Caractérisation de la protéine Naf1 chez Saccharomyces cerevisiae et chez l'homme
267
0
0
Texte intégral
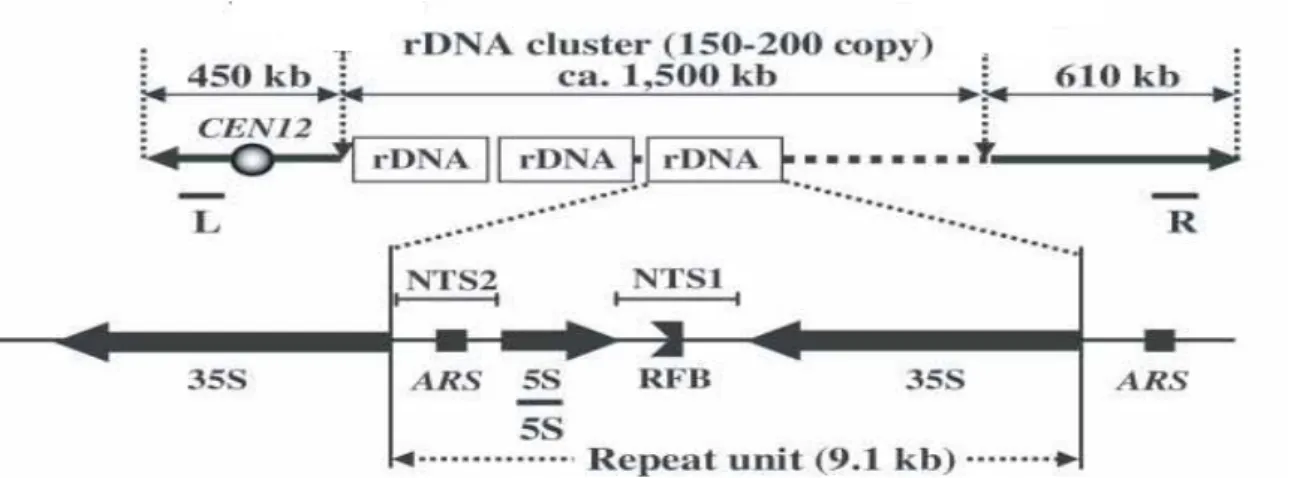
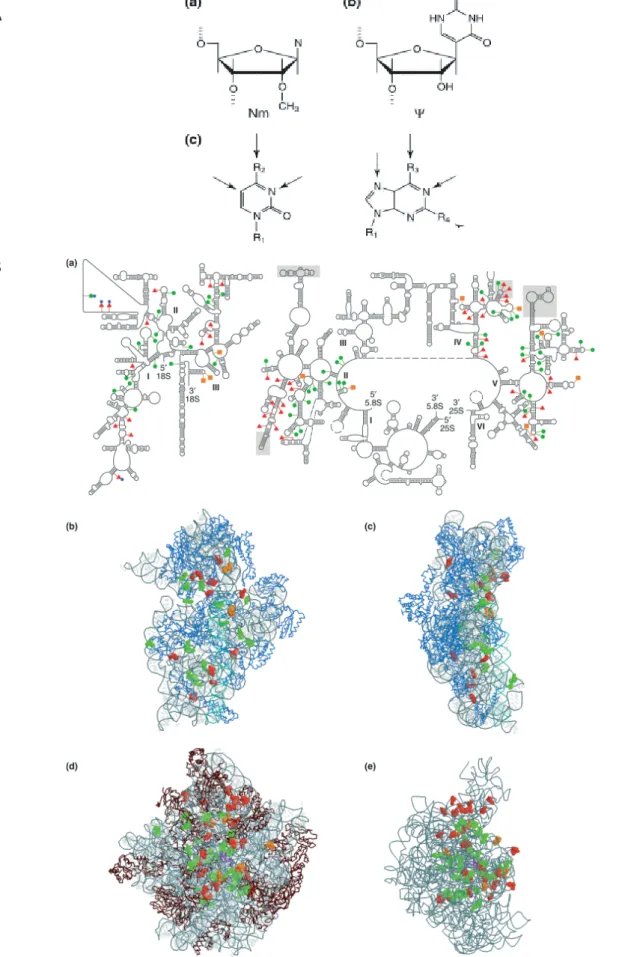
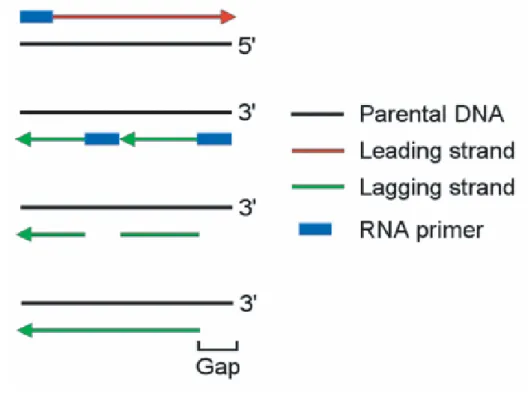
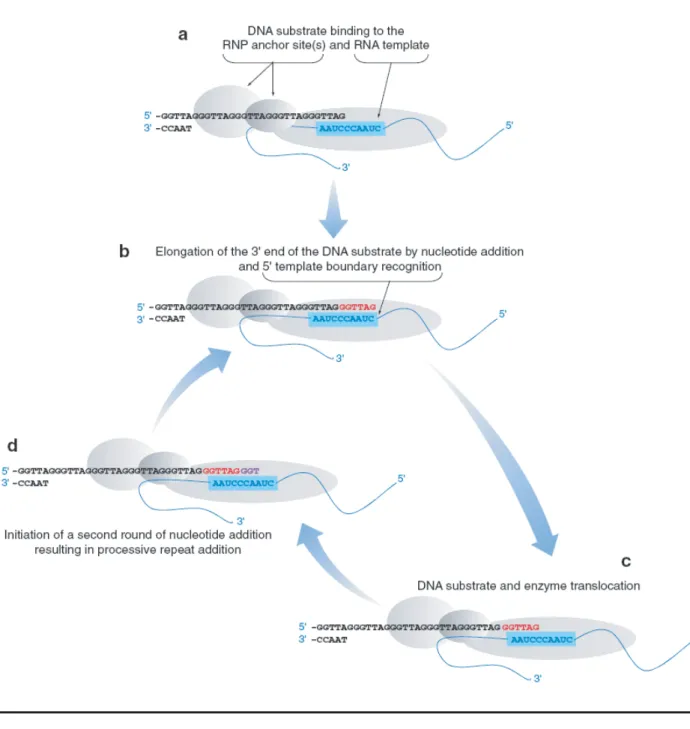
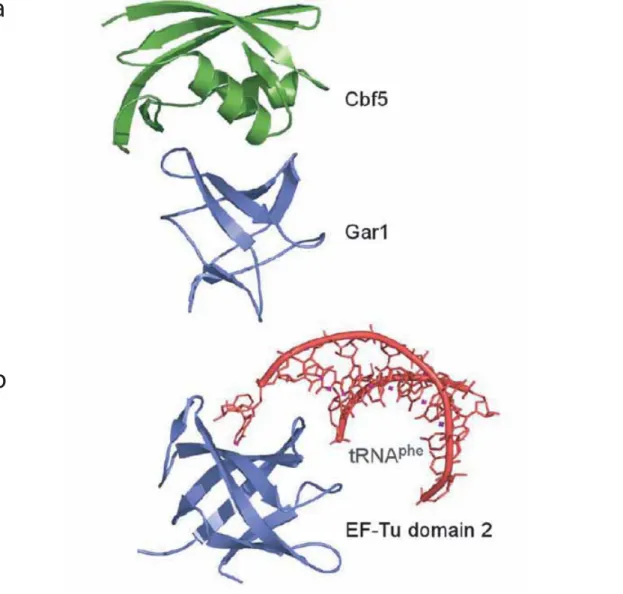
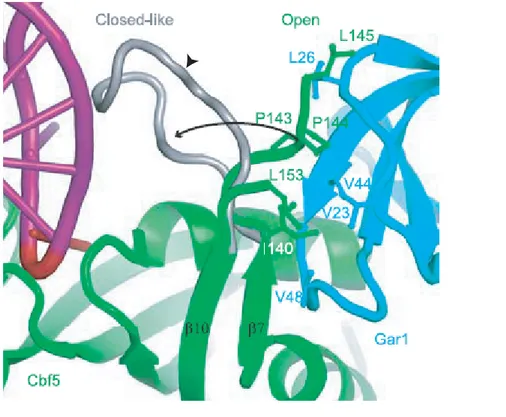
Figure
+7
Documents relatifs