholinergiques, de façon générale, sont responsables de médier les interactions entre les deux sous-groupes de MSN, en inhibant l'activité endocannabinoïde de ceux-ci et modulant donc à la baisse la dépression synaptique à long terme (LTD). Par ailleurs, l'activité tonique des interneurones cholinergiques, régulée positivement par la présence de dopamine, facilite ainsi la neurotransmission glutamatergique corticostriatale (Calabresi et al., 2014). Les interneurones cholinergiques agissent également en amplifiant la libération de DA via l'activation de récepteurs nicotiniques terminaux (Exley & Cragg, 2008).
Enfin, les interneurones GABAergiques striataux sont divisés en trois sous-classes: les neurones à décharge rapide exprimant la parvalbumine, les neurones exprimant la calrétinine, et les neurones exprimant la NO synthase et produisant du monoxyde d'azote, impliqué dans la LTD des MSN. La figure 6 de la prochaine section reprend de façon graphique la composition
de la base (figure 6A). Par ailleurs, le vSTR est divisé en deux sous-structures, le cœur (core) et la coquille (shell), dont les populations de MSN diffèrent quant à la densité de leurs épines, la complexité de leurs arborisations et leurs projections. Ces structures sont toutes deux impliquées dans les circuits de la récompense, mais diffèrent quant à leurs afférences et à leurs rôles comportementaux précis (Ito & Hayen, 2011), caractéristiques qui ne seront pas détaillées ici.
Les principales afférences du striatum sont les projections corticostriées glutamatergiques faisant synapse sur l'arborisation dendritique des MSN, responsables de l'excitation de ceux-ci (figure 6C). Des afférences glutamatergiques striatales proviennent aussi du complexe parafasciculaire centromédian du thalamus et du noyau sous-thalamique, et le GPe fournit des afférences GABAergiques. Par ailleurs, le striatum reçoit une innervation dopaminergique des groupes A9 et A10, dont le patron d'activité est à la fois tonique et phasique (Calabresi et al., 2014). L'activité électrophysiologique tonique assure des niveaux basaux de DA, alors que l'activation phasique du système mésostrié se fait sous forme de décharges en bouffées, sous l'influence des afférences glutamatergiques nigrales des noyaux pédonculopontin (PPN) et sous-thalamique (STN) (Shimo & Wichmann, 2009), et permet une modulation localisée des niveaux de DA. De plus, une sous-population de neurones DA de la VTA présentant une capacité de cotransmission glutamatergique fournit une innervation glutamatergique dans le vSTR (Chuhma, Mingote, Moore, & Rayport, 2014; L.-É. Trudeau et al., 2014). Enfin, il est à noter que l'on distingue traditionnellement deux compartiments neurochimiques à l'intérieur du striatum, qui ont d'abord été identifiés par immunomarquage contre l'acétylcholinestérase (Graybiel & Ragsdale, 1978), soit le compartiment striosomal et le compartiment matriciel, et ont ensuite été caractérisés selon leur contenu en récepteurs opioïdes et en calbindin (Damier et al., 1999a). Le profil neurochimique, les afférences préférentielles et le rôle précis de ces compartimentalisations sont encore l'objet de recherches actives.
Aspects moléculaires
La DA et ses voies de synthèse et de dégradation
La DA est une catécholamine qui fait aussi partie de la voie de synthèse de la noradrénaline. Elle est synthétisée par le neurone dopaminergique en transformant d'abord l'acide aminé tyrosine en 3,4-dihydroxyphénylalanine (L-DOPA), catalysée par l'enzyme TH, ce qui constitue l'étape limitante de la voie de synthèse (Laatikainen, Sharp, Harrison, & Tunbridge, 2013; Levitt, Spector, Sjoerdsma, & Udenfriend, 1965). La L-DOPA est ensuite transformée en DA par la dopamine décarboxylase (DDC), également appelée décarboxylase des acides aminés aromatiques. Elle peut par ailleurs être catabolisée par l'action conjointe des enzymes monoamine oxydase (MAO), de la catéchol-O-méthyltransférase (COMT), et de l'aldéhyde déshydrogénase (AH), selon deux voies possibles. Dans la première, la COMT introduit un groupement méthyl sur la DA pour générer la 3-méthoxytyramine (3-MT), elle-même oxydée par la MAO située sur la membrane externe des mitochondries pour former la 3-méthoxy-4-hydroxyphénylacétaldéhyde (MHPA), à son tour oxydée par l'AH pour former de l'acide homovanilique (HVA). Dans la seconde, la MAO oxyde la DA pour former la dihydrophénylacétaldéhyde (DHPA), elle-même oxydée par l'AH pour former l'acide 3,4-dihydroxyphénylacétique (DOPAC), sur lequel la COMT catalyse l'ajout d'un groupement méthyl pour générer l'HVA (Meiser, Weindl, & Hiller, 2013). Par ailleurs, dans les neurones noradrénergiques, l'enzyme dopamine -hydroxylase permet la conversion de la DA en noradrénaline.
La terminaison dopaminergique
Une fois synthétisée, la DA est emmagasinée dans des vésicules synaptiques via le transporteur vésiculaire des monoamines de type 2 (VMAT2), présent à la surface des vésicules.
survient lors de l'entrée massive de l'ion Ca++ dans la cellule secondaire à l'arrivée d'un potentiel
d'action membranaire. Par ailleurs, l'innervation dopaminergique s'effectue à la fois au moyen de varicosités dopaminergiques synaptiques et asynaptiques, c'est-à-dire dépourvues de spécialisations membranaires permettant l'établissement d'une synapse. Les terminaisons dopaminergiques synaptiques sont associées à la présence du transporteur vésiculaire du glutamate de type 2 (VGLUT2), ce qui leur conférerait la propriété de pouvoir co-libérer du glutamate (Descarries et al., 2008; El Mestikawy, Wallén-Mackenzie, Fortin, Descarries, & Trudeau, 2011; L.-E. Trudeau & Gutiérrez, 2007). De façon générale, il a été rapporté qu'environ 60-70% des terminaisons dopaminergiques striatales étaient asynaptiques et que les 30-40% restants étaient de type synaptique (Descarries, Watkins, Garcia, Bosler, & Doucet, 1996; Freund, Powell, & Smith, 1984).
Suite à sa relâche par exocytose, dans la fosse synaptique s'il s'agit d'une terminaison synaptique ou dans le milieu extracellulaire s'il s'agit d'une terminaison asynaptique, la DA est libre d'agir sur ses cibles, les récepteurs dopaminergiques, qui seront décrits plus loin. L'activité de la DA est principalement interrompue par sa recapture au moyen d'un transporteur membranaire spécifique, le DAT, qui est uniformément réparti sur la membrane neuronale des neurones DA (Nirenberg et al., 1997). Le DAT permet donc la clairance extracellulaire et le recyclage de la DA par les neurones dopaminergiques.
Les récepteurs de la dopamine
La DA agit sur les récepteurs dopaminergiques, pouvant être situés sur les cellules DA, et sont alors qualifiés d'autorécepteurs, ou être situés sur d'autres cellules, agissant alors à titre d'hétérorécepteurs. Il en existe cinq sous-types (étiquetés D1 à D5), et ils sont séparés en deux groupes selon leur type de couplage aux protéines G: le groupe D1 (récepteurs D1 et D5) et le groupe D2 (récepteurs D2, D3 et D4) (Beaulieu, Espinoza, & Gainetdinov, 2015). Les récepteurs du groupe D1 sont des récepteurs métabotropes couplés à la protéine Gs/olf, qui activent
l'adénylate cyclase et induisent la production d'adénosine monophosphate cyclique (AMPc). Les récepteurs D1 se retrouvent dans le striatum préférentiellement sur les neurones épineux moyens (MSN) de la voie directe. On les retrouve aussi au niveau des autres structures cibles des voies dopaminergiques mésencéphaliques, plus précisément dans le tubercule olfactif, l'hypothalamus, le thalamus et les structures limbiques (Beaulieu et al., 2015). Les récepteurs D2, quant à eux, sont couplés aux protéines Gi/o, et inhibent la production d'AMPc. Ils se
retrouvent en tant qu'hétérorécepteurs sur les neurones de la voie indirecte, et en tant qu'autorécepteurs sur les terminaisons axonales des neurones DA (Beaulieu et al., 2015; Calabresi et al., 2014). De plus, dans la SN, la DA est libérée à partir du compartiment dendritique des neurones dopaminergiques, ce qui permettrait l'activation d'autorécepteurs et l'autorégulation de leur activité (Bjorklund & Dunnett, 2007). La prochaine section présente une description de la maladie de Parkinson et des bases neuronales de sa symptomatologie, et détaille du même fait les voies directe et indirecte du mouvement ci-haut mentionnées, en plus de d'illustrer les interactions neuronales physiologiques et pathologiques au niveau du striatum (figure 6).
La maladie de Parkinson
La maladie de Parkinson a été décrite pour la première fois en 1817 par James Parkinson, qui la désignait alors sous le nom de paralysis agitans. Par la suite, Jean-Martin Charcot et son collègue Leopold Ordenstein ont repris et précisé la définition et la symptomatologie de la maladie dans leur thèse Sur la paralysie agitante et la sclérose en plaques généralisée, publiée en 1868. Dans cette publication, les auteurs en décrivaient avec justesse les principaux symtômes, mais considéraient la maladie comme une névrose, n'évoquant pas la possibilité d'une étiologie pathologique organique sous-jacente. Il faudra plus de deux décades avant que Jean-Baptiste Charcot, Georges Marinesco et Paul Oscar Blocq, poursuivant la foulée des recherches de Jean-Martin Charcot pour identifier la cause de la maladie, en arrivent à suspecter l'implication de la SN dans la maladie, postulat officiellement formulé pour la première fois en 1893 lors d'une conférence de Blocq à la Salpêtrière (Martin Parent & Parent, 2010). Dans la présente section, nous aborderons d'abord la symptomatologie, la pathophysiologie et les stratégies de traitement de la maladie, avant de présenter les différents modèles animaux utilisés pour la modéliser. Bien que la cause des symptômes de parkinsonisme ait été identifiée depuis le début du 20e siècle comme étant la dysfonction du circuit mésostrié secondairement à une
dégénérescence des neurones dopaminergiques mésencéphaliques (Martin Parent & Parent, 2010), l'étiologie précise à l'origine de cette neurodégénérescence dans le cadre de la maladie de Parkinson demeure obscure dans la grande majorité des cas, malgré l'identification graduelle de divers éléments pathophysiologiques potentiellement contributifs.
Symptomatologie de la maladie de Parkinson
La maladie de Parkinson se manifeste sur le plan clinique par quatre symptômes locomoteurs cardinaux, soit la rigidité, le tremblement, la bradykinésie et l’instabilité posturale, qui peuvent être associés à un grand nombre d'autres symptômes, qui seront évoqués plus loin.
Symptômes cardinaux
D'abord, la rigidité consiste en une raideur des membres et une résistance au mouvement causée par l’augmentation du tonus musculaire associée à une contraction excessive et discordante. Cette rigidité est fréquemment accompagnée de douleurs musculaires résultant de la contraction aberrante de groupes musculaires antagonistes. Les tremblements, d'une fréquence de 4 à 6 hertz, affectent davantage les groupes musculaires distaux que proximaux, présentent un patron d'apparition asymétrique (un hémicorps étant atteint de façon plus importante que l'autre) et sont surtout présents au repos, s'estompant lors de mouvements volontaires et durant le sommeil. Le signe classique du pill-rolling, caractéristique de la maladie, est une manifestation précoce de tremblement de repos (Jankovic, 2008). Par ailleurs, la rigidité et le tremblement de repos contribuent à engendrer le signe de la «roue dentée» à la supination rapide de l'avant-bras, où est rencontrée une résistance spontanée et erratique à la mobilisation passive de l'avant-bras (Jankovic, 2008). La bradykinésie est caractérisée par des difficultés tant dans la planifiction que l’amorce et l’exécution des mouvements, surtout lorsque ceux-ci sont séquentiels ou simultanés, résultant en une lenteur dans les activités de la vie quotidienne et une augmentation du temps de réaction. La bradykinésie entraîne à son tour une série de symptômes tels que la perte de la gestuelle spontanée, l'hypersialorrhée par atteinte de la déglutition, la dysarthrie, la perte d'expression faciale et la perte du balancement normal des bras à la marche (Jankovic, 2008).
De façon intéressante, l'état émotionnel plus particulièrement l'anticipation d'une contrainte temporelle imminente peuvent mener au pénomène de kinesia paradoxica, grâce auquel le patient parkinsonien peut temporairement échapper à la bradykinésie (Distler, Schlachetzki, Kohl, Winkler, & Schenk, 2016). Enfin, l'instabilité posturale, causée par une perte des réflexes posturaux, survient habituellement plus tardivement dans la maladie, et est un facteur de risque de chute non-négligeable (Jankovic, 2008; Tolosa, Wenning, & Poewe, 2006).
Autres symptômes moteurs
Les blocs moteurs, ou épisodes de «freezing», surviennent chez environ 50% des patients, et sont caractérisés par des périodes d'akinésie de quelques secondes, transitoires, surtout lors de la marche. Au niveau de la démarche, la combinaison de la rigidité et de l'instabilité posturale contribue à créer un patron de marche dit de festination, caractérisé par des pas de faible amplitude et une diminution du déroulement plantaire. Par ailleurs, des déformations posturales secondaires à la rigidité sont aussi fréquentes, tant au niveau tronculaire (position de flexion antérieure du tronc secondaire à la rigidité du cou et du tronc) qu'au niveau des membres (déformations dites «striatales» de la main, du pied ou des orteils, coudes et genoux en flexion), et survenant plus tard dans la maladie (Jankovic, 2008). La micrographie est aussi un signe caractéristique d'une atteinte motrice de type parkinsonienne.
Des signes d'atteinte bulbaire, tels que la dysarthrie, l'hypophonie et la dysphagie, s'ajoutent souvent au tableau moteur, et découlent probablement d'une bradykinésie et d'une rigidité atteignant les muscles orofaciolaryngés. Enfin, des signes neuro-ophtalmologiques ne sont pas rares, et incluent une diminution de la fréquence de clignement, une xerophtalmie, une atteinte de la poursuite oculaire et une apraxie de l'ouverture des paupières, entre autres (Jankovic, 2008).
Symptômes sensitifs, végétatifs et cognitifs
Sur le plan sensitif, plusieurs études ont démontré qu'une hyposmie était présente chez près de 90% des patients, et ce précocément au cours de l'installation de la maladie (Tolosa et al., 2006). Par ailleurs, plusieurs signes de dysfonction végétative sont souvent retrouvés, et incluent l'hypotension orthostatique, la sudation excessive, la dysfonction sexuelle et les troubles sphinctériens (Jankovic, 2008). De plus, au niveau gastrointestinal, le reflux gastro-oesophagien, la gastroparésie et la constipation sont des problèmes fréquents, s'ajoutant aux
difficultés nutritionnelles reliées à la dysphagie (Barichella, Cereda, & Pezzoli, 2009). Au niveau du sommeil, une perturbation du stade REM (rapid eye movement), caractérisée par une augmentation des rêves violents et une perte de l'atonie physiologique, est fréquemment et précocément retrouvée, dysfonction à laquelle peut s'ajouter une somnolence diurne (Tolosa et al., 2006).
Enfin, les fonctions cognitives sont elles aussi souvent affectées par la maladie de Parkinson, tant au point de vue des fonctions exécutives que du langage, de la mémoire et des aptitudes visuospatiales. De plus, une atteinte de la capacité de planification, de la flexibilité cognitive, de la pensée abstraite et du choix d'actions appropriées est souvent présente, tout comme la fluctuation de l'attention et le ralentissement psychique. Un tableau dépressif associé n'est pas rare. Dans 25 à 30% des cas, les troubles cognitifs légers évoluent graduellement vers une démence, l'atteinte exécutive prédominant habituellement l'atteinte mnésique (Caballol, Martí, & Tolosa, 2007). La démence à corps de Lewy, classiquement définie comme entité clinique distincte de la maladie de Parkinson, est de plus en plus considérée comme une présentation clinique alternative de la même maladie, avec des observations neuropathologiques similaires atteignant toutefois davantage le lobe temporal, et un tableau clinique de démence précoce et fluctuante, et d'hallucinations visuelles (Caballol et al., 2007; Tolosa et al., 2006).
Évolution clinique
Les symptômes végétatifs, l'anosmie et la perturbation du sommeil sont souvent les premières manifestations de la maladie, des années avant l’apparition des symptômes moteurs et l’établissement provisoire du diagnostic, par ailleurs seulement confirmé avec certitude au moment de l'autopsie. L'installation de la maladie est presque toujours asymétrique, ce qui reflète les études en imagerie nucléaire de la déplétion dopaminergique striatale, elle aussi asymétrique. Suit l'apparition d'une bradykinésie progressive, d'une rigidité et de troubles de la
l'instabilité posturale et la dystonie arrivent habituellement plus tard dans le cours de la maladie, et mettent le patient à risque augmenté de chutes. Par ailleurs, le traitement à la L-DOPA entraîne des fluctuations motrices et des dyskinésies apparaissant chez une proportion significative des patients traités. Des comportements de dépendance, d'abus de médicaments, d'hyperphagisme, d'hypersexualité et de jeu pathologique, peuvent aussi apparaître en cours de traitement, surtout lorsque des agonistes dopaminergiques sont utilisés (Jankovic, 2005). Enfin, une composante de dysfonction respiratoire chronique (obstructive par rigidité du cou ou restrictive par rigidité de la paroi thoracique, est associée à un risque plus élevé de pneumonies, facteur significatif de morbidité et de mortalité (Jankovic, 2008).
Diagnostic différentiel des syndromes parkinsoniens
Il est à noter que plusieurs pathologies différentes de la maladie de Parkinson peuvent présenter un tableau de parkinsonisme chevauchant plus ou moins la symptomatologie décrite ci-haut, la paralysie supranucléaire progressive, la dégénération corticobasale et l'atrophie multisystémique en étant des exemples (Tolosa et al., 2006). De plus, un syndrome parkinsonien peut être causé par des causes secondaires identifiables, notamment pharmacologiques (prise d'agents à action antidopaminergique tels que les antipsychotiques), infectieuses, toxiques (MPTP, manganèse, cyanure), mécanique (par compression tumorale ou malformative des voies dopaminergiques), métaboliques (maladie de Wilson, hypoxie, myélinolyse extrapontine), ou congénitales (syndrome d'hémiatrophie-hémiparkinsonisme), pour n'en évoquer que quelques causes (Tolosa et al., 2006). Pour conclure, la confirmation du diagnostic de maladie de Parkinson en présence d'un syndrome parkinsonien ne peut se faire hors de tout doute qu'à la pathologie, devant une perte des populations DA mésencéphaliques et en l'absence de cause secondaire identifiable.
Stratégies de traitement
En absence de tests de dépistage suffisament fiables et de thérapies éprouvées pouvant freiner la dégénérescence dopaminergique nigrale, le but des traitements actuellement disponibles pour les patients atteints de la maladie est d'en contrôler les symptômes le plus longtemps possible tout en minimisant les effets secondaires.
Précurseur de la dopamine
À l'heure actuelle, la première ligne de traitement des symptômes moteurs est habituellement la L-DOPA, prise en combinaison avec de la carbidopa (un inhibiteur de la dopa-décarboxylase périphérique). La L-DOPA/carbidopa est administrée en doses graduellement plus hautes, et permet habituellement un contrôle des symtômes moteurs de la maladie sur une période de 4 à 6 ans environ. Après cette période, tel que mentionné précédemment, une proportion croissante de patients (40% à 6 ans suivi de 10% supplémentaires par année) développeront des fluctuations motrices, alternant entre des périodes de bon contrôle (périodes «on»), et périodes d'exacerbation des symptômes moteurs et non-moteurs (périodes «off»), ainsi que des périodes de dyskinésie induite (Jankovic, 2008; Stocchi et al., 2010).
Inhibiteurs des enzymes de dégradation
Les inhibiteurs de la MAO-B tels que la sélégiline peuvent être considérés à un stade précoce de la maladie ou en traitement adjuvant à la L-DOPA/carbidopa à des stades plus avancés, réduisant significativement la durée des périodes «off» (Caslake et al., 2009; Rascol et al., 2005; The Parkinson Study Group, 2005). Les inhibiteurs de la COMT (entacapone, tolcapone), prolongent aussi l'effet de la L-DOPA en en inhibant la dégradation enzymatique périphérique, réduisant la durée des périodes «off», mais avec une augmentation importante des dyskinésies (Stocchi et al., 2010). En somme, les agents inhibiteurs de la MAO-B et de la COMT comportent des effets secondaires non négligeables, dont les plus importants sont l'exacerbation
des dyskinésies, des hallucinations visuelles, des troubles du sommeil et de l'hypotension (Jankovic, 2005), ce qui limite leur fenêtre thérapeutique.
Agonistes dopaminergiques
Des agonistes dopaminergiques (ropinirole, pramipexole ou rotigotine) peuvent aussi être utilisés en monothérapie précoce ou de façon adjuvante dans la maladie moyenne à avancée afin de réduire les fluctuations motrices. Par ailleurs, l'apomorphine en injection sous-cutanée peut permettre un soulagement temporaire et rapide des périodes d'akinésie transitoire chez les patients qui y font face. Toutefois, leurs effets secondaires incluent de la somnolence, des hallucinations visuelles, de la confusion, et des comportements compulsifs, en plus de dyskinésies, qui sont cependant un effet secondaire moins commun (Jankovic, 2005).
Autres traitements pharmacologiques
Enfin, d'autres agents pharmacologiques peuvent être utilisés pour traiter certains symptômes. L'amantadine, un aminoadamantane d'abord utilisé comme agent antiviral dont le mécanisme antiparkinsonien demeure méconnu, est approuvée pour le traitement adjuvant des dyskinésies et des fluctuations motrices, mais augmente le risque de déclencher des troubles du comportement compulsifs (Weintraub et al., 2010). Par ailleurs, les agents anticholinergiques ont été démontrés efficaces pour diminuer les tremblements, mais présentent un risque non-négligeable d'effets secondaires neuropsychiatriques (Katzenschlager, Sampaio, Costa, & Lees, 2002). Plusieurs autres agents pharmacologiques, qui ne seront pas abordés ici, sont utilisés pour traiter les symptômes non-moteurs de la maladie.
Stimulation cérébrale profonde
Une seconde approche de traitement chez les patients réfractaires à l'approche pharmacologique consiste en la stimulation cérébrale profonde (DBS), via l'implantation neurochirurgicale d'électrodes au niveau des ganglions de la base. À ce jour, les structures pour