Déterminants intrinsèques et extrinsèques du
comportement de vigilance et ses conséquences sur la
valeur adaptative des femelles de la chèvre de
montagne
Mémoire
Florent Déry
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Déterminants intrinsèques et extrinsèques du
comportement de vigilance
et ses conséquences sur la valeur adaptative
des femelles de la chèvre de montagne
Mémoire
Florent Déry
Sous la direction de :
Sandra Hamel, codirectrice de recherche
Steeve D. Côté, directeur de recherche
Résumé
La vigilance permet de détecter des menaces de congénères et de prédateurs pour les éviter, mais peut aussi diminuer la valeur adaptative en réduisant le temps alloué à d’autres activités améliorant la performance individuelle. J’ai étudié les facteurs extrinsèques et intrinsèques qui modulent la vigilance ainsi que son effet sur la valeur adaptative des femelles adultes marquées de la population de chèvres de montagne (Oreamnos
americanus) de Caw Ridge (Alberta, Canada). J’ai cerné les facteurs influençant la vigilance
à partir d’observations cibles récoltées sur 12 ans. J’ai ensuite utilisé une approche par modèle multivarié bayésien pour estimer la corrélation latente au niveau individuel entre trois traits de vigilance et un trait de la valeur adaptative. Ma première hypothèse était que l’ajustement de la vigilance dépend soit du risque de prédation à lui seul ou soit du compromis entre les besoins nutritifs et le risque de prédation. Ma deuxième hypothèse était que la vulnérabilité du chevreau influence la vigilance de sa mère. Finalement, je m’attendais à ce que la vigilance favorise la survie annuelle des femelles adultes et de leur chevreau. J’ai observé que les femelles étaient plus vigilantes en forêt ouverte, lorsqu’elles étaient isolées de leurs voisins et quand le risque d’interactions augmentait, une fois l’effet confondant des voisins pris en compte. Les femelles reproductrices étaient plus vigilantes et leurs alertes tendaient à s’allonger quand leur chevreau était plus loin. La fréquence et le temps total en alerte des mères étaient positivement corrélés à la probabilité de survie de leur chevreau, mais la vigilance n’était pas corrélée à la survie des femelles adultes. Mes résultats indiquent que le risque de prédation et la vulnérabilité du chevreau sont les mécanismes modulant la vigilance et que cette tactique améliore probablement la valeur adaptative des femelles car leurs jeunes survivent mieux.
Abstract
Vigilance allows animals to quickly detect threats from conspecifics and predators to avoid or minimize costly encounters, but it can often limit other fitness-enhancing activities, such that fitness returns are expected when vigilance is properly adjusted. I studied drivers of vigilance and the link between vigilance and fitness in a wild population of individually marked mountain goats (Oreamnos americanus) at Caw Ridge (Alberta, Canada). I first investigated the proximate factors influencing vigilance by using focal observations recorded over 12 years. Then I used a joint modelling framework to estimate the latent correlations at the individual level among three vigilance traits and two fitness components. I first hypothesized that vigilance adjustment would either be influenced by predation risk only or by the trade-off between predation risk and daily energetic requirements. My second hypothesis was that offspring vulnerability would drive mother’s vigilance. Finally, I hypothesized that vigilance would favor annual survival of adult females and of their offspring. I observed that females increased vigilance in open forest, when they were not surrounded by conspecific neighbours, and when the risk of social interactions increased, once accounting for a cofounding effect of surrounding neighbours. Alerts of reproductive females were more frequent and longer than alerts of females without offspring. Mothers also tended to perform longer alerts when their offspring was >10 meters away than at shorter distances. There was no association between vigilance and annual survival of adult females, whereas offspring from mothers that were more often alert and spent more time vigilant survived better. Predation risk and offspring vulnerability were the main mechanisms behind variation in vigilance. My results indicate that vigilance is driven both by predation risk and offspring vulnerability, and that this behavioral tactic likely improved adult female fitness by increasing their offspring’s chances of survival.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des figures ... vi
Liste des tableaux ... viii
Liste des annexes ... ix
Remerciements ... x
Avant-propos ... xii
Introduction ... 1
Risque de prédation et tactiques anti-prédatrices ... 1
Le comportement de vigilance ... 2
Déterminants extrinsèques de la vigilance ... 3
Déterminants intrinsèques de la vigilance ... 5
Contexte de l’étude... 7
Objectifs et hypothèses ... 10
Aire d’étude ... 10
Suivi à long terme et observations comportementales ... 11
Chapitre 1: Linking proximate drivers and fitness returns of vigilance: offspring from more vigilant mothers survive better ... 13
1.1 Résumé ... 13
1.2 Abstract ... 13
1.3 Introduction ... 14
1.4 Methods ... 16
1.4.1 Study area and captures ... 16
1.4.2 Reproduction and survival ... 17
1.4.3 Behavioural observations ... 18 1.4.4 Predation risk ... 20 1.4.5 Statistical Analyses ... 20 1.5 Results ... 24 1.6 Discussion ... 28 1.7 Acknowledgements ... 32 Conclusion ... 33
Perspectives de recherche ... 37 Bibliographie ... 39 Annexe A ... 48 Annexe B ... 49 Annexe C ... 50 Annexe D ... 51 Annexe E ... 52
Liste des figures
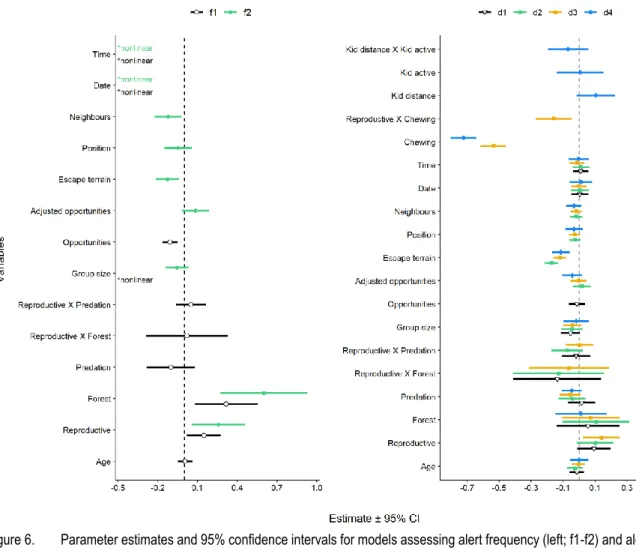
Figure 1. Illustration d’une chèvre de montagne en comportement d'alimentation (A) et de vigilance (B), ainsi que des mesures différentes pour quantifier la vigilance (C; les séquences noires représentent des alertes). .. 2 Figure 2. Observations de prédateurs et taille de la population de chèvres de montagne à Caw Ridge (Alberta, Canada; 1989 – 2018). A) Cougar responsable de la prédation du chevreau #456 de la chèvre #354; 1er juillet 2015. B) Loup transportant le chevreau de la chèvre #413; 13 juillet 2015. C) Mère grizzly et son jeune s’alimentant de racines; 24 mai 2015. D) Variation annuelle (1989-2018) de la fréquence d'occurrence des observations de prédateurs (axe de gauche et barres verticales) et la taille maximale de la population de chèvres de montagne de Caw Ridge (axe de droite et courbe noire). La fréquence est comptabilisée de façon à ce qu’une observation de prédateur compte pour une unité, peu importe si plusieurs prédateurs sont observés ensemble (p. ex. une observation d’une meute de sept loups est comptabilisée comme une occurrence de loups). La catégorie « Ours » regroupe les grizzlis et les ours noirs, la catégorie « Canidés » regroupe les coyotes et les loups, et la catégorie « Autres » regroupe les autres prédateurs tels que le carcajou (Gulo gulo) et le lynx du Canada (Lynx canadensis). ... 9 Figure 3. Vue aérienne de l'aire d'étude de Caw Ridge (Alberta, Canada), à partir de l’ouest. ... 10 Figure 4. Scénario d’observation cible de la femelle #440 dans un groupe de neuf chèvres (A). L’environnement social de #440 à ce moment précis se définirait comme suit : #440 est dans un groupe de neuf chèvres avec un voisin dans son rayon de 4m et elle est en position périphérique (B). ... 11 Figure 5. Sampling design across time showing the structure of the datasets in relation with the independent variables available. A) Annual variation in occurrence of predator sightings (number of predator sightings reported per 100 field days; connected solid black dots, left y-axis) and in focal observations sample size (grey bars, right y-axis). B) Structure of the datasets highlighting the independent variables available over the years (listed in the white boxes) in relation with the vigilance responses evaluated (listed in the black boxes; data #1 = 411 focal obs. on 115 individuals; data #2 = 330 focal obs. on 91 individuals; data #3 = 135 focal obs. on 21 individuals).* Variables only available for reproductive females, which were assessed in model d4 that included only vigilance response of reproductive females. ... 19 Figure 6. Parameter estimates and 95% confidence intervals for models assessing alert frequency (left; f1-f2) and alert duration (right; d1-d4) in adult female mountain goats at Caw Ridge (AB, Canada), 1994―2018. Reproductive represents the difference for reproductive females compared with barren females (reference level). Forest represents the difference when a female was in open forest compared to alpine habitat (reference level). Predation is the number of predator sightings reported per 100 field days. Opportunities are the opportunities for social interactions and are adjusted for the number of neighbours when both variables are included in a model (see Methods). Neighbours is the number of neighbours within a 4 m radius. Position is spatial position within the group (ranging from 0 to 1, with 1 = central, 0.5 = mid-periphery, 0 = periphery). Escape terrain is the distance to nearest escape terrain. Time is the number of minutes until sunset. Estimates for variables with a nonlinear influence are illustrated in Annexe B, and intercept estimates and variance for random effects and residuals are given in Annexe C. ... 26 Figure 7. Effects of habitat, distance to escape terrain, number of neighbours, reproductive status, and mother-kid distance on vigilance of adult female mountain goats at Caw Ridge (AB, Canada), 1994―2018. A) Gardner-Altman plot (Ho et al., 2019) illustrating the effect of habitat type on alert frequency (“open” = alpine habitat, green, “forest” = open forest, orange). B) Effect of distance to escape terrain on alert frequency (curve and green zone present predictions ± 95% confidence interval (CI) from model f2). C) Gardner-Altman plot illustrating the effect of reproductive status on alert frequency (“Barren” = barren females, light blue; “Repro.“ = reproductive females, dark blue). D) Effect of the numbers of neighbours on alert frequency, with green dots and overlapping vertical lines (left of boxplots) presenting predicted values ± 95% CI from models f2. Boxplots are summarizing the raw data and the grey dots are the only 3 observations for “4 neighbours”. The lines in the boxes represent medians, and boxes represent lower to upper quartile ranges, with whiskers extending up to 1.5 times the
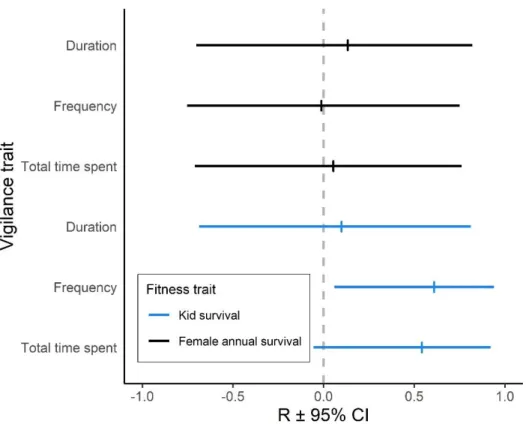
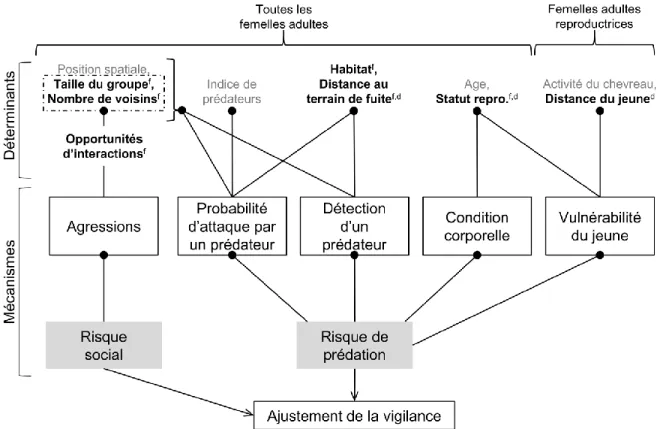
interquartile range and data beyond that distance represented by black dots. E) Effect of distance to escape terrain on alert duration (curve and green zone present predictions ± 95% CI of model d2). F) Effect of mother-offspring distance on alert duration for reproductive females (curve and blue zone present predictions ± 95% CI from model d4). For all panels, dots or squares represent raw data. ... 27 Figure 8. Correlations (R 95% confidence interval (CI)) at the individual level between each vigilance trait – alert duration, alert frequency, and total time spent alert – and two measures of fitness – adult female (black) and offspring survival to one year (blue) – in mountain goats at Caw Ridge (AB, Canada), 1994―2018. ... 28 Figure 9. Schéma conceptuel résumant les relations entre les variables d’intérêt mesurées et les mécanismes influençant l’ajustement de la vigilance des femelles de la chèvre de montagne de Caw Ridge (Alberta, Canada, 1994-2018). Les variables en gras sont celles dont l’intervalle de confiance à 95% excluait ou chevauchait légèrement zéro. Les lettres en indice indiquent quel trait de la vigilance était influencé par une variable (f : fréquence des alertes, d : durée d’une alerte). Les variables en gris sont celles n’ayant pas influencées le comportement de vigilance. « Statut repro. » signifie statut reproducteur. La date et l’heure de la journée ont aussi influencé la fréquence des alertes mais ces deux variables ne sont pas schématisées car elles ont été incluses à titre de co-variables. ... 34
Liste des tableaux
Liste des annexes
Means, standard deviations and transformations of variables of each dataset included in models f1, f2, d1, d2, d3, and d4. Mean (bold, first line) and standard deviation (second line) used to standardize each continuous independent variable included in the six models analysing vigilance, with the third line in italic indicating when a variable was transformed with the type of transformation. Dashes represent cases where the variable was not included in the analysis and/or was not transformed. ... 48 Predictions ± 95% confidence interval (CI) for variables having a nonlinear influence on alert frequency (models f1, left panels, and f2, right panels; colors are consistent with models presented in figure 6). Time is hours until sunset. ... 49 Intercepts and variance estimates of random effects for the six vigilance models. ... 50 Mean (bold, first line) and standardized deviation (second line) of continuous independent variables in each dataset included in multivariate Bayesian models of vigilance and fitness relationships (m1 and m2). Dashes represent cases where the variable was not included in the analysis. ... 51 Variance-covariance matrix of vigilance and fitness traits of (A) multivariate model (m1) of vigilance traits and annual survival of adult female, and B) multivariate model (m2) of vigilance traits and kid survival to one year old. The diagonals of the matrices represent the trait variances (italic), latent correlations between traits are above the diagonals, and trait covariance are given below the diagonals. Each estimate is given at the individual level. ... 52
Remerciements
J’ai eu la chance de partager mes embuches, mes succès, mes doutes et mes niaiseries avec une brochette bien garnie de gens que j’apprécie énormément. Je tiens à les remercier d’avoir contribué à mon cheminement de façon directe ou indirecte.
Tout d’abord, merci à Sandra et Steeve de m’avoir encadré pendant mes travaux au cours des dernières années. J’ai eu la chance de vous connaître sous une autre couture dès le début, à Caw Ridge en 2015, et vous m’avez transmis la passion qu’il fallait pour entreprendre mes études graduées. Les centaines d’histoires rocambolesques et les étoiles dans vos yeux sur le terrain ont contribuées à faire grandir mon sentiment d’appartenance pour Caw Ridge pendant les quatre étés où j’y ai travaillé.
Sandra, ta rigueur, ton écoute et ta disponibilité (même en Norvège!) auront été les moteurs de ma réussite. Tu as su canaliser mon énergie et ma créativité, une tâche à la fois, m’aidant ainsi à tracer le chemin de ma maîtrise. Ton énergie, ton positivisme, tes conseils sur l’équilibre vie-travail, tes blagues et expressions, nos riches discussions et la position dans laquelle tu laves la vaisselle dans la cabane à Caw Ridge m’auront marqué au fer rouge. Collaborer avec toi fut un honneur !
Steeve, merci de m’avoir donné la chance de participer à ce projet, de m’avoir fait confiance tant au niveau de la recherche qu’en tant que responsable sur le terrain. L’effet papillon de cette opportunité fait en sorte que ma vie est maintenant scindée en deux parties : l’avant et l’après Caw Ridge. Je suis très fier d’avoir pu contribuer à l’étude à long terme de Caw, à laquelle j’ai tenté de laisser une empreinte indélébile de mon passage. Merci de m’avoir fait connaître la brown ale de la brasserie Big Rock, et je reste sceptique sur tes observations de #386 en 2017 et 2018.
Merci aux nombreux coéquipiers qui ont fait honneur à mes muffins aux bananes sur le terrain, en ordre de saison de terrain : mes goat masters, Andrea et Édouard, ma marmot
princess Béatrice, et les « chèvriers.ères » et « marmotteux.ses » Giulia, Frédéric, Sonia,
Dominik, David, Marie-Ève. Vous êtes responsables des fous-rires, des bonheurs, des quelques heures passées à réparer des quatre-roues, quelques lendemains sur la petite
gear, mais surtout de mes bons souvenirs sur le terrain.
Je suis extrêmement reconnaissant pour l’aide de mes collègues au labo, que ce soit en me posant des questions à m’en gratter le cuir chevelu au vif, en discutant de mes problèmes,
d’analyses, de faits divers ou encore de la prochaine occasion pour fêter. Mentions spéciales (en ordre alphabétique) à Alexis, Béatrice, Emilie, Marianne, Martin, Michaël et Sabrina pour avoir lu mes travaux, les avoir commentés, pour avoir assisté à mes présentations et répondu à mes questions durant mon parcours. J’ai eu la chance de développer de belles relations d’amitié; des relations que j’entends bien entretenir !
J’ai pu compter sur bon nombre de personnes-ressources durant ma maîtrise. Merci à Jean-Pierre Tremblay et Nadia Aubin-Horth d’avoir commenté mes travaux lors d’étapes cruciales à mon cheminement et de faire partie du jury pour l’évaluation de ce mémoire. Merci au personnel de soutien du département de biologie, pensée spéciale à Josée et Marie-Soleil qui ont répondu à mes innombrables questions. Réaliser mes travaux aurait été impossible sans l’aide des professionnels de recherche Caroline, Julien, Catherine et Marie-Claude. Merci à vous pour le support logistique avant, durant et après mes étés sur le terrain, votre travail est essentiel et j’en suis très reconnaissant.
Merci à ma famille pour leur soutien, leur écoute, leur aide, leur intérêt, ou simplement d’avoir arrêté de me demander « Pis? Quand est-ce ‘tu finis? ». Merci Mom - pour le support, l’aide, la bouffe (même si c’était tout le temps trop!), m’avoir enduré débarquer à la maison puis passer une fin de semaine complète à pigrasser dans le garage et dévaliser le frigo à maintes reprises.
Finalement, merci à Béatrice, ma compagne, mon p’tit paintbrush sur lequel je suis tombé au début de cette aventure, avant même d’avoir en tête une maîtrise. Merci d’avoir chamboulé ma vie dès notre premier été, de m’inonder de ton intelligence, ta bonne humeur, ton audace et ton support. Je ne sais pas comment tu as fait pour m’endurer durant les derniers mois de ma maîtrise. Merci de m’avoir encouragé, de me faire rire, sourire et me donner la force de surmonter toutes les embuches que j’ai rencontré. On forme une belle équipe et sans toi mon parcours aurait été radicalement différent.
Avant-propos
Ce mémoire aborde quels déterminants intrinsèques et extrinsèques modulent l’ajustement de la vigilance et comment l’ajustement de ce comportement anti-prédateur influence l’aptitude phénotypique des femelles adultes de la chèvre de montagne. Le mémoire comporte trois sections : une introduction générale, le chapitre principal sous la forme d’un article scientifique rédigé en anglais, et une conclusion générale. Le mémoire inclus également des annexes qui seront jointes à l’article scientifique qui sera soumis dans le cadre de mes travaux de maîtrise.
Je soumettrai le chapitre principal de ce mémoire au processus de publication scientifique. J’ai mené la conception, les analyses statistiques, l’interprétation des résultats et la rédaction de ce chapitre en tant que premier auteur en collaboration avec ma codirectrice Sandra Hamel (professeure agrégée au Département de biologie de l’Université Laval) et mon directeur Steeve D. Côté (professeur titulaire au Département de biologie de l’Université Laval; Tableau 1).
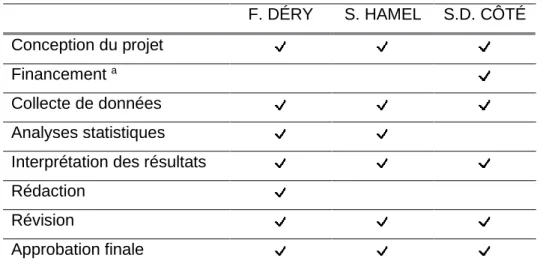
Tableau 1. Implications des coauteurs pour l’article présenté dans le mémoire.
F. DÉRY S. HAMEL S.D. CÔTÉ Conception du projet
Financement a Collecte de données Analyses statistiques Interprétation des résultats Rédaction
Révision
Approbation finale
a Mon projet a été financé par les fonds de recherche de S. D. Côté obtenus principalement via le
programme de subventions à la découverte du Conseil de recherches en sciences naturelles et génie du Canada (CRSNG).
Introduction
Risque de prédation et tactiques anti-prédatrices
L’étude des déterminants de la dynamique des populations est un thème central en écologie. Parmi ces facteurs, la prédation peut réguler la démographie (Sinclair, 1985; Skogland, 1991) en ayant un impact sur le recrutement (Gaillard, Festa-Bianchet, & Yoccoz, 1998) ou, dans de plus rares cas, en affectant la survie des adultes (p. ex. Grange et al., 2015). En plus des conséquences au niveau populationnel, la présence de prédateurs dans un environnement a d’abord des répercussions au niveau individuel, notamment en modulant le comportement d’une proie. Comme la prédation est un facteur pouvant directement affecter la survie et la valeur adaptative d’un individu, le risque de prédation dans un environnement donné pourrait favoriser le développement de tactiques comportementales constituant la stratégie anti-prédatrice d’un individu. Chaque tactique comporte des coûts et bénéfices et, pour qu’une tactique soit adaptative, ses coûts doivent être inférieurs à ses bénéfices. Ultimement, le bilan du compromis de l’ensemble des tactiques sélectionnées devrait augmenter la valeur adaptative d’une proie en présence de prédateurs.
Les tactiques comportementales anti-prédatrices sont nombreuses et varient selon l’échelle temporelle considérée et selon leurs conséquences sur la valeur adaptative. Par exemple, la fuite (M. Altmann, 1958) est une tactique de dernier recours qui a le potentiel d’augmenter les chances de survie et donc de procurer des bénéfices majeurs par rapport à un investissement minimal de temps à long terme. À l’inverse, sélectionner des lieux à faible risque de prédation se fait sur une période temporelle étendue, ce qui peut limiter l’utilisation d’habitats de meilleure qualité d’un point de vue nutritif ou énergétique (Dussault et al., 2005) et ainsi diminuer la condition corporelle et potentiellement la valeur adaptative de l’individu. La vie en groupe peut occasionner des coûts en augmentant la probabilité d’être repéré par un prédateur (Ruxton & Johnsen, 2016) ou en limitant l’acquisition des ressources individuelles due à la compétition entre les individus du groupe (Black, Carbone, Wells, & Owen, 1992; Gompper, 1996; Sansom, Cresswell, Minderman, & Lind, 2008). Néanmoins, se regrouper permet à un individu de diminuer le risque de prédation car celui-ci peut obtenir de l’information sur la présence de prédateurs à partir de ses congénères (Beauchamp, 2010). Vivre en groupe diminue aussi le risque de mortalité en réduisant la probabilité individuelle d’être attaqué par un prédateur (Lima & Dill, 1990). Également, vivre en groupe
permet de diminuer le temps passé en vigilance, un état d’alerte interne mesurable voué à la détection de menaces dans l’environnement (Beauchamp, 2015) pouvant favoriser la survie d’un individu ou de sa progéniture, et ainsi sa valeur adaptative.
Le comportement de vigilance
Il y a deux fonctions à la vigilance : surveiller ses congénères et détecter les prédateurs. La vigilance sociale permet de surveiller ses voisins et ainsi obtenir de l’information sur le risque de prédation (van der Post, de Weerd, Verbrugge, & Hemelrijk, 2013; Zhao, Lyu, Sun, & Zhou, 2019), les ressources disponibles (Beauchamp, 2001), ou encore éviter une interaction intra-spécifique potentiellement coûteuse (Treves, 2000). La vigilance anti-prédatrice quant-à-elle permet de détecter directement les prédateurs, et ainsi réduit les chances d’être attaqué par un prédateur (FitzGibbon, 1989) ou diminue le temps de réaction pour fuir à temps (Lima, 1994).
Malgré ces bénéfices, le temps passé en vigilance peut diminuer le temps disponible à d’autres activités essentielles telles que l’alimentation (Creel, Schuette, & Christianson, 2014) ou le toilettage (Blanchard, Pays, & Fritz, 2017). Ainsi, la valeur adaptative d’un individu pourrait être réduite si celui-ci alloue trop de temps à la vigilance. Devant cette contrainte, il existe une variation individuelle dans l’allocation du temps à la vigilance et la distribution des alertes pour une période de temps donnée. Bien entendu, il devient alors nécessaire de quantifier la vigilance en mesurant la durée de chaque alerte, la fréquence de ces alertes, ou encore le temps total passé en vigilance (Fig. 1). La présence d’un compromis opposant l’alimentation à la vigilance implique également que ces comportements soient mutuellement exclusifs. Dans de tels cas, être plus vigilant peut réduire le taux d’alimentation (Barnier et al., 2016) et potentiellement affecter la condition physique. Or, chez les grands herbivores, combiner le temps en alerte
Figure 1. Illustration d’une chèvre de montagne en comportement d'alimentation (A) et de vigilance (B), ainsi que des mesures différentes pour quantifier la vigilance (C; les séquences noires représentent des alertes).
avec la mastication permettrait de réduire une partie du coût en temps de la vigilance (Fortin, Boyce, Merrill, & Fryxell, 2004; Illius & Fitzgibbon, 1994). Néanmoins, les alertes combinées à la mastication sont moins efficaces (Blanchard & Fritz, 2007) car le bruit produit en mâchant réduit vraisemblablement la probabilité de détection auditive d’un prédateur (Molinari-Jobin, Molinari, Loison, Gaillard, & Breitenmoser, 2004).
Peu d’études ont réellement évalué les conséquences de la vigilance sur la valeur adaptative, probablement parce que ceci nécessite un suivi individuel en milieu naturel et des données comportementales répétées au cours de la vie d’un animal (Beauchamp, 2015). Effectivement, la relation entre la vigilance et la valeur adaptative a reçu peu d’attention, particulièrement chez les grands herbivores (Say-Sallaz, Chamaillé-Jammes, Fritz, & Valeix, 2019). Néanmoins, deux études ont explicitement testé cette relation chez le chevalier gambette (Tringa totanus) et la perdrix grise (Perdix perdix). Les chevaliers qui passaient plus de temps en vigilance dans les habitats de qualité et à haut risque de prédation avaient une plus grande probabilité de survie, mais cette tendance s’inversait dans les habitats de faible qualité et risque de prédation (Sansom, Lind, & Cresswell, 2009). Les perdrix plus vigilantes durant la période de couvée avaient un taux de survie inférieur, et ce même si la vigilance réduit le risque de prédation à court terme (Watson, Aebischer, & Cresswell, 2007). Pour ces deux études, l’effet négatif de la vigilance sur la survie s’expliquerait par la diminution du temps passé à s’alimenter, ce qui augmentait le risque de mourir d’inanition, ou par une baisse de la condition corporelle augmentant la vulnérabilité à la prédation (Sansom et al., 2009; Watson et al., 2007). Il reste que l’étude du lien entre la vigilance et la valeur adaptative a reçu peu d’attention, probablement due au fait que ces deux traits ne sont pas mesurés sur la même échelle temporelle et varient en fonction de nombreux déterminants, ceux-ci formant d’ailleurs la majorité de la littérature sur la vigilance.
Déterminants extrinsèques de la vigilance
La présence de prédateurs et l’habitat d’une proie sont des déterminants majeurs de l’ajustement de la vigilance puisqu’ils créent un paysage de la peur (Brown, 1999). Plus le nombre de prédateurs est élevé, plus la probabilité de rencontre augmente. Les proies augmentent donc leur taux de vigilance dans les situations à haut risque, par exemple lorsqu’ils détectent un prédateur (Eccard, Meißner, & Heurich, 2017) ou après la
réintroduction de prédateurs dans un site (Hunter & Skinner, 1998; Laundré, Hernández, & Altendorf, 2001). Selon l’écologie d’une proie et de son prédateur, le niveau de vigilance d’une proie varie en fonction de la visibilité associée à un habitat. Lorsque la visibilité est réduite, la probabilité d’une proie à détecter un prédateur diminue (Bednekoff & Blumstein, 2009; Whittingham, Butler, Quinn, & Cresswell, 2004). Ces habitats sont également davantage utilisés par les prédateurs chassant en embuscade (p. ex. le cougar, Puma
concolor; Pierce & Bleich 2003). Ainsi, Ebensperger et Hurtado (2005) ont montré que le
dègue du Chili (Octodon degus) est plus vigilant lorsqu’il est entouré de hautes herbes, en plus d’augmenter l’intensité de sa vigilance en adoptant une position bipède plutôt que quadrupède. Chez les ongulés, le springbok (Antidorcas marsupialis) est plus vigilant loin des arbustes (Burger, Safina, & Gochfeld, 2000) et le mouflon méditerranéen (Ovis gmelini
musimon x Ovis sp.) est plus vigilant dans les habitats à visibilité réduite (Benoist, Garel,
Cugnasse, & Blanchard, 2013). L’ajustement de la vigilance peut aussi dépendre de la probabilité de rencontrer un type de prédateur dans un habitat donné (Scannell, Roberts, & Lazarus, 2001). Les individus dont le patron de vigilance est régulier optimisent la détection de prédateurs « coureurs », qui attaquent de façon aléatoire, alors que les individus dont la vigilance est irrégulière ont plus de chances de détecter un prédateur embusqué (Scannell et al., 2001). Effectivement, lorsque la variabilité dans l’intervalle entre les alertes est imprévisible, il devient plus difficile pour un prédateur embusqué d’identifier le moment optimal pour lancer une attaque.
Lorsqu’un habitat offre un refuge protégeant une proie, la vigilance est généralement positivement corrélée à la distance à cet habitat. Ainsi, les marmottes des Rocheuses (Marmotta caligata) et les marmottes à ventre jaune (Marmotta flaviventris) sont plus souvent en alerte lorsqu’elles sont loin de leur terrier (Holmes, 1984; Monclús, Anderson, & Blumstein, 2015). Les ongulés alpins sont aussi généralement plus vigilants lorsqu’ils sont loin d’un terrain de fuite, c’est-à-dire des pentes abruptes et des falaises (Frid 1997; mais voir Hopewell et al., 2005), ces habitats étant généralement associés à un risque de prédation plus bas.
Chez les espèces grégaires, la position spatiale dans un groupe, la taille du groupe et le nombre de voisins à proximité sont des déterminants socio-environnementaux qui interagissent de concert pour influencer la vigilance. Les individus en périphérie d’un groupe s’exposent à un plus grand risque de prédation si le prédateur utilise une tactique de prédation marginale en attaquant par l’extérieur (Hamilton 1971; Krause 1994).
Conséquemment, les individus en marge d’un groupe sont généralement plus vigilants (Beauchamp, 2007), tel qu’observé chez le vacher à tête brune (Molothrus ater; Fernández-Juricic & Beauchamp 2008) et le springbok (Burger et al., 2000). La vigilance d’un individu diminue généralement avec la taille du groupe car les chances de détecter un prédateur sont plus élevées dans les plus grands groupes (Beauchamp, 2017; Pulliam, 1973) et le risque de prédation est dilué (Beauchamp, 2017; Pulliam, 1973). Cet effet a été observé chez un grand nombre de taxons chez les oiseaux et les mammifères (revu par Beauchamp 2008, 2019). Le temps alloué à la vigilance dépend aussi des congénères à proximité d’un individu (Bertram, 1980). Effectivement, un individu peut diminuer le temps passé en vigilance s’il est entouré de voisins puisque l’information produite par un voisin vigilant ou qui fuit lui signale la présence d’un prédateur (van der Post et al., 2013).
Bien que le temps passé en vigilance puisse diminuer dû aux bénéfices de l’environnement social d’une proie, la compétition intraspécifique peut quant-à-elle augmenter le temps passé en vigilance (Beauchamp, 2014). La relation entre la vigilance et la taille du groupe peut s’inverser ou se stabiliser passé un certain seuil, particulièrement si les voisins posent une menace pouvant mener à des blessures ou à la mort (Treves 2000). La compétition pour des sites de qualité ou des ressources alimentaires peut pousser un individu à être plus vigilant pour éviter ces interactions potentiellement coûteuses (Treves, 2000) ou pour défendre ses ressources. La visibilité peut aussi diminuer lorsqu’il y a plus de voisins à proximité, forçant un individu à être plus vigilant pour surveiller le comportement de son voisin afin de récolter de l’information sur le risque de prédation (Zhao et al., 2019). La vigilance peut aussi augmenter dans les groupes plus nombreux à des fins de chapardage : en surveillant les producteurs, le chapardeur a accès à de l’information sur l’emplacement et la quantité de ressources alimentaires (Beauchamp, 2001).
Déterminants intrinsèques de la vigilance
Le statut reproducteur est l’un des plus importants facteurs expliquant la variation individuelle du comportement de vigilance. Le coût énergétique des soins parentaux peut contraindre le temps alloué à la vigilance (Beauchamp, 2015). Chez les mammifères, l’allaitement peut entrainer un compromis entre l’alimentation et la vigilance (Fortin et al., 2004; Illius & Fitzgibbon, 1994) puisque les femelles pourraient devoir passer plus de temps en alimentation afin de compenser pour cette contrainte énergétique (Ruckstuhl, Festa-Bianchet, & Jorgenson, 2003). Alternativement, le niveau de vigilance d’une femelle qui allaite pourrait demeurer stable si celle-ci passe le même temps en alimentation qu’une
femelle sans jeune tout en augmentant son rythme d’alimentation (Gélin, Wilson, Coulson, & Festa-Bianchet, 2013), ou encore si elle alloue plus de temps à l’alimentation aux dépends d’une autre activité (p. ex. le repos; Hamel & Côté 2008).
La vulnérabilité du jeune peut aussi causer une augmentation du temps passé en vigilance chez les femelles reproductrices (Burger & Gochfeld, 1994; Caro, 1987). Chez les ongulés, les femelles sont généralement plus vigilantes lorsqu’elles ont un jeune que lorsqu’elles sont seules (Burger & Gochfeld, 1994). L’augmentation du temps alloué en vigilance serait donc un soin maternel qui devrait permettre de favoriser la survie du jeune et ainsi la valeur adaptative de la femelle (Beauchamp, 2015). Puisque la capacité de fuite augmente avec l’âge d’un jeune, les mères pourraient être moins vigilantes lorsque leur jeune devient moins vulnérable en vieillissant, tel qu’observé chez le guépard (Acinonyx jubatus; Caro 1987). Dépendamment de la relation définissant le lien mère-jeune, l’activité du jeune et la distance le séparant de sa mère peut également influencer l’ajustement de la vigilance chez les femelles reproductrices (Lent 1974). Dans le cas où le jeune suit sa mère et que celle-ci peut le défendre d’une éventuelle attaque de prédateur ou de congénères (stratégie de talonnage; Lent 1974), la mère devrait être plus vigilante lorsque son jeune est loin. Si la mère et le jeune ne sont unis que pour la durée de l’allaitement et que le jeune reste immobile et camouflé lors des autres périodes de la journée (stratégie de « cachette »; Lent 1974). Ainsi, les mères dont le jeune est camouflé dans l’environnement devraient être plus vigilantes lorsque leur jeune est actif car il est plus facile à détecter par les prédateurs, tel qu’observé chez le bouquetin d’Espagne (Capra pyraneica; Alados & Escos 1988). Néanmoins, le lien explicite entre la vigilance en tant que soin maternel et la survie du jeune reste peu exploré dans la littérature.
De plus, en altérant l’état de satiété et la vulnérabilité à la prédation d’un individu, la condition corporelle influence la vigilance. Les individus en mauvaise condition doivent passer plus de temps à acquérir des ressources énergétiques que les individus en bonne condition, ce qui limite le temps disponible pour la vigilance (Bachman 1993). À l’opposé, si les individus en meilleur condition sont simplement ceux qui allouent davantage de temps à l’alimentation, ces individus auraient un taux de vigilance inférieur car le temps à s’alimenter devrait réduire le temps disponible à la vigilance. D’ailleurs, un individu en bonne condition est généralement moins vulnérable à la prédation, ce qui pourrait aussi expliquer un niveau de vigilance inférieur à celui d’un individu en mauvaise condition (Brivio, Grignolio, Brambilla, & Apollonio, 2014). Une plus faible disponibilité des ressources peut aussi
augmenter le temps nécessaire à l’acquisition de ressources et affecter la vigilance. Par exemple, dans une expérience manipulant la nourriture réalisée avec le cratérope écaillé (Turdoides squamiceps; Wright et al. 2001) et le geai à gorge blanche (Aphelocoma
coerulescens; Bednekoff and Woolfenden 2003), les individus moins bien nourris étaient
moins vigilants que ceux qui étaient mieux nourris. L’effet de la condition corporelle sur la vigilance est intimement relié à l’âge et la croissance. En effet, chez plusieurs espèces (Grue cendrée, Grus grus; Alonso & Alonso 1993; Avilés & Bednekoff 2007; mouflon méditerranéen; Benoist et al., 2013; wapiti, Cervus elaphus; Childress & Lung 2003) les coûts énergétiques élevés associés à la croissance réduisent le temps disponible pour la vigilance au profit du temps passé à s’alimenter. Le manque d’expérience des juvéniles peut aussi faire en sorte qu’ils sont plus ou moins vigilants que les adultes (Hanson & Coss, 2001; Hollén, Clutton-Brock, & Manser, 2008; Li, Monclús, Maul, Jiang, & Blumstein, 2011; Loehr et al., 2005; Loughry, 1992). La personnalité d’un individu peut également influencer le niveau de vigilance d’un individu. Chez une population de kangourous gris (Macropus
giganteus), par exemple, les femelles prudentes passent plus de temps en vigilance que les
femelles téméraires (Edwards, Best, Blomberg, & Goldizen, 2013). Chez le chevalier gambette par contre, la vigilance n’était pas corrélée à l’agressivité ou aux comportements exploratoires (Couchoux & Cresswell, 2011). Pour ces deux espèces, la proportion de la variation du niveau de vigilance expliquée seulement par la témérité était toutefois relativement faible par rapport à l’ensemble des déterminants de la vigilance (Couchoux & Cresswell, 2011; Edwards et al., 2013).
Contexte de l’étude
La prédation est le facteur de mortalité le plus important chez la chèvre de montagne, principalement par le cougar (Fig. 2a), le loup (Canis lupus; Côté, Peracino, & Simard, 1997; Fig. 2b) et le grizzly (Ursus arctos; Côté & Beaudoin, 1997; Fig. 2c). Cette espèce sélectionne des terrains de fuite (Gross, Kneeland, Reed, & Reich, 2002), des habitats qui sont composés de pentes abruptes et de falaises qui permettent aux chèvres de fuir et déjouer les prédateurs (Côté & Festa-Bianchet, 2003). Elles utilisent davantage des milieux alpins où le risque de prédation est plus faible et la probabilité de détection d’un prédateur est plus grande qu’en forêt ouverte (Côté & Beaudoin, 1997; Festa-Bianchet, Urquhart, & Smith, 1994).
Les chèvres de montagne sont aussi grégaires, une tactique comportementale généralement associée à une réduction du risque de prédation. Néanmoins, se regrouper
expose aussi les chèvres de montagne à un risque social puisqu’elles sont très agressives envers leurs congénères et interagissent fréquemment. De plus, leurs cornes sont pointues et acérées, ce qui augmente le risque de blessures ou de mortalité suivant une interaction (Côté, 2000). Une étude a montré que la taille du groupe diminue le temps passé en vigilance jusqu’à un seuil de 12 individus chez la chèvre de montagne (Risenhoover & Bailey, 1985). Bien que la compétition intraspécifique puisse expliquer pourquoi la taille du groupe n’influence plus la vigilance une fois passé un certain seuil (Quenette, 1990), les travaux de Risenhoover et Bailey (1985) n’incluaient que la taille du groupe comme composante sociale. L’effet des voisins à proximité ou des interactions sociales sur la vigilance n’ont pas été pris en compte et pourraient expliquer la présence de ce seuil.
En plus de vivre en groupe dans un environnement intégrant de multiples prédateurs, les chèvres de montagne occupent des habitats au climat rigoureux (Côté & Festa-Bianchet, 2003). En milieu alpin, la courte période de qualité et de disponibilité des ressources alimentaires exerce une contrainte temporelle sur l’acquisition de ressources énergétiques, ce qui pourrait potentiellement exacerber le coût en temps de la vigilance pour la chèvre de montagne. Ainsi, la phénologie des ressources alimentaires en milieu alpin combinée à la pression de multiples prédateurs forme le contexte idéal pour étudier l’ajustement de la vigilance face au compromis entre le risque de prédation et l’alimentation.
Puisque les femelles ongulés produisent généralement qu’un seul jeune par évènement de reproduction, la survie du jeune est le déterminant majeur du succès reproducteur à vie des femelles ongulés (Clutton-Brock, Guiness, & Albon, 1982). La prédation des jeunes peut donc grandement affecter la dynamique de population. Le coût de la reproduction et la vulnérabilité du chevreau à la prédation favorisent également l’utilisation de tactiques anti-prédatrices et alimentaires au niveau individuel. Les juvéniles sont plus vulnérables à la prédation que les adultes (Festa-Bianchet et al., 1994). De plus, puisque les femelles ne produisent qu’un seul jeune par année, qu’elles sont primipares en moyenne à 4.6 ans malgré une maturité sexuelle à 3 ans, et que la reproduction est coûteuse (Côté and Festa-Bianchet 2001; Festa-Festa-Bianchet et al. 2019), la prédation d’un jeune peut avoir une influence considérable sur la valeur adaptative d’une femelle. Par rapport aux femelles sans jeune, les femelles reproductrices s’alimentent dans des habitats plus sécuritaires (Hamel & Côté, 2007), augmentent le temps passé à s’alimenter et à ruminer (Hamel & Côté, 2008) et ont un taux d’alimentation plus rapide (Hamel & Côté, 2009a).
La combinaison de ces tactiques comportementales permet aux femelles de répondre aux besoins énergétiques de l’allaitement. Cependant, les mécanismes de l’ajustement du comportement de vigilance des femelles adultes – la classe d’âge avec le plus d’élasticité
Figure 2. Observations de prédateurs et taille de la population de chèvres de montagne à Caw Ridge (Alberta, Canada; 1989 – 2018). A) Cougar responsable de la prédation du chevreau #456 de la chèvre #354; 1er
juillet 2015. B) Loup transportant le chevreau de la chèvre #413; 13 juillet 2015. C) Mère grizzly et son jeune s’alimentant de racines; 24 mai 2015. D) Variation annuelle (1989-2018) de la fréquence d'occurrence des observations de prédateurs (axe de gauche et barres verticales) et la taille maximale de la population de chèvres de montagne de Caw Ridge (axe de droite et courbe noire). La fréquence est comptabilisée de façon à ce qu’une observation de prédateur compte pour une unité, peu importe si plusieurs prédateurs sont observés ensemble (p. ex. une observation d’une meute de sept loups est comptabilisée comme une occurrence de loups). La catégorie « Ours » regroupe les grizzlis et les ours noirs, la catégorie « Canidés » regroupe les coyotes et les loups, et la catégorie « Autres » regroupe les autres prédateurs tels que le carcajou (Gulo gulo) et le lynx du Canada (Lynx canadensis).
sur les paramètres démographiques (Hamel, Côté, Smith, & Festa-Bianchet, 2006) – et le lien entre la vigilance de la mère et la survie de son jeune restent inexplorés. L’étude de cette tactique pourrait améliorer notre compréhension du rôle d’un comportement anti-prédateur sur les populations évoluant en sympatrie avec de multiples anti-prédateurs dont la densité peut grandement varier dans un court laps de temps.
Objectifs et hypothèses
Mon mémoire a comme premier objectif d’évaluer l’influence du risque de prédation et du compromis entre la vulnérabilité du jeune et la demande énergétique associées aux soins maternels sur le comportement de vigilance des femelles de la chèvre de montagne. Le second objectif est de vérifier si la vigilance améliore la valeur adaptative des femelles en procurant des bénéfices directs au niveau de leur survie et de celle de leur chevreau. J’ai émis l’hypothèse que le mécanisme expliquant l’ajustement de la vigilance soit le risque de prédation à lui seul (H1) ou encore que le mécanisme soit plutôt le compromis entre le risque de prédation et le besoin de s’alimenter (H2). Chez les femelles reproductrices, j’ai fait l’hypothèse que la vulnérabilité du chevreau est le principal facteur modulant la vigilance (H3). Finalement, ma dernière hypothèse était que les femelles plus vigilantes augmenteraient leur chance de survie et celle de leur chevreau (H4).
Aire d’étude
Mon étude repose sur une population de chèvres de montagne étudiée depuis 1989. Cette population est située à Caw Ridge, un complexe de montagnes isolées par la forêt boréale (Fig. 3) dans le piedmont des Rocheuses du Canada, au centre-ouest de l’Alberta (54°N; 119°W; Festa-Bianchet & Côté, 2008). L’habitat de la chèvre de montagne y occupe environ 28 km2 (Festa-Bianchet & Côté, 2008) et est principalement caractérisé de toundra alpine ainsi que de forêt subalpine en dessous de la limite altitudinale de la forêt (~1900 m). Le climat
alpin-subalpin caractérisant Caw Ridge réduit la période de croissance de végétation à trois mois, de la mi-mai à la mi-août. Les températures sous zéro et les chutes de neige y sont probables à n’importe quel moment de l’année (Festa-Bianchet & Côté, 2008). Les arbres
Figure 3. Vue aérienne de l'aire d'étude de Caw Ridge (Alberta, Canada), à partir de l’ouest.
rabougris et éparses de l’étage subalpin sont principalement composés d’épinettes d’Engelmann (Picea engelmannii) et de sapins des Rocheuses (Abies lasiocarapa; Hamel et al. 2009b).
Suivi à long terme et observations comportementales
L’étude à long terme de Caw Ridge a débuté en 1989 et comprend un suivi où chaque individu est capturé au moins une fois dans sa vie. Le site de trappe est appâté avec des blocs de sel, et les individus sont capturés dans des trappes de type Stephenson ou Clover. Lors de la capture, chaque individu est sexé, âgé, pesé, puis muni d’un collier visuel (Fig. 4a) et d’une combinaison d’étiquettes à oreilles numérotées. Le marquage permet ensuite de reconnaitre les individus pour suivre leur survie et leur reproduction, mais également pour observer leur comportement de la mi-mai jusqu’à la mi-septembre. L’identité des chevreaux est attribuée selon les observations de soins maternels, d’associations mère-jeune, ainsi que par des analyses génétiques (Mainguy, 2008). Nous avons considéré qu’une femelle avait survécu lorsqu’elle était observée le printemps suivant. La survie d’un chevreau a été déterminée soit par son association avec sa mère le printemps suivant ou par analyse génétique s’il n’était pas associé.
De 1994 à 2005 et en 2017-2018, nous avons réalisé des observations cibles de 30 minutes où nous avons quantifié cinq comportements : vigilance, alimentation, interactions sociales, repos, et autres. Ces observations nous ont permis de mesurer la durée de chacune des alertes, le nombre d’alertes et le temps total passé en alerte. Nous avons noté l’habitat, la taille du groupe, les opportunités d’interactions (voir « Behavioural observations » dans la section « Methods » du chapitre principal), de même que le statut reproducteur et l’âge de
Figure 4. Scénario d’observation cible de la femelle #440 dans un groupe de neuf chèvres (A). L’environnement social de #440 à ce moment précis se définirait comme suit : #440 est dans un groupe de neuf chèvres avec un voisin dans son rayon de 4m et elle est en position périphérique (B).
la femelle pour toutes les observations cibles. Entre 2002-2005 et 2017-2018, nous avons noté à chacune des alertes la distance au terrain de fuite le plus près, le nombre de voisins dans un rayon de 4m, la position spatiale dans le groupe (périphérie, mi-périphérie et centrale; Fig. 4), et pour les femelles reproductrices, à quel distance le jeune se trouvait-il et si celui-ci était actif ou non. Ces données ont permis d’analyser plus précisément les variables qui influencent la durée des alertes (Fig. 5). En 2017-2018, nous avons également récolté ces mêmes informations à chaque minute d’une observation pour obtenir une moyenne de l’environnement d’alerte durant les 30 minutes d’une observation, ce qui m’a permis de réaliser une analyse plus précise sur les variables influençant la fréquence d’alerte.
Chapitre 1: Linking proximate drivers and fitness
returns of vigilance: offspring from more vigilant
mothers survive better
1.1 Résumé
La vigilance permet de détecter et éviter les menaces de congénères et prédateurs, mais peut diminuer la valeur adaptative en réduisant le temps pour d’autres activités améliorant la performance individuelle. Nous avons examiné les facteurs influençant la vigilance des femelles adultes et estimé la corrélation entre la vigilance et la survie de la femelle ou de son chevreau à partir d’une population de chèvres de montagne suivie depuis 1989. Les femelles étaient plus vigilantes en forêt ouverte, lorsqu’elles étaient isolées de leurs congénères ou lorsque le risque d’interaction intraspécifique augmentait. Les mères étaient plus vigilantes quand leur jeune était plus loin d’elles. La survie des chevreaux et la vigilance de leur mère étaient positivement corrélées, mais cette association était absente pour la survie annuelle des femelles adultes. Ces résultats suggèrent qu’ajuster la vigilance selon le risque de prédation et la vulnérabilité du chevreau permet aux femelles d’augmenter leur valeur adaptative.
1.2 Abstract
Vigilance allows animals to quickly detect threats from conspecifics and predators to avoid or minimize costly encounters. Time spent vigilant is usually traded-off against other fitness-enhancing activities, such that fitness returns are expected when vigilance is properly adjusted. We used individually marked mountain goats to assess the link between vigilance and fitness in the wild. Our goal was to determine the proximate factors influencing vigilance and to test whether vigilance leads to fitness returns. We first investigated which extrinsic and intrinsic factors influenced alert duration and frequency in adult females using 411 focal observations (30-min each) recorded over 12 years on 115 adult females. We then used a joint modelling framework to estimate the latent correlations at the individual level among vigilance traits (alert duration, alert frequency, and total time spent vigilant) and fitness components (adult female survival and offspring survival) while accounting for the influence of extrinsic and intrinsic factors influencing vigilance. Females adjusted their vigilance in relation to predation risk: they increased vigilance in open forest compared with alpine habitat and were more often vigilant when they were not surrounded by conspecific neighbours. Alerts of reproductive females were more frequent and longer than alerts of females without offspring. Mothers also tended to perform longer alert when their offspring was >10 meters away than at shorter distances, suggesting that variation in offspring’s vulnerability to predation influences mother’s vigilance. There was no association between vigilance and annual survival of adult females, whereas offspring from mothers that were more often alert and spent more time vigilant survived better. Predation risk and offspring vulnerability were the main mechanisms behind variation in vigilance. Adult female survival is high and mostly age-dependent whereas offspring are highly vulnerable to predation, which likely explain why vigilance was strongly correlated with offspring survival but only weakly correlated with female survival. Our results indicate that vigilance is a non-trivial maternal care and that its consequences support the conservative reproductive strategy of long-lived species, because offspring rather than female mortality is correlated with maternal vigilance.
1.3 Introduction
In predator-prey systems, direct and indirect effects of predation can modulate prey life-history traits. The direct effects of predation are lethal, whereas the indirect effects are non-lethal consequences that may reduce individual performance (Creel, Christianson, Liley, & Winnie, 2007). Indirect effects can be induced by physiological changes, such as increased stress hormones levels (Cooke, Bohnert, Reis, & Cappellozza, 2013), or behavioural decisions, such as feeding in poorer but safer habitat (Altendorf, Laundré, López González, & Brown, 2001) or reducing forage intake through changes in activity budgets (Barnier et al., 2016). These decisions can decrease body condition, which can in turn lead to lower reproductive success (Beauplet & Guinet, 2007; Cam, Link, Cooch, Monnat, & Danchin, 2002). Thus, antipredator behavioural strategies may result in large impacts on life-history traits such as survival and reproductive success (Preisser, Bolnick, & Benard, 2005).
Antipredator behavioural tactics are selected in the context of a trade-off opposing energy gain with safety (Brodin & Johansson, 2004). An activity budget that maximizes both daily energetic gains and antipredator tactics should also favor higher fitness returns (Brown, 1999). Because time is limited, allocation to antipredator behaviour is generally incompatible with other fitness-enhancing activities such as foraging (Barnier et al., 2016, but see Fortin et al., 2004). One antipredator tactic that has evolved in this context is vigilance, a measurable behaviour that serves to detect threats early to successfully avoid costly encounters with predators or conspecifics (Beauchamp, 2015). Vigilance is affected by extrinsic factors such as predation risk and social environment. Prey often increase vigilance at high predation risk (e.g. predator odor; Eccard et al., 2017), following the reintroduction of predators (Hunter & Skinner, 1998; Laundré et al., 2001), in habitats with lateral obstruction (Benoist et al., 2013), or when they are far from a refuge habitat (Holmes, 1984; Lazarus & Symonds, 1992). The presence of predators and certain habitat characteristics directly impact how prey perceive predation risk, and thereby modulate their level of vigilance. The social environment also influences vigilance levels because it also serves to detect threats from conspecifics (Beauchamp, 2015). Generally, vigilance decreases with group size (“many eyes” and “dilution” effects hypotheses; Beauchamp, 2017; Lima & Dill, 1990; Pulliam, 1973), but this relation may be mitigated or even reversed in larger groups due to increased intra-specific competition (Treves, 2000). Individuals surrounded by close neighbours may increase vigilance to avoid interactions or to collect social information on the perceived risk of their neighbours (Zhao et al., 2019). Vigilance can also depend on the
spatial position relative to conspecifics (Fernández-Juricic & Beauchamp, 2008; Morrell & Romey, 2008): individuals at the margin of a group are usually more vigilant than surrounded individuals because they are exposed to a greater predation risk (marginal predation; Hamilton, 1971).
Vigilance is also driven by intrinsic factors such as reproductive status, body condition, and age (Beauchamp, 2015). Because offspring are more vulnerable to predation, mothers generally have higher vigilance than females without offspring (Beauchamp, 2015; Burger & Gochfeld, 1994; Caro, 1987). Mothers either increase alert frequency, duration, or both to improve offspring protection (Childress & Lung, 2003). As offspring ages and become less vulnerable, reproductive females can spend less time vigilant (Caro, 1987). Still, the increased nutritional demand of lactation in mammals and maternal care might constrain vigilance levels of mothers (Gélin et al., 2013). Furthermore, individuals in good body condition can allocate more time to vigilance because they have a lower time constraint to meet their energy demand than individuals in poor condition (Bachman, 1993; Wright et al., 2001). Alternatively, as individuals in good condition face lower predation risk, they may also spend less time vigilant (Brivio et al., 2014). Juveniles can either allocate more or less time to vigilance than adults as they experiment high energetic costs due to growth, forage less efficiently, are more vulnerable to predation, and have been less exposed to predation risk (reviewed for many species in Beauchamp, 2015).
Variation of vigilance according to extrinsic and intrinsic factors can impact fitness. Although the factors determining vigilance have been extensively studied, the question of whether or not vigilance contributes to fitness remains largely unexplored, especially in large herbivores (Say-Sallaz et al., 2019). This likely arises because it requires knowledge on factors determining vigilance combined with individual-based monitoring to estimate fitness (Beauchamp, 2015). We fill this gap by studying the relationships among vigilance and fitness traits in individually marked mountain goats (Oreamnos americanus) of a population monitored since 1989. Mountain goats cope with numerous predators (the main cause of mortality; Festa-Bianchet et al., 1994) in a harsh alpine climate that limits resources availability, two conditions that may affect trade-offs and factors determining vigilance. Female mountain goats are capital breeders that evolved a conservative reproductive strategy of generally producing singletons and often skipping reproduction as a result of reproductive costs (Festa-Bianchet, Côté, Hamel, & Pelletier, 2019). Mothers cope with predation risk and the energetic needs of lactation by foraging in safer habitats, spending
more time foraging and ruminating, and increasing bite rate (Hamel & Côté, 2007, 2008, 2009a) compared with barren females. Still, little is known about how mountain goats adjust vigilance according to extrinsic and intrinsic factors, and whether vigilance provides fitness returns. We first hypothesized that predation risk alone explains the variations in alert frequency and duration (H1), predicting that alert frequency and duration would both increase with predation risk. Alternatively, a trade-off between predation risk and daily energetic needs could constrain vigilance levels (H2). In this case, we expected that only one trait of vigilance (e.g. alert duration) would increase while the other one (e.g. alert frequency) would remain stable or decrease when predation risk increases, such that total time spent vigilant would stay stable through alert duration and frequency adjustment (McVean & Haddlesey, 1980). Third, we hypothesized that vigilance of reproductive females depends on the vulnerability of their offspring (H3), predicting that alert frequency and duration would increase when the offspring is active or far from its mother. Finally, we hypothesized that greater vigilance would reduce predation risk and thereby improve fitness (H4), predicting that more vigilant females would have a higher survival probability and increased offspring survival.
1.4 Methods
1.4.1 Study area and captures
We studied mountain goats at Caw ridge, which is a complex of rolling ridges surrounded by boreal forest, in the foothills of the Canadian Rockies, west-central Alberta, Canada (54°N; 119°W). Mountain goats use a 28 km2 area at elevations between 1750 and 2170 m. Climate is characterised by long cold winters and short cool summers, and snowfalls are possible anytime of the year. The main habitats above the tree line are alpine meadows surrounded by rocky slopes and small cliffs, whereas sparse stunted trees mark the transition between the alpine tundra and the dense forest. Mountain goats typically forage in sub-alpine forest or alpine tundra close to escape terrain. These escape terrains are typically composed of sheer cliffs and steep slopes (Gross, Kneeland, Reed, & Reich, 2002). Their main predators include grizzly bear (Ursus arctos; Côté & Beaudoin, 1997), wolf (Canis
lupus; Côté et al., 1997), and cougar (Puma concolor), but other potential predators are
black bear (Ursus americanus), coyote (Canis latrans), wolverine (Gulo gulo), and golden eagle (Aquila chrysaetos; Côté & Festa-Bianchet 2003; Hamel & Côté 2009b).
Since 1989, we have captured mountain goats using salt-baited box traps and self-tripping Clover traps. Adults were immobilized by using xylazine hydrochloride reversed with an injection of Idazoxan (2.4 - 7.5 mg/kg; Haviernick et al., 1998). Most goats were captured as juveniles (1 and 2 years old), and adult females or kids were not captured after 1996 to avoid kid abandonment (Côté, Festa-Bianchet, & Fournier, 1998). Captured individuals were marked with collars (colored symbols) and ear-tag combinations. Since 1993, more than 98% of the population ≥1 year old has been marked. During handling, we aged and sexed each individual, attributing the age of goats first captured as adults (>2 years old) by counting horn annuli, a technique accurate up to 7 years old (Stevens & Houston, 1989). Some females captured early in the study were older than 7 years old (14% of focal observations) and were given the minimum age of 8 at first captured. Because including these individuals did not change the outcome of analyses, we kept all individuals to maximize sample size. More details about the study area, captures, and handling of mountain goats can be found in Festa-Bianchet & Côté (2008). We followed guidelines for the care, welfare and use of animals (permit # 2018033-2, Laval University).
1.4.2 Reproduction and survival
From mid-May to late September, we observed goats almost daily using spotting scopes (15– 45x). A few days prior to parturition, females usually isolate themselves to give birth to a single offspring. We intensively searched the study area each day during the parturition season (mid-May to early June) to determine birthdates, either by direct observations of births or by daily observations of presence/absence of a kid for each female. In some cases (62%), we estimated kid birthdates within five days from the characteristics of the umbilical cord, and by comparing the kid’s behaviour and body size with those of known-age kids (Côté & Festa-Bianchet, 2001a). Each year, a female observed nursing a kid was considered reproductive unless its offspring died during the summer. In such cases, a female was recorded as non-reproductive after the disappearance of her kid and we did not collect behavioural data for two weeks to avoid bias from potential stress due to offspring loss.
We determined annual survival of females and their offspring by their presence in the study area the following summer. Because emigration was seldomly reported for these age and sex-classes (2 females since 1989; Festa-Bianchet & Côté, 2008), we are confident that adult females and offspring not observed the following spring died over winter. Because 60% of female mountain goats are still associated with their yearling in spring (Gendreau, Côté,
& Festa-Bianchet, 2005), we defined the identity of unmarked kids that survived to one year by observing mother-offspring associations (Gendreau et al., 2005) and through DNA analyses (Mainguy, 2008).
1.4.3 Behavioural observations
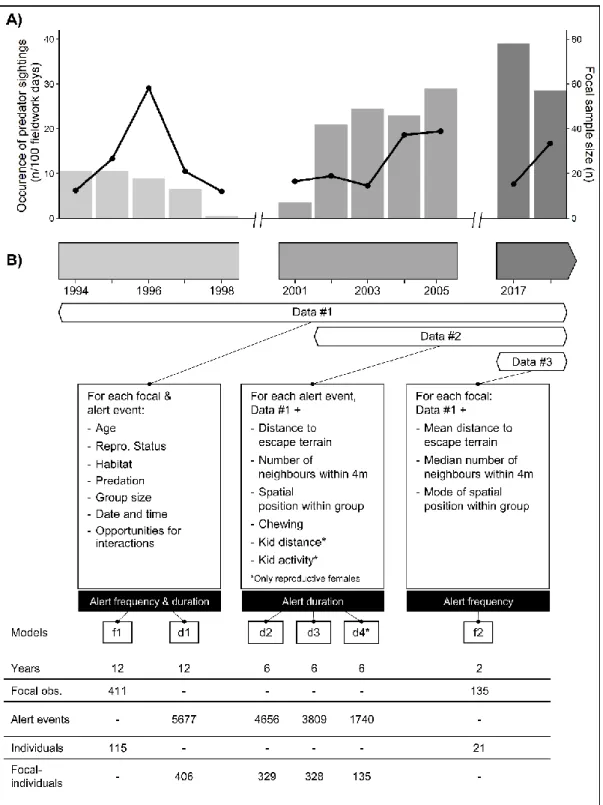
We collected vigilance data using focal-animal sampling (Altmann, 1974). For each observation, we monitored the behaviour of a single female continuously for 30 minutes. We classified the focal female’s behaviour in five categories: vigilant, foraging, lying, social (when interacting with another individual or her offspring), and other behaviours (walking, running, etc.). We defined a goat as vigilant when it was in upright position, ears raised and staring in one direction. From this, we computed three measures of vigilance: alert frequency (total number of alerts during the entire 30 min focal), alert duration (the number of seconds for each alert event during the focal), and total time spent vigilant (total number of seconds in vigilance for the entire 30 min focal). We recorded 411 focal observations on 115 adult females in 12 years spread over three time periods, with 16% of individuals sampled more than once a year (N=306 female*years; Fig. 5).
For all focal observations (Fig. 5), we recorded whether the focal female was in open area (no visual obstruction) or forested area (typically open subalpine forest providing visual obstruction). We also recorded group size and opportunities for interactions, that is the number of times another goat entered within 4 m of the focal individual during the entire focal duration (see Côté, 2000). Starting in 2002 (Fig. 5), we recorded for each alert event the number of neighbours within a 4 m radius, the spatial position within the group (ranging from 0 to 1, with 1 = central, surrounded on four sides; 0.5 = mid-periphery, surrounded on three sides; 0 = periphery, surrounded on one or two sides) and the distance (in m) between the female and the nearest escape terrain for each vigilance event after 2001 (Fig. 5). We also recorded whether or not the individual was chewing for each alert event (only when the mouth was visible), as chewing while being alert is a way to reduce vigilance cost (Fortin et al., 2004; Illius & Fitzgibbon, 1994). Distances were approximated by using known distances between landmarks in the field and “number of goat lengths” (see Hamel & Côté, 2007). For reproductive females, we also recorded whether its offspring was active or bedded and distance to its mother (in m; Fig. 5).
Figure 5. Sampling design across time showing the structure of the datasets in relation with the independent variables available. A) Annual variation in occurrence of predator sightings (number of predator sightings reported per 100 field days; connected solid black dots, left y-axis) and in focal observations sample size (grey bars, right y-axis). B) Structure of the datasets highlighting the independent variables available over the years (listed in the white boxes) in relation with the vigilance responses evaluated (listed in the black boxes; data #1 = 411 focal obs. on 115 individuals; data #2 = 330 focal obs. on 91 individuals; data #3 = 135 focal obs. on 21 individuals).* Variables only available for reproductive females, which were assessed in model d4 that included only vigilance response of reproductive females.