THESE
en vue de l’obtention du
DOCTORAT DE L’UNIVERSITE DE TOULOUSE délivré par l’Université Toulouse III − Paul Sabatier
Discipline : Ecologie Microbienne présentée et soutenue
par Amaia IRIBAR le 26 octobre 2007
Titre :
Composition des communautés bactériennes dénitrifiantes au sein
d’un aquifère alluvial et facteurs contrôlant leur structuration:
relation entre structure des communautés et dénitrification
Directeurs de thèse :
Jose Miguel SÁNCHEZ-PÉREZ et Frédéric GARABETIAN
JURY
I. ANTIGUEDAD Professeur Université du Pays Basque – EHU, Bilbao Examinateur
F. GARABETIANProfesseur Université de Bordeaux I Co-directeur de thèse J. GARNIERDirecteur de Recherche CNRS, UMR 7619 Sisyphe Rapporteur
Je remercie Monsieur Jean Luc Rols, Directeur du Laboratoire d’Ecologie des Hydrosystèmes UMR 5177, pour m’avoir accueillie au sein de son laboratoire pour les 3 premières années de cette thèse. Je remercie également Monsieur Eric Chauvet, Directeur du Laboratoire d’Ecologie Fonctionelle (ECOLAB) UMR5245, pour m’avoir permis de continuer et terminer cette thèse au sein de son laboratoire depuis janvier 2007.
A mon co-directeur de thèse Monsieur Jose Miguel Sánchez Pérez, j’adresse mes remerciements pour m’avoir donné l’occasion de découvrir l’hydro-géochimie et pour son accueil au sein de son équipe, à Toulouse. Je remercie mon autre co-directeur de thèse, Monsieur Frédéric Garabetian, pour sa contribution rigoureuse à ma formation scientifique en microbiologie, ses qualités humaines et son caractère aux accents latins.
Je remercie également Madame Josette Garnier du laboratoire de Structure et Fonctionnement des Systèmes Hydriques Continentaux (SISYPHE) UMR 7619 et Monsieur Philippe Mérot du laboratoire de l’INRA UMR Sol Agronomie Spatialisation de Rennes pour avoir accepté de juger ce travail comme rapporteurs. Je remercie Madame Michelle Trémolières Université de Strasbourg d’avoir présidé le jury et Monsieur Iñaki Antiguedad Université du Pays Basque d’avoir accepté de juger un document qui n’est pas rédigé dans sa langue maternelle.
Je remercie Madame Sara Hallin de m’avoir accueillie au sein de son groupe du département de Microbiologie de l’Université Suédoise d’Uppsala (SLU) pour un séjour de 2 mois dans le cadre d’un échange ATUPS. Je la remercie pour l’intérêt qu’elle a porté à mon travail jusqu'à avoir accepte de faire partie du jury de cette thèse. Au delà je remercie l’ensemble du département de Microbiologie de la SLU pour son accueil et en particulier Madame Karin Enwall pour son aide au laboratoire.
Je suis redevable envers l’ancien plateau technique composé par Madame Claude Mur et Monsieur Daniel Dalger pour les dosages chimiques des eaux, pour leur aide toujours sympathique et efficace et de maints encouragements. Je remercie Mademoiselle Isabelle Vitte et Monsieur Frédéric Julien pour leur soutien sur le terrain et leur participation aux analyses en chromatographie ; Je remercie Monsieur Nicolas Poulet, pour sa disponibilité inconditionnelle et son apport a ce travail en matière d’analyses statistiques qui doivent beaucoup à ses compétences. Enfin Monsieur Yvan Nicaise m’a apporté son soutien inconditionel dans la partie «Biologie Moléculaire ». Sa générosité incommensurable et sa
Un regard spécial à ma familia francesa pour son aide, sa générosité et l’amour donné. Le file crée, qu’il ne se rompe jamais. « Bai pozik eta arro nagoela zuek ezagutu izanaz » (qui dans ma seconde langue se traduirait par « ¡Que contenta y orgullosa estoy de haberos conocido !).
Euskal Herriko nere Iribar sendia, Ama Aita et Asier, nere betiko lagunak Oihana, Nora, Larraitz, Laura eta Amaia urrutitik gertura ibili direnak ere eskertzen ditut. Argiñe, Haritz, Uxu, Aiora…mil esker zuen adiskidetasuna Toulousera gerturatzeagatik. Denbora izango det laxter denoi bixita egiteko !
Azkenerako utzi dut gehien eskertzeko dudan pertsona, Laurent hitz nahikorik ez nuke aurkituko egindako guztia eskertzeko, bikote, zuzentzaile eta lagun bezela. Nere eguneroko maitasuna dezu trukean. Hemendik aurrera gurea da etorkizuna !
Euskal Herriko nere Iribar sendia, Ama Aita et Asier, nere betiko lagunak Oihana, Nora, Larraitz, Laura eta Amaia urrutitik gertura ibili direnak ere eskertzen ditut. Argiñe, Haritz, Uxu, Aiora…mil esker zuen adiskidetasuna Toulousera gerturatzeagatik. Denbora izango det laxter denoi bixita egiteko !
Azkenerako utzi dut gehien eskertzeko dudan pertsona, Laurent hitz nahikorik ez nuke aurkituko egindako guztia eskertzeko, bikote, zuzentzaile eta lagun bezela. Nere eguneroko maitasuna dezu trukean. Hemendik aurrera gurea da etorkizuna !
Sommaire
Sommaire ... 1
Liste des abréviations... 6
Introduction ... 8
Chapitre I. Relation entre la composition des communautés bactériennes et le fonctionnement des écosystèmes : état de l’art... 14
1. Les communautés bactériennes... 15
1.1. Définition ... 15
1.2. Diversité des communautés bactériennes... 16
1.2.1. Définition de la biodiversité... 16
1.2.2. La diversité bactérienne ... 16
1.2.3. La diversité fonctionnelle bactérienne ... 19
1.2.4. La mesure de la diversité... 20
1.3. Facteurs qui affectent la composition d’une communauté bactérienne dans un aquifère... 21
1.3.1 Facteurs abiotiques... 21
1.3.2. Facteurs biotiques... 23
2. Rôle de la diversité sur le fonctionnement d’un écosystème ... 24
2.1. Des modèles théoriques aux études empiriques... 25
2.2. Comment mesurer l’intensité de la relation diversité/fonction ? Quels processus et communautés bactériennes ont été étudiés ? ... 27
3. Le modèle d’étude : les communautés dénitrifiantes des agrégats naturels... 28
3.3.2. Les rôle des communautés bactériennes dans les aquifères alluviaux . 37
3.3.3. Justification du modèle... 39
Chapitre II. Méthodologies... 42
1. Les Biofilms hétérotrophes (BH) de l’aquifère alluvial... 43
1.1. Site d’étude... 43
1.1.1. La Garonne... 43
1.1.2. Le site de Monbéqui... 44
1.2. Échantillonnage... 48
1.2.1. Variabilité spatiale et temporelle de la dénitrification ... 48
1.2.2. Composition des communautés bactériennes libres et attachées ... 50
1.2.3. Expérience de colonisation... 51
1.2.3.1. Choix des piézomètres ... 51
1.2.3.2. Préparation et installation des sachets contenant le substrat artificiel ... 52
1.3. Paramètres mesurés en analyse d’eau ... 53
1.3.1. Paramètres physiques ... 53
1.3.2. Paramètres chimiques... 53
1.3.2.1. Flux de carbone organique dissous et d’azote inorganique dissous (FCOD et FNID) ... 54
1.4. Descripteurs de sédiments... 54
1.4.1. Granulométrie... 54
1.4.2. Matière sèche (MS) et matière sèche sans cendre (MSSC) ... 55
1.5. Mesure de la dénitrification... 55
1.5.1. Dénitrification in situ (DNT)... 57
1.5.2. Activité enzymatique dénitrifiante (DEA) ... 58
1.5.3. Calculs de production de N2O... 58
1.5.3.1. Concentrations de N2O in situ... 59
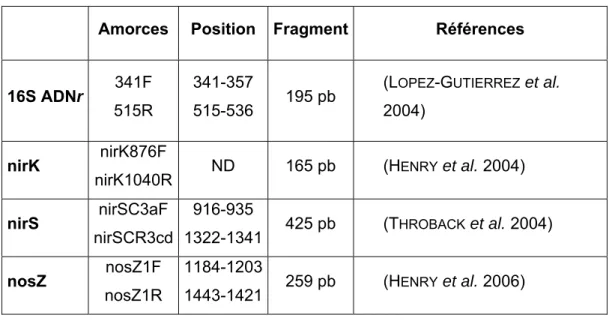
1.6.2.2. Choix des marqueurs : 16S ARNr et nosZ... 63
1.6.2.3. Analyse moléculaire : PCR-DGGE... 64
1.6.2.3.1. Extraction d’ADN ... 64
1.6.2.3.2. PCR (Polymerase Chain Reaction) ... 65
1.6.2.3.3. DGGE...67
1.6.2.3.4. Séquençage et analyse phylogénétique des OTUs de DGGE...68
1.6.2.3.5. Quantification par PCR en temps-réel ... 69
1.6.2.4. Diversité métabolique : BIOLOG EcoPlate ... 70
1.7. Unités d’expression des résultats ... 71
1.8. Méthodes Statistiques... 71
1.8.1. Tests non paramétriques... 71
1.8.2. Régression linéaire multiple pas à pas ... 71
1.8.3. Test d’imbrication (Nestedness test) ... 72
1.8.3. Test de Mantel... 73
1.8.4. Carte auto-organisées (Self-Organizing Map SOM)... 74
1.8.5. La Régression partielle aux moindres carrés (PLS) ... 75
1.8.6. Analyse Canonique des Correspondances (ACC)... 76
2. Les Biofilms Phototrophes du lit de la rivière... 78
2.1. Echantillonnage... 78
2.2 Descripteurs de la biomasse épilithique ... 80
2.3. Activité dénitrifiante : DEA limitée en carbone... 80
2.4. Typage des communautés bactériennes ... 81
2.5 Quantification par PCR en temps réel... 82
et DEA) ... 91
2.3. Variabilité temporelle de la dénitrification (DNT et DEA) ... 96
3. Discussion ... 102
4. Conclusions et perspectives ... 105
Chapitre IV. Rôle fonctionnel des communautés bactériennes attachées et libres dans l’aquifère alluvial... 108
1. Introduction ... 109
2. Analyse des différences entre la composition des communautés attachées et libres à l’intérieur (IHD) et à l’extérieur (OHD) de hot spots de dénitrification au sein d’un aquifère alluvial ... 111
3. Publication... 113
4. Résultats complémentaires à la publication ... 152
5. Conclusion et perspectives ... 155
Chapitre V. Etude de la structuration des communautés bactériennes attachées à travers une expérience de colonisation au sein de l’aquifère alluvial... 157
1. Introduction ... 158
2. Relation entre la structuration des communautés bactériennes dénitrifiantes et la dénitrification au cour d’une colonisation... 160
3. Publication... 162
4. Résultats et analyses complémentaires ... 189
4.1. Relation colonisation/flux nutritif ... 189
4.2. Communautés 16S ARNr... 190
4.3. Imbrication des communautés 16S ARNr de BH ... 192
4.4. Relation entre les communautés 16S ARNr et nosZ... 193
4.5. Structure des communautés dénitrifiantes... 193
6. Conclusion et perspectives ... 203
Chapitre VI. Relation structure fonction au sein d’un d’assemblage phototrophe: BP de rivière ... 206
1. Introduction ... 207
2. Résultats ... 208
3. Discussion ... 219
4. Conclusion et perspectives ... 220
Conclusion et perspectives générales... 223
Annexes ... 231
Liste des abréviations
ACC : analyse canonique des correspondances ACoP : analyse en coordonnées principales ADN : acide désoxyribonucléique
ARB : du latin Arbor (arbre)
ARNr : acide ribonucléique ribosomique AWCD : average well color development BH : biofilm hétérotrophe
BMU : best matching unit BP : biofilm phototrophe
CLPP : community level physiological profiles CPG : chromatographe en phase gaseuze DAPI : 4',6-diamidino-2-phenylindole
DEA : activité enzymatique dénitrifiante ou dénitrification potentielle DEA-LC : denitrification enzyme activity limited in carbone
DESUPS : diplôme des études supérieures de l’Université Paul Sabatier DGGE : denaturing gradient gel electrophoresis
DNT : denitrification in situ
EDTA : l'acide éthylène-diamine-tétraacétique FCOD : flux de carbone organique dissous FNID : flux d’azote inorganique dissous
INRA : institut national de la recherche agronomique MS : matière sèche
MSSC : matière sèche sans cendre OTU : operational taxonomic unit PCR : polymerase chain reaction PLS : Partial Least Square PRS : phosphate réactive soluble
PT : phosphate total
Ppmv : parties par million en volume
PRIP : prévention et réduction intégrées de la pollution Pb : paire de base
QMJ : débit moyen journalier
RDNA : réduction dissimilative du nitrate en ammonium Rpm : révolution par minute
SOM : self organizing map
T-RFLP : terminal-restriction fragment length Polymorphism TE : tris d'acide éthylène-diamine-tétraacétique
TS : température du système UV : ultra-violet
Introduction
L’intervention de l’Homme sur le cycle de l’azote a continuellement augmenté depuis 1860 avec l’évolution des pratiques agricoles et le développement de l’industrie. Ce processus s’est surtout intensifié depuis la seconde partie du XXème siècle. Les quantités des différentes formes d’azote réactif (e.g. NH4+, NH3, NOx,
HNO3, N2O, NO3-) générées par les pratiques agricoles ont été multipliées par 10 au
cours des dernières décennies (GALLOWAY et al. 2003). Dans les écosystèmes
aquatiques, l’augmentation d’un facteur 2 à 4 des concentrations en nitrate est clairement associée aux activités humaines (SEITZINGER et KROEZE 1998) et a
considérablement augmenté le risque d’eutrophisation des eaux douces et marines (MCISAAC et al. 2001 ; NIXON 1995). Parmi les différents écosystèmes aquatiques,
les zones riveraines revêtent un intérêt particulier par leur capacité à atténuer les apports de nitrate du bassin versant à la rivière et, en conséquence, dans les écosystèmes situés à l’aval (GALLOWAY et al. 2003 ; GILLIAM 1994). En effet, les
zones alluviales des grands fleuves constituent des zones d’interface entre le fleuve et les écosystèmes terrestres adjacents et notamment les agroécosystèmes. Elles constituent donc un des réceptacles du flux de nitrate d’origine agricole (fertilisants azotés d’origine industrielle et organique). Ces zones de contact entre les eaux souterraines et les eaux de rivière sont identifiées comme des zones clef pour la rétention des contaminants diffus (LOWRANCE et al. 1984 ; PETERJOHN et CORRELL
1984 ; DAHM et al. 1998). Leur capacité à purifier les eaux souterraines de certains
de leurs polluants comme les nitrates (MAITRE et al. 2002) peut résulter de trois
processus principaux : l’absorption racinaire (JORDAN et al. 1993 ; O'NEILL et
Ammonium) (TIEDJE 1988) peuvent concurrencer la dénitrification mais ils ont été
peu étudiés en milieu d’eau douce (BURGIN et HAMILTON 2007).
La dénitrification complète consiste en la réduction des nitrates (NO3-) en
azote gazeux (N2) via quatre séquences réductrices (NO3-→NO2-→NO→N2O→N2).
Ce processus a été identifié chez les bactéries depuis le XIXème siècle, par Gayon and Dupetit (GAYON et DUPETIT 1886) et aujourd’hui est étudié dans des milieux très
différents, de la colonne d’eau marine (JAYAKUMAR et al. 2004) jusqu’à l’estomac
des vers de terre (HORN et al. 2006). Lors d’une réduction incomplète des nitrates, la
dénitrification peut générer de l’oxyde nitrique (NO) et du protoxyde d’azote (N2O).
Dans le cas d’une dénitrification complète (production de N2), le processus peut être
considéré comme un « service écologique » réduisant les charges de nitrates dans les ressources d’eau, dans cas d’une dénitrification incomplète, au contraire, l’émission de NO et N2O a un impact négatif, ces deux gaz contribuant à l’effet de serre et à la
destruction de l’ozone stratosphérique (BOTHE et al. 2007). Dans les écosystèmes
riverains, la dénitrification est une fonction qui permet l’élimination d’une partie de l’excès d’azote dissous, diminuant les risques d’eutrophisation (GROFFMAN et TIEDJE
1989 ; WILLEMS et al. 1997 ; SABATER et al. 2003 ; VAN CLEEMPUT et al. 2007). On
estime que dans ces zones s’effectue par dénitrification l’élimination de 15 à 100% des nitrates transportés à travers les sédiments alluviaux de l’aquifère (MCMAHON et
BÖHLKE 1996 ; RUFFINONI et al. 2003). De nombreuses études utilisant l’approche
biogéochimique ont permis au cours des années antérieures de mieux comprendre l’importance et les facteurs de contrôle de la dénitrification dans les zones alluviales. Elles ont par ailleurs suscité de nombreuses questions concernant les acteurs biologiques de ce processus : les bactéries dites dénitrifiantes. Ce travail se base sur certaines de ces questions : Quelle est la différence entre les communautés dénitrifiantes libres et attachées ? Quelle est la dynamique spatiale et temporelle (hautes et basses eaux) de ces communautés bactériennes ? Comment ces communautés se structurent-elles, quels sont les facteurs qui contrôlent leur
structuration ? Quelles est la relation entre la composition des ces communautés et la dénitrification ?
Une nouvelle voie en Ecologie fonctionnelle consiste à décrire comment interagissent les fonctions de l’écosystème, la dynamique des communautés et les conditions abiotiques (LOREAU et al. 2001). Cette question concerne naturellement
les bactéries dont la contribution au fonctionnement des écosystèmes est reconnue
(SCHIMEL 1995). Or la contribution de la diversité bactérienne au fonctionnement des
écosystèmes est encore peu étudiée (BELL et al. 2005). Ce travail s’inscrit dans cette
logique en prenant les communautés bactériennes dénitrifiantes de différents assemblages bactériens (bactéries libres et fixées de l’aquifère, bactéries des biofilms phototrophes de rivière) comme modèle. La dénitrification est un trait assez répandu, puisqu’on estime que 5 à 15 % des bactéries que l’on rencontre dans des milieux naturels peuvent dénitrifier (TIEDJE 1988 ; CASELLA et PAYNE 1996 ; HENRY et al.
2006). Les différentes étapes de dénitrification (NO3-→NO2-→NO→N2O→N2) sont
catalysées par différentes enzymes que l’on regroupe sous l’appellation générique de N réductases dont la distribution chez les Bactéries est polyphylétique (ZUMFT
1997) ; (PHILIPPOT et al. 2001). Les bactéries dénitrifiantes constituent un groupe
fonctionnel intéressant car composé par des bactéries de différents genres comme
Pseudomonas, Ralstonia, Alcaligenes, Paracoccus, Rhodobacter, Rubrivivax, Thauera, Burkholderia, Bacillus et Streptomyces (ZUMFT 1992), sans que le
processus soit « trop » généraliste (par opposition, la dégradation des composés carbonés est un processus redondant (tel que défini par WALKER (1992). D’autre
part, la dénitrification est relativement facile à mesurer en laboratoire (TEISSIER et M.
En combinant des méthodes biogéochimiques et des méthodes de typage moléculaire, nous avons étudié le rôle fonctionnel des communautés bactériennes dénitrifiantes principalement au sein des Biofilms Hétérotrophes (BH) d’un aquifère alluvial.
Ce manuscrit se compose de deux parties (Figure 1). La première partie divisée en trois chapitres présente une synthèse bibliographique axée successivement sur les études de la composition des communautés bactériennes, les facteurs qui influencent leur composition et l’importance de la diversité sur les capacités fonctionnelles des communautés. Le modèle d’étude, la communauté dénitrifiante et la dénitrification sont aussi présentés. Dans le deuxième chapitre, nous décrivons les caractéristiques métaboliques, moléculaires et écologiques des bactéries dénitrifiantes afin de présenter en détail le modèle utilisé et les connaissances sur son rôle dans le cycle de l’azote. Pour terminer cette partie, nous avons introduit le rôle écologique des bactéries dénitrifiantes dans les systèmes aquatiques étudiés dans ce travail.
La seconde partie est divisée en cinq chapitres. Le premier décrit la méthodologie utilisée pour cette étude. Le deuxième porte sur la description des patrons de la dénitrification dans une zone alluviale de la Garonne avec des comparaisons de mesures de l’activité in situ (Dénitrification iN siTu-DNT) et de l’activité enzymatique dénitrifiante (Denitrification Enzyme Activity - DEA). Afin de pouvoir identifier les rôles respectifs des communautés bactériennes associées aux compartiments eau et sédiment, nous avons comparé la composition des communautés attachées au sédiment (biofilm) et libres dans l’eau souterraine au sein des zones alluviales de forte et faible dénitrification. Ceci nous a dirigé pour le quatrième chapitre vers l’étude de la structuration des communautés impliquées dans la dénitrification dans l’aquifère alluvial, les communautés dénitrifiantes au sein du BH. L’objectif était d’étudier la dynamique des ces communautés au cours d’une
explique l’hétérogénéité (patchiness) observée au niveau de la dénitrification dans l’aquifère alluvial.
Figure 1 : Schéma récapitulatif des problématiques abordées dans la partie expérimentale du travail (chapitre III, IV, V et VI). DNT : dénitrification in situ ; DEA : activité enzymatique dénitrifiante et DEA-LC : activité enzymatique dénitrifiante limitée en carbone.
Enfin, dans le chapitre VI nous avons étudié la relation entre la diversité dénitrifiante et la dénitrification sur le biofilm phototrophe de rivière (BP) afin de déterminer comment l’écosystème (rivière) et la composition de l’assemblage (présence d’organismes phototrophes) agissent sur la sélection des bactéries dénitrifiantes et les conséquences de cette action sur la dénitrification. La dernière partie de ce travail est une synthèse des résultats acquis et une présentation des
Chapitre III, IV et V Chapitre VI
Dénitrification (DNT et DEA) Rôle fonctionnel des bactéries attachées et libres BIOFILM HETEROTROPHE Structuration de communautés bactériennes Dénitrification (DEA-CL) BIOFILM PHOTOTROPHE Diversité communautés Diversité taxonomique Diversité fonctionnelle Diversité taxonomique Diversité fonctionnelle
Chapitre III, IV et V Chapitre VI
Dénitrification (DNT et DEA) Rôle fonctionnel des bactéries attachées et libres BIOFILM HETEROTROPHE Structuration de communautés bactériennes Dénitrification (DEA-CL) BIOFILM PHOTOTROPHE Diversité communautés Diversité taxonomique Diversité fonctionnelle Diversité taxonomique Diversité fonctionnelle
Chapitre I. Relation entre la
composition des communautés
bactériennes et le fonctionnement
des écosystèmes : état de l’art
Ce premier chapitre a pour objectif de présenter, avant la description des différentes parties expérimentales, le contexte général dans lequel s’inscrit ce travail. Il décrit les modèles utilisés, les communautés bactériennes dénitrifiantes, et donne un aperçu de l’environnement dans lequel ces communautés ont été étudiées : les aquifères alluviaux.1. Les communautés bactériennes
1.1.
Définition
Il est difficile de déterminer quand le terme de communauté a été utilisé pour la première fois avec les bactéries, mais de nombreuses études ont décrit la composition de communautés bactériennes présentes dans différents milieux, aquatiques (CRUMP et al. 1999 ; DANG et LOVELL 2000 ; ARAYA et al. 2003) comme
terrestres (RANJARD et al. 2000 ; WEBSTER et al. 2003 ; RICH et MYROLD 2004).
En Ecologie, la communauté est généralement définie comme un ensemble de populations, i.e. d’individus de différentes espèces qui coexistent en même temps et au même endroit (BEGON et al. 1996). Mais cette définition est trop générale pour
être appliquée directement aux communautés bactériennes. En Ecologie microbienne, le terme communauté peut désigner des assemblages d’espèces bactériennes qui sont en interaction dans un même écosystème, milieu, habitat et/ou qui partagent une même fonction. Dans notre manuscrit, le terme communauté sera utilisé avec une connotation fonctionnelle en parlant par exemple de la communauté
dénitrifiante qui rassemble l’ensemble des bactéries partageant la capacité à
dénitrifier. Les assemblages qui sont présents dans un même milieu mais dans différents micro-habitats peuvent également être désignés comme des communautés différentes, par exemple les communautés libres (i.e. bactéries en suspension dans le milieu aquatique) et les communautés attachées (i.e. bactéries incluses dans un biofilm à la surface d’un substrat solide) décrites dans certains travaux (LEHMAN et
1.2. Diversité des communautés bactériennes
1.2.1. Définition de la biodiversité
“La diversité biologique est la variabilité entre les êtres vivants de toutes origines ; ceci inclut la diversité au sein des espèces, entre les espèces et des écosystèmes”. Cette définition est celle retenue lors de la Convention de la Diversité Biologique signée par plus de 150 nations à Rio de Janeiro en 1992 (CBD 2001). Dans son acception la plus large, le terme de biodiversité fait donc référence à l’amplitude de la variabilité du vivant. Il s’applique aux différents niveaux d’organisation biologique, des gènes jusqu’aux écosystèmes en passant par les individus, populations et communautés.
1.2.2.
La
diversité
bactérienne
Les bactéries seraient plus de 1030 individus sur Terre (WHITMAN et al. 1998)
et même si l’on considère généralement que la diversité bactérienne ne représentent qu’un faible pourcentage de la biodiversité globale (sur toutes les espèces identifiées 0,2% seraient des bactéries (CBD 2001), la grande diversité des ressources énergétiques et voies métaboliques qu’elles utilisent leur confère un rôle majeur dans le fonctionnement des écosystèmes. En microbiologie, la notion d’espèce (Bergey's
Manual of Systematic Bacteriology 2nd edition, (2001)) est utilisée pour désigner une
collection de souches microbiennes qui partagent plusieurs propriétés similaires et différentes de celles d’autres groupes de souches. Les Bactéries peuvent être classées en trois groupes par exemple sur la base de leur morphologie : les Gram + (Bactéries avec une paroi homogène composé de peptidoglycane), les Gram – (Bactéries avec une paroi plus complexe composée d’une couche de petidoglycane et d’une membrane externe) et celles qui ne sont pas dotées de paroi. Les autres caractéristiques utilisées sont les caractéristiques biochimiques, la forme de la cellule, la morphologie des colonies etc. Actuellement, la classification
phylogénétique basée sur les séquences de l’ARN 16S ribosomal (16S ARNr) illustre l’hétérogénéité existant dans le domaine des Bactéries (Figure I.1).
L’étude de la diversité bactérienne est importante pour connaître la composition génétique de la communauté, comprendre les patrons de distribution des bactéries, améliorer la connaissance de ces communautés et gérer la diversité pour permettre le fonctionnement et la viabilité des écosystèmes (OVREAS 2000).
Traditionnellement, le terme de diversité se réfère à une diversité dont l’unité de base est l’espèce, cependant chez les bactéries plus que chez les autres organismes, définir l’espèce est problématique en absence de véritable barrière aux flux de gènes. L’utilisation du niveau taxonomique d’espèce chez les bactéries a cependant été considérée comme pratique, pragmatique, commode et arbitraire (WARD 1998). La
diversité bactérienne décrit le nombre d’espèces bactériennes et leur abondance relative dans une communauté, ou la quantité et la distribution de l’information (génétique) dans la communauté (définition proposée par Atlas, 1984 cité par
Figure I.1. Arbre phylogénétique basé les séquences de 16S ARNr montrant les principaux phyla bactériens. Les triangles représentent les groupes d’organismes apparentés, l’angle à la racine indique approximativement le nombre de séquences disponibles et les limites représentent la plus courte et la plus longue branche à l’intérieur du groupe. L’arbre a été reconstruit, évalué et optimisé en utilisant l’outil parcimonie dans le logiciel ARB. Seule les positions de séquence partageant des résidus identiques à au moins 50% de toutes les séquences bactériennes ont été inclues dans le calcul. L’arbre a été enraciné en utilisant toutes les séquences homologues disponibles et complètes chez les Archaea et les Eucarya prises comme référence hors-groupe (représenté par la flèche). La longueur des triangles représente la profondeur phylogénétique des lignées principales. La barre correspond à 10% de divergence de la séquence estimée (extrait de LUDWIG et KLENK
(2001)).
La diversité des bactéries peut être étudiée au niveau phénotypique, sur la base de leur composition en acides gras (pour revue cf. OVREAS 2000) mais aussi sur
la base de leurs propriétés métaboliques comme l’utilisation de différentes sources de carbone (en utilisant le kit commercial Biolog par exemple, GARLAND et MILLS
Le développement des techniques de biologie moléculaire a recemment permis d’aborder la diversité génotypique des communautés bactériennes. Le développement des méthodes d’empreinte génétique - i.e. la DGGE (Denaturing Gradient Gel Electrophoresis (MUYZER et al. 1997), la T-RFLP
(Terminal-Restriction Fragment Length Polymorphism (LIU et al. 1997) ou le clonage (MEAD
et al. 1991) – a permis de s’affranchir des techniques de culture très contraignantes
(accès à seulement 1% de la population totale (AMANN et al. 1995) et associées à des
biais inévitables (effet de sélection). Ces techniques de biologie moléculaire ont également donné accès à une diversité bactérienne plus large que la seule diversité cultivable dans les écosystèmes étudés. Elles restent cependant encore loin de pouvoir fournir une idée de la diversité bactérienne dans sa globalité (WHITMAN et
al. 1998).
1.2.3. La diversité fonctionnelle bactérienne
Les mécanismes par lesquels la diversité influence le fonctionnement de l’écosystème sont liés aux attributs des espèces plutôt qu’au nombre total d’espèces présentes (GILLER et al. 2004). Certaines bactéries sont autotrophes, d’autres
hétérotrophes, certaines sont aérobies strictes, d’autres aérobies facultatives et d’autre encore anaérobies. Parmi les hétérotrophes, on trouve même des photosynthétiques. Les bactéries sont donc responsables de la majorité des processus biogéochimiques dans les écosystèmes terrestres et aquatiques (BELL et al. 2005).
Elles recyclent la matière organique pour les plantes et autres organismes autotrophes et sont même capables de dégrader des composés xénobiotiques. Au sein des écosystèmes aquatiques, une de leur fonction est de dégrader la matière organique et de recycler des éléments inorganiques (soufre, phosphore, azote) par des voies
communauté est capable de réaliser (GASTON et SPICER 1998). L’isolation de
souches bactériennes a permis d’identifier les capacités fonctionnelles de communautés bactériennes depuis (WINOGRADSKY 1948). Mais, depuis le
développement des marqueurs fonctionnels (gènes utilisés en typage moléculaire des communautés bactériennes pour étudier la diversité fonctionnelle (TORSVIK et
OVREAS 2002) en biologie moléculaire, i.e. amoA pour les bactéries
ammonio-oxydantes, nirS bactéries dénitrifiantes, la diversité fonctionnelle des communautés bactériennes cultivables et non-cultivables est accessible.
1.2.4. La mesure de la diversité
La description de la diversité impose que celle-ci soit quantifiée. Si en théorie cette question peut paraître simple et se résumer à déterminer quelles entités sont présentes (espèces, allèles) et par combien d’unités chacune est représentée (nombre d’individus, nombre de copies) (GASTON et SPICER 1998), dans la pratique cette
quantification est généralement plus difficile. Par exemple, s’agissant de la variabilité au sein d’une espèce, plusieurs critères peuvent être retenus pour définir les différentes entités et la variabilité peut aussi bien être considérée sous l’angle de la biochimie (ex. sérotypes), de l’écologie, (ex. la Brocadia anammoxidans capable de nitrifier et dénitrifier (MULDER et al. 1995)), de la morphologie (ex. Vibrio
halioticoli à formes bacillaire et coco-bacillaire selon le type de milieu de culture
utilisé) ou encore au regard du rôle que cette variabilité joue sur les aspects fonctionnels de l’espèce au sein d’une communautés donnée (ex. prédateur, proie). Il n’existe pas de mesure unique qui puisse embrasser cette diversité (GASTON et
SPICER 1998).
Les méthodes moléculaires basées sur le marqueur 16S ARNr se sont substituées aux méthodes de cultures en donnant accès aux bactéries non cultivables (AMANN et al. 1995 ; MADSEN 2004). Néanmoins, la magnitude de la diversité
de celle-ci reste parmi les problèmes de l’écologie bactérienne actuelle (TORSVIK et
al. 2002).
La diversité α, ou diversité dans l’échantillon, est exprimée généralement à l’aide de différents indices mathématiques. Parmi ceux-là, les indices de Simpson
(SIMPSON 1949) et de Shannon (SHANNON et WEAVER 1963) sont les plus utilisés
pour les communautés bactériennes. Ce sont des indices qui se basent sur le nombre d’espèces et l’abondance relative de chacune : D = Σ (NS / NT)² (Simpson) et H = - Σ [(NS / NT) x log2 (NS / NT)] (Shannon) ; NS :nombre d’individus par taxon et NT=nombre total de taxons.
Pour comparer directement des échantillons deux à deux, et ainsi obtenir la diversité β on utilise souvent les indice de Jaccard : J = [c / (a+b-c)] x 100 où a : bandes présentes seulement dans l’échantillon A ; b : bandes présentes seulement dans l’échantillon B et c : bandes totales communes aux échantillons A et B.
1.3. Facteurs qui affectent la composition d’une
communauté bactérienne dans un aquifère
1.3.1
Facteurs
abiotiques
Comprendre les changements de composition des communautés bactériennes passe par la détermination des facteurs qui affectent ces communautés. A l’échelle locale, dans un écosystème, la disponibilité des ressources nutritives est un facteur important. Une des premières études sur les communautés dénitrifiantes de l’aquifère
aquifère alluvial dépend des concentrations en nutriments organiques et inorganiques dans l’eau, en accord avec JUDD et al. (2006) qui a observé une forte relation entre le
taux de carbone organique dissous et la composition des communautés bactériennes de rivière. Dans le cas des communautés bactériennes de lacs, plutôt que la concentration, c’est le type de carbone disponible qui agit sur la composition des communautés bactériennes d’après EILER et al. (2003).
Dans le cas des aquifères, la composition des communautés bactériennes peut également varier en fonction de la disponibilité en carbone et des niveaux de contamination par des composés xénobiotiques. RÖLING et al. (2001) montre que des
zones polluées par du benzène, du toluène, de l’éthyl-benzène et du xylène ont des compositions bactériennes qui diffèrent de celles des zones non contaminées. Dans une étude en milieu souterrain comparant deux milieux caractérisés par des concentrations en herbicides contrastées, DE LIPTHAY et al. (2004) observe un impact
sur la structure (morphotypes, formes de colonies) mais pas sur les diversités génétique et métabolique.
La perturbation de l’écosystème – i.e. un changement brutal et imprévisible des caractéristiques de l’habitat – cause des changements importants sur les communautés des différents organismes. Dans le cas des aquifères, une crue est considérée comme une perturbation (BOULTON et al. 1998). En général, les
communautés bactériennes répondent aux perturbations du milieu comme les macro-organismes, mais leurs taux de croissance généralement plus élevés permettent une réponse plus rapide. Au cours de la perturbation ou dans les situations post-perturbation, tous les taxons de la communauté ne sont pas égaux. Certains, par leur capacité à s’adapter à la nouvelle situation, leur l’abondance relative avant la perturbation peuvent prendre le dessus sur les autres et en conséquence accroitre leur représentation au sein de la communauté. Dans une étude de l’effet des degrés et types de perturbation (artificiellement crées) sur la composition des communautés
composant une communauté pour le maintient d’un processus dans un écosystème. La stabilité de la communauté, c’est à dire l’absence d’extinction ou d’explosion démographique de certaines espèces suite à une perturbation, dépend de la diversité dans la communauté : plus la diversité est importante plus la communauté est stable
(LOREAU et al. 2003). L’hypothèse d’assurance temporelle propose que la
biodiversité joue le rôle d’une « assurance » face aux variations environnementales si les espèces sont fonctionnellement complémentaires dans le temps. L’hypothèse d’assurance spatiale applique cette idée dans une métacommunauté (somme de communautés reliées par la dispersion), en considérant que la complémentarité temporelle peut venir d’une complémentarité spatiale et de taux de dispersion appropriés entre les communautés (LOREAU et al. 2003).
1.3.2.
Facteurs
biotiques
Parmi les facteurs biotiques qui affectent la composition des communautés bactériennes, on identifie la plasticité métabolique et physiologique des communautés. Ces facteurs biotiques ont souvent été étudiés à travers la manipulation des concentrations en éléments nutritifs ou des conditions physiques du milieu dans des expériences au laboratoire (PEACOCK et al. 2004 ; FINDLAY et al.
2003). Certain auteurs ont aussi testé la résistance des communautés (i.e. le maintien de la diversité et de la fonction) face à différentes concentrations de contaminants comme le nickel (LAWRENCE et al. 2004). Pour les communautés bactériennes de
biofilms hyporhéiques, la plasticité du métabolisme azoté, très répandue chez les bactéries, permet l’assimilation de différentes formes d’azote et induit que les variations des concentrations en azote sous ses différentes formes affectent la composition métabolique mais pas la structure taxonomique (F et al. 2003).
l’histoire de la communauté : une espèce installée ayant de ce seul fait un avantage sur d’autres espèces arrivant ultérieurement lorsque les ressources ne sont pas limitantes. Cependant, cette étude a été réalisée avec des ciliés (Colpidium sp.,
Paramecium sp. et Tetrahymena sp.) et à notre connaissance il n’existe pas d’étude
similaire conduite sur des communautés bactériennes.
Les organismes unicellulaires grâce à leur petite taille, leur abondance et leur plasticité métabolique sont peu contraints dans leur dispersion. Au début du XXème siècle, les scientifiques ont commencé à s’intéresser à la distribution spatiale des communautés bactériennes (BEIJERINCK 1913 ; BAAS BECKING 1934). Depuis, la
distribution des bactéries est considérée comme ubiquiste mais récemment plusieurs études ont démontré l’existence de l’endémisme chez les procaryotes (pour revue cf.
RAMETTE et TIEDJE (2006)). La distribution géographique des communautés
bactériennes dépend de la spéciation, des extinctions et de la dispersion (RAMETTE et
TIEDJE 2006). La distribution des communautés bactériennes au sein d’un
écosystème donné est aussi la conséquence des exigences de niche des membres de cette communauté. Les processus qui agissent sur la géographie des communautés bactériennes comparée aux eucaryotes ont été récemment abordés par HORNER
-DEVINE et al. (2004).
2. Rôle de la diversité sur le fonctionnement d’un écosystème
“It appeared that a plot of ground sown with several different types of grass was more productive than a similar plot with just one species of grass” remarquait
Darwin dans On the origin of species by mean of natural selection (1859). Il faisait référence au travail de George Sinclair, chef des jardiniers du Duc de Bedford au début du XIXème siècle qui, à partir de plantations combinant différentes espèces végétales et différents types de sols dans le jardin de Woburn Abbey, a réalisé la première étude expérimentale sur la relation entre biodiversité et fonctionnement
d’un écosystème (HECTOR et HOOPER 2002). Cependant, ce n’est que cent ans plus
tard, avec la publication de Biodiversity and ecosystem functioning (SCHULZE et
MOONEY 1994), que l’étude de la relation entre biodiversité et fonctionnement d’un
écosystème est devenue un enjeu majeur d’une discipline en passe de s’imposer comme telle : l’ « Écologie fonctionnelle ».
Depuis que les extinctions d’espèces et leur introduction dans de nouveaux écosystèmes connaissent un rythme inégalé du fait des activités humaines, la biodiversité a été placée au centre de questions scientifiques et d’enjeux de politique et de gestion (LOREAU et al. 2001 ; HOPPER et al. 2005 ; SRIVASTAVA et VELLEND
2005 ; DIVERSITAS 2005). Ceci est d’autant plus vrai que les changements de
biodiversité s’accompagnent parfois d’un impact clairement négatif d’un point de vue anthropocentrique et utilitariste. Ainsi, la qualité des biens et services délivrés par les écosystèmes (ecological service and goods sensu CHAPIN III et al. (2000))
peut être négativement affectée par une modification de leur biodiversité.
2.1. Des modèles théoriques aux études empiriques
L’observation répétée d’une relation entre modification de la biodiversité et altération des biens et services produits par un écosystème conduit à s’interroger sur la nature de cette relation et sur les mécanismes qui la déterminent ; ceci particulièrement afin de prédire les conséquences de gains et pertes de biodiversité. Dans ce contexte, la biodiversité additionnée ou perdue doit être abordée sur le plan fonctionnel et l’étude de la relation entre la structure des communautés et le fonctionnement de l’écosystème devient centrale. VITOUSEK et HOOPER (1993) ont
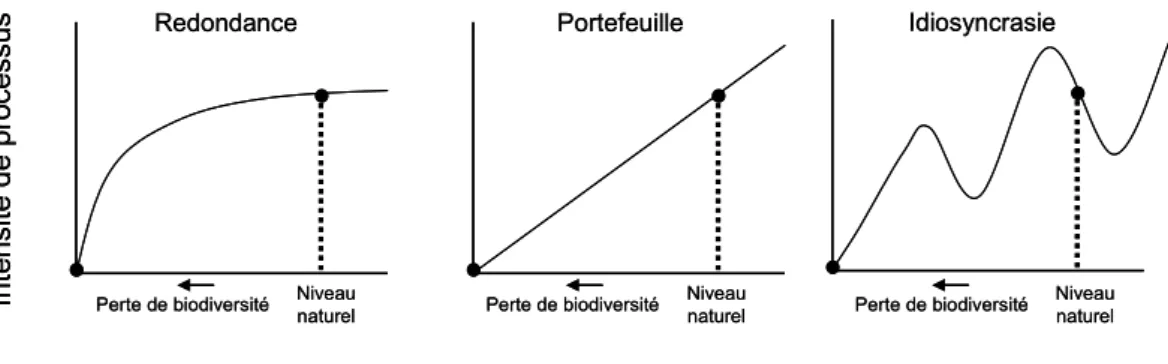
représentant l’intensité d’un processus en fonction de trois modèles de relation à la biodiversité. Dans le cas de l’hypothèse de redondance (i.e. redondance fonctionnelle entre les espèces sensu (WALKER 1992), le processus étudié est peu affecté par
l’érosion de la biodiversité, chaque espèce étant capable d’assurer le processus considéré à la place des espèces supprimées. Dans le cas de l’hypothèse du portefeuille, par référence à un portefeuille d’actions, le principe retenu est celui d’une additivité simple : chaque espèce assure une part du processus de façon irremplaçable. Enfin, l’idiosyncrasie propose que les conséquences de la disparition d’une espèce dépendent des conditions abiotiques du milieu, donc l’impact sur le processus considéré sera variable.
Figure I.2. Représentations graphiques des hypothèses de la redondance, du portefeuille et de l’idiosyncrasie régissant la relation entre biodiversité et fonction d’un écosystème (modifié d’après (NAEEM et al. 2002).
Si ces modèles théoriques sont disponibles, leur validation empirique est insuffisante. Notamment il reste à préciser dans quelles conditions et quels types d’écosystèmes chacun peut être appliqué.
L’intérêt de coupler la biodiversité au fonctionnement de l’écosystème a été récemment promu par (KINZIG et al. 2001). Toutefois, l’étude de la relation
biodiversité/fonction de l’écosystème et encore peu développée, surtout s’agissant du domaine de l’écologie des micro-organismes.
Biodiversité In te ns ité d e pr oc es su
s Redondance Portefeuille Idiosyncrasie
Niveau naturel
Perte de biodiversité Perte de biodiversité Niveau naturel Niveau naturel Perte de biodiversité Biodiversité In te ns ité d e pr oc es su
s Redondance Portefeuille Idiosyncrasie
Niveau naturel
Perte de biodiversité Perte de biodiversité Niveau naturel Niveau
naturel Perte de biodiversité
2.2. Comment mesurer l’intensité de la relation
diversité/fonction
? Quels processus et communautés
bactériennes ont été étudiés ?
S’agissant des études de relation diversité/fonction, le but n’est généralement pas de rechercher une relation entre espèces et processus, pour plusieurs raisons. D’une part parce qu’une telle relation serait difficile à établir chez les bactéries pour lesquelles la validité du concept d’espèce est mise en question. D’autre part, parce que les méthodes de typage moléculaire citées plus haut ne permettent pas évaluer le nombre d’espèces et de mesurer la diversité des complexes chez les procaryotes (BENT et al. 2007). En revanche, ces techniques révèlent des informations nouvelles
sur l’écologie, la biogéographie et la microbiologie appliquée des assemblages bactériens – i.e. les interaction conditions environnementales/communautés bactériennes, le rôle fonctionnel des bactéries, la distribution des communautés bactériennes, les interaction entre les différentes espèces, etc.
La plupart des études qui ont mis en évidence l’effet de la biodiversité sur le fonctionnement de l’écosystème ont été réalisées dans des écosystèmes terrestres, avec des plantes (TILMAN 1996 ; NAEEM et al. 1996). Pour les communautés
bactérienne, la relation entre structure de communautés et activité a été plus étudiée dans les milieux terrestres tels que les sols (CAVIGELLI et ROBERTSON 2000 ;
HOLTAN-HARTWIG et al. 2000 ; HORZ et al. 2004). Les premières études qui ont
montré des couplages entre la diversité des communautés dénitrifiantes et activité dénitrifiantes se sont basé sur des mesures de cinétiques de dénitrification dans des sols géomorphologiquement similaires mais avec une occupation végétale et des
respiration, la minéralisation de composants simples de carbone ou l’immobilisation du carbone) n’a pas permis de trouver un couplage entre diversité et activité. De plus, les études sont souvent réalisées sur des diversités bactériennes très complexes à analyser (en terme de nombre d’espèce), ex. diversité des bactéries dans les sols (RICH et MYROLD 2004 ; ENWALL et al. 2005). Récemment, une expérimentation sur
le rôle fonctionnel des bactéries en milieu contrôlé avec des assemblages « artificiels » a été conduite avec l’objectif de trouver une relation espèce/processus (BELL et al. 2005). A travers l’étude du processus de la respiration bactérienne, ces
auteurs confirment que les interactions synergiques entre les espèces bactériennes ainsi que la composition des communautés sont des facteurs importants qui jouent sur un rôle fonctionnel dans l’écosystème. Mais manipuler la biodiversité sans tenir compte des conditions environnementales reste artificiel ou trompeur (WELLNITZ et
POFF 2001). La réponse fonctionnelle des bactéries dépend de l’assemblage dont
elles font partie (HANSEN et al. 2007) donc cette approche ne donne qu’une idée
partielle de la réalité.
3. Le modèle d’étude : les communautés dénitrifiantes des
agrégats naturels
3.1. La dénitrification
3.1.1.
Définition
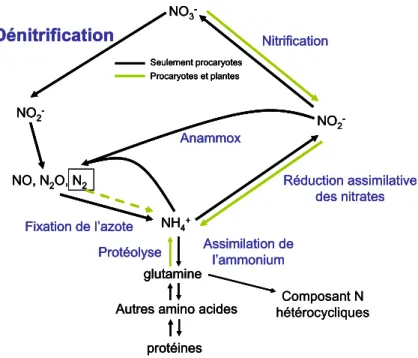
La dénitrification est un processus biologique qui participe au cycle de l’azote
(KNOWLES 1982 ; BOTHE et al. 2007) (Figure I.3). Même si certains eucaryotes
comme les champignons (ZUMFT 1997) et les foraminifères (RISGAARD-PETERSEN et
al. 2006) sont capable de dénitrifier, ce processus est principalement assuré par les
bactéries (ZUMFT 1997). Ce processus hétérotrophe, véritable « respiration des
concentrations d’oxygène ou en son absence, les nitrates jouent le rôle d’accepteur final d’électron.
Figure I.3. Le cycle biologique de l’azote (extrait de BOTHE et al. (2007)).
La dénitrification consiste en une réduction séquentielle, en quatre étapes, des nitrates en azote gazeux. Dans chaque étape différentes enzymes interviennent (Figure I.4). La première étape consiste en la transformation des nitrates en nitrites. Elle est catalysée par deux types de molybdo-enzymes : une nitrate-réductase liée à la membrane (Nar) et une nitrate-réductase periplasmique (Nap).
NO3 -NO2 -NO, N2O, N2 NH4+ NO2 -Dénitrification Nitrification Anammox Fixation de l’azote glutamine Autres amino acides
protéines
Protéolyse Assimilation de l’ammonium
Réduction assimilative des nitrates Seulement procaryotes Procaryotes et plantes Composant N hétérocycliques NO3 -NO2 -NO, N2O, N2 NH4+ NO2 -Dénitrification Nitrification Anammox Fixation de l’azote glutamine Autres amino acides
protéines
Protéolyse Assimilation de l’ammonium
Réduction assimilative des nitrates Seulement procaryotes Procaryotes et plantes Composant N hétérocycliques NO3 -NO2 -NO, N2O, N2 NH4+ NO2 -Dénitrification Nitrification Anammox Fixation de l’azote glutamine Autres amino acides
protéines
Protéolyse Assimilation de l’ammonium
Réduction assimilative des nitrates Seulement procaryotes Procaryotes et plantes NO3 -NO2 -NO, N2O, N2 NH4+ NO2 -Dénitrification Nitrification Anammox Fixation de l’azote glutamine Autres amino acides
protéines
Protéolyse Assimilation de l’ammonium
Réduction assimilative des nitrates Seulement procaryotes Procaryotes et plantes Composant N hétérocycliques
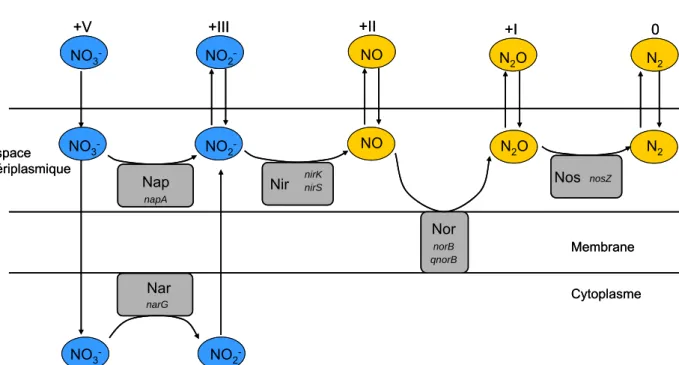
Figure I.4. Séquences réductrices de la dénitrification avec une représentation de la localisation des enzymes par rapport à la membrane cytoplasmique. Nar : nitrate réductase ; Nap : nitrate réductase liée à la membrane ; Nir : nitrite réductase ; Nor : oxyde nitrique réductase, Nos : oxyde nitreux réductase. Les noms de gènes en italique codent pour les sous unités catalytiques. Les composé dissous sont représentés en bleu et les produits gazeux en orange. Les chiffres romains correspondent à la valence de l’azote dans chaque molécule azotée. (modifié d’après (WALLENSTEIN et al. 2006).
La réduction des nitrites en oxyde nitrique est catalysée par deux enzymes de structure et métaux prosthétiques différents : l’une avec du cuivre (Cu-Nir) et l’autre avec du cytochrome cd1 (Cd-Nir). L’étape suivante consiste en la transformation de l’oxyde nitrique en protoxyde d’azote. Cette étape est aussi catalysée par deux enzymes : l’une reçoit l’électron du cytochrome c ou de la pseudoazurin (cNor) et l’autre d’un pool de quinol (qNor). La dernière étape de la dénitrification, la transformation du protoxyde d’azote en azote gazeux, est une étape pour laquelle on ne connaît qu’une seule enzyme (Nos) localisée dans le périplasme des bactéries Gram-. L’identification de cette enzyme a constitué une découverte méthodologique majeure dans l’étude de la dénitrification (YOSHINARI et KNOWLES 1976).
Espace Périplasmique Nor Nos NO3- NO2- NO N2O N2 NO3- NO2 -NO2- NO N 2O N2 Membrane Cytoplasme norB qnorB nosZ NO3 -Nap napA
Nir nirKnirS
Nar narG +V +III +II +I 0 Espace Périplasmique Nor Nos NO3- NO2- NO N2O N2 NO3- NO2 -NO2- NO N 2O N2 Membrane Cytoplasme norB qnorB nosZ NO3 -Nap napA
Nir nirKnirS
Nar
narG
La dénitrification est étudiée depuis longtemps car c’est un processus simple à mesurer au laboratoire (TIEDJE 1994). Elle a été très étudiée dans les milieux
terrestres et aquatiques (pour revue cf. SEITZINGER et al. (2006)). Même si l’une des
premières études en milieu naturel a été réalisée dans le sol (NÕMMIK 1956), ce
processus est largement étudié dans les systèmes aquatiques (BRADLEY et al. 1992 ;
LOWRANCE 1992 ; BEGTSSON et BERGWALL 1995 ; ADDY et al. 2002 ; CLEMENT et
al. 2003, KELLOGG et al. 2005, LOWRANCE 1992). SMITH et DUFF (1988) ont été
parmis les premiers à identifier l’importance de la dénitrification dans les aquifères. La plupart des études ont eu comme objectif la description des patrons de dénitrification à l’échelle de l’écosystème (PINAY et al. 1993 ; BAKER et VERVIER
2004) et à l’échelle régionale (BURT et al. 2002 ; PINAY et al. 2007).
3.1.2. Où a lieu la dénitrification ?
Les modèles globaux qui décrivent la distribution spatiale de la dénitrification au niveau des écosystèmes montrent que les sédiments des plateaux continentaux assurent la plus grande part de la dénitrification (44%) suivis par les sols (22%) et les zones océaniques en condition d’anoxie (12%) (SEITZINGER et al. 2006). Les
systèmes d’eau douce (aquifères, lacs, rivières) réaliseraient 20% de la dénitrification totale et les estuaires 1% d’après (SEITZINGER et al. 2006). Un tel processus est aussi
utilisé comme mécanisme d’épuration dans des sites comme les zones humides artificielles (VAN CLEEMPUT et al. 2007 ; KJELLIN et al. 2007).
Dans le continuum bassin versant/océan, les zones riveraines sont des intermédiaires obligatoires lors du transport des nitrates provenant des terres
SANCHEZ PEREZ et al. 2003a ; PINAY et al. 2007). Cette information est indispensable
pour mesurer l’impact de l’activité humaine sur le cycle de l’azote et gérer et réduire les conséquences environnementales associées aux pollutions azotées. Mais selon BOYER et al. (2006) la modélisation de ce processus reste encore ardue à cause de la
variabilité des contrôles dominants de la dénitrification dans le temps et l’espace (qui favorisent l’existence de « hotspots » et « hot moments » (MCCLAIN et al.
2003)) et du manque de connaissances sur les communautés bactériennes responsables de ce processus.
3.1.3. Les facteurs contrôlant la dénitrification
dans le milieu aquatique
Les conditions requises pour que la dénitrification ait lieu sont d’abord la présence de donneurs d’électrons, c’est à dire (généralement) de sources de carbone organique, ensuite une anaérobiose stricte ou modérée qui régule l’activité enzymatique (concentration en O2 environ <0,2 mg L-1), la présence de formes
azotées oxydantes comme les nitrates, les nitrites, les oxydes nitriques ou le protoxyde d’azote et enfin la présence des bactéries capables de réaliser ce processus. L’intensité de la dénitrification est donc susceptible d’être conditionnée par des variations de ces différents facteurs abiotiques et biotiques, comme décrit ci-dessous.
L’influence de chaque facteur sur la dénitrification est dépendante du type d’écosystème au sein duquel les interactions conditions environnementales/dénitrification sont analysées. Parmi les facteurs environnementaux qui affectent le taux de dénitrification, la production de N2O et/ou
de N2, le pH, le carbone, la température, la disponibilité des nitrates et nitrites, la concentration d’oxygène ont été identifiés (KNOWLES 1982) mais aussi l’humidité, la
porosité, l’alternance gel/dégel, les cycles de séchage/remouillage dans le cas des sols où la plupart de ces facteurs sont étudiés (pour revue cf. PHILIPPOT et al. (2007)).
démontré que la dénitrification est plus limitée par la disponibilité du carbone que par les nitrates, BRADLEY et al. (1992) ont réalisé un des premiers travaux identifiant
le carbone comme principal facteur limitant de la dénitrification, parmi d’autres comme les concentrations en nitrates et le pH ; ce résultat a été confirmé ensuite par (HILL et al. 2000). Dans cette interface terrestre/aquatique, les processus
bio-géochimiques comme la dénitrification sont contrôlés par deux facteurs dominants l’azote et le carbone (MCCLAIN et al. 2003). Ces facteurs sont eux même sous
influence de l’hydrologie ou d’autres conditions abiotiques (flux, profondeur, vitesse, mélanges des eaux) qui modifient leur répartition et disponibilité. La dynamique des flux qui distribuent les nutriments dans l’aquifère dépend fortement de sa topographie (PFEIFFER et al. 2006). Ainsi la dénitrification diminue quand
l’éloignement à la rivière augmente (KELLOGG et al. 2005) car avec l’éloignement les
apports en carbone assurés par les eaux de surface diminuent (BRUGGER et al. 2001).
Les facteurs abiotiques régulent le taux de dénitrification de façon instantanée, mais ils agissent aussi à long terme sur la composition des communautés dénitrifiantes (sensu distal control (WALLENSTEIN et al. 2006)). Depuis que
CAVIGELLI et ROBERTSON (2000) et HOLTAN-HARTWIG et al. (2000) ont suggéré que
la composition des communautés dénitrifiantes est importante pour la dénitrification
in situ et l’émission de N2O, certains travaux ont conduit à des conclusions
différentes. Par exemple RICH et MYROLD (2004) étudiant le taux de dénitrification
et les communautés dénitrifiantes de différents types de sols, indiquent que la composition des communautés dénitrifiantes n’explique pas la variabilité de l’activité. Ce travail concernait des communautés issues de différents écosystèmes (sédiments de rivière et sols). Ces mêmes auteurs ont également réalisé une étude sur
3.1.4. Service écologique et incidence
agronomique de la dénitrification
La dénitrification en tant que processus écologique peut être perçue sous différents angles. Elle peut être vue comme un « service écologique » puisque en transférant une partie de l’azote des sols et des eaux polluées dans l’atmosphère sans les faire transiter par le continuum aquatique rivière/océans (GALLOWAY et al. 2003),
elle épure les eaux. Par là, elle contribue à sauvegarder les écosystèmes aquatiques et à préserver la santé humaine des effets délétères des nitrates. Car les nitrates sont la principale cause d’eutrophisation dans les eaux continentales et marines (MCISAAC et
al. 2001 ; NIXON 1995) et leur toxicité aiguë et chronique justifie qu’en Europe les
doses maximales autorisées pour déclarer une eau potable soient de 50 mg L-1.
Le processus de dénitrification n’est cependant pas qu’un service écologique et peut aussi être considéré sous un angle négatif. Ainsi, outre le fait que la dénitrification est susceptible de réduire la production des cultures en dissipant les amendements azotés (MOOGE et al. 1999), celle-ci est la principale source de N2O,
un gaz dont l’effet serre est 300 fois plus important que celui du CO2 et qui participe
à la destruction de l’ozone. 25% du N2O produits par dénitrification restent dans la
troposphère pendant plus de 100 ans (GALLOWAY et al. 2003). Les milieux
océaniques sont les principaux émetteurs de N2O mais d’autres milieux aquatiques
(rivières, estuaires, plateaux continentaux) représentent une part non négligeable (35% soit 1,9 Tg N a-1) de cette émission (SEITZINGER et al. 2000). Dans les milieux
terrestres, les sols fertilisés sont aussi une source importante de N2O (jusqu'à 25% de
l’émission totale selon le PRIP - Prévention et Réduction Intégrées de la Pollution (1994).
3.2. Les bactéries dénitrifiantes
La dénitrification est un trait biologique assez répandu chez les procaryotes. Il a été identifié dans des groupes phylogénétiquement éloignés comme les Aquificae,
Deinococcus-Thermus, Firmicutes, Actinobacteria, Bacteroides et Proteobacteria
(ZUMFT 1997). En conséquence, une simple affiliation taxonomique ne suffit pas à
dire si telle bactérie est dénitrifiante ou non.
De plus, parmi les bactéries dénitrifiantes, tous les genres ne sont pas capables de réaliser l’ensemble des séquences de la dénitrification (TIEDJE 1994) ;
(PHILIPPOT et al. 2007). Certaines bactéries non dénitrifiantes (les bactéries nitrate
réductrices) partagent avec les dénitrifiantes la capacité à transformer les nitrates en nitrites, l’une des étapes de la dénitrification, mais ne peuvent réaliser le processus dans sa totalité. Des bactéries impliquées dans la dénitrification participent aussi à d’autres processus comme la nitrification ou la fixation de l’azote (PHILIPPOT et al.
2007). Les bactéries dénitrifiantes peuvent être organotrophes, lithotrophes, ou même phototrophes, suivant la nature de la source d’énergie utilisée. Dans le cas présent, nous nous intéressons à la dénitrification effectuée par une grande partie des bactéries anaérobies facultatives chimio-hétérotrophes.
Une des premières études sur la phylogénie des communautés dénitrifiantes a été réalisée par (BETLACH 1982) qui s’est intéressé à la présence des différents gènes
codant pour les enzymes de la dénitrification chez les procaryotes. Il suggére que les dénitrifiantes auraient un ancêtre commun, une bactérie pourpre photosynthétique. Aujourd’hui, la diversité dénitrifiante est déterminée à l’aide de marqueurs
nombre de genres bactériens dénitrifiants (PHILIPPOT et HOJBERG 1999). Dans le
génome bactérien, ce sont les seuls gènes de dénitrification qui peuvent être représenté par plus d’une copie (PHILIPPOT 2002). Ces gènes ont été décrits dans les
bactéries α-, β-, γ- et ε-proteobacteria ; les dénitrifiantes narG sont incapable d’assurer une autre étape de la dénitrification (CHENEBY et al. 2003).
Les gènes nirS et nirK codent pour les cd1- et Cu-nitrite réductases qui
participent à la transformation des nitrites en monoxyde d’azote (PHILIPPOT 2002).
La présence de monoxyde d’azote et de concentrations basses en oxygène stimule l’expression de nirS mais cette expression ne requière pas une anaérobiose complète, contrairement au gène nirK (PHILIPPOT et al. 2001). Ces gènes ont été les plus
utilisés pour la caractérisation des communautés dénitrifiantes dans les milieux terrestres et aquatiques (BRAKER et al. 2000 ; PRIEME et al. 2002 ; YOSHIE et al.
2004). Parmi les gènes les moins utilisés pour l’étude des communautés dénitrifiantes, norB et qnorB codent pour les sous-unités catalytiques des deux réductases qui participent à la transformation du monoxyde d’azote en protoxyde d’azote, une réaction biologique particulière à cause de la création de liaison N-N.
(PHILIPPOT 2002). Comme pour le gène nirS, la présence de monoxyde d’azote
favorise l’expression de ces gènes. Peu d’auteurs utilisent ces gènes pour étudier la diversité dénitrifiante (BRAKER et TIEDJE 2003 ; DANDIE et al. 2006). Enfin, parmi
les gènes qui codent pour la protoxyde d’azote réductase, un seul gène est identifié comme marqueur fonctionnel. Cette étape de la dénitrification est la seule à avoir cette caractéristique. La phylogénie des gènes nosZ est congruente avec les arbres phylogénétiques 16S ARNr, sauf quelques exceptions expliquées par des transferts latéraux (ZUMFT et KÖRNER 2007).
Le développement d’amorces adaptées à chaque gène (BRAKER et al. 1998 ;
HALLIN et LINDGREN 1999 ; BRAKER et TIEDJE 2003 ; THROBACK et al. 2004) a aidé
2004 ; HENRY et al. 2004 ; RICH et MYROLD 2004) ou de deux gènes qui codent une
même étape i.e. nirS et nirK (BRAKER et al. 2000 ; PRIEME et al. 2002) ; HEYLEN et
al. 2006). Seuls quelques récents travaux font exception (HENRY et al. 2006 ;
KANDELER et al. 2006) en intégrant à leur analyse les communautés dénitrifiantes
décrites avec chacun des gènes impliqués dans ces différentes étapes de la dénitrification.
3.3. Justification du choix du modèle
: les
communautés dénitrifiantes dans le biofilm hétérotrophe
d’un aquifère alluvial
3.3.1.
Les
aquifères alluviaux et leur
fonctionnement
Les zones riveraines constituent des zones de transition et de régulation des flux de matière et d’énergie entre le cours d’eau et les zones adjacentes, entre les eaux de surface et les eaux souterraines (PETERJOHN et CORRELL 1984). L’aquifère
alluvial est l’un des compartiments de la zone riveraine. Il est constitué par des dépôts sédimentaires perméables de type gravier et sable, dont la porosité est saturée d’une eau qui peut provenir soit de la nappe phréatique soit de la rivière (CREUZE DES
CHATELLIERS et al. 1994). Pour un instant donné, chaque point de l’aquifère est donc
caractérisé par une valeur de mélange de ces eaux d’origines différentes. Dans le dépôt alluvial, les interactions entre les eaux provenant de la surface et les eaux d’origine phréatique favorisent des processus biotiques qui régulent les stocks en azote, carbone et phosphore, eux même conditionnés par l’hydrologie du système
sur un pourcentage en eau de surface hyporhéiques qui doit être supérieur à 98%. Dans notre cas, ce pourcentage variant d’un piézomètre à l’autre, nous avons préféré utiliser le terme d’aquifère ou de zone alluviale.
Au sein d’une zone alluviale, la connectivité locale et instantanée varie. Cette connectivité peut être appréciée à travers l’amplitude et la dynamique des variations de niveau de nappe (STANDFORD et WARD 1993). Ainsi les zones fortement
connectées à la rivière ont des hiérogramme de profils similaires à celui de la rivière ;
a contrario, dans les zones le moins connectées le niveau de nappe ne varie pas ou la
variation est décalée dans le temps (WENG et al. 2003). Ainsi les zones alluviales
sont une composante du continuum hydrologique. Les conditions environnementales qui règnent au sein d’un aquifère alluvial sont plus stables que celles qui caractérisent la rivière. Les eaux qui y circulent sont peu turbulentes, ont des temps de résidence longs (la vitesse moyenne de l’eau varie entre 0,3 et 1 mètre par jour dans l’aquifère étudié (WENG et al. 2003), les sédiments y ont une structure stable, la
température y est peu variable et l’obscurité permanente (BRUNKE et GONSER 1997).
Dans ces zones de transition, la qualité de l’eau est fortement influencée par l’activité microbiologique (PUSCH et al. 1998).
3.3.2.
Les
rôle
des communautés bactériennes
dans les aquifères alluviaux
Au sein des aquifères alluviaux, les bactéries sont impliquées dans la transformation de la matière organique et participent aux grands cycles géochimiques des nutriments, notamment celui de l’azote (PUSCH et al. 1998). Le carbone
provenant de la rivière contient une fraction importante de carbone labile qui soutient l’activité bactérienne (HENDRICKS 1996). Spatialement, les densités de ces bactéries
diminuent au fur et à mesure qu’augmente la distance avec la rivière (ELLIS et al.
1998 ; CRAFT et al. 2002). Toutefois, l’analyse des communautés bactériennes ainsi
récente par rapport aux communautés d’autres milieux, terrestres et aquatiques
(GHIORSE et WILSON 1988 ; CHAPELLE 1993 ; MADSEN et GHIORSE 1993). Plusieurs
études se sont intéressé à la différence entre communautés planctoniques et communautés attachées au sédiment, montrant que la majeure partie de la biomasse dans les aquifères vit attachée aux particules de sédiment (HIRSCH et RADES
-ROHKOHL 1990 ; ALFREIDER et al. 1997 ; RÖLING et al. 2001 ; GRIEBLER et al.
2002 ; BESEMER et al. 2005), comme elles vivent attachées à des supports dans la
plupart des milieux naturels (WATNICK et KOLTER 2000). Une étude récente a porté
sur la composition des deux types de communauté (attachée et libre) et les facteurs qui influencent leur structuration dans un plaine alluviale : (BESEMER et al. 2005) qui
montraient l’importance de l’hydrologie et de l’interaction eau de rivière/eau souterraine sur la régulation de la composition des communautés bactériennes. Cependant, les connaissances sur leur composition, les facteurs qui influencent leur composition et leur rôle fonctionnel restent peu précisées. L’accès difficile au milieu souterrain pose un problème lors de l’échantillonnage et a sévèrement limité l’avancement des travaux dans ce milieu (MADSEN et GHIORSE 1993). Plusieurs
auteurs ont contourné cet obstacle en introduisant des échantillons de milieux stérilisé ou des substrats artificiels comme de billes en verre (HIRSCH et RADES
-ROHKOHL 1990 ; DODDS et al. 1996 ; CLARET 1998 ; GRIEBLER et al. 2001).
3.3.3.
Justification
du
modèle
Pour ce travail de thèse, nous avons réalisé des expérimentations in situ avec des agrégats microbiens fixés (biofilm) afin de déterminer le rôle fonctionnel des communautés bactériennes dénitrifiantes. Les biofilms, sont distribués de façon