L'encodage de la forme par le toucher haptique
Texte intégral
Figure
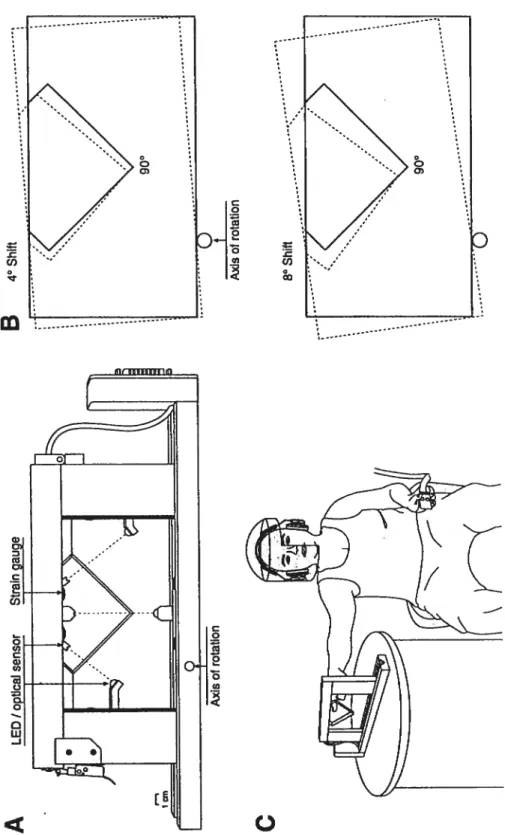


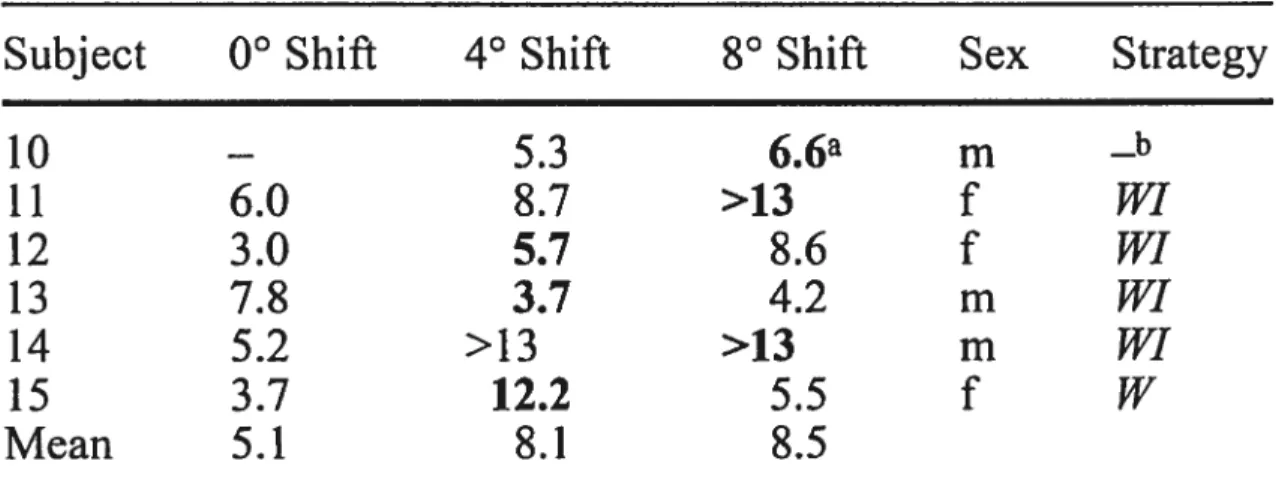

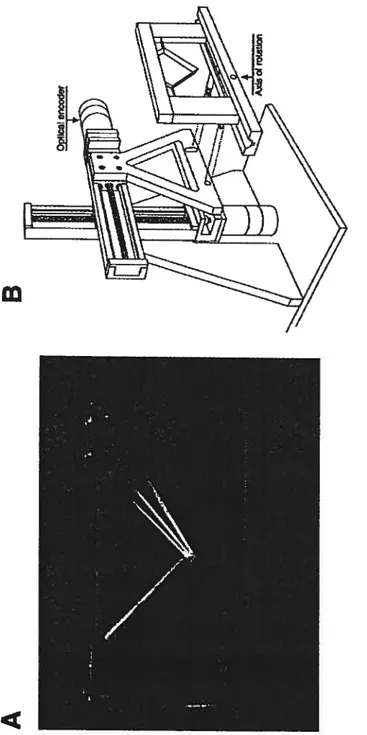

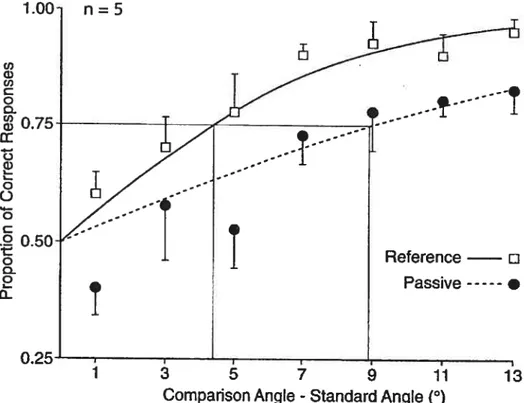
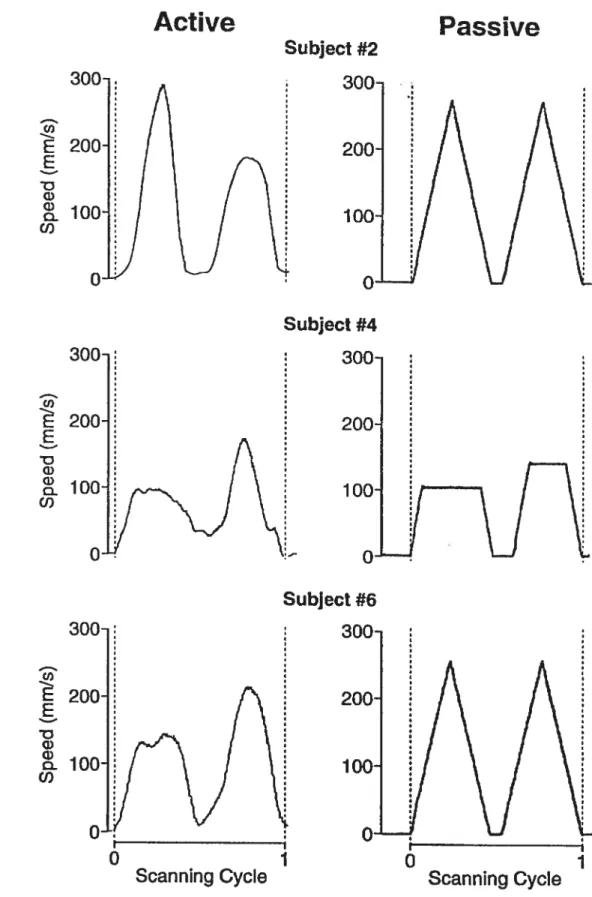
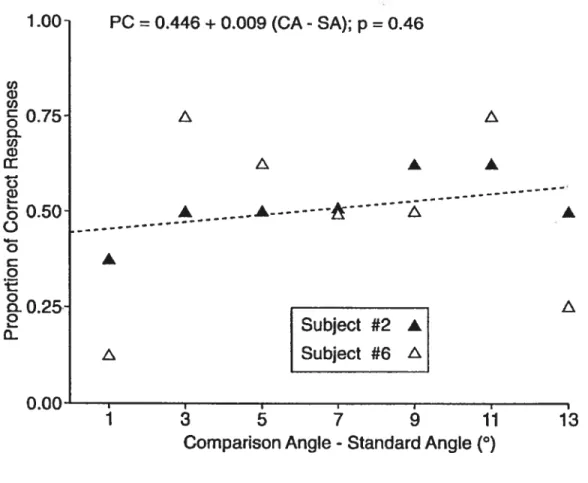
Documents relatifs
A l’analyse par hexagone, l’amplitude et la latence pour l’œil droit et pour l’œil gauche de chaque sujet étudié ont été comparées avec la moyenne de l’amplitude et de
En application des dispositions du chapitre IV du titre préliminaire du livre premier du code de l’urbanisme et sur la base des informations fournies par la personne
En application des dispositions du chapitre IV du titre préliminaire du livre premier du code de l’urbanisme et sur la base des informations fournies par la personne
Sur la base des informations fournies par la personne publique responsable, la procédure de modification n°4 du plan local d’urbanisme intercommunal (PLUi) de l’agglomération
• au vu de l’ensemble des informations fournies par la personne publique responsable, le projet de révision du zonage d’assainissement des eaux usées de la
En application de la section deux du chapitre II du titre II du livre premier du code de l’environnement, et sur la base des informations fournies par la personne publique
En application de la section deux du chapitre II du titre II du livre premier du code de l’environnement, et sur la base des informations fournies par la personne publique
En application de la section deux du chapitre II du titre II du livre premier du code de l’environnement, et sur la base des informations fournies par la personne publique
