HAL Id: inserm-00796724
https://www.hal.inserm.fr/inserm-00796724
Submitted on 4 Mar 2013
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
Cell therapy using tolerogenic dendritic cells in
transplantation.
Aurelie Moreau, Emilie Varey, Laurence Bouchet-Delbos, Maria-Cristina
Cuturi
To cite this version:
Aurelie Moreau, Emilie Varey, Laurence Bouchet-Delbos, Maria-Cristina Cuturi. Cell therapy using
tolerogenic dendritic cells in transplantation.. Transplant Res, 2012, 1 (1), pp.13.
�10.1186/2047-1440-1-13�. �inserm-00796724�
R E V I E W
Open Access
Cell therapy using tolerogenic dendritic cells in
transplantation
Aurélie Moreau, Emilie Varey, Laurence Bouchet-Delbos and Maria-Cristina Cuturi
*Abstract
Organ transplantation is the main alternative to the loss of vital organ function from various diseases. However, to avoid graft rejection, transplant patients are treated with immunosuppressive drugs that have adverse side effects. A new emerging approach to reduce the administration of immunosuppressive drugs is to co-treat patients with cell therapy using regulatory cells. In our laboratory, as part of a European project, we plan to test the safety of tolerogenic dendritic cell (TolDC) therapy in kidney transplant patients. In this mini-review, we provide a brief summary of the major protocols used to derive human TolDC, and then focus on the granulocyte
macrophage-TolDC generated by our own team. Proof of safety of TolDC therapy in the clinic has already been demonstrated in patients with diabetes. However, in transplantation, DC therapy will be associated with the administration of immunosuppressive drugs, and interactions between drugs and DC are possible. Finally, we will discuss the issue of DC origin, as we believe that administration of autologous TolDC is more appropriate, as demonstrated by our experiments in animal models.
Keywords: Clinical trial, Human, Tolerogenic dendritic cells, Transplantation Introduction
Dendritic cells (DC) are potent antigen-presenting cells with dual functions; they can be either immunogenic or tolerogenic in nature. Several protocols of human DC generation have been described and both types of DC have clinical applications. Immunogenic DC are used in patients with cancer to reduce tumor development [1]. Tolerogenic DC (TolDC) therapy appears to be a prom-ising strategy for the treatment of autoimmune diseases and transplantation. In this mini-review, we will focus on human TolDC and their potential clinical application. Tolerogenic dendritic cells in humans
In the literature, several protocols have been described for the generation of human TolDC. In these studies, TolDC have been derived from monocytes (MoDC) using the cytokines granulocyte macrophage colony-stimulating factor (GM-CSF) and IL-4. However, as described for tolerogenic bone marrow-derived DC (BMDC) in animal models, different drugs or cytokines could be added to GM-CSF/IL-4 culture to manipulate
human DC in vitro, to obtain TolDC with specific fea-tures [2]. Among those methods, TolDC can be gener-ated with vitaminD3 (VitD3). VitD3-tregener-ated DC have the properties of tolerogenic DC; the cells are maturation-resistant, produce IL-10 after stimulation and induce a low proliferation of allogeneic T cells [3-5]. More re-cently, Raïch-Regué et al. showed that VitD3-DC gener-ated from the monocytes of healthy volunteers or patients with relapsing remitting multiple sclerosis have similar properties; a semi-mature phenotype, an anti-inflammatory profile and a low capacity to induce allo-geneic T cell proliferation [6]. Furthermore, these cells seem to show potential for clinical application since hyporesponsiveness of myelin-reactive T cells from patients with relapsing remitting multiple sclerosis was observed when these T cells were cultured with autolo-gous TolDC loaded with myelin peptides [6]. In parallel, several studies have investigated the generation of VitD3-TolDC together with dexamethasone (Dex) in order to increase their tolerogenic potential [7]. Prior to the clinical application of these dexamethasone/VitD3 TolDC in rheumatoid arthritis, Harry and colleagues compared the generation of TolDC from healthy volun-teers with those from patients with rheumatoid arthritis;
* Correspondence:[email protected]
INSERM, U1064, ITUN, CHU HôtelDieu, 30 Boulevard Jean Monnet, NANTES, France
© 2012 Moreau et al.; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
a similar phenotype and function was observed between the two groups [8]. In order to favor their migration to the draining lymph nodes and their antigen presentation to T cells, VitD3-DC or Dex/VitD3-DC can be matured in vitro with lipopolysaccharide (LPS). Such cells are described as alternatively activated DC [9,10] and induce memory T cell hyporesponsiveness and naive T cell liferation associated with low IFN-γ and high IL-10 pro-duction [9]. Other maturation stimuli such as a cytokine cocktail or monophosphoryl lipid A have also been ana-lyzed [11].
By contrast to Dex-DC and VitD3-DC, rapamycin-treated DC (Rapa-DC) express CD83 and CD86 markers and produce low amounts of IL-10 and high levels of IL-12p40/p70, characteristics of a mature DC phenotype [12]. However, Rapa-DC induce low-level proliferation of allogeneic T cells, similar to Dex-DC and VitD3-DC [13]. Furthermore, Rapa-DC secrete high levels of IL-12 after LPS stimulation, thereby promoting the induction of Treg Foxp3+cells in mice [14]a.
Another important molecule used to generate tolero-genic DC is IL-10. Two protocols have been used and lead to the differentiation of different types of TolDC depending on whether IL-10 is present from the initi-ation of culture or added at the end. In fact, DC gener-ated with IL-10 added at the end of culture have an immature phenotype and display resistance to matur-ation stimuli [15,16]. These DC induce a state of anergy in CD4+ T cells [16] and CD8+ T cells [17] in an antigen-specific manner [18]. More recently, DC derived from macaque monocytes in the presence of VitD3 and IL-10 were described as having tolerogenic properties, including resistance to maturation and low-level induc-tion of T cell proliferainduc-tion [19]. The authors demon-strated the safe intravenous injection of these DC to major histocompatibility complex (MHC)-mismatched recipient macaques treated with antihistamine drug and CTLA4Ig (Cytotoxic T lymphocyte Antigen-4 Ig). A transient increase in donor antigen-specific T cell prolif-eration was detected in these animals without any in-crease in anti-donor antibodies [19]. Another protocol to generate TolDC with IL-10 consists of culturing monocytes with IL-10 (in addition to GM-CSF and IL-4) from the initiation of culture. In this case, TolDC (called DC10) express CD83, CD80 and CD86, similar to acti-vated/mature cells, but also Ig-like transcript (ILT)2, ILT3, ILT4 and human leukocyte antigen G, similar to Tol-DC. Furthermore, DC10 secrete high levels of IL-10 and induce hyporesponsiveness in allogeneic T cells [20]. A key characteristic of DC generated with IL-10 is their ability to induce the differentiation of Tr1 regulatory T cells [20,21]b. Unfortunately, another property of
IL-10-producing DC is a decreased trafficking of these cells to the lymph nodes. The chemokine CCR7 participates in
the migration of DC to the lymph nodes, and generating mouse DC with IL-10 down-regulates their expression of CCR7 and impairs their in vivo homing to lymph nodes [22]. In a model of mouse cardiac allotransplantation, Garrod et al. showed that injection of DC co-expressing IL-10 and CCR7 induced a significant prolongation of graft survival. However, DC expressing either IL-10 only or CCR7 only had no effect [23].
Alternative protocols to generate tolerogenic antigen-presenting cells have been described by other teams, with the resulting cells being referred to as myeloid-derived suppressor cells [24]c, mesenchymal stem cells [25,26]d or regulatory macrophages [27]e. Clinical trials in transplantation using mesenchymal stem cells and regulatory macrophages have already been performed.
From the studies described in this section, some TolDC were generated using a clinical grade protocol prior to an application in the clinic [6,8,11,13]. In our center, we chose to generate human TolDC using a sim-ple protocol in which monocytes are cultured with low-dose GM-CSF without any additional cytokines or drugs. This protocol is in accordance with our work performed in mice [28] and is compatible with a clinical approach. Generation of human granulocyte macrophage
tolerogenic dendritic cells
Over the last few years, we have generated and charac-terized tolerogenic BMDC in rats, mice and non-human primates. In these different models, injection of tolero-genic BMDC leads to a reduced immune response in vivoor an induction of tolerance in transplant models [29-31]. Based on this expertise in TolDC generation in animals, we decided to derive TolDC in humans from monocytes in the presence of GM-CSF only. Indeed, the conventional cytokines used to derive dendritic cells from precursors are GM-CSF and IL-4. However, a study performed in mice in 2000 showed that DC generated with a low dose of GM-CSF in the absence of IL-4 have the properties of immature tolerogenic DC. These cells have a high capacity of antigen capture and presentation and induce a low proliferation of allogeneic T cells. Fur-thermore, they are maturation-resistant and lead to an increase of graft survival after in vivo injection [32]. In parallel, more recently, human MoDC generated in the presence of GM-CSF and without IL-4 were described to have tolerogenic properties in vitro [33]. Human GM-TolDC are derived from monocytes (0.5 million/mL) cultured with a low dose of GM-CSF (100 U/ml) for 6 days. Different doses of GM-CSF were tested and the best tolerogenic phenotype was obtained in the presence of a low dose. In our protocol, medium and cytokines do not have to be renewed as no difference in the pheno-type or function of the cells was observed with or with-out medium and/or cytokines replacement. On day 6,
Moreau et al. Transplantation Research 2012, 1:13 Page 2 of 8
cells are harvested and characterized on their phenotype and their function.
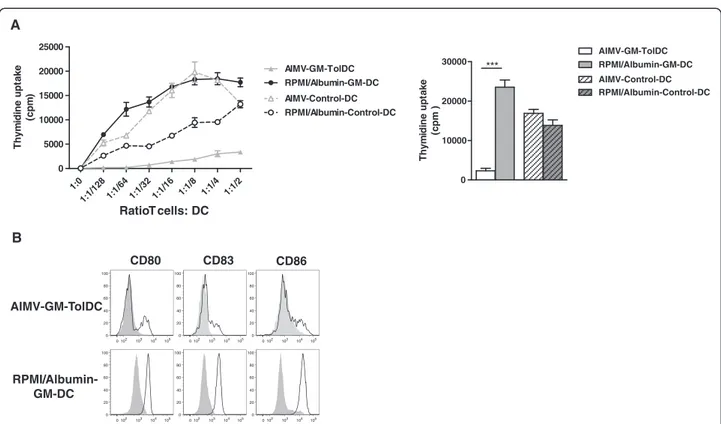
To set up this protocol, we tested methods of mono-cyte selection. The most common method to obtain monocytes is to positively select CD14-positive cells using microbeads. Another possibility is to enrich mono-cytes from peripheral blood by elutriation. This purifica-tion technique is based on the separapurifica-tion of cells according to their size and density [34]. The latter tech-nique, which has been adapted to Good Manufacturing Practice facilities, is much cheaper and isolates less manipulated monocytes. We derived TolDC in AIMV medium (Gibco Life Technologies) and GM-CSF (Cell-Genix) using these two techniques from the same donor. Analysis of the phenotype, function and maturation re-sistance of the cells generated from both monocyte isola-tion protocols gave similar results. Thus, all subsequent experiments were performed with elutriated monocytes. We next tested different clinical-grade culture media. We compared GM-TolDC cultured with GM-CSF alone (100 U/mL) in Roswell Park Memorial Institute (RPMI)/ human albumin medium and in AIMV medium for six days. In the literature, CellGroDC or X-VIVO 15 me-diums have also been used to derive human TolDC in clinical grade conditions [8,13]. Control non-tolerogenic DC were generated in parallel in the presence of GM-CSF (100 U/mL) and IL-4 (200 U/mL) in both types of media. After 6 days of culture, the DC were all non-adherent in RPMI/albumin medium whereas half of the cells were adherent in AIMV medium. Analysis of the phenotype and function of the cells revealed major differ-ences as the DC generated with RPMI/albumin did not have tolerogenic properties. As shown in Figure 1A, DC cultured with RPMI/albumin induced a strong prolifera-tion of allogeneic T cells, similar to that observed when T cells were cultured with control DC (either in RPMI/al-bumin or in AIMV media). By contrast, GM-TolDC in-duce a very low stimulation of allogeneic T cells, a feature that we described previously in rat and macaque tolerogenic BMDC [29,30]. Furthermore, a higher ex-pression of CD80 was detected at day 6 in RPMI/albu-min-GM-DC compared with the AIMV-GM-TolDC. This difference in phenotype was much greater after mat-uration with LPS/IFNγ, as RPMI/albumin-GM-DC highly over-expressed CD80, CD86 and CD83 (Figure 1B).
In summary, GM-TolDC were more than 90% MHC-IIlow CD80low CD86low CD40low with less than 2% con-tamination with T cells, B cells or natural killer cells. This immature phenotype of human TolDC is in accord-ance with the findings of our previous studies in rats, mice and non-human primates [28-30]. Furthermore, as we showed previously in rats [35], human GM-TolDC also expressed the tolerogenic marker Epstein-Barr virus induced gene 3 protein. As regards to their function,
GM-TolDC induced a weak stimulation of allogeneic T cells compared with control DC. We also found these cells to be semi-resistant to maturation induced by LPS/ IFNγ (as shown in Figure 1). In terms of cytokine re-lease, GM-TolDC produced IL-10 but no IL-12 when stimulated with LPS/IFNγ. Like their phenotype, the in vitro function (that is, T cell proliferation induction, maturation-resistance and cytokine production) of human GM-TolDC was similar to that of tolerogenic BMDC described in animal models [28-30]. The similar-ities between the in vitro features of animal TolDC, whose efficacy has been proven in vivo, with those obtained in humans, is encouraging for their potential use in the clinic.
Clinical application of tolerogenic dendritic cells
Whereas clinical trials using immunogenic DC to treat cancer have been developed over the last 15 years [1,36], TolDC therapy is only just emerging in the clinical arena. This was initiated by the pioneer study published in 2001 demonstrating the safety of injecting autologous immature DC into healthy volunteers [37]. Injections of these DC by the subcutaneous route were well-tolerated without signs of toxicity or development of autoimmun-ity. Inhibition of antigen-specific effector T cell function and induction of antigen-specific CD8 Tregs in vivo were detected in DC-treated volunteers [37,38]. More recently, the first phase I clinical trial using TolDC and including 10 patients was reported in type 1 diabetes [39]. Control DC generated in the presence of GM-CSF and IL-4 were injected into three patients while seven patients received immunosuppressive DC generated in the presence of GM-CSF, IL-4 and antisense oligonu-cleotides targeting CD40, CD80 and C86 transcripts. In this trial, intra-dermal injections of both control and im-munosuppressive autologous TolDC were well-tolerated and safe [39]. Furthermore, two clinical trials in rheuma-toid arthritis are on-going, one by Thomas’s group in Australia (University of Queensland) and the other one by Hilkens’s and Isaacs’s group in the UK (University of Newcastle). These different studies highlight the emer-gence of tolerogenic DC therapy as a new approach to treat autoimmune diseases.
As part of a European project, we plan to test the safety of autologous monocyte-derived TolDC in patients who have had a kidney transplant. To avoid graft rejection, transplant patients receive life-long im-munosuppressive drugs (IS). That means that in this clinical trial, TolDC will be injected into patients receiv-ing three IS commonly used in transplantation: tacroli-mus, mycophenolate mofetil (MMF) and prednisolone. However, as summarized in Table 1, several studies per-formed in mice and humans have shown that generation of DC in the presence of these IS modifies the DC
phenotype and function. In particular, generation of mouse BMDC with tacrolimus reduces the ability of DC to process and/or present antigens [40,41]. All the drug-treated DC have been shown to induce hypoproliferation of allogeneic T cells. Interestingly, a study published in 2005 showed that neither migration nor survival of injected BMDC were affected by tacrolimus-treatment of recipient rats [42]. Furthermore, co-treatment with donor DC and tacrolimus increased the hyporesponsive-ness of recipient T cells. In a model of heart allotrans-plantation, donor DC alone did not delay rejection while tacrolimus or tacrolimus + donor DC induced a pro-longation of graft survival [42]. This work was per-formed using donor-derived mature BMDC. Prior to the clinical trial, we will test the absence of toxicity of our TolDC in IS-treated animals in a mouse skin graft model. In parallel, graft survival will be monitored in animals treated with IS without TolDC therapy. So far, we have noted that injection of MMF induces a pro-longation of graft survival and injection of TolDC does not impair this effect. In fact, a slight increase in graft survival was actually detected (Segovia et al., manuscript in preparation). Similar experiments using the two other
IS or the combination of both IS associated or not with DC therapy are on-going. These studies should identify any potential side effects of DC therapy on the IS treat-ment used in the clinic.
Others parameters related to DC injections should be considered before doing a clinical trial, such as the time of cell product injection, the numbers of DC injected, and also the number of injections as well as the route of administra-tion. Regarding this last parameter, experiments performed in mice have shown that intravenous injection of Dex/LPS-treated BMDC prolongs cardiac transplant survival whereas subcutaneous injection of the same Dex/LPS-treated BMDC does not increase graft survival [58]. Our prelimin-ary experiments in macaques show that intradermal injec-tion of autologous TolDC prime an immune response while intravenous injection does not (unpublished results). A study also performed in monkeys confirmed the fact that intravenous injection of TolDC is well-tolerated [19]. The potential of autologous tolerogenic dendritic cells in transplantation
The clinical trials described in the previous section of this review either have already been performed or are
Thym id in e upt a ke (c p m ) 1:1/ 2 1:1/ 4 1:1/8 1:1/ 16 1:1/ 32 1:1/6 4 1:1/1 28 1:0 0 5000 10000 15000 20000 25000 AIMV-GM-TolDC RPMI/Albumin-GM-DC AIMV-Control-DC RPMI/Albumin-Control-DC Thy m idi ne up ta k e (c p m ) 0 10000 20000 30000 AIMV-GM-TolDC RPMI/Albumin-GM-DC RPMI/Albumin-Control-DC AIMV-Control-DC *** RPMI/Albumin-GM-DC 0 102 103 104 105 0 20 40 60 80 100 0 102 103 104 105 0 20 40 60 80 100 0 102 103 104 105 0 20 40 60 80 100 AIMV-GM-TolDC CD83 CD86 CD80 0 102 103 104 105 0 20 40 60 80 100 0 102 103 104 105 0 20 40 60 80 100 0 102 103 104 105 0 20 40 60 80 100 A B Ratio T cells: DC
Figure 1 Influence of culture medium on human dendritic cell differentiation in vitro. Four populations of DC were generated in two different culture media, either RPMI/albumin or AIMV and with two different cytokine conditions, either low-dose GM-CSF (GM-DC) or GM-CSF and IL-4 (Control DC) for 6 days. (A) The four DC populations were cultured with allogeneic T cells at different ratios for 6 days. A representative experiment is shown on the left-hand panel. On the right, the results of the ratio of one DC to four Tcells are expressed as the mean T cell proliferation + standard error of the mean for three different donors (*** P < 0.001, paired T tests). (B) The four populations were un-stimulated (grey solid histogram) or stimulated with 200 ng/mL LPS and 50 ng/mL IFN-γ (black line) for 48 hours. Cell surface expression of CD80, CD83 and CD86 was analyzed after 48 hours of culture by flow cytometry. Data shown are representative of two experiments performed..
Moreau et al. Transplantation Research 2012, 1:13 Page 4 of 8
on-going in autoimmune diseases. In transplantation, another parameter has to be taken into consideration; this is the question of whether TolDC should be derived from the donor or from the recipient. Most of the stud-ies described in rodents have been performed using donor TolDC or recipient TolDC loaded with donor peptides and administrated one week or more before transplantation [2]. In this last part, we discuss the rele-vance of using autologous TolDC from a safety and effi-cacy point of view.
Firstly and very importantly, the risk of donor sensitization due to the presence of a slight contaminant
cell product or the destruction of the injected cells by non-self recognition cannot be excluded using donor TolDC therapy [59]; we hypothesize that this risk is minimized using autologous TolDC. Furthermore, to be efficient, donor TolDC (or donor pulsed recipient TolDC) need to be sufficiently activated using LPS or other cytokine cocktails in order to migrate to lymphoid organs and present the antigen to T cells [9-11]. To avoid activation-induced maturation, TolDC are also modified using Dex, VitD3 or IL-10 as described in the first section of this review. By contrast, in accordance with our previous work in rodents [30], autologous Table 1 Effects of tacrolimus, mycophenolate mofetil and prednisolone in tolerogenic dendritic cells generated in vitro
Drugs Models Effects of the drugs on DC and therapeutic effects of the modified DC References Tacrolimus Mouse BMDC +/−
Tacrolimus
- Reduction in MHC Class I and Class II-restricted presentation of antigen (role of tacrolimus on Ag processing or presentation)
[40,41,43,44] - Reduction in pro-inflammatory cytokine (IL-6 and IL-12) secretion and
expression of CD40 and CD86 by BMDC after maturation
- Tacrolimus-treated BMDC induce a low proliferation of allogeneic T cells - Low CD69 expression and IFN-γ, IL-2 and IL-4 secretion by T cells stimulated with tacrolimus-treated BMDC
Human MoDC +/− Tacrolimus
- Low expression of CXCL10 [45-50] - Reduction in pro-inflammatory cytokine (TNFα and IL-12)
secretion by BMDC after maturation
- Decrease in CD83 and CD86 expression after LPS maturation in the presence of high levels of tacrolimus only
- Tacrolimus-treated MoDC induce a low level of allogeneic T cell proliferation, in favor of Th2 cells
- Low CD69 expression and IFN-γ, IL-2 and IL-4 secretion by T cells stimulated with tacrolimus-treated MoDC (immature or mature) Mycophenolate
mofetil
Mouse BMDC +/− MMF
- Decrease in co-stimulatory markers CD80, CD86 and CD40 expression [51,52] - Decrease in IL-12 production
- MMF-treated BMDC induce a low proliferation of allogeneic T cells - Decrease in DTH response and increase in allograft survival after injection of MMF-treated-BMDC
Human MoDC +/− MMF
- Decrease in co-stimulatory marker expression [53] - Induction of LPS-maturation resistance (in terms of both
phenotype and cytokine release)
- MMF-treated MoDC induce a low proliferation of allogeneic T cells - Reduction in endocytic capacity in mature MMF-treated MoDC (related to mannose receptor expression)
Prednisolone Mouse BMDC +/− prednisolone
- IL-10/methylprednisolone-treated BMDC increase survival of skin allografts [54] Human MoDC +/−
prednisolone
- Increase in endocytic capacity [55-57] - Induction of LPS maturation resistance (absence of
CD80/CD86/CD83 up-regulation)
- Increase in anti-inflammatory cytokines (IL-10 and TGF-β) and decrease in pro-inflammatory cytokines (IL-6, IL-12, IL-23 and TNFα)
- Prednisolone-treated MoDC induce a low-level proliferation of allogeneic T cells (which acquire suppressive functions)
BMDC: bone marrow-derived dendritic cells; CD: cluster of differentiation; CXCL10: chemokine (C-X-C motif) ligand 10; IFN-γ: interferon gamma; IL: interleukin; LPS: lipopolysaccharide; MHC: major histocompatibility complex; MMF: mycophenolate mofetil; MoDC: monocyte-derived dendritic cells; TNF α: tumor necrosis factor alpha.
TolDC do not require activation or pulsing to be effi-cient, leading to a reduced risk of cell maturation. In ac-cordance with this hypothesis, the first clinical trial using human TolDC performed with MoDC generated with GM-CSF and IL-4 showed no toxicity and no ad-verse effects in the patients injected with these DC [39], supporting the theory that autologous TolDC do not be-come immunogenic after injection, correlated with an absence of maturation of the cells.
Secondly, as regards the efficacy of the cells, some studies performed recently in mice by Morelli’s group demonstrated that injected donor DC die quickly after in vivoinjection. Even if administration of donor TolDC induces tolerance to a transplant, donor TolDC are un-able to directly regulate an immune response in vivo [60]. In this context, donor DC mediate their suppres-sive effects on T cells through endogenous conventional DC from the recipient mouse [61]. We believe the mechanisms of action to be different when using autolo-gous TolDC. Indeed, we detected injected recipient rat TolDC in the spleen at least two weeks post-injection [30]. Studies performed in mice and humans have also shown that DC accumulate in the spleen after intraven-ous injection [62,63]. Furthermore, our experiments showed that donor-derived MHC ClassII+cells from the graft are present in the spleen of the recipient 3 to 5 days after transplantation. Stainings performed in the spleens of these animals suggest that the donor cells interact with the injected TolDC. Moreover, depletion of graft passenger leukocytes from the donor organ before trans-plantation prevents any effect of the autologous TolDC injection (Segovia et al. submitted manuscript). These results validate the hypothesis that injected autologous un-pulsed TolDC are able to migrate to the spleen where they capture and process the donor antigen from graft passenger leukocytes (Segovia et al. submitted manuscript) leading to antigen-specific graft acceptance [31].
Conclusions
As demonstrated by the clinical trial in patients with diabetes [39], the use of autologous tolerogenic DC appears to be a potential safe method that may promote alloantigen-specific Tcell unresponsiveness and trans-plant survival. Our experiments performed in animals suggest that, to be efficient, injected autologous TolDC have to be in contact with donor antigens and the ad-ministration of TolDC at the time of the graft is import-ant. In the clinical trial, patients will be treated with autologous TolDC at the time of the transplant and will be additionally treated with low-dose IS (tacrolimus, prednisolone, MMF). As this will be a phase I trial, we will test the safety and toxicity of TolDC therapy in transplantation. However, we cannot exclude the
possibility that IS will have a negative effect on the func-tion of the injected DC, notably by inhibiting antigen presentation as already observed when TolDC were gen-erated with tacrolimus. An alternative could be to change the drugs used or to wait until a decrease in the immunosuppressive treatment is possible before inject-ing the TolDC.
Ethical approval
Human leukapheresis samples were collected from healthy donors following institutional-approved proto-cols (Etablissement Français du Sang, Nantes, France). All animal experiments were performed under specific pathogen-free conditions in accordance with the Euro-pean Union Guidelines and in compliance with the eth-ical rules of the INSERM.
Endnotes
a
Camilla Macedo;bGiada Amodio;cNatasa Obermajer;
d
Martin Hoogduijn and Elke Eggenhofer;eJames Hutch-inson and Paloma Riquelme participated in The One Study Workshop 2012 and described Rapa-DC, DC10, myeloid-derived suppressor cells, mesenchymal stem cells and regulatory macrophages respectively in other mini-reviews.
Abbreviations
BMDC: Bone-marrow derived dendritic cells; CD: Cluster of differentiation; DC: Dendritic cells; Dex: Dexamethasone; GM-CSF: Granulocyte macrophage colony-stimulating factor; IFN-γ: Interferon gamma; Ig: Immunoglobulin; ILT: Ig-like transcript; IS: Immunosuppressive drugs; LPS: Lipopolysaccharide; MHC: Major histocompatibility complex; MoDC: Monocyte derived DC; MMF: Mycophenolate Mofetil; Rapa: Rapamycin; RPMI: Roswell Park Memorial Institute; TolDC: Tolerogenic DC; VitD3: Vitamin D3.
Competing interests
The authors declare that they have no competing interests. Authors’ contributions
AM participated in the design of the study and drafted the manuscript. EV helped to draft the manuscript. LBD carried out the cell culture and the different assays. MCC conceived the study, participated in its design and coordination, revised and gave the final approval of the manuscript. All the authors read and approved the final manuscript.
Acknowledgments
The authors are grateful for funding support from The One Study, Centaure and Progreffe Foundation.
Received: 2 May 2012 Accepted: 4 August 2012 Published: 28 September 2012
References
1. Correale P, Campoccia G, Tsang KY, Micheli L, Cusi MG, Sabatino M, Bruni G, Sestini S, Petrioli R, Pozzessere D, Marsili S, Fanetti G, Giorgi G, Francini G: Recruitment of dendritic cells and enhanced antigen-specific immune reactivity in cancer patients treated with hr-GM-CSF (Molgramostim) and hr-IL-2. results from a phase Ib clinical trial. Eur J Cancer 2001,
37(7):892–902.
2. Morelli AE, Thomson AW: Tolerogenic dendritic cells and the quest for transplant tolerance. Nat Rev Immunol 2007, 7(8):610–621.
3. Canning MO, Grotenhuis K, de Wit H, Ruwhof C, Drexhage HA: 1-alpha, 25-Dihydroxyvitamin D3 (1,25(OH)(2)D(3)) hampers the maturation of
Moreau et al. Transplantation Research 2012, 1:13 Page 6 of 8
fully active immature dendritic cells from monocytes. Eur J Endocrinol 2001, 145(3):351–357.
4. Penna G, Adorini L: 1 Alpha,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol 2000, 164(5):2405–2411.
5. Piemonti L, Monti P, Sironi M, Fraticelli P, Leone BE, Dal Cin E, Allavena P, Di Carlo V: Vitamin D3 affects differentiation, maturation, and function of human monocyte-derived dendritic cells. J Immunol 2000, 164(9):4443–4451.
6. Raïch-Regué D, Grau-Lopez L, Naranjo-Gomez M, Ramo-Tello C, Pujol-Borrell R, Martinez-Caceres E, Borras FE: Stable antigen-specific T-cell
hyporesponsiveness induced by tolerogenic dendritic cells from multiple sclerosis patients. Eur J Immunol 2012, 42(3):771–782. 7. Pedersen AE, Gad M, Walter MR, Claesson MH: Induction of regulatory
dendritic cells by dexamethasone and 1alpha,25-Dihydroxyvitamin D(3). ImmunolLett 2004, 91(1):63–69.
8. Harry RA, Anderson AE, Isaacs JD, Hilkens CM: Generation and
characterisation of therapeutic tolerogenic dendritic cells for rheumatoid arthritis. Ann Rheum Dis 2010, 69(11):2042–2050.
9. Anderson AE, Sayers BL, Haniffa MA, Swan DJ, Diboll J, Wang XN, Isaacs JD, Hilkens CM: Differential regulation of naive and memory CD4+ T cells by alternatively activated dendritic cells. J LeukocBiol 2008, 84(1):124–133. 10. Anderson AE, Swan DJ, Sayers BL, Harry RA, Patterson AM, von Delwig A,
Robinson JH, Isaacs JD, Hilkens CM: LPS activation is required for migratory activity and antigen presentation by tolerogenic dendritic cells. J LeukocBiol 2009, 85(2):243–250.
11. Raïch-Regué D, Naranjo-Gomez M, Grau-Lopez L, Ramo C, Pujol-Borrell R, Martinez-Caceres E, Borras FE: Differential effects of monophosphoryl lipid A and cytokine cocktail as maturation stimuli of immunogenic and tolerogenic dendritic cells for immunotherapy. Vaccine 2012, 30(2):378–387.
12. Turnquist HR, Cardinal J, Macedo C, Rosborough BR, Sumpter TL, Geller DA, Metes D, Thomson AW: mTOR and GSK-3 shape the CD4+ T-cell stimulatory and differentiation capacity of myeloid DCs after exposure to LPS. Blood 2010, 115(23):4758–4769.
13. Naranjo-Gomez M, Raïch-Regué D, Onate C, Grau-Lopez L, Ramo-Tello C, Pujol-Borrell R, Martinez-Caceres E, Borras FE: Comparative study of clinical grade human tolerogenic dendritic cells. J Transl Med 2011, 9:89. 14. Turnquist HR, Raimondi G, Zahorchak AF, Fischer RT, Wang Z, Thomson AW:
Rapamycin-conditioned dendritic cells are poor stimulators of allogeneic CD4+ T cells, but enrich for antigen-specific Foxp3+ T regulatory cells and promote organ transplant tolerance. J Immunol 2007,
178(11):7018–7031.
15. Buelens C, Verhasselt V, De Groote D, Thielemans K, Goldman M, Willems F: Human dendritic cell responses to lipopolysaccharide and CD40 ligation are differentially regulated by interleukin-10. Eur J Immunol 1997, 27(8):1848–1852.
16. Buelens C, Willems F, Delvaux A, Pierard G, Delville JP, Velu T, Goldman M: Interleukin-10 differentially regulates B7-1 (CD80) and B7-2 (CD86) expression on human peripheral blood dendritic cells. Eur J Immunol 1995, 25(9):2668–2672.
17. Steinbrink K, Jonuleit H, Muller G, Schuler G, Knop J, Enk AH: Interleukin-10-treated human dendritic cells induce a melanoma-antigen-specific anergy in CD8(+) T cells resulting in a failure to lyse tumor cells. Blood 1999, 93(5):1634–1642.
18. Steinbrink K, Graulich E, Kubsch S, Knop J, Enk AH: CD4(+) and CD8(+) anergic T cells induced by interleukin-10-treated human dendritic cells display antigen-specific suppressor activity. Blood 2002, 99(7):2468–2476.
19. Zahorchak AF, Kean LS, Tokita D, Turnquist HR, Abe M, Finke J, Hamby K, Rigby MR, Larsen CP, Thomson AW: Infusion of stably immature monocyte-derived dendritic cells plus CTLA4Ig modulates alloimmune reactivity in rhesus macaques. Transplantation 2007, 84(2):196–206. 20. Gregori S, Tomasoni D, Pacciani V, Scirpoli M, Battaglia M, Magnani CF,
Hauben E, Roncarolo MG: Differentiation of type 1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10-dependent ILT4/HLA-G pathway. Blood 2010, 116(6):935–944.
21. Levings MK, Gregori S, Tresoldi E, Cazzaniga S, Bonini C, Roncarolo MG: Differentiation of Tr1 cells by immature dendritic cells requires IL-10 but not CD25 + CD4+ Tr cells. Blood 2005, 105(3):1162–1169.
22. Takayama T, Morelli AE, Onai N, Hirao M, Matsushima K, Tahara H, Thomson AW: Mammalian and viral IL-10 enhance C-C chemokine receptor 5 but down-regulate C-C chemokine receptor 7 expression by myeloid dendritic cells: impact on chemotactic responses andin vivohoming ability. J Immunol 2001, 166(12):7136–7143.
23. Garrod KR, Chang CK, Liu FC, Brennan TV, Foster RD, Kang SM: Targeted lymphoid homing of dendritic cells is required for prolongation of allograft survival. J Immunol 2006, 177(2):863–868.
24. Obermajer N, Muthuswamy R, Lesnock J, Edwards RP, Kalinski P: Positive feedback between PGE2 and COX2 redirects the differentiation of human dendritic cells toward stable myeloid-derived suppressor cells. Blood 2011, 118(20):5498–5505.
25. MISOT Study Group, Hoogduijn MJ, Popp FC, Grohnert A, Crop MJ, van Rhijn M, Rowshani AT, Eggenhofer E, Renner P, Reinders ME, Rabelink TJ, van der Laan LJ, Dor FJ, Ijzermans JN, Genever PG, Lange C, Durrbach A, Houtgraaf JH, Christ B, Seifert M, Shagidulin M, Donckier V, Deans R, Ringden O, Perico N, Remuzzi G, Bartholomew A, Schlitt HJ, Weimar W, Baan CC, Dahlke MH: Advancement of mesenchymal stem cell therapy in solid organ transplantation (MISOT). Transplantation 2010, 90(2):124–126.
26. Roemeling-van Rhijn M, Weimar W, Hoogduijn MJ: Mesenchymal stem cells: application for solid-organ transplantation. CurrOpin Organ Transplant 2012, 17(1):55–62.
27. Hutchinson JA, Riquelme P, Sawitzki B, Tomiuk S, Miqueu P, Zuhayra M, Oberg HH, Pascher A, Lutzen U, Janssen U, Broichhausen C, Renders L, Thaiss F, Scheuermann E, Henze E, Volk HD, Chatenoud L, Lechler RI, Wood KJ, Kabelitz D, Schlitt HJ, Geissler EK, Fändrich F: Cutting edge:
immunological consequences and trafficking of human regulatory macrophages administered to renal transplant recipients. J Immunol 2011, 187(5):2072–2078.
28. Segovia M, Cuturi MC, Hill M: Preparation of mouse bone marrow-derived dendritic cells with immunoregulatory properties. Methods MolBiol 2011, 677:161–168.
29. Moreau A, Hill M, Thebault P, Deschamps JY, Chiffoleau E, Chauveau C, Moullier P, Anegon I, Alliot-Licht B, Cuturi MC: Tolerogenic dendritic cells actively inhibit T cells through heme oxygenase-1 in rodents and in nonhuman primates. FASEB J 2009, 23(9):3070–3077.
30. Peche H, Trinite B, Martinet B, Cuturi MC: Prolongation of heart allograft survival by immature dendritic cells generated from recipient type bone marrow progenitors. Am J Transplant 2005, 5(2):255–267.
31. Beriou G, Peche H, Guillonneau C, Merieau E, Cuturi MC: Donor-specific allograft tolerance by administration of recipient-derived immature dendritic cells and suboptimal immunosuppression. Transplantation 2005, 79(8):969–972.
32. Lutz MB, Suri RM, Niimi M, Ogilvie AL, Kukutsch NA, Rossner S, Schuler G, Austyn JM: Immature dendritic cells generated with low doses of GM-CSF in the absence of IL-4 are maturation resistant and prolong allograft survival in vivo. Eur J Immunol 2000, 30(7):1813–1822. 33. Chitta S, Santambrogio L, Stern LJ: GMCSF in the absence of other
cytokines sustains human dendritic cell precursors with T cell regulatory activity and capacity to differentiate into functional dendritic cells. ImmunolLett 2008, 116(1):41–54.
34. Berger TG, Strasser E, Smith R, Carste C, Schuler-Thurner B, Kaempgen E, Schuler G: Efficient elutriation of monocytes within a closed system (Elutra) for clinical-scale generation of dendritic cells. J Immunol Methods 2005, 298(1–2):61–72.
35. Hill M, Thebault P, Segovia M, Louvet C, Beriou G, Tilly G, Merieau E, Anegon I, Chiffoleau E, Cuturi MC: Cell therapy with autologous tolerogenic dendritic cells induces allograft tolerance through interferon-gamma and Epstein-Barr virus-induced gene 3. Am J Transplant 2011, 11(10):2036–2045.
36. Redman BG, Chang AE, Whitfield J, Esper P, Jiang G, Braun T, Roessler B, Mule JJ: Phase Ib trial assessing autologous, tumor-pulsed dendritic cells as a vaccine administered with or without IL-2 in patients with metastatic melanoma. J Immunother 2008, 31(6):591–598. 37. Dhodapkar MV, Steinman RM, Krasovsky J, Munz C, Bhardwaj N:
Antigen-specific inhibition of effector T cell function in humans after injection of immature dendritic cells. J Exp Med 2001, 193(2):233–238. 38. Dhodapkar MV, Steinman RM: Antigen-bearing immature dendritic cells
induce peptide-specific CD8(+) regulatory T cells in vivo in humans. Blood 2002, 100(1):174–177.
39. Giannoukakis N, Phillips B, Finegold D, Harnaha J, Trucco M: Phase I (safety) study of autologous tolerogenic dendritic cells in type 1 diabetic patients. Diabetes Care 2011, 34(9):2026–2032.
40. Imai A, Sahara H, Tamura Y, Jimbow K, Saito T, Ezoe K, Yotsuyanagi T, Sato N: Inhibition of endogenous MHC class II-restricted antigen presentation by tacrolimus (FK506) via FKBP51. Eur J Immunol 2007, 37(7):1730–1738. 41. Lee YR, Yang IH, Lee YH, Im SA, Song S, Li H, Han K, Kim K, Eo SK, Lee CK: Cyclosporin A and tacrolimus, but not rapamycin, inhibit MHC-restricted antigen presentation pathways in dendritic cells. Blood 2005,
105(10):3951–3955.
42. Azhipa O, Kimizuka K, Nakao A, Toyokawa H, Okuda T, Neto JS, Alber SM, Kaizu T, Thomson AW, Demetris AJ, Murase N: Comparative analysis of the fate of donor dendritic cells and B cells and their influence on alloreactive T cell responses under tacrolimus immunosuppression. ClinImmunol 2005, 114(2):199–209.
43. Matsue H, Yang C, Matsue K, Edelbaum D, Mummert M, Takashima A: Contrasting impacts of immunosuppressive agents (rapamycin, FK506, cyclosporin A, and dexamethasone) on bidirectional dendritic cell-T cell interaction during antigen presentation. J Immunol 2002, 169(7):3555–3564.
44. Morelli AE, Antonysamy MA, Takayama T, Hackstein H, Chen Z, Qian S, Zurowski NB, Thomson AW: Microchimerism, donor dendritic cells, and alloimmune reactivity in recipients of Flt3 ligand-mobilized hemopoietic cells: modulation by tacrolimus. J Immunol 2000, 165(1):226–237. 45. Cos J, Villalba T, Parra R, Gallardo D, Bilbao I, Margarit C, Massuet L: FK506 in
the maturation of dendritic cells. Haematologica 2002, 87(7):679–687. discussion 687.
46. Duperrier K, Velten FW, Bohlender J, Demory A, Metharom P, Goerdt S: Immunosuppressive agents mediate reduced allostimulatory properties of myeloid-derived dendritic cells despite induction of divergent molecular phenotypes. MolImmunol 2005, 42(12):1531–1540. 47. Monti P, Mercalli A, Leone BE, Valerio DC, Allavena P, Piemonti L:
Rapamycin impairs antigen uptake of human dendritic cells. Transplantation 2003, 75(1):137–145.
48. Shimizu K, Fujii S, Fujimoto K, Kawa K, Yamada A, Kawano F: Tacrolimus (FK506) treatment of CD34+ hematopoietic progenitor cells promote the development of dendritic cells that drive CD4+ T cells toward Th2 responses. J LeukocBiol 2000, 68(5):633–640.
49. Szabo G, Gavala C, Mandrekar P: Tacrolimus and cyclosporine A inhibit allostimulatory capacity and cytokine production of human myeloid dendritic cells. J Investig Med 2001, 49(5):442–449.
50. Tiefenthaler M, Hofer S, Ebner S, Ivarsson L, Neyer S, Herold M, Mayer G, Fritsch P, Heufler C: In vitrotreatment of dendritic cells with tacrolimus: impaired T-cell activation and IP-10 expression. Nephrol Dial Transplant 2004, 19(3):553–560.
51. Han CH, Li HF, Wang YX, Zhang M, Wang Y, Yin M, Min ZL, Zheng KL: The influence of mycophenolate mofetil upon the maturation and allostimulatory activity of cultured dendritic cell progenitors and the effects of tolerance induction in allograft recipients. Zhonghua Yi XueZaZhi 2005, 85(19):1327–1332.
52. Mehling A, Grabbe S, Voskort M, Schwarz T, Luger TA, Beissert S: Mycophenolatemofetil impairs the maturation and function of murine dendritic cells. J Immunol 2000, 165(5):2374–2381.
53. Colic M, Stojic-Vukanic Z, Pavlovic B, Jandric D, Stefanoska I: Mycophenolate mofetil inhibits differentiation, maturation and allostimulatory function of human monocyte-derived dendritic cells. ClinExpImmunol 2003, 134(1):63–69.
54. Li X, Dou KF, Liu HL, Zhang FQ, Cai L: Immune tolerance induced by IL-10 and methylprednisolone modified dendritic cellsin vitro. Xi Bao Yu Fen ZiMian Yi XueZaZhi 2007, 23(5):436–438.
55. Luther C, Adamopoulou E, Stoeckle C, Brucklacher-Waldert V, Rosenkranz D, Stoltze L, Lauer S, Poeschel S, Melms A, Tolosa E: Prednisolone treatment induces tolerogenic dendritic cells and a regulatory milieu in myasthenia gravis patients. J Immunol 2009, 183(2):841–848.
56. Rozkova D, Horvath R, Bartunkova J, Spisek R: Glucocorticoids severely impair differentiation and antigen presenting function of dendritic cells despite upregulation of Toll-like receptors. ClinImmunol 2006, 120(3):260–271.
57. Vanderheyde N, Verhasselt V, Goldman M, Willems F: Inhibition of human dendritic cell functions by methylprednisolone. Transplantation 1999, 67(10):1342–1347.
58. Emmer PM, van der Vlag J, Adema GJ, Hilbrands LB: Dendritic cells activated by lipopolysaccharide after dexamethasone treatment induce donor-specific allograft hyporesponsiveness. Transplantation 2006, 81(10):1451–1459.
59. Yu G, Xu X, Vu MD, Kilpatrick ED, Li XC: NK cells promote transplant tolerance by killing donor antigen-presenting cells. J Exp Med 2006, 203(8):1851–1858.
60. Divito SJ, Wang Z, Shufesky WJ, Liu Q, Tkacheva OA, Montecalvo A, Erdos G, Larregina AT, Morelli AE: Endogenous dendritic cells mediate the effects of intravenously injected therapeutic immunosuppressive dendritic cells in transplantation. Blood 2010, 116(15):2694–2705.
61. Wang Z, Divito SJ, Shufesky WJ, Sumpter T, Wang H, Tkacheva OA, Wang W, Liu C, Larregina AT, Morelli AE: Dendritic cell therapies in transplantation revisited: deletion of recipient DCs deters the effect of therapeutic DCs. Am J Transplant 2012, 12(6):1398–1408.
62. Eggert AA, Schreurs MW, Boerman OC, Oyen WJ, de Boer AJ, Punt CJ, Figdor CG, Adema GJ: Biodistribution and vaccine efficiency of murine dendritic cells are dependent on the route of administration. Cancer Res 1999, 59(14):3340–3345.
63. Mackensen A, Krause T, Blum U, Uhrmeister P, Mertelsmann R, Lindemann A: Homing of intravenously and intralymphatically injected human dendritic cells generatedin vitrofrom CD34+ hematopoietic progenitor cells. Cancer ImmunolImmunother 1999, 48(2–3):118–122.
doi:10.1186/2047-1440-1-13
Cite this article as: Moreau et al.: Cell therapy using tolerogenic dendritic cells in transplantation. Transplantation Research 2012 1:13.
Submit your next manuscript to BioMed Central and take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit
Moreau et al. Transplantation Research 2012, 1:13 Page 8 of 8