Effets de la composition de la ration sur la synthèse
apparente de vitamines B dans le rumen de la vache laitière
Mémoire
Valérie Beaudet
Maîtrise en sciences animales
Maître ès sciences (M.Sc.)
Québec, Canada
iii
Résumé
Au cours des dernières années, l’intérêt de la communauté scientifique en ce qui a trait à la synthèse de vitamines B dans le rumen des vaches laitières s’est accentué, et une des stratégies employées afin d’augmenter cette synthèse endogène réside dans la formulation des rations. Plusieurs paramètres alimentaires influencent la quantité de vitamines B qui est acheminée à l’intestin. Ce projet de maîtrise a été entrepris pour évaluer les effets de deux de ces paramètres sur la synthèse apparente de vitamines B dans le rumen de la vache laitière : d’une part, la teneur en graines de lin extrudées de la ration, d’autre part, le niveau d’azote et la source d’énergie.
Le premier essai s’est déroulé en deux expériences qui se différenciaient par leur ration de base; la première contenait du foin alors que la deuxième contenait de l’ensilage de maïs. Chacune d’entre elles a été entreprise avec quatre vaches Holstein qui ont ingéré des rations supplémentées à raison de 0, 5, 10 ou 15% en graines de lin extrudées sur une base de MS. L’augmentation de la concentration de graines de lin extrudées dans la ration a induit un plus grand flux duodénal de vitamine B6 et de folates dans les deux expériences. Pour l’expérience conduite avec un régime à base d’ensilage de maïs, il y a eu une réduction de la dégradation de vitamine B6 dans le rumen lorsque la concentration de graines de lin a augmenté dans la ration. La synthèse apparente de folates a augmenté avec un plus grand apport de graines de lin extrudées pour les deux expériences de ce projet. La thiamine, la riboflavine, la niacine et la vitamine B12 n’ont pas été affectées par les traitements.
Le deuxième essai s’est effectué avec quatre vaches Holstein qui ont reçu des traitements alimentaires composés de deux niveaux d’azote (haut: 14% de protéines brutes; bas: 11% de protéines brutes) et de deux sources d’énergie (riche en amidon ou riche en fibres). Le niveau d’azote alimentaire le plus faible a entraîné la diminution du flux duodénal de vitamine B6 ainsi qu’une dégradation accrue de cette vitamine dans le rumen. Il y a également eu une réduction du flux duodénal et de la synthèse ruminale apparente de folates. Le régime alimentaire riche en amidon a augmenté le flux duodénal et la synthèse apparente dans le rumen de riboflavine, de vitamine B6 et des folates alors qu’au contraire, pour la vitamine B12, c’est le régime riche en fibres qui a augmenté la synthèse dans le rumen et le flux duodénal.
Les résultats de ces travaux démontrent que, chez la vache laitière, les caractéristiques de la ration influencent différemment les apports de chacune des vitamines B.
v
Abstract
Over the past years, scientific community interest on B-vitamin synthesis in the rumen of dairy cows has increased, and strategies employed to enhance their endogenous synthesis reside in the formulation of diets. Several dietary factors influence the amount of B vitamins reaching the intestine and this project was undertaken to evaluate the effects of two of these parameters on B-vitamin apparent ruminal synthesis (ARS) in dairy cows. On one hand, the dietary concentration of extruded linseed, and on the other hand, the nitrogen level and the energy source of the diet were studied.
The first trial was conducted in two phases which differed in their basal forage diet. In the first one, cows received hay based diet while in the second one, cows received corn silage based diet. Each experiment was performed with four Holstein cows which ingested extruded linseed supplements at 0, 5, 10 or 15% on DM basis. Increasing extruded linseed concentration in the diet induced a greater duodenal flow of vitamin B6 and folates in both experiments. Vitamin B6 degradation in the rumen decreased with the increased concentration of linseed only in the experiment with corn silage-based diets. Folate ARS increased with a higher supply of extruded linseed in both phases. Thiamin, riboflavin, niacin and vitamin B12 were not affected by treatments.
The second trial was carried out with four Holstein cows receiving dietary treatments comparing two nitrogen levels (high: 14% of crude protein; low: 11% crude protein) and two energy sources (high in starch or high in fiber). Decreasing dietary nitrogen concentration reduced vitamin B6 duodenal flow and increased its ruminal degradation. It also reduced folate duodenal flow and ARS. The rich-starch diets increased duodenal flow and ruminal balance of riboflavin, vitamin B6 and folates while the rich-fiber diets increased vitamin B12 duodenal flow and ARS.
In summary, the results from the current study demonstrate that, in lactating dairy cows, dietary characteristics influence the individual supply of B vitamins differently.
vii
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... xi
Liste des figures ... xiii
Liste des abréviations... xv
Dédicace ... xvii
Remerciements ... xix
Avant-propos ... xxi
INTRODUCTION ... 1
CHAPITRE 1 : REVUE DE LITTÉRATURE ... 3
1.1 Les vitamines du complexe B ... 3
1.1.1 Thiamine (B1) ... 3 1.1.2 Riboflavine (B2) ... 4 1.1.3 Niacine (B3)... 5 1.1.4 Vitamine B6 ... 6 1.1.5 Biotine (B8) ... 7 1.1.6 Acide folique (B9) ... 8 1.1.7 Vitamine B12 ... 9 1.2 Mise en contexte ... 11 1.3 Objectifs de l’étude ... 12
1.4 Synthèse de vitamines B dans le rumen de la vache laitière ... 12
1.4.1 Influence des caractéristiques de la ration sur la synthèse dans le rumen ... 13
1.4.2 Caractéristiques de la ration susceptibles d’affecter la synthèse apparente de vitamines B dans le rumen ... 21
1.4.2.1 Niveau de lipides ... 21
1.4.2.2 Source de glucides et niveau d’azote ... 23
1.5 Conclusion ... 26
1.6 Références ... 27
CHAPITRE 2 ... 33
Effects of Increasing Amounts of Extruded Linseed in the Diet on Apparent Ruminal Synthesis of B vitamins in Dairy Cows ... 33
2.1. Abstract ... 35
2.2. Résumé ... 35
2.3. Introduction ... 36
2.4. Materials and methods ... 37
2.4.1. Animals and treatments ... 38
Experiment 1 ... 38
Experiment 2 ... 38
2.4.2. Sampling and data collection ... 39
2.4.3. Laboratory analyses ... 40 2.4.3.1. Thiamin ... 40 2.4.3.2. Riboflavin ... 41 2.4.3.3. Niacin ... 41 2.4.3.4. Vitamin B6 ... 42 2.4.3.5. Folates ... 43 2.4.3.6. Vitamin B12 ... 43
2.4.4. Calculations and Statistical analyses ... 44
2.5 Results ... 44
2.5.1. Thiamin, Riboflavin, Niacin and Vitamin B12... 47
2.5.2. Vitamin B6 and Folates ... 47
Vitamin B6 ... 47 Folates... 48 2.6 Discussion ... 48 2.7. Conclusion ... 50 2.8. Acknowledgments ... 50 2.9 References ... 51 CHAPITRE 3 ... 55
Effects of Dietary Nitrogen Levels and Energy Sources on Apparent Ruminal Synthesis of B Vitamins in Dairy Cows ... 55
ix
3.4.2. Sample and data collection ... 62
3.4.3. Laboratory analyses ... 62
3.4.4. Calculations and Statistical analyses ... 62
3.5 Results and discussion ... 63
3.5.1. Thiamin (B1) ... 65 3.5.2. Riboflavin (B2) ... 66 3.5.3. Niacin (B3) ... 66 3.5.4. Vitamin B6 ... 66 3.5.6. Folates (B9) ... 67 3.5.7. Vitamin B12 ... 68 3.6. Conclusion ... 68 3.7. Acknowledgments ... 69 3.8 References ... 69 CONCLUSION ... 73
xi
Liste des tableaux
Tableau 1.1 : Teneur en vitamines B d’ingrédients utilisés dans les rations des vaches laitières1 ... 14 Tableau 1.2 : Teneur en vitamines B de matières premières destinées aux animaux d'élevage1 ... 14 Tableau 1.3 : Synthèse apparente de vitamines B dans le rumen de la vache laitière 1 ... 15 Tableau 1.4 : Niveaux moyens1 de vitamines B dans le rumen de bouvillons alimentés avec six rations
différentes2 ... 19 Tableau 1.5 : Digestibilité ruminale de vaches laitières soumises à un régime de graines de lin1 ... 22 Table 2.1 : Ingredient, chemical composition and B-vitamin composition of the diets fed to dairy cows receiving an increasing contribution of extruded linseed in Experiments 1 and 2... 39 Table 2.2: B-vitamin concentrations (mg/kg DM ± SD; n=4) in dietary ingredients of natural grassland hay diets used in Experiment 1... 45 Table 2.3: B-vitamin concentrations (mg/kg DM ± SD; n=4) in dietary ingredients of corn silage diets
Experiment 2. ... 45 Table 2.4: Intake, duodenal flow and apparent ruminal synthesis (ARS) of thiamin, riboflavin, niacin, vitamin B6, folates and vitamin B12 of lactating dairy cows fed natural grassland hay (H) diets supplemented with extruded linseed at 0% (H0), 5% (H5), 10% (H10) or 15% (H15) of DM (Experiment 1) ... 46 Table 2.5: Intake, duodenal flow and apparent ruminal synthesis (ARS) of thiamin, riboflavin, niacin, vitamin B6, folates and vitamin B12 of lactating dairy cows fed corn silage (CS) diets supplemented with extruded linseed at 0% (CS0), 5% (CS5), 10% (CS10) or 15% (CS15) of DM (Experiment 2) ... 47 Table 3.1: Ingredient, chemical composition, nutritive value, and vitamin composition of the diets fed to dairy cows receiving high-starch or high-fiber concentrate with a low or high N level ... 61 Table 3.2 : B-vitamin concentrations (mg/kg DM ± SD; n=4) in the dietary ingredients fed to the dairy cows .. 64 Table 3.3 : Intake, duodenal flow and apparent ruminal synthesis (ARS) of thiamin, riboflavin, niacin, vitamin B6, folates and vitamin B12 of cows receiving high-starch or high-fiber concentrate at a low or high N level ... 65
xiii
Liste des figures
Figure 1.1 : Structure chimique de la thiamine (Royal Society of Chemistry, 2013) ... 4
Figure 1.2 : Structure chimique de la riboflavine (Royal Society of Chemistry, 2013) ... 5
Figure 1.3 : Structure chimique de la niacine (Royal Society of Chemistry, 2013) ... 6
Figure 1.4 : Structure chimique de la vitamine B6 (Royal Society of Chemistry, 2013) ... 7
Figure 1.5 : Structure chimique de la biotine (Royal Society of Chemistry, 2013) ... 8
Figure 1.6 : Structure chimique de l'acide folique (Royal Society of Chemistry, 2013) ... 9
Figure 1.7 : Structure chimique de la vitamine B12 (Royal Society of Chemistry, 2013) ... 10
Figure 1.8 : Influence du rapport fourrage : concentré sur les concentrations de thiamine et de riboflavine dans le rumen de veaux alimentés avec du foin de mil comme unique source de fourrage (*=P<0.01; Adaptée de Conrad et Hibbs, 1954). ... 17
Figure 1.9 : Influence du niveau de la prise alimentaire sur la synthèse ruminale de vitamines B (Adaptée de Zinn et al., 1987) ... 18
Figure 1.10: Contenu en riboflavine (a), acide nicotinique (b), acide pantothénique (c) et biotine (d) par gramme de matériel ruminal sec chez la vache laitière (Adaptée de Lardinois et al., 1944) ... 25
xv
Liste des abréviations
ADF Acid Detergent Fiber ALC Acide Linoléique Conjugué ARS Apparent Ruminal Synthesis ATP Adénosine Triphosphate BW Body Weight
CP Crude Protein CS Corn Silage DIM Days In Milk DM Dry Matter DMI Dry Matter Intake FA Fatty Acid
FAD Flavine Adénine Dinucléotide FMN Flavine Mononucléotide
H Hay
HPLC High Performance Liquid Chromatography MS Matière Sèche
N Nitrogen
NA Nicotinic Acid
NADH Nicotinamide Adénine Dinucléotide Hydrogène
NADPH Nicotinamide Adénine Dinucléotide Phosphate Hydrogène
NAM Nicotinamide
NE Net Energy
NDF Neutral Detergent Fiber OM Organic Matter O/R Ochrosomonas/Radio-essai P5P Pyridoxal-5’-Phosphate PAL Pyridoxal PAM Pyridoxamine PYR Pyridoxine TMP Thiamine Monophosphate TPP Thiamine Pyrophosphate TTP Thiamine Triphosphate UV Ultraviolet
xvii
À Simon;
À Papa & Maman, mes fiers commanditaires
xix
Remerciements
Christiane Girard; mes premiers remerciements te reviennent puisque sans toi, je ne serais certainement pas là où je suis aujourd’hui. Tu m’as engagée pour ce qui ne devait être qu’un stage d’été, mais tu as cru en moi et m’as laissée savoir à ce moment-là que j’avais les capacités pour faire une maîtrise, ce dont je doutais. L’idée a sans doute germé dans mon esprit avec le temps, car à la fin de mon baccalauréat en biologie, j’ai eu envie de poursuivre aux études supérieures. Mes connaissances en agronomie étaient alors quasi inexistantes, mais deux ans plus tard, je repars avec de nombreuses nouvelles connaissances et j’ai le sentiment du devoir accompli. J’ai beaucoup apprécié ton attention, parfois quasi maternelle, ta patience, ta gentillesse et ta disponibilité à tout instant. Merci d’avoir croisé ma route et d’être une personne aussi exceptionnelle!
Rachel Gervais; merci pour ta confiance presque aveugle malgré la distance; tu es énergique, rafraîchissante et je me sens vraiment choyée de t’avoir eue comme directrice de maîtrise. J’admire beaucoup ton approche, ta façon de travailler et ta franchise, le tout combiné à une immense gentillesse qui a grandement facilité le travail que j’avais à faire. Merci énormément!
Chrystiane Plante; merci pour ton aide technique sans pareille, ta rigueur scientifique, ta gentillesse, ta bonne humeur et ton dévouement!
Mes collègues étudiants Douglas De Souza Castagnino (maintenant demi-Brésilien, quart Québécois et quart Français) & Mélissa Duplessis; merci d’avoir contribué à ma compréhension du monde de l’agronomie et d’avoir rendu ces deux années plus joyeuses et mémorables. Un merci spécial à Douglas, avec qui j’ai partagé tellement de belles discussions et de rires lors de nos voyages vers l’Université Laval… là où le temps passé sur la route était plus long que le cours en soit!
Merci à tous les gens du centre de recherche à Agriculture et Agroalimentaire Canada de m’avoir aidée à un moment ou à un autre lorsque j’avais «une ptite question», spécialement à ceux du labo 218 Michelle Guillette, Isabelle Audet & Véronique Roy.
Merci à mes meilleures amies, mes âmes sœurs, Annie-Claude, Chloé, Marilyne & Vanessa.
Simon; merci à toi qui durant toutes ces années universitaires m’as supportée, m’as «bottée» le derrière quand j’en avais besoin, m’as préparé de merveilleux caffè latte au Coureur des Bois qui me remontaient tellement le moral! Toi qui m’as aimée, respectée et attendue sans compter les heures, les jours voire même les années… Merci de partager ma vie et de faire de moi ta petite princesse.
Mes fiers supporteurs de tous les instants, mes parents, Yvan & Angèle; merci d’être toujours là pour moi, merci de m’avoir transmis de bonnes valeurs, dont la persévérance qui me permet d’écrire ces lignes aujourd’hui. La femme que je suis maintenant ne serait pas la même sans vous, vous avez toute mon admiration et ma reconnaissance… et plus encore! Merci pour tout, je vous aime énormément!
xxi
Avant-propos
Ce mémoire contient deux chapitres rédigés sous forme d’article scientifique. Je suis l’auteure principale de ces articles, les coauteurs sont P.Y. Chouinard, M. Doreau, A. Fanchone, R. Gervais, C.L. Girard, B. Graulet, C. Martin et P. Nozière. Ces personnes se sont impliquées dans les travaux. Le premier article porte le titre «Effects of increasing amounts of extruded linseed in the diet on apparent ruminal synthesis of B vitamins in dairy cows» et sera soumis pour publication dans la revue «Animal». Le deuxième article porte le titre «Effects of dietary nitrogen levels and energy souces on apparent ruminal synthesis of B vitamins in dairy cows» et sera soumis pour publication dans la revue «Journal of Dairy Science».
1
INTRODUCTION
Au début du 20e siècle, plusieurs travaux démontrèrent qu’il existe une synthèse apparente de vitamines B dans le rumen des vaches laitières. À partir de ces données, on conclut que les ruminants ayant une bonne alimentation et un rumen fonctionnel pouvaient satisfaire leurs besoins en vitamines B (Bechdel et Honeywell, 1927; Bechdel et al., 1928; Kon et Porter, 1954) et donc que les carences en ces nutriments étaient quasi impossibles. Cela explique pourquoi, pendant plusieurs décennies, les recherches dans ce domaine ont été très limitées. Cependant, plusieurs chercheurs ont récemment montré que les vaches laitières hautes productrices pouvaient bénéficier de suppléments vitaminiques (Jaster et Ward, 1990; Girard et Matte, 1998; Shaver et Bal, 2000; Girard et Matte, 2006; Graulet et al., 2007). Malgré cela, la réponse à ces suppléments alimentaires est très variable puisque ceux-ci ne sont généralement pas protégés de la dégradation dans le rumen et, conséquemment, les quantités de vitamines réellement disponibles pour la vache sont inconnues.
Le rumen étant un environnement affecté par les caractéristiques de la ration (Hunt et al., 1941; 1943; Lardinois et al., 1944; Conrad et Hibbs, 1954; Hayes et al., 1966), il y a des variations dans la synthèse apparente de vitamines B selon le rapport fourrage:concentré, le niveau d’ingestion, la taille et le traitement des particules ou encore avec les additifs alimentaires. Pareillement, il y a des facteurs qui sont susceptibles d’avoir un impact sur la synthèse de vitamines B puisqu’ils sont connus pour affecter la microflore ruminale. Par exemple, les glucides facilement fermentescibles sont plus efficaces que les autres glucides pour promouvoir la croissance microbienne (Bach et al., 2005). Ensuite, le métabolisme de l'azote peut dépendre de la nature de l'écosystème bactérien (Belanche et al., 2012). Par ailleurs, il y a une préoccupation croissante en ce qui concerne les impacts environnementaux de l’élevage. Outre le fait qu’il est un gaz à effet de serre puissant qui contribue au réchauffement climatique de la planète, le méthane émis par les vaches laitières représente aussi une perte d’énergie pour celles-ci (Moss et al., 2000). Une des stratégies étudiées afin de réduire les émissions entériques de méthane des vaches laitières est la supplémentation alimentaire de graines de lin (Martin et al., 2008). En plus de réduire les émissions de méthane, Gómez-Cortés et al. (2009) ont montré que l’ajout de graines de lin extrudées dans la ration permet un gain énergétique résultant en une augmentation de la production laitière. Néanmoins, les effets de ces facteurs alimentaires sur le bilan des vitamines B sont inconnus et l’information n’est pas disponible dans la littérature.
Ce mémoire présente les résultats de deux projets de recherche bien distincts qui ont évalué des facteurs alimentaires différents, mais qui ont pour objectif commun d’évaluer les effets de ces facteurs alimentaires sur la synthèse apparente des vitamines B dans le rumen de la vache laitière. Les données
recueillies pourront se joindre aux résultats d’autres études et seront profitables ultérieurement à l’élaboration d’un modèle de prédiction de la disponibilité des vitamines B pour la vache laitière en fonction des conditions de régie nutritionnelle.
L’hypothèse de ce projet de maîtrise soutient que les caractéristiques de la ration influencent la synthèse apparente de vitamines B dans le rumen, et au final, les quantités disponibles pour la vache. Les objectifs spécifiques sont de déterminer l’apport alimentaire, le flux duodénal et la synthèse apparente de thiamine, riboflavine, niacine, vitamine B6, folates et vitamine B12 dans le rumen de vaches soumises à différentes conditions nutritionnelles, soit des niveaux croissants de graines de lin extrudées ou deux niveaux d’azote et deux sources d’énergie.
3
CHAPITRE 1 : REVUE DE LITTÉRATURE
1.1 Les vitamines du complexe B
Les vitamines sont des molécules organiques essentielles à l’organisme qui sont requises seulement en faibles quantités, car elles répondent à des besoins spécifiques et peuvent être «recyclées». Les scientifiques ont découvert les vitamines B au début du 20e siècle et les ont classées de façon empirique. Elles montrent généralement des ressemblances fonctionnelles et structurelles, autant entre elles qu’entre les différentes formes de chacune des vitamines (Combs, 2012). Plus précisément, les vitamines B sont des composés hydrosolubles dont les besoins pour l’hôte sont définis comme la quantité minimale devant être présente dans l’alimentation afin d’éviter les symptômes de carences.
Il existe huit vitamines B : thiamine, riboflavine, niacine, acide pantothénique, vitamine B6, biotine, acide folique et vitamine B12. Dans l’organisme, les vitamines B agissent à titre de cofacteurs ou de coenzymes, c'est-à-dire qu’elles participent à une réaction enzymatique et retrouvent leur état initial à la fin de cette réaction. Ces dernières possèdent toutes des caractéristiques qui leur sont propres et qui complexifient leur détermination analytique dans les aliments et les échantillons biologiques. En étant des cofacteurs ou des coenzymes, les vitamines B sont généralement liées à des protéines. Une extraction enzymatique est nécessaire afin de cliver les liens qui les retiennent à d’autres molécules. Les enzymes utilisées sont généralement la papaïne (elle fait la digestion de substrats protéiques plus intensivement que les protéases pancréatiques), l’α-amylase (elle hydrolyse les liens α des polysaccharides) et la phosphatase acide (une hydrolase acide utilisée pour déphosphoryler les protéines et les acides nucléiques). Cela permet d’augmenter la concentration de vitamines détectée dans un échantillon.
1.1.1 Thiamine (B
1)
La thiamine est la première d’une longue série de vitamines B à être découverte. C’est d’abord la capacité d’une substance à guérir le béribéri qui a été constatée par un médecin néerlandais, Christiaan Eijkman, puis cette substance contenue dans le riz entier a été isolée par C. Funk en 1910 (Le Grusse et Watier, 1993). Ce dernier la nomma vitamine à cause de son groupement amine. Concrètement, cette molécule est composée d’un cycle pyrimidique, d’un cycle thiazolique azoté et soufré, le tout relié par un pont méthylène (Figure 1.1). La thiamine peut être sous forme libre ou sous forme de thiamine monophosphate (TMP), pyrophosphate (TPP) ou triphosphate (TTP). La TPP, ou cocarboxylase, est la forme la plus abondante dans l’organisme et joue un rôle de cofacteur important dans le métabolisme glucidique. La TTP
joue, pour sa part, un rôle vital dans la transmission nerveuse. La thiamine est naturellement et largement distribuée, en faible concentration, et est synthétisée par les bactéries, les champignons et les plantes.
Figure 1.1 : Structure chimique de la thiamine (Royal Society of Chemistry, 2013)
Des techniques récentes utilisant la chromatographie en phase liquide à haute performance (HPLC) équipée d’un détecteur à fluorescence permettent aujourd’hui un dosage de la vitamine beaucoup plus rapide, précis et reproductible que d’autres méthodes analytiques comme les essais microbiologiques (De Leenheer et Lambert, 2000). Suite à l’hydrolyse enzymatique, les échantillons sont soumis à du ferricyanure de potassium [K3Fe(CN)6] afin de transformer les trois formes de thiamine phosphorylées en thiamine libre qui sera détectée par le fluorimètre. Cela augmente la précision du dosage et diminue la marge d’erreur.
1.1.2 Riboflavine (B
2)
La riboflavine a été différenciée de la thiamine en 1920. Elle doit son nom à sa structure, soit une molécule de ribose associée à une flavine, à laquelle est lié un hétérocycle azoté à trois noyaux (Figure 1.2). Les deux formes métaboliquement actives dérivent de la riboflavine : flavine mononucléotide (FMN) et flavine adénine dinucléotide (FAD) et ont pour noms les coenzymes flaviniques. Les trois formes sont abondamment retrouvées dans l’alimentation, autant d’origines animales que végétales, mais la riboflavine libre est la forme la moins abondante. Le FMN et le FAD sont les coenzymes des déshydrogénases (réactions anaérobiques) et des oxydases (réactions aérobiques; Combs, 2012). Ces coenzymes flaviniques sont fondamentaux pour le métabolisme des glucides, des lipides et apportent une protection cellulaire antioxydante.
5 Figure 1.2 : Structure chimique de la riboflavine (Royal Society of Chemistry, 2013)
Tout comme la thiamine, les concentrations de vitamine B2 sont déterminées par HPLC. L’extraction enzymatique est effectuée avant de faire la quantification, car il a été montré que les enzymes permettent la libération de vitamines dans les échantillons (C. Plante, communication personnelle). Pour la riboflavine par exemple, on observe 32 et 21% plus de riboflavine, dans le solide duodénal et la ration totale mélangée respectivement, que sans l’utilisation d’enzymes. Pareillement, une dérivatisation précolonne est réalisée avec de la phosphatase acide pour transformer les formes de FMN et FAD en riboflavine. C’est cette forme libre qui est quantifiée par fluorescence.
1.1.3 Niacine (B
3)
La niacine a été découverte dans la première moitié du 20e siècle et a au préalable été nommée vitamine PP, pour pellagra preventive factor, car les scientifiques de l’époque ont montré qu’une carence en vitamine B3 causait la pellagre (Le Grusse et Watier, 1993). La niacine correspond à deux composés : l’acide nicotinique (acide pyridine 3-carboxylique) et la nicotinamide (amide de l’acide nicotinique; Figure 1.3). Cette vitamine est disponible dans une grande quantité d’aliments, généralement liée à des protéines, et est davantage sous forme d’acide nicotinique chez les plantes et de nicotinamide dans les tissus animaux. Cependant, en plus de l’apport alimentaire de cette vitamine, il est possible qu’il y ait synthèse endogène à l’aide d’un acide aminé essentiel : le tryptophane. La vitamine B3 est nécessaire à la biosynthèse du NADH et du NADPH, qui ont un rôle clé dans des réactions d’oxydoréduction de l’organisme, notamment le métabolisme des glucides, des lipides, le cycle de Krebs et la synthèse d’hormones sexuelles.
Nicotinamide Acide nicotinique
Figure 1.3 : Structure chimique de la niacine (Royal Society of Chemistry, 2013)
Le maïs possède une caractéristique pouvant avoir un impact important en nutrition bovine. En effet, en plus d’être pauvre en tryptophane, l’acide nicotinique qui s’y trouve est complexé à un ester ce qui le rend indisponible pour l’absorption, à moins d’être soumis à un traitement thermique. La capacité des formes acide nicotinique et nicotinamide à s’interconvertir en conditions acides complexifie le dosage de la niacine qui se fait également à l’aide du HPLC. Suite à l’extraction enzymatique, les échantillons sont disposés dans l’appareil pour leur quantification. À l’exception des autres vitamines B quantifiées par HPLC, la mesure de la niacine nécessite une dérivatisation postcolonne, c’est-à-dire que les échantillons sont exposés à une lumière UV (réaction photochimique) d’une longueur d’onde se situant entre 300 et 400 nm excluant 254 nm. Cela permet ainsi à la vitamine d’être détectée par le détecteur fluorimétrique.
1.1.4 Vitamine B
6La vitamine B6 a été découverte en 1935 par P. Györgyi. On retrouve trois formes vitaminiques qui dérivent de la pyridine : une forme amine (pyridoxamine), une forme aldéhyde (pyridoxal) et une forme alcool (pyridoxine; Figure 1.4). Elles peuvent toutes être phosphorylées, mais la plus importante d’entre elles est le phosphate de pyridoxal, puisqu’il s’agit de la forme métaboliquement active de la vitamine B6 (Combs, 2012). Cette dernière joue le rôle critique de coenzyme dans plusieurs réactions biologiques. Le phosphate de pyridoxal joue un rôle majeur dans le métabolisme des acides aminés, mais également dans celui des glucides, des lipides et dans la synthèse de neurotransmetteurs comme la sérotonine ou la dopamine. La
7
Pyridoxamine Pyridoxal Pyridoxine
Figure 1.4 : Structure chimique de la vitamine B6 (Royal Society of Chemistry, 2013)
La quantification de la vitamine B6 dans les échantillons requiert l’utilisation du HPLC pour sa grande précision. Cette vitamine pouvant apparaître sous trois formes, le dosage doit être assez spécifique pour pouvoir atteindre le seuil de détection et permettre d’obtenir la concentration de toutes les formes présentes dans les échantillons. Suite à l’extraction enzymatique effectuée avant le dosage, une étape de dérivatisation précolonne est réalisée avec de la phosphatase acide pour transformer la forme active de la vitamine B6, le phosphate de pyridoxal, en pyridoxal.
1.1.5 Biotine (B
8)
La vitamine B8 a été découverte autour des années 1920 alors que les chercheurs tentaient de trouver la cause de la maladie du blanc d’œuf. Quelques années plus tard, deux études parallèles aboutirent à la conclusion que dans l’œuf cru, la biotine est fortement liée à une glycoprotéine appelée avidine, ce qui empêche la biotine d’être absorbée, créant ainsi un effet de carence. Le lien covalent qui unit les deux molécules peut être brisé par la chaleur libérant ainsi la vitamine (Le Grusse et Watier, 1993). La biotine est formée par la fusion de deux cycles : un cycle imidazolidone et un cycle tétrahydrothiophène (Figure 1.5). La vitamine B8 est présente dans un large éventail d’aliments et y apparaît sous deux formes : une libre (moins abondante) et une liée aux protéines par un résidu lysine. Cette dernière forme est communément appelée biocytine. Il s’agit d’un lien amide qui unit la biotine à la lysine, ce qui implique que les peptidases intestinales ne peuvent effectuer leur travail. Ce lien doit être clivé par une enzyme spécifique naturellement présente dans les cellules de la bordure en brosse de l’intestin grêle, dans le pancréas et dans certaines bactéries: la biotinidase (Schwab et al., 2006). La biotine agit en tant que coenzyme des carboxylases de l’organisme en transférant un groupement CO2 vers divers substrats. Elle opère dans le métabolisme des glucides, des acides gras et des acides aminés.
Figure 1.5 : Structure chimique de la biotine (Royal Society of Chemistry, 2013)
Due à son fort lien avec la lysine, le dosage de la biotine est une tâche laborieuse. En effet, la biotinidase nécessaire au clivage du lien amide n’est pas disponible commercialement. La quantification de la biotine libre n’est donc pas représentative de la quantité totale de vitamine présente dans l’échantillon. Cette quantification doit être faite en évaluant la biotine et la biocytine. Schwab et al. (2006) ont constaté qu’environ 93% de la biotine totale dans divers échantillons était présente sous forme de biocytine. Cependant, les caractéristiques de cette vitamine rendent le procédé plus complexe et c’est pourquoi des méthodes de dosage fiables ne sont pas encore au point.
1.1.6 Acide folique (B
9)
L’acide folique a été découvert dans les années 1940. Cet acide ptéroylglutamique a pour structure une combinaison d’un noyau ptéridine, d’un acide para-aminobenzoïque et d’un acide glutamique (Figure 1.6). La vitamine B9 comprend par contre une multitude de dérivés qui sont rassemblés sous le terme de folates. En effet, la molécule d’acide folique peut compter une ou plusieurs molécules d’acide glutamique, formant des polyglutamates. Une fois réduits, ces polyglutamates constituent les formes biologiquement actives (ex. dihydrofolates et tétrahydrofolates). Les folates sont bien répandus dans les aliments de sources animales et végétales. Comme toutes les vitamines B, la vitamine B9 est fondamentale dans divers processus métaboliques puisqu’elle participe à la synthèse des bases puriques et pyrimidiques en vue de former les acides nucléiques, à la division cellulaire, à l’élaboration des acides aminés, au cycle des méthylations et à la
9 Figure 1.6 : Structure chimique de l'acide folique (Royal Society of Chemistry, 2013)
Étant donné la multitude de formes que peuvent prendre les folates, leur dosage est plus complexe. La méthode utilisant le HPLC n’est pas encore définie alors que la méthode par radio-essai ne donne pas les résultats escomptés. Cette dernière a été développée pour des échantillons humains donc la forme de folates qui est détectée est principalement le 5-méthyltétrahydrofolate puisque c’est la forme active de l’organisme. Pour l’analyse d’aliments et des contenus ruminaux ou duodénaux, ce n’est pas approprié, car beaucoup de formes de folates passent inaperçues ou sont surévaluées. Le dosage par microbiologie se fait depuis des décennies, et récemment, une technique en plaque microbiologique a été mise au point. Celle-ci utilise
Lactobacillus rhamnosus, un microorganisme dont la croissance dépend de l’apport en folates. L’intensité du
métabolisme ou de la croissance de ce microorganisme exprime la quantité totale de folates contenus dans l’échantillon.
1.1.7 Vitamine B
12L’histoire de la vitamine B12 commence en 1925 et prendra près de 50 ans de recherche avant que les scientifiques en comprennent toutes les facettes et qu’ils arrivent à en faire la synthèse. La structure moléculaire de cette vitamine est volumineuse et son poids moléculaire est élevé. La vitamine B12 est composée d’un noyau tétrapyrrolique qui renferme en son centre un atome de cobalt relié à quatre atomes d’azote. On retrouve audessous un groupement pseudonucléotidique et audessus un groupement R (CN, -OH, -CH3, 5’-désoxyadénosine; Figure 1.7). La synthèse de la vitamine B12 est presque exclusivement effectuée par les bactéries. Ainsi, les aliments qui en contiennent sont les tissus animaux où la vitamine dérive de la microflore ruminale ou intestinale, et les aliments fermentés. De plus, cette synthèse dépend d’un apport adéquat en cobalt, élément essentiel de la molécule.
La vitamine B12 a de multiples fonctions dans l’organisme, mais elle est particulièrement importante pour deux enzymes. L’adénosylcobalamine est coenzyme de la méthylmalonyl-CoA mutase qui catalyse la
transformation du méthylmalonyl-CoA en succinyl-CoA dans le cycle de Krebs, et donc joue un rôle majeur dans le métabolisme énergétique chez les ruminants. En second lieu, la méthylcobalamine est coenzyme de la méthionine synthétase qui catalyse la méthylation de l’homocystéine pour régénérer la méthionine. Cette dernière réaction est intimement liée avec le 5’-méthyltétrahydrofolate (5-CH3-THF; forme de la vitamine B9). En effet, c’est d’abord le 5-CH3-THF qui transfère son groupement méthyle vers la cobalamine pour devenir la méthylcobalamine, puis il y a un don immédiat du groupement méthyle pour convertir l’homocystéine en méthionine (Combs, 2012).
Figure 1.7 : Structure chimique de la vitamine B12 (Royal Society of Chemistry, 2013)
La détermination de la concentration de vitamine B12 dans un échantillon se fait à l’aide d’un radio-essai. Le tampon utilisé pendant ce dosage, lorsqu’ajouté aux échantillons, transforme toutes les formes de cobalamine en cyanocobalamine grâce au cyanure qu’il contient et c’est cette forme qui est quantifiée. Le principe de ce test permet d’obtenir la quantité de «vraie» vitamine B12 et non pas celle comprenant ses analogues. Les analogues sont des molécules similaires morphologiquement, mais qui n’ont pas les mêmes
11
1.2 Mise en contexte
Au début du 20e siècle, les travaux de Bechdel et al. (1928) ont révélé qu’un veau peut croître normalement et produire des descendants normaux en consommant une ration ayant un apport insuffisant en vitamines B, et qu’une vache laitière peut produire du lait ayant une concentration de vitamines B adéquate indépendamment des quantités de vitamines ingérées (Bechdel et Honeywell, 1927). Ces travaux contribuèrent à établir qu’il existe une synthèse microbienne de vitamines B dans le rumen, ce qui fut appuyé plus tard par d’autres études, dont celle de Kon et Porter (1954). La communauté scientifique a alors accepté le dogme selon lequel les ruminants ayant un rumen fonctionnel et une bonne alimentation peuvent satisfaire leurs besoins en vitamines B et qu’une carence est pratiquement impossible. La recherche dans ce domaine a donc été au ralenti pendant de nombreuses années puisqu’elle semblait «inutile».
En s’y penchant de plus près, certains chercheurs ont cependant constaté qu’avec les années, il y avait une augmentation des performances animales et que les vaches étaient de plus en plus productives. Par exemple, en 1976, une vache avait une production moyenne de 5770 kg de lait par an alors qu’en 2010, cette production était de 9111 kg de lait par an (Hutjens, 2012). Cela représente une augmentation de 58%. Les scientifiques se sont aperçus que l’efficacité métabolique peut être affectée par cette production accrue, car il y a nécessairement une augmentation de la demande en enzymes pour produire davantage de lait et de composantes, ce qui sollicite aussi plus de cofacteurs, comme les vitamines B. De nombreuses études ont ensuite montré les effets bénéfiques des supplémentations en vitamines B, laissant supposer que l’apport aux animaux n’était effectivement pas suffisant. Par exemple, une supplémentation alimentaire en acide folique augmente la production laitière de vaches multipares (Girard et Matte, 1998); il semble qu’une supplémentation alimentaire combinée de vitamines B9 et B12 améliore l’efficacité métabolique (Girard et Matte, 2006; Graulet et al., 2007; Preynat et al., 2009); Shaver et Bal (2000) ont montré qu’une plus grande concentration de thiamine dans la ration des vaches laitières permet à celles-ci d’avoir un meilleur rendement en lait; un apport accru en niacine a pour effet de diminuer les cétoses par la capacité de l’acide nicotinique à réduire la mobilisation des acides gras libres en bloquant la lipolyse, et à augmenter la concentration de glucose sanguin (Fronk et Schultz, 1979; Jaster et Ward, 1990). Des veaux supplémentés en acide folique ont eu une croissance légèrement plus rapide, leur permettant d’atteindre le poids du marché entre trois et cinq jours plus tôt, de même qu’une plus grande concentration hépatique de folates comparée à celle des animaux témoins (Lévesque et al., 1993). Ces résultats ont contribué à raviver l’intérêt pour la recherche sur les vitamines B chez les vaches laitières.
1.3 Objectifs de l’étude
La supplémentation peut être envisagée afin d’augmenter la teneur en vitamines B disponibles pour les vaches laitières. Toutefois, malgré qu’elle semble simple et intéressante, cette pratique est confrontée à des obstacles d’ordre technique et économique. Des études sur la supplémentation en vitamines B ont montré que les suppléments alimentaires ne résistent pas au passage dans le rumen et sont détruits en grande partie (Zinn et al., 1987; Majee et al., 2003; Santschi et al., 2005a) alors que des injections intramusculaires impliquent un engagement plus important et plus complexe par le producteur pour soigner ses animaux.
Une connaissance plus approfondie des facteurs intervenant dans la synthèse apparente des vitamines B dans le rumen pourrait contribuer à apporter une alternative et/ou un complément à la supplémentation. Le présent projet tentera de :
déterminer les apports alimentaires, la synthèse apparente ainsi que le flux duodénal des vitamines B sous diverses conditions de régies nutritionnelles;
identifier les facteurs alimentaires ayant le plus d’influence sur les quantités de vitamines B disponibles pour l’animal.
Ultimement, ces résultats pourront être jumelés à d’autres études afin d’ériger une banque de données dans le but de développer un modèle de prédiction des apports en vitamine B pouvant être intégré dans les logiciels de formulation de ration pour maximiser l’efficacité métabolique des vaches laitières.
1.4 Synthèse de vitamines B dans le rumen de la vache laitière
À partir du moment où il a été établi qu’il pouvait y avoir synthèse de vitamines par les bactéries du rumen, les scientifiques se penchèrent sur la question afin de découvrir si des éléments pouvaient influencer cette synthèse. La réponse fut positive. L’apport de vitamines B à une vache laitière dépend de leur teneur dans la ration, de même que de la quantité consommée par l’animal, de la synthèse et de la dégradation des vitamines dans le rumen, sans oublier leur biodisponibilité (la capacité à être absorbée).13 essentiellement associées à la fraction bactérienne du contenu ruminal et que seules d’infimes quantités sont détectables dans le liquide environnant. Cela concorde avec ce qui avait été mentionné par Rerat et al. (1959), soit que les vitamines B sont principalement séquestrées dans les bactéries du rumen. Pareillement, il a été démontré, à l’aide de marqueurs, que ni la thiamine ni la vitamine B12 ne sont absorbées en quantités appréciables dans le rumen (Smith et Marston, 1970; Hoeller et al., 1977; 1979). Girard et al. (2001) ont déterminé qu’il n’y a pas de flux net détectable de folates et de vitamine B12 à travers la paroi ruminale avant perfusion de vitamines, et que suite à des perfusions, l’efficacité des vitamines à atteindre la circulation sanguine est très faible. Toutes ces études laissent donc présager qu’il n’y a pas vraiment d’absorption à travers la paroi ruminale et que la synthèse apparente dans le rumen fournit une approximation assez juste de l’apport en vitamines pour l’animal par la population microbienne. Ainsi, la quantité de vitamines B qui disparait dans le rumen est associée à la dégradation par les microorganismes, soit par destruction simple ou pour le bon fonctionnement de leur métabolisme.
L’efficacité de synthèse des vitamines B dans le rumen peut avoir un impact non seulement sur la production laitière, mais également sur la croissance, la reproduction et la santé des vaches laitières. Ainsi, il est important d’identifier les moyens de moduler la synthèse des vitamines B afin de maximiser les performances de production des animaux.
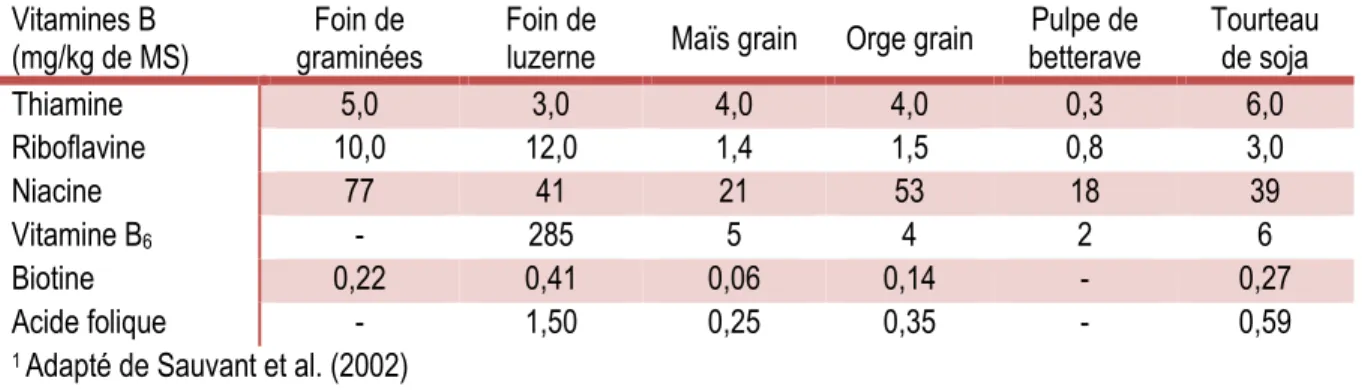
1.4.1 Influence des caractéristiques de la ration sur la synthèse dans le rumen
La composition de la ration est une préoccupation constante dans le domaine des sciences animales. Celle-ci influence la synthèse de vitamines B dans le rumen et de nombreux facteurs nutritionnels ont été évalués au cours des dernières décennies chez plusieurs espèces de ruminants comme la chèvre, le mouton ou le bovin de boucherie. Schwab et al. (2006) ont établi la teneur en vitamines B de différents ingrédients fréquemment utilisés dans l’industrie bovine (Tableau 1.1).Tableau 1.1 : Teneur en vitamines B d’ingrédients utilisés dans les rations des vaches laitières1 Vitamines B
(mg/kg de MS) Ensilage de maïs graminées Foin de Luzerne Foin de moulu Maïs moulue Orge de soja Coque betterave Pulpe de Tourteau de soja
Thiamine 0,57 0,89 1,90 2,70 3,90 1,80 0,62 7,10 Riboflavine 3,5 9,9 17,5 1,2 1,1 2,4 2,0 4,3 Acide nicotinique 22,5 11,8 26,4 7,0 18,9 34,2 59,8 16,0 Nicotinamide 1,5 0,3 7,6 - 18,0 194,7 1,1 26,1 Pyridoxamine 0,24 0,25 0,82 1,60 0,73 0,27 - 2,00 Pyridoxal 0,44 0,15 0,61 1,80 0,28 0,48 5,80 0,84 Pyridoxine 1,90 1,80 4,50 0,21 0,69 1,20 0,61 1,10 Biotine 7,3 7,6 7,4 6,4 6,2 7,2 5,7 6,9
1 Adapté de Schwab et al. (2006)
Il en ressort que les teneurs varient entre les divers aliments, alors que la niacine a la particularité d’être en concentration beaucoup plus élevée dans la pulpe de betterave ainsi que dans la coque et le tourteau de soja. La vitamine B12 n’a pas été évaluée, car comme mentionné précédemment, elle est exclusivement synthétisée par les bactéries. En comparaison, le Tableau 1.2 présente les vitamines B contenues dans quelques-unes des matières premières destinées aux animaux d’élevage.
Tableau 1.2 : Teneur en vitamines B de matières premières destinées aux animaux d'élevage1 Vitamines B
(mg/kg de MS) graminées Foin de Foin de luzerne Maïs grain Orge grain betterave Pulpe de Tourteau de soja
Thiamine 5,0 3,0 4,0 4,0 0,3 6,0 Riboflavine 10,0 12,0 1,4 1,5 0,8 3,0 Niacine 77 41 21 53 18 39 Vitamine B6 - 285 5 4 2 6 Biotine 0,22 0,41 0,06 0,14 - 0,27 Acide folique - 1,50 0,25 0,35 - 0,59
1 Adapté de Sauvant et al. (2002)
Ce tableau présente des concentrations de niacine plus élevées dans tous les aliments comparativement à l’étude de Schwab et al. (2006), bien que l’acide nicotinique et la nicotinamide n’aient pas
15 Santschi et al. (2005a) ont conclu que le flux duodénal de la majorité des vitamines B est plus élevé que le niveau de vitamines ingéré par l’animal, ce qui suggère une synthèse apparente dans le rumen.
Tableau 1.3 : Synthèse apparente de vitamines B dans le rumen de la vache laitière 1
Vitamines B
Synthèse nette dans le rumen Flux total 2,3
(mg/jour) Synthèse dans le rumen, % du flux total mg/kg d’ingestion de MS mg/jour 2 Thiamine 2 51 96 53 Riboflavine 12 274 397 69 Niacine totale 63 1425 2470 58 Vitamine B6 1 21 139 15 Biotine - - 157 - Folates 1 19 30 63 Vitamine B12 4 88 88 100
1 Adapté de Weiss et Ferreira (2006), à partir des données de Santschi et al. (2005a) et Schwab et al. (2006) 2 Basée sur une consommation de matière sèche de 22,7 kg/jour
3 Flux mesuré au duodénum, soit la somme de la consommation de vitamines et de la synthèse nette
Les teneurs en thiamine, riboflavine, niacine et vitamine B6 ont été déterminées par HPLC, la biotine par ÉLISA, alors que les vitamines B9 et B12 par radio-essai. Le Tableau 1.3 montre une synthèse apparente dans le rumen chez la vache laitière de six des sept vitamines étudiées. Le flux duodénal total de la thiamine, la riboflavine, la niacine, les folates et la vitamine B12 provient à plus de 50% de cette synthèse, ce qui est considérable.
Des études ont montré que les suppléments alimentaires de vitamines B ne résistent pas au passage dans le rumen et sont détruits (Zinn et al., 1987; Majee et al., 2003; Santschi et al., 2005a). Étant plutôt inefficace, la supplémentation alimentaire simple entraîne des coûts inutiles aux producteurs. Afin d’atteindre l’intestin et d’être assimilées de façon optimale, les vitamines B doivent être administrées par injection ou être protégées de la dégradation dans le rumen. Leur protection peut se faire avec une matrice d’acides gras saturés, ce qui évite de nuire à l’équilibre ruminal (Sacadura et al., 2008). Sacadura et al. (2008) ont montré qu’un supplément de vitamines B protégées de la dégradation dans le rumen permettait d’augmenter la production de lait ainsi que le rendement en protéines et en gras du lait chez des vaches en début de lactation. Utilisant la même procédure de matrice de graisse que Sacadura et al. (2008), Juchem et al. (2012) ont donné une supplémentation d’un mélange de vitamines B protégées de la dégradation dans le rumen à des vaches multipares hautes productrices en début de lactation. Cette équipe de recherche a conclu que cette récente technique de supplémentation affecte positivement les performances reproductives tout en réduisant le taux de réforme des vaches hautes productrices en début de lactation. Néanmoins, il est
important de miser sur le meilleur apport en vitamines B possible dans la ration, mais surtout sur des facteurs qui favorisent leur synthèse dans le rumen.
La majorité des études ne mesurent que les concentrations des vitamines B dans différentes fractions du contenu ruminal. Par contre, les concentrations de ces vitamines dans le contenu ruminal ne permettent pas de quantifier la synthèse microbienne totale ou les quantités de vitamines atteignant les sites intestinaux où ces vitamines sont absorbées, puisque ces valeurs ne tiennent pas compte du taux de remplissage du rumen ou du taux de passage des digestas. La concentration ruminale en vitamines B est énormément influencée par les taux de remplissage et de passage, qui eux sont grandement influencés par de nombreux facteurs présentés ci-dessous. C’est donc dire que la quantité de vitamines B disponible pour la vache laitière est aussi variable selon l’ingestion, la dégradation ou la synthèse de ces vitamines.
Premièrement, l’effet du rapport fourrage:concentré dans la ration a été évalué chez le bouvillon par Hunt et al. (1941). Les résultats présentés par cette équipe relatent une plus grande concentration de riboflavine dans le rumen lorsque les animaux sont nourris avec un mélange de maïs, de foin de luzerne et d’un supplément protéique qu’avec une ration constituée exclusivement de foin. Hollis et al. (1954) ont conclu la même chose pour l’acide nicotinique, la riboflavine et l’acide pantothénique, soit que la synthèse est relativement faible lorsque le mouton est nourri avec du foin uniquement. Chez le bouvillon Angus, une diète comprenant exclusivement des concentrés en comparaison à une diète composée d’un mélange de maïs et de foin induit une plus forte concentration de thiamine, de niacine, d’acide pantothénique, de folates et de vitamine B12 dans le liquide ruminal (Hayes et al., 1966).
La Figure 1.8 montre les concentrations de thiamine et de riboflavine chez des veaux alimentés avec du foin de mil comme unique source de fourrage jumelé à un mélange de grains. L’analyse du liquide ruminal révéla que le rapport entre le fourrage et les concentrés influence les concentrations de ces vitamines B retrouvées dans le rumen. La thiamine a une concentration optimale à un rapport fourrage:concentré de 4:1, alors que la concentration de riboflavine plafonne à un rapport de 2:1 (Conrad et Hibbs, 1954).
17 Figure 1.8 : Influence du rapport fourrage : concentré sur les concentrations de thiamine et de riboflavine dans le rumen de veaux alimentés avec du foin de mil comme unique source de fourrage (*=P<0.01; Adaptée de Conrad et Hibbs, 1954).
Ensuite, la famille de fourrage qui compose ces rations affecte la digestibilité. Récemment, une étude de Kammes et Allen (2012a) a démontré par exemple que la luzerne augmente le pH ruminal, le taux de digestion et de passage des fibres NDF potentiellement digestibles, mais diminue le temps de rumination par unité de fibres NDF des fourrages consommée par rapport au dactyle. Voelker Linton et Allen (2008) avaient auparavant établi que l’ingestion de luzerne induit un temps de rétention ruminale plus court pour les fibres NDF dans le rumen, ce qui réduit les effets physiques de remplissage comparativement aux animaux nourris avec du dactyle. Cela a nécessairement un effet sur la population microbienne du rumen et donc l’apport final de vitamines B pour la vache risque d’être différent. Seck et al. (2010) ont évalué la synthèse apparente dans le rumen de la niacine et de la vitamine B6 chez des vaches laitières ayant consommé de la luzerne ou du dactyle. Il s’est avéré que la synthèse est plus importante lorsque du dactyle est ingéré comparativement à de la luzerne. Une seconde étude réalisée avec les mêmes animaux a permis d’établir que la synthèse apparente de riboflavine dans le rumen est également supérieure lorsque le dactyle remplace la luzerne dans la ration des vaches laitières (Castagnino et al., 2013).
Troisièmement, le niveau d’ingestion d’une ration est un autre facteur influençant la synthèse de vitamines B dans le rumen. En 1972, une équipe de recherche s’est attardée sur le sujet en évaluant le niveau de production de vitamine B12 chez le mouton en fonction du niveau d’ingestion alimentaire (Sutton et Elliot,
0 10 20 30 40 50 60 Foin 12:1 6:1 4:1 2:1 1:1 0,6:1 Grain C onc entrati on rumi nal e (µg / g de M S) Thiamine Riboflavine * * *
1972). Les animaux de cette étude ingéraient un niveau faible, moyen ou élevé de matière sèche digestible correspondant respectivement à une moyenne de 527, 698 et 884 g/jour. La production de vitamines B12 a été mesurée par microbiologie (Ochrosomonas) et par radio-essai. Les résultats obtenus avec Ochrosomonas indiquèrent que la production de vitamine B12 augmente à mesure que l’ingestion de matière sèche digestible augmente. Cependant, aucune variation n’a été observée lorsque cette production était évaluée avec le radio-essai. Ainsi, le rapport duodénal O/R (interprétant le rapport entre la vitamine B12 biologiquement active et la vitamine B12 totale) croît avec le niveau d’ingestion, ce qui indique une réduction relative de la synthèse des analogues de la vitamine B12.
Figure 1.9 : Influence du niveau de la prise alimentaire sur la synthèse ruminale de vitamines B (Adaptée de Zinn et al., 1987)
Une quinzaine d’années plus tard, Zinn et al. (1987) ont évalué dans quelle mesure la prise alimentaire influence la synthèse de thiamine, riboflavine, niacine, vitamine B6 et vitamine B12 dans le rumen
0 10 20 30 40 50 60 70 1,2 1,6 1,8 2,2 Sy nthès e vit ami nes (mg / j our)
Consommation, % de poids vif
Thiamine Riboflavine Niacine (1/10) Vitamine B6 Vitamine B12
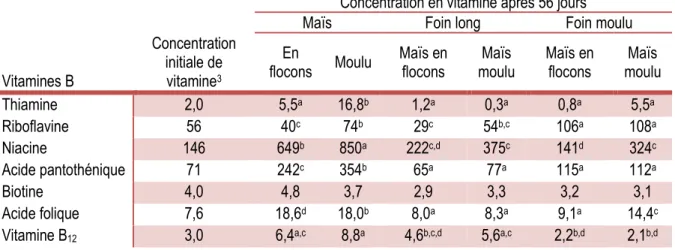
19 Quatrièmement, le traitement des aliments et la taille des particules présentes dans les rations des animaux sont des éléments qui peuvent affecter notamment le temps de mastication et de rumination, ce qui a un effet ultime sur la digestion ruminale. Dans la première moitié du 20e siècle, Hunt et al. (1943) ont montré que le maïs moulu induit une plus grande concentration ruminale de riboflavine qu’une même quantité de maïs entier. Quelques années plus tard, ce résultat a été corroboré par une autre équipe de recherche qui utilisa 48 bouvillons Angus d’environ 340 kg pour mesurer la synthèse de thiamine, riboflavine, niacine, acide pantothénique, vitamine B6, biotine, acide folique et vitamine B12 dans le rumen (Tableau 1.4; Hayes et al., 1966). Six traitements ont été testés, soit 1) maïs en flocons, 2) maïs moulu, 3) maïs en flocons et foin long, 4) maïs moulu et foin long, 5) maïs en flocons et foin moulu et 6) maïs moulu et foin moulu.
Tableau 1.4 : Niveaux moyens1 de vitamines B dans le rumen de bouvillons alimentés avec six rations différentes2
Concentration en vitamine après 56 jours
Maïs Foin long Foin moulu
Vitamines B
Concentration initiale de vitamine3
En
flocons Moulu Maïs en flocons moulu Maïs Maïs en flocons moulu Maïs
Thiamine 2,0 5,5a 16,8b 1,2a 0,3a 0,8a 5,5a Riboflavine 56 40c 74b 29c 54b,c 106a 108a Niacine 146 649b 850a 222c,d 375c 141d 324c Acide pantothénique 71 242c 354b 65a 77a 115a 112a Biotine 4,0 4,8 3,7 2,9 3,3 3,2 3,1 Acide folique 7,6 18,6d 18,0b 8,0a 8,3a 9,1a 14,4c
Vitamine B12 3,0 6,4a,c 8,8a 4,6b,c,d 5,6a,c 2,2b,d 2,1b,d
1 Microgramme par 100 mL de liquide ruminal 2 Adapté de Hayes et al. (1966)
a,b,c,d En excluant les concentrations initiales, les moyennes dans la même ligne avec un exposant différent sont significativement différentes (P < 0,05)
3 Teneurs en vitamines dans le liquide ruminal durant la phase préliminaire (15 jours)
En résumé, le maïs moulu permet une meilleure production de vitamines que le maïs en flocons, à l’exception de la biotine. Par ailleurs, la concentration de riboflavine est plus grande avec une ration contenant du foin moulu plutôt que du foin long. Beaucoup plus récemment, un article évaluant la taille des particules de fourrage sur l’ingestion de matière sèche, la production de lait, la mastication et la fermentation ruminale a été publié par l’équipe de Kammes et Allen (2012b). Selon cette publication, la longueur des particules d’herbe a tendance à augmenter le temps de mastication et d’ingestion, ce qui tend à diminuer l’ingestion de matière sèche (qui peut être limitée par le remplissage du rumen). Cette modification de l’équilibre ruminal affecte inévitablement le métabolisme de fermentation. En effet, le pH du rumen des vaches laitières nourries avec du foin moulu est plus faible que celui des vaches nourries avec du foin haché (Beauchemin et al., 2003).
Le même constat a été fait par l’équipe de recherche de Cao et al. (2010) avec cette fois une ration composée d’ensilage de maïs. Il y a une augmentation linéaire du pH de 6,20 à 6,39 avec l’augmentation de la taille des particules d’ensilage. Ces deux études montrent également que le pH plus élevé observé lorsque les particules sont plus longues est probablement en lien avec un temps de mastication et de rumination plus grand. La concentration en acides gras volatils semble elle aussi affectée par la taille des particules. Il est possible de remarquer que dans une étude, cette concentration est inversement proportionnelle à la taille des particules (Kononoff et al., 2003) alors qu’à l’inverse, dans l’étude de Cao et al. (2010), les teneurs en acides gras volatils étaient positivement corrélées à la longueur des particules. Par ailleurs, la taille des particules a un effet quadratique sur le rapport acétate:propionate dans le rumen (Kononoff et al., 2003; Cao et al., 2010). Les exemples ci-dessus montrent que la fermentation ruminale est affectée par la taille des particules alimentaires laissant ainsi croire que la synthèse de vitamines B puisse l’être également.
Cinquièmement, l’ajout d’additifs dans les rations des ruminants peut modifier les conditions du rumen, comme le pH ou l’osmolarité, et par le fait même l’équilibre microbien. Le cobalt est un minéral qui est fréquemment ajouté à la ration des ruminants. Il s’agit d’un élément chimique qui fait partie intégrante de la molécule de la vitamine B12, et donc son apport est essentiel à la synthèse de cette vitamine par les bactéries du rumen. En effet, la présence de vitamine B12 est nettement plus importante chez des moutons recevant du cobalt per os que ceux ayant une diète carencée en cobalt (Dawbarn et al., 1957). Selon Stangl et al. (1999), une carence en cobalt (200 µg/kg vs. 83 µg/kg) induit chez des bovins de boucherie une diminution de l’ingestion volontaire de matière sèche de 18%, un gain de poids 31% plus faible ainsi qu’une réduction de la teneur en vitamine B12 dans le sérum et le foie, ce dernier étant davantage affecté. Cependant, selon quelques études, lorsque la concentration de cobalt administrée excède la quantité requise par les microorganismes afin de produire la molécule de vitamine B12 dite «active», la proportion de cet élément chimique qui est aussi utilisé pour former des analogues de la cobalamine changent. Les bactéries utilisent alors le cobalt pour produire plus de ces analogues non biologiquement actifs qu’en concentration normale (Ford et al., 1953; Dawbarn et al., 1957; Kawashima et al., 1997; Girard et al., 2009).
21 leur part examiné les effets du monensin en plus de la chlortétracycline. Le monensin n’a pas donné de résultats significatifs sur les quantités de vitamines B aux différents sites de synthèse ou d’absorption, mais a eu un effet négatif sur la digestibilité de la matière organique dans le rumen. La chlortétracycline a eu peu de conséquences sur les vitamines, néanmoins, à plus forte teneur, l’absorption apparente tout comme les quantités dans le contenu duodénal, iléal et fécal de la thiamine et de la niacine ont diminué.
1.4.2 Caractéristiques de la ration susceptibles d’affecter la synthèse apparente de
vitamines B dans le rumen
1.4.2.1 Niveau de lipides
Chez les ruminants, les lipides sont importants, car ils occupent plusieurs fonctions. Par exemple, ils sont une bonne source énergétique et ils contribuent à la structure et à la fluidité des membranes cellulaires. Les lipides sont divisés en plusieurs classes dépendamment de leur structure moléculaire. Les acides gras sont un constituant majeur de plusieurs classes de lipides. Les acides gras possèdent une chaîne hydrocarbonée qui peut être sans liaisons doubles, les acides gras saturés, ou avec une ou plusieurs liaisons doubles, les acides gras mono- ou polyinsaturés. Les acides gras insaturés sont reconnus pour être néfastes pour certaines populations microbiennes présentes dans le rumen (Harfoot et Hazlewood, 1997). Afin de contrer leurs effets, les microorganismes du rumen peuvent effectuer de la biohydrogénation. Cela consiste à saturer les chaînes d’acides gras en ajoutant des molécules d’hydrogène. Ce procédé biochimique se déroule en plusieurs étapes où chacune d’entre elles génère des intermédiaires. Néanmoins, les acides gras insaturés peuvent être bénéfiques pour la santé humaine de plusieurs façons. Ils contribuent par exemple à une meilleure santé cardiovasculaire, immunologique et inflammatoire (Williams, 2000). De plus, certains acides linoléiques conjugués (ALC), isomères de la biohydrogénation de certains acides gras polyinsaturés, peuvent avoir des effets biologiques positifs
Chez les ruminants, une ration normale contient une teneur plutôt faible en lipides, de l’ordre de 2 à 5% sur une base de matière sèche (Doreau et Ferlay, 1994). Comme mentionné précédemment, l’addition d’une source lipidique dans la ration d’une vache laitière peut avoir plusieurs avantages autant sur la santé animale que la santé humaine. Un des suppléments qui peuvent être utilisés par l’industrie alimentaire bovine est la graine de lin. Il est possible de la retrouver sous forme de graine crue, micronisée ou extrudée. L’huile peut aussi être extraite et ajoutée directement à la ration. En théorie, le procédé d’extrusion soumet la graine de lin à un traitement thermique afin de briser les cellules végétales, ce qui rend possiblement la graine plus accessible pour la dégradation ruminale et ses lipides plus rapidement disponibles pour la lipolyse et la
biohydrogénation (Doreau et al., 2009). Cette équipe de recherche a ainsi montré que la supplémentation en graines de lin extrudées permet d’augmenter le flux duodénal d’intermédiaires insaturés par rapport à une supplémentation en graines de lin roulées. Gonthier et al. (2004) ont montré chez la vache laitière qu’une supplémentation en graines de lin extrudées, à raison de 12,6 % sur une base de matière sèche, n’induit pas d’effets sur la digestibilité ruminale de la matière sèche et de la matière organique entre les traitements témoins et ceux supplémentés en graines de lin. Par contre, la graine de lin extrudée entraîne une meilleure digestibilité ruminale de la matière sèche et de la matière organique que la graine de lin micronisée (Tableau 1.5). Pour sa part, l’huile de lin peut être néfaste sur la digestibilité de la matière organique dans le rumen du mouton (Ikwuegbu et Sutton, 1982). Cependant, Ueda et al. (2003) n’ont pas observé ces effets négatifs pour la digestibilité chez des vaches laitières en lactation dont la ration était supplémentée avec 3% d’huile de lin.
Tableau 1.5 : Digestibilité ruminale de vaches laitières soumises à un régime de graines de lin1
Digestibilité ruminale (% de l’apport)
Traitements alimentaires
SEM Témoin Graine de lin crue Graine de lin micronisée Graine de lin extrudée
Matière sèche 66,9 64,2 57,9 68,7 1,89
Matière organique 57,7 57,8 52,0 61,3 1,70
1 Adapté de Gonthier et al. (2004)
En outre, la graine de lin extrudée semble être la forme à privilégier lors d’une supplémentation alimentaire. C’est d’ailleurs sous cette forme que l’équipe de Gómez-Cortés et al. (2009) a exposé que cette source lipidique permet une augmentation de la production laitière. Cette augmentation semble attribuée à une meilleure offre énergétique, ce qui pourrait aussi expliquer les effets bénéfiques sur les performances reproductives puisque les vaches sont en meilleures conditions pour la prochaine gestation. Cependant, une contribution en graines de lin extrudées supérieure à 15% de l’apport en matière sèche induit une diminution de la teneur en matière grasse du lait (Ferlay et al., 2013). L’approvisionnement accru en graines de lin extrudées dans la ration résulte en une augmentation de la teneur en matières grasses alimentaires. Ces modifications peuvent avoir un effet inhibiteur sur les protozoaires, ce qui a pour conséquence de réduire la production de méthane. Martin et al. (2008) ont ajouté 5,7 % de lipides provenant de la graine de lin dans la ration de vaches laitières ce qui a effectivement entraîné une diminution des émissions de méthane.
23 base de matière sèche, sont ajoutées à la ration (Gonthier et al., 2004). En 2010, l’équipe de Côrtes et al. (2010) a montré que l’ajout de graines de lin a modifié le profil en acides gras dans le liquide ruminal. Pour ce qui est de la microflore, un examen superficiel dans le rumen des moutons a révélé des changements marqués dans la population microbienne (Czerkawski, 1973). Dans cette expérience, lorsque la teneur en acides gras libres dérivés d'huile de lin était plus élevée dans la ration, il y avait une chute drastique des protozoaires. Puis, lorsque la ration témoin a été servie de nouveau aux moutons, le nombre de bactéries a diminué alors que celui des protozoaires a remonté, sans par contre retrouver sa concentration initiale. Il semble que le nombre inférieur de protozoaires soit associé à l’augmentation du nombre de bactéries présentes dans le rumen et à des valeurs plus élevées de synthèse de protéines microbiennes. Cela pourrait donc aussi entraîner une variation de la synthèse apparente de vitamines B.
1.4.2.2 Source de glucides et niveau d’azote
Afin d’obtenir de l’énergie pour combler leurs besoins d’entretien, de croissance, de reproduction et de production laitière, les vaches transforment l’énergie contenue dans leur alimentation en un combustible organique appelé adénosine triphosphate (ATP). Cette énergie alimentaire se retrouve sous forme de glucides majoritairement (entre 60 et 70%), mais aussi sous forme de protéines et de matières grasses (NRC, 2001). Afin de dégrader les glucides dans le rumen, la population microbienne procède à l’hydrolyse extracellulaire des polymères glucidiques puis à la fermentation intracellulaire des glucides simples. Cette fermentation produit des acides gras volatils, principalement les acides acétique, propionique, butyrique et valérique, à partir du pyruvate formé lors de la glycolyse des glucides. Le rendement énergétique du métabolisme microbien dépend de la voie métabolique utilisée. Ces acides gras volatils servent ensuite à fournir de l’énergie pour entre autre le bon fonctionnement des tissus de l’animal et la production de lait.
Les glucides de la ration fournissent de l’énergie aux microorganismes du rumen ainsi qu’aux tissus de l’animal. Les glucides non fibreux se composent de sucres simples, d’amidon, d’acides organiques et de glucides de réserve. Les glucides non fibreux constituent la partie la plus digestible par la vache, quoiqu’il reste peu de cette fraction glucidique à digérer après leur passage dans le rumen. Les glucides fibreux sont, pour leur part, des éléments essentiels de la ration puisqu’ils contribuent au bon fonctionnement du rumen. Les fibres sont contenues dans les parois cellulaires des végétaux. Elles sont composées de glucides structuraux, sont beaucoup moins digestibles que les glucides non fibreux et comprennent de la cellulose, de l’hémicellulose et de la lignine (NRC, 2001). Subséquemment, les sources de glucides utilisées dans la ration des vaches laitières influencent le taux de fermentation de ceux-ci. Par exemple, l’amidon, qui est un glucide facilement fermentescible, est facilement attaqué par des amylases sécrétées par la microflore amylolytique