© Julius Baya Mdzomba, 2021
The role of Nogo-A in the visual deficits induced by
retinal injury
Thèse
Julius Baya Mdzomba
Doctorat en neurobiologie
Philosophiæ doctor (Ph. D.)
The role of Nogo-A in the visual deficits
induced by retinal injury
Thesis
Julius Baya Mdzomba
Under the supervision of:
iii
Résumé
Le manque de thérapies efficaces pour les pathologies oculaires affectant les neurones rétiniens conduit suivant à un déclin de la vision et même à la cécité. Par exemple, le glaucome, la rétinopathie diabétique et l'ischémie rétinienne entraînent la perte des cellules ganglionnaires rétiniennes, les seules cellules transmettant des informations visuelles de l'œil au cerveau. Cette perte est aggravée par le fait que le système nerveux central mature (SNC), dont fait partie la rétine, a des capacités très limitées à se régénérer après une blessure. Le principal mécanisme par lequel ces neurones sont perdus, dans la plupart de ces pathologies, est l'excitotoxicité due à la sur-activation du récepteur N-méthyl-D-aspartate (NMDAR). En effet, la sur-stimulation du récepteur conduit à un afflux massif d'ions calcium dans le neurone, activant les voies apoptotiques et nécrotiques et finalement la mort cellulaire. Nogo-A est un puissant inhibiteur de la croissance neuritique et est depuis longtemps étudié dans les maladies neurodégénératives et dans les tentatives de régénération après des pathologies telles que les lésions de la moelle épinière, la sclérose en plaques et les accidents vasculaires cérébraux (AVC). Il a également été impliqué dans la maladie d'Alzheimer et même la maladie de Parkinson en raison de son lien avec les protéines associées à ces maladies. Dans les études sur les lésions de la moelle épinière chez le rat, la neutralisation de Nogo-A à l'aide d'anticorps bloquant pouvait aider à la récupération des fonctions motrices perdues. Actuellement, chez les humains souffrant des lésions de la moelle épinière, des études cliniques de deuxième phase sont en cours d’investigation. De même, des études prouvent que la neutralisation de Nogo-A peut également être utilisée comme thérapie après un AVC. Ces données montrent que la neutralisation de Nogo-A améliore la vascularisation autour du site des infarctus, réduit la gravité de la blessure et contribue ainsi à la récupération fonctionnelle après un AVC ischémique.
Il y a eu peu d'études sur les implications de Nogo-A dans les pathologies du système visuel, en particulier celles qui affectent la rétine. Dans ma thèse, nous proposons que Nogo-A soit impliquée dans la physiopathologie des maladies oculaires et nous avons étudié le rôle que Nogo-A jouait dans le système visuel et après une lésion rétinienne. Dans cette thèse, nous montrons que l'expression de Nogo-A n'est pas seulement
iv
augmenter dans la vitré des souris après une lésion rétinienne, mais que cette augmentation est corrélée à la quantité de dommages à la rétine. En outre, la neutralisation de Nogo-A conduit à une récupération spontanée des fonctions visuelles qui était plus rapide et complète par rapport aux animaux de contrôle (WT) après une lésion rétinienne. De plus, l'activité des cellules ganglionnaires rétiniennes (RGC) vers le cerveau est améliorée par rapport aux animaux WT après une lésion rétinienne. L'augmentation de l'acuité visuelle des yeux intacts des souris KO pour Nogo-A, témoigne de la présence de la plasticité corticale accrue. Remarquablement, le traitement avec l'anticorps a montré des résultats similaires dans la récupération de la fonction visuelle et la propagation du signal vers le cerveau et ces améliorations se sont maintenues sur une période de plusieurs semaines. Nous montrons également que Nogo-A affecte la réaction inflammatoire de la rétine après une blessure. Cette inflammation pourrait être impliquée dans l'exacerbation de la blessure. Le blocage de Nogo-A conduit à une régulation négative des molécules inflammatoires, dont le TNFα. Le niveau de TNFα est resté basse pendant plusieurs jours après la neutralisation de Nogo-A et pourrait être impliquée dans la récupération fonctionnelle. De plus, nous montrons également que l'expression de Nogo-A est fortement régulée à la hausse dans le tissu rétinien humain de donneurs souffrant de rétinopathie diabétique.
Nos résultats montrent donc que Nogo-A est impliqué dans la physiopathologie des maladies oculaires et que sa neutralisation, en utilisant ici la KO et l'anticorps bloquant, peut-être une nouvelle thérapie non seulement pour les maladies oculaires mais pourrait être pertinente dans le traitement des maladies qui présentent une augmentation de la réponse inflammatoire.
v
Abstract
The lack of effective therapies for ocular pathologies affecting retinal neurons lead most often to vision decline and even blindness. For example, glaucoma, diabetic retinopathy, and retinal ischemia, leads to the loss of retinal ganglion cells, the only cells relaying visual information from the eye to the brain. This loss is compounded by the fact that the mature central nervous system (CNS), of which the retina is part of, has very limited abilities to regenerate after injury, leading to irreversible blindness. The main mechanism by which these neurons are lost, in most of these pathologies, is excitotoxicity due to the over activation of N-methyl-D-aspartate receptor (NMDAR). Indeed, the over stimulation of the receptor leads to a massive influx of calcium ions into the neuron, activating apoptotic and necrotic pathways and ultimately cell death. Nogo-A is a potent neurite outgrowth inhibitor that has been studied for a long time in neurodegenerative diseases and in regenerative efforts after pathologies like spinal cord injuries, multiple sclerosis and stroke. It has also been implicated in Alzheimer’s disease and even Parkinson’s disease due to its link to the proteins associated with these diseases. In the spinal cord injury studies in rats, Nogo-A neutralization using function blocking antibodies has been shown to rescue some motor function. Currently, second stage clinical studies are investigated in humans suffering from spinal cord injuries. Likewise, there is growing evidence that Nogo-A neutralization can also be used as a therapy after stroke. This evidence shows that Nogo-A neutralization improves vascularization around the infarcts site, reduces the severity of the injury and thus helps in function recovery after an ischemic stroke.
There has been little study of the implications of Nogo-A in pathologies of the visual system especially those that affect the retina. In my thesis we hypothesize that Nogo-A is involved in ocular disease pathophysiology and we reveal this by investigating the role that Nogo-A plays in the visual system and after retinal injury. In this thesis, we show that Nogo-A expression is not only upregulated after retinal injury, but this upregulation is correlated to the amount of damage to the retina. Furthermore, neutralization of Nogo-A in knockout (KO) mice led to spontaneous recovery of visual functions that was faster and complete as compared to wildtype (WT) animals after retinal
vi
injury. Moreover, retinal ganglion cells (RGCs) activity to the brain was improved when compared to WT animals after retinal injury. Cortical plasticity was also observed to be heightened in the Nogo-A KO mice as compared to WT animals as seen by the increase in the visual acuity of intact eyes of these mice. Remarkably, treatment with Nogo-A function-blocking antibody showed similar results in both visual function recovery and signal propagation to the brain. These improvements were sustained over a period of several weeks. We also show that Nogo-A affects the inflammatory reaction of the retina after injury, which could be implicated in the exacerbation of the injury. Blocking Nogo-A leads to a down-regulation of inflammatory molecules including, TNFα, which was sustained over several days, and could help in function recovery. We also show that Nogo-A expression is highly upregulated in human retinal tissue from donors suffering from diabetic retinopathy.
Overall, our results show that Nogo-A is implicated in the pathophysiology of ocular diseases and that its neutralization, here using KO and function blocking antibody, can be a novel therapy not only for ocular diseases but could be relevant in the treatment of diseases that exhibit increase in inflammatory response.
vii
Contents
Résumé ... iii
Abstract ... v
List of Figures ... xi
List of Tables ... xiii
List of abbreviations ... xiv
Acknowledgments ... xix
Avant-propos ... xxii
INTRODUCTION ... 1
1. The visual system ... 1
1.1. Anatomy of the visual system, difference between human and mice ... 1
1.1.1. Retina: cytoarchitecture and expression of Nogo-A in the different cell types. ... 1
1.2. Brain visual system: Optic nerve, LGN, superior colliculus and visual cortex... 4
1.2.1. Optic nerve ... 4
1.2.2. Dorsal lateral geniculate nucleus ... 5
1.2.3. Superior colliculus ... 8
1.2.4. Visual cortex ... 9
2. Reticulon family tree in mammals, characteristics, features, and function of the family ... 11
Reticulon 1 (RTN-1) ... 12
Reticulon 2 (RTN-2) ... 13
Reticulon 3 (RTN-3) ... 13
The reticulon 4 (Nogo-A) ... 14
3. Nogo-A ... 15
3.1 Nogo-A Receptors and signaling pathway ... 16
3.1.1. NGR1, PIRB, S1PRs ... 16
3.1.2. LINGO1, p75, TROY ... 16
3.2. The role of Nogo-A in in the development of the central nervous system in mammals ... 19
3.3. Nogo-A implications in neuronal functions ... 19
3.3.1. Neuronal cytoskeletal modelling ... 20
3.3.2. Neuronal gene and protein expression ... 20
3.3.3. Memory formation ... 21
3.3.4. Neuronal outgrowth and cell migration ... 22
3.3.5. Axon myelination ... 23
viii
4. Implications of Nogo-A in the maintenance of neuronal function in aging and after CNS
insult ... 24
4.1. The expression and role of Nogo-A in the aging cortex and after cortical insults and pathologies. ... 24
4.1.1. Alzheimer’s disease ... 24
4.1.2. Parkinson’s disease ... 25
4.1.3. Multiple sclerosis ... 26
4.1.4. Ischemic stroke ... 27
4.1.5. Spinal cord injury ... 27
4.1.6. Aging ... 28
4.3. The role of Nogo-A in the maturation of the visual pathway ... 29
4.4. Implication in visual maturation, segregation and retinotopic map ... 29
4.4.1. In the retina... 29 4.4.2. In the brain ... 30 5. Implication in inflammation ... 32 5.1. Neuro-inflammation ... 32 5.2. Immune cells ... 32 5.2.1 Macrophages ... 33 5.2.2 Microglia ... 34 5.3. Cytokines ... 35 5.3.1. Pro inflammatory ... 35
5.4. Implications of Nogo-A on inflammation ... 37
6. Ocular pathologies ... 38
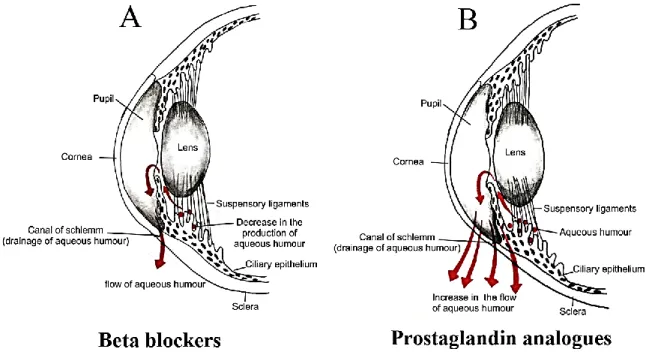
6.1. Glaucoma ... 38
6.2. Diabetic retinopathy ... 40
6.3. Retinal ischemia ... 42
OBJECTIVES AND HYPOTHESIS ... 43
Chapter 1: Nogo-A inactivation improves visual plasticity and recovery after retinal injury .... 45
1.1. Résumé ... 46
1.2. Abstract ... 47
1.3. Introduction ... 48
1.4. Materials and methods ... 50
Animals ... 50
Intraocular injections ... 50
ix
ERG recording ... 51
LFP recording in the visual cortex ... 52
RGC survival analysis on retinal flat-mounts ... 52
Retina processing and immunostaining ... 53
Semi-qRT-PCR ... 53
1.5. Results ... 55
Dose-dependent effects of NMDA on visual loss and recovery ... 55
Endogenous visual recovery after retinal damage ... 55
Nogo-A is not significantly downregulated in retinal glia after reversible injury... 56
Nogo-A neutralization improves visual recovery and plasticity after retinal injury ... 57
The electrophysiological response of V1 neurons is modified by Nogo-A inactivation... 58
1.6. Discussion ... 60
The intrinsic recovery properties of the injured mouse visual system ... 60
The involvement of Nogo-A signaling in the inhibition of retinal neuron recovery ... 61
Nogo-A inactivation enhances visual brain plasticity ... 62
The role of glial vs neuronal Nogo-A on vision recovery and plasticity ... 62
The modulation of neuronal plasticity in ocular disease treatments ... 63
1.7. Conclusion ... 64
Chapter 2: Nogo-A-targeting antibody promotes visual recovery and inhibits neuroinflammation after retinal injury. ... 80
2.1. Résumé ... 81
2.2. Abstract ... 82
2.3. Introduction ... 83
2.4. Materials and methods ... 85
Animals ... 85
Intraocular injections ... 85
Optokinetic reflex test ... 85
Electroretinogram (ERG) recording ... 86
Tissue preparation for histological analysis ... 86
Retinal ganglion cell (RGC) and amacrine cell survival ... 86
Immunofluorescence on retinal and optic nerve cryosections ... 87
Western blot analysis ... 87
Semi-qRT-PCR ... 88
Human retina histology and vitreous analyses ... 89
x
Retinal excitotoxicity increases the level of Nogo-A proteins in the vitreous of mice ... 90
Targeting Nogo-A-Delta 20 with a function-blocking antibody improves visual recovery after retinal injury ... 91
Nogo-A antibody inhibits monocyte-mediated inflammation in the injured retina ... 92
Nogo-A modulates phosphorylation signalings controlling retinal gliosis ... 93
Nogo-A is upregulated in the retina of patients with diabetic retinopathy ... 94
2.6. Discussion ... 95
Retinal excitotoxicity increases vitreal Nogo-A proteins ... 95
Intraocular inhibition of Nogo-A with blocking antibody promotes visual recovery ... 96
Switch in microglia and Müller glia reaction to injury ... 97
Therapeutic potential of Nogo-A-targeting antibodies in the treatment of retinal diseases ... 99
2.7. Conclusion ... 100
DISCUSSION AND PERSPECTIVES ... 119
1. Recapitulation on the results ... 119
2. Perspectives ... 122
2.1. Induced diabetic retinopathy ... 122
2.1.1. Streptozotocin (model for type I diabetes) ... 123
2.1.2. db/db mice (model for type II diabete) ... 123
2.1.3. Expected result ... 124
2.2. GLAST model of glaucoma ... 125
2.2.1. Expected results ... 126
GENERAL CONCLUSION ... 128
xi
List of Figures
Figure 1. Illustration of the reticulon Family tree. Figure 2. Illustration of Nogo-A signaling in Neurons. Figure 3. Illustration of the eye (a) and retina (b).
Figure 4. Illustration of the retinofugal projections of the mouse and human. Figure 5. Illustration of the mouse and human dLGN..
Figure 6. Illustration of the superior colliculus. Figure 7. Illustration of the visual cortex.
Figure 8. Illustrations of topical treatment to lower intraocular pressure. Figure 9. Schematic illustration of retinal changes due to diabetic retinopathy. Figure 10. Reversible and irreversible patterns of OKR deficits are induced in a dose-dependent manner by intravitreal injections of NMDA.
Figure 11. Marked electroretinographic alterations and histological damages are associated with irreversible vision deficits.
Figure 12. NMDA-induced gene expression changes in the retina.
Figure 13. Nogo-A gene ablation improves visual recovery and plasticity after NMDA-induced excitotoxicity.
Figure 14. The electrophysiological response of V1 neurons is modified in the absence of Nogo-A.
Figure 15. Intravitreal injection of Nogo-A-neutralizing antibody 11C7 changes V1 neuron response after retinal injury.
Figure 16. RGC survival quantification on retinal flat-mounts. Figure 17. Inflammation gene expression after intravitreal injection.
Figure 18. Comparison of LFP wave amplitudes between WT and Nogo-A KO mice and after antibody delivery.
Figure 19. Retinal function and RGC survival analyses after intravitreal injection of NMDA and antibodies.
Figure 20. Retinal injury induces Nogo-A protein release in the mouse vitreous. Figure 21. Intravitreal administration of Nogo-A blocking antibody enhances visual recovery after retinal injury.
xii
Figure 22. Antibody-based neutralization of Nogo-A mitigates inflammation in the injured retina.
Figure 23. Antibody-mediated Nogo-A blockade decreases the number of cells expressing TNFα without affecting the number of Iba1-positive monocytes.
Figure 24. 11C7 treatment inhibits inflammatory processes in injured retinal glia. Figure 25. Nogo-A protein is increased in the vitreous and in the retina of patients affected by diabetic retinopathy.
Figure 26. Electroretinogram (ERG) recordings.
Figure 27. Histological analysis of oligodendrocytes in the optic nerve. Figure 28. Histological analysis of RGC axons in the optic nerve.
xiii
List of Tables
Table 1. Primer sequences used for qRT-PCR measurements in retinal lysates. Table 2. Antibodies used for immunofluorescence (IF) and western blotting (WB). Table 3. Primer sequences used for qRT-PCR measurements.
xiv
List of abbreviations
aa Amino acid
AD Alzheimer’s disease
ANOVA Analysis of variance
AP Adaptor-related protein
APP Amyloid β peptides
BACE1 BBB BDNF
Beta-site amyloid precursor protein cleaving enzyme 1 Blood brain barrier
Brain derived neurotrophic factor cAMP Cyclic adenosine monophosphate ChAT Choline acetyltransferase
CME Cystoid macular edema
CNS Central nervous system
CREB cAMP response element binding protein CRMP-2 Collapsin response mediator protein-2 CSPG Chondroitin sulfate proteoglycan C-terminus
DAMPS
Carboxyl-terminal
Damage associated molecular patterns dLGN Dorsal lateral geniculate nucleus
DR Diabetic retinopathy
DSGC Direction selective ganglion cells EAAC1 Excitatory Amino Acid Transporters 3
ER endoplasmic reticulum
ERG Electroretinogram
Erk1/2 Extracellular signal-regulated kinase 1/2
GABA Gamma aminobutyric acid
Gap43 Growth associated protein 43
GAPDH D-glycéraldéhyde-3-phosphate déshydrogénase
GCL Ganglion cell layer
GFAP Glial fibrillary acidic protein
GIRK G protein-gated inwardly rectifying K+ channels GLAST Glutamate/aspartate transporter
GS Glutamine synthetase
GTP Guanosine triphosphate
HEK 293 Human embryonic kidney 293 HSPG Heparan sulfate proteoglycans
IgG Immunoglobulin G INL IFN-γ IL-1β IL-4 IL-6 IL-10 IL-13
Inner nuclear layer Interferon‐gamma Interleukin 1β Interleukin 4 Interleukin 6 Interleukin 10 Interleukin 13
xv
ipRGCs JAK
Intrinsically photosensitive melanopsin-containing retinal ganglion cells
Janus kinase
KO Knock-out
LFP Local field potentials LINGO1
LIF
Leucine rich repeat and Immunoglobin-like domain-containing protein 1
Leukemia inhibitory factor
LTD Long-term depression
LTP MAPK
Long-term potentiation
Mitogen-activated protein kinase
MD Monocular deprivation
MS Multiple sclerosis
mTOR MYD88
mammalian target of rapamycin
Myeloid differentiation primary response protein 88 NFL
NF-κB
Nerve fiber layer Nuclear factor kappa B
NgR1 Nogo Receptors 1
NMDAR N-methyl-D-aspartate receptor N-terminus Amino terminal
OC Optic chiasma
OD Ocular dominance
OKR PAMPs
Optokinetic response
Pathogen associated molecular patterns
P.Stat3 Phosphorylated Signal transducer and activator of transcription 3 PBS Phosphate-buffered solution
PD Parkinson’s disease
PirB PRR
Paired immunoglobulin-like receptor B Pattern recognition receptors
PSD95 Postsynaptic density protein 95
PV+ Parvalbumin positive
qRT-PCR Quantitative real time reverse transcription polymerase chain reaction
RBPMS RNA-binding protein with multiple splicing RGC retinal ganglion cells
RHD Reticulon Homology Domain
RhoA Ras homolog family member A
RhoGEF Rho guanine exchange factor
RNA Ribonucleic acid
ROCK Rho-associated coiled-coil containing protein kinase
RTN Reticulon
S1PR2 Sphingolipid 1 phosphate receptors 2 SALMs Synaptic adhesion-like molecules SC
SOCS3
Superior colliculus
Suppressor of cytokine signalling 3 Sprr1a
STAT-1
Small proline rich protein 1a
xvi
STAT-3 TGF-β TIR TLRs
Signal transducer and activator of transcription 3 Transforming growth factor
Toll/IL-1 receptor Toll like receptors
TNFα Tumor necrosis factor alpha
TROY Tumor necrosis factor-α (TNFα) receptor superfamily member 19
Tubb3 β3tubulin
V1 visual cortex
WT Wildtype
αRGC Alpha retinal ganglion cells
xvii
Dedication
xviii
Epigraph
Lamukani asena hukashomeni, Lamukani asena fukashomeni, Hindeni hosini asena,
Hindeni ache na alume, Hindeni hosini fukapate elimu.
xix
Acknowledgments
This journey, started four years ago and coming to an end now, has been the effort of many people. Blood, sweet, and tears have been shaded in the realization of this manuscript. I hope I do not forget anyone.
First of all, I would like to thank Dr Pernet for the opportunity he gave me to learn all that I have needed to realize this thesis. The scientific ethics and culture that make a great scientist. The camaraderie of the team both in the realization of this and many other projects in the team. The patience of teaching and supervising someone like me. It takes an exceptional person to do all that. Thank you, sir.
Even Dr Pernet could not do well all that he does without the help of Dr Sandrine Joly. Your help in virtually every aspect of this thesis is paramount in its accomplishment. From the very first day finding me by the coded door, to the lessons of immunofluorescence and western blotting. I would like to visit Switzerland one day and from your stories of the place, I would like it.
I have always been fascinated by electrophysiology and getting to use that technique in my project has been one of the best experimental time pasted in the lab. This could not have been possible without the help from Dr Frederic Bretzner who not only allowed me to use his set-up but helped me in the setting up of the experiments. Thank you very much sir.
I also want to thank Dr. M. E. Schwab for providing us with anti-Nogo-A antibodies and Nogo-A KO mice, Dr. E. Planel, Prof. Serge Rivest and Dr. S. Proulx for allowing us to use their epifluorescence and confocal microscopes and Dr. S. Hébert for sharing his qRT-PCR machine with us. Also Dr. Ali Dirani, Dr. Patricia Lassiaz and Dr. Francine Behar-Cohen for the collaboration in this project and the human data we used.
All this work needed financial support and I would like to thank the following organization for their confidence and support. The Fonds de recherche du Québec-Santé (FRQS), The Canadian Research Health Institutes (CIHR), The Vision Health Research Network (VHRN) The Fonds de recherche du Québec-Santé, the Canada Foundation for
xx
Innovation (CFI), The Velux Stiftung, the foundation CHU de Quebec, The Neurosciences axis CRCHUQ-UL, the Centre Thématique de la Recherche en Neurosciences (CTRN) the faculty of medicine Université Laval and department ophthalmology.
I would like to thank the other members of team Pernet, Sabrina, Emilie and former members Mahmoud. You guys have been the best colleagues in this endeavor for it is with good humor and comraderies like yours that I have had the best time to achieve this.
My sister from another mister. Petite crevette. You know all that has gone into the accomplishment of this project. From Québec to Hawaii, to being bitten by mosquitos in the forests during camping. You have been the voice of urgency and the close help in everything about this project and more. Thanks for all of it.
A big thanks also goes to Dr. Emmanuelle Boucher for her cheerful demeanor, a great friend and an expert in her field. Some say cooking is like carrying out an experiment but for the life of me, I have not understood why you excel in one and are terrible in the other. It is not for the lack of protocols.
Those friends that crossed my path, in the lab, on the football pitch, in the swimming pool, you are too many to put your names down but here goes. Maude, Marie-Kim, Fernando, Mickael, Andreanne, Chloé, Marie, Remi, Serena, Ophélie, Nicolas, Melody, Mathilde, Antho, Julia, Mémé, Quentin, Ben, Hortense, Robert, Sy, Cedric. Pap, I forget a lot more but you guys rock.
A special thanks to my Bru. Those encouraging words, prayers and company you have given me in the past two years nearly. I can’t thank you enough for the PhD memes and those lessons of how to take a great selfie. You know what has been sacrificed in the realization of this project.
The Sirya family, for believing that I had what it took to do this. Baba Sirya, pembe za khulungu thizino. Mama Sirya, nothing that I have been able to do ever would not have happened without you. Jerry, Laure, Jean and Gabriel you guys have seen the
xxi
start of this journey and have been there for me, love you guys. James, Pricilla and Matsezi you guys are the best. Thii for being there with me when I was down and out, hot oil and flour season. Johnny and Hariri for all the times you stood by me when I was down and helped me out of those difficult moment.
A big thanks goes to the animal facility staff, France, Sonia, Steve, Kim, Mathieu, Karine, René and Denis.
I have never lived with housemates until I come to Quebec and boy did I get the best. Guylaine, Réal and even Charlie, thank you for the cozy and friendly place we have.
And Finally, I thank God for all He has enabled me to achieve, His revealing of the secrets of His creation and all the big and small miracles that have been rampant during this journey.
xxii
Avant-propos
Brief introduction and contributions towards this thesis.
The breakdown of this thesis is conventional to the norms of Université Laval. The introduction part is divided into three parts, a summary of the family tree of our protein of interest, Nogo-A, and the function of each member and isoform. Followed by a more detailed examination of Nogo-A and its implications in different aspect, from maturation to its involvement in several diseases of the central nervous system. Lastly, a brief description of the mouse and human visual system in the context of Nogo-A influence, molecularly, structurally, and functionally.
The articles that have been included in this thesis have both already been published in Cell Death and Disease Journal in 2018 and 2020, respectively. They have been a collaboration between our team and other both locally and internationally. Other articles that I have taken part in their realization in the team and are not included in this thesis are listed below.
Chapter 1: Nogo-A inactivation improves visual plasticity and recovery after retinal injury
Mdzomba J. B.*, Jordi N.*, Rodriguez L., Joly S., Bretzner F. and Pernet P.
Cell Death and Disease (2018) 9:727.
The initial planning and set-up of the project was done by Dr Pernet and subsequently we refined the plans. Initial observations of visual acuity and NMDA induced excitotoxicity test had been done in Zurich by my co-author, Noemie Jordi. I participated in all the experiments that took place in Québec. I participated in tissue collection and carried out all the immunofluorescent marking and data collection. With the help of Dr Bretzner and Dr Pernet, I set up the protocol and carried out all the visual evoked potential recordings that were done. In collaboration with Dr Pernet and Dr Joly, I interpreted the data and took part in the marking of some of the figures. Dr Pernet wrote the article and together with the other members of the team, I took part in the correction and layout of the article.
xxiii
Chapter 2: Nogo-A-targeting antibody promotes visual recovery and inhibits neuroinflammation after retinal injury.
Mdzomba J. B., Joly S., Rodriguez L., Dirani A., Lassiaz P., Behar-Cohen F. and Pernet V.
Cell Death and Disease (2020) 11:101.
The initial planning and set-up of the project was done by Dr Pernet, Dr Joly and me. I participated in all the experiments that took place in Québec. I participated in tissue collection and carried out some of the immunofluorescent marking and data collection for all mouse tissue. I carried out all the electroretinogram and visual acuity recordings that were done. In collaboration with Dr Pernet and Dr Joly, I interpreted the data and took part in the marking of some of the figures. Dr Pernet wrote the article and together with the other members of the team, I took part in the correction and layout of the article.
Other published or submitted articles that are not included in this thesis.
Human Tau expression does not induce mouse retina neurodegeneration, suggesting differential toxicity of Tau in brain vs. retinal neurons.
Rodriguez L., Mdzomba J. B., Joly S., Boudreau-Laprise M., Planel E., Pernet V.
Front Mol Neurosci. 2018; 11: 293.
Tau gene deletion does not influence axonal regeneration and retinal neuron survival in the injured mouse visual system.
Rodriguez L., Joly S., Mdzomba J.B., Pernet V.
Int. J. Mol. Sci. 2020, 21(11), 4100.
Tau modulates visual plasticity in adult and aging mice.
Rodriguez L., Joly S., Zine-Eddine F., Mdzomba J.B., Pernet V.
1
INTRODUCTION
1. The visual system
1.1. Anatomy of the visual system, difference between human and mice
1.1.1. Retina: cytoarchitecture and expression of Nogo-A in the
different cell types.
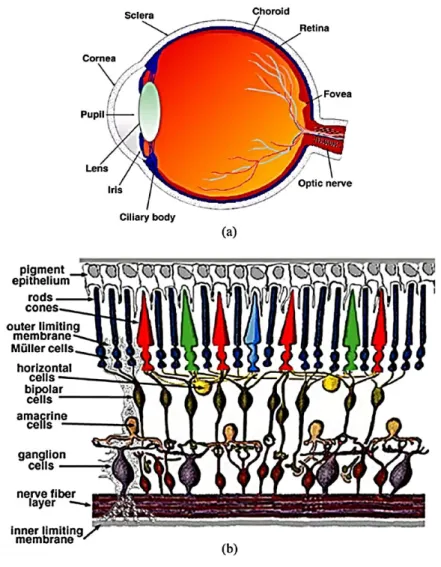
The retina is part of the central nervous system that is responsible for the collection processing and propagation of visual information. It is composed of several specialized cells, from neurons, interneurons and glial cells, organized in functionally optimized layers (Hoon
et al., 2014). There are about six major layers; photoreceptor layer, outer plexiform layer,
inner nuclear layer, inner plexiform layer, the ganglion cell layer, and the nerve fiber layer (Jeon, Strettoi and Masland, 1998). Photoreceptors, are composed of neurons that can convert light stimuli to electrical signals through a process called phototransduction. They line the back of the retina in the photoreceptor layer. These neurons are divided into two types, rod and cone photoreceptors, because of their form (Fig.3). Rod photoreceptors also differ from cone photoreceptors by their sensibility to light intensity, rods are more sensitive to light and can be activated at lower light intensity and shorter wavelengths, scotopic vision, while cones are less sensitive to light and are activated by high light intensity and longer wavelengths, photopic vision (Hoon et al., 2014; Lamb, 2016). In humans, there are three types of cone photoreceptors each more sensitive to a specific wave length of light conferring the ability to see in color (Hoon et al., 2014; Lamb, 2016)(Fig.3b). These cones photoreceptors make up only 5% of all the photoreceptors while the remaining 95% is made up of rod photoreceptors (Lamb, 2016). In mice, there are two types of cone photoreceptors, one expressing a short wavelength sensitive opsin, known as the S opsin, and the other expressing both the S opsin and a middle wavelength sensitive opsin, known as the M opsin. 95% of all murin photoreceptors express both M and S opsins (Hoon et al., 2014).
Downstream of the photoreceptors are the bipolar interneurons (Fig.3). They are found in the inner nuclear layer and form synaptic connections with photoreceptors and horizontal interneurons in outer plexiform layer. The synaptic connection formed by bipolar cells to the photoreceptors in their vicinity, are modulated by the horizontal interneurons
2
(Jeon, Strettoi and Masland, 1998). There are in general two types of bipolar cells, one connected to the rod photoreceptors thus called rod bipolar cells and the other connected to the cone photoreceptors and thus called cone bipolar cells (Jeon, Strettoi and Masland, 1998; Hoon et al., 2014). Bipolar cells can further be divided into ON and OFF depending on if they depolarize or hyperpolarize at the increment of light intensity, respectively (Hoon et al., 2014). The processed electrical signals propagate down to the next layer of integration, the inner plexiform layer, where the bipolar cells form synaptic connections to the retinal ganglion cells (RGCs). In this layer, the amacrine interneurons modulate the synaptic activity of bipolar and the RGC synaptic connection (Hoon et al., 2014). All the retinal interneurons are found in the inner nuclear layer, apart from displaced amacrine cells that can be found in the ganglion cell layer (Fig.3).
Retinal ganglion cells are the only neurons that transmit visual signals to the brain and are located in the ganglion cell layer (Hoon et al., 2014). There exist several types of retinal ganglion cells classified in many categories ranging from morphology to their function (Jeon, Strettoi and Masland, 1998). One of the most studied types of RGCs are the direction selective ganglion cells (DSGC) who as their name suggests are activated by the direction of stimuli (Kerschensteiner and Guido, 2017). They are further divided into ON and ON-OFF DSGC according to their preference to light increment or light increment and decrement (Sanes and Masland, 2015). ON DSGCs activate preferentially to one of three directions that are in line with the orientation of the inner ear, while ON-OFF DSGCs activate to motion in the four cardinal directions (Kerschensteiner and Guido, 2017). A part from their preferred activation cues, these RGCs do not target the same nuclei in the brain (Kerschensteiner and Guido, 2017). The other major RGC type is the alpha RGC (αRGC) which is characterized as having a big soma and very wide dendritic arbor (Sanes and Masland, 2015). These αRGCs are also further divided into three types, sustained ON, sustained OFF, and transient OFF αRGCs according to their activation preference (Sanes and Masland, 2015). Another group of RGCs that is noteworthy are the intrinsically photosensitive melanopsin-containing retinal ganglion cells (ipRGCs). Thanks to their melanopsin content these RGCs are the other photo-sensitive cells in the retina. These RGCs are characterized by their large dendritic arborization and their photo-sensibility. They are also divided into five types M1 to M5 and of note is the M4 group which is also the sustained ON αRGC. They project to several nuclei
3
in the brain and are responsible for the circadian rhythm among other functions (Sanes and Masland, 2015).
The RGC axon fibers carpet the inner surface of the retina, the nerve fiber layer, and converge in the middle to form the optic nerve head, the most apical segment of the optic nerve (Jeon, Strettoi and Masland, 1998). Specialized glial cell known as the Müller cells span the whole retina from the photoreceptors to the nerve fiber layer where they form structures known as the end feet (Masland, 2011; Hoon et al., 2014).
Figure 1. Illustration of the human eye (a) and retina (b).
The photoreceptors are responsible for photo-transduction. Electrical impulses are then propagated inwards to the bipolar cells and finally the ganglion cells. Horizontal and amacrine cells modulate these impulses and help in the processing of the visual information. Axons from the ganglion cells form the optic nerve which sends the pre-processed information to cortical and sub-cortical targets. Modified from (MacGillivray et al., 2014).
4
1.2. Brain visual system: Optic nerve, LGN, superior colliculus and
visual cortex
1.2.1. Optic nerve
The optic nerve is a consolidation of RGC axons, the myelin sheath and the oligodendrocytes forming the sheath, astrocytes, and connective tissue. In humans, about 0.5 to 1.2 million RGC axons form the optic nerve (Mukherjee, El-Dairi and Bhatti, 2013). The oligodendrocytes start to form the myelin sheath around every axon of the RGCs after they pass the lamina cribrosa. This lamina cribrosa is a load bearing structure of the optic nerve whose function is both to provide structural support and nutrients to RGC axons leaving the retina (Downs and Girkin, 2017). At this structure and between the axon fascicules are located astrocytes that not only bundle together the RGC axons as they leave but also provide homeostatic support to them (Cooper, Collyer and Calkins, 2018). Passed the lamina cribosa, the optic nerve is covered with the meningeal sheaths, bathed in cerebral spinal fluid and irrigated by the ophthalmic artery as it continues towards the various cortical and sub-cortical targets of the retina (Cooper, Collyer and Calkins, 2018).
Another noteworthy structure of the optic nerve is the optic chiasma. This is the region of the optic nerve whereby projections form the two retinas cross over. This percentage of RGC axon decussation depends on the laterality of the eyes on the head of the animal, which in turn influences the size of the binocular visual field (Herrera and Garcia-Frigola, 2008). In species with more forward facing eyes like humans, the percentage of crossing axons is close to 50% of axons, while in more lateral facing eyes like in mice, about 95% cross over (Herrera and Garcia-Frigola, 2008)(Fig.4). The decussation at the optic chiasma contributes to the stereopsis and binocular vision (Ireland and Carter, 2020).
5
Figure 2. Illustration of the retinofugal projections of the mouse and human.
R (Retina) OC (Optic Chiasma, LGN (Lateral Geniculate Nucleus, SC (Superior Colliculus) VC (Visual Cortex). In animals with lateral facing eyes most of the retinal projections cross over at the optic chiasma as compared to animals with frontal facing eye that have approximately the same number that crosses over. The visual receives and sends inputs to both the LGN and SC.
1.2.2. Dorsal lateral geniculate nucleus
One of the major targets for RGC axons input is the dorsal lateral geniculate nucleus of the thalamus (dLGN). In rodents, about 40 % of RGC axons and in humans and non-human
6
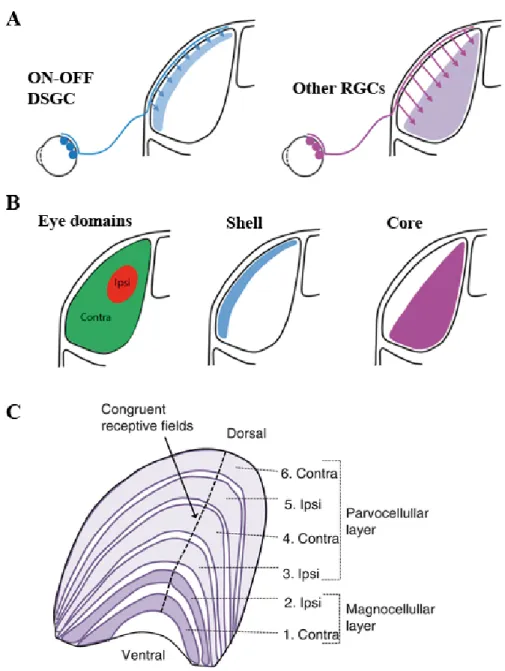
primates about 90% project to the dLGN, forming synaptic connections with relay neurons of the dLGN (Pasternak, Bisley and Calkins, 2003; Baker, 2012; Guido, 2018). These relay neurons in turn transmit the processed visual information to their target neurons in the visual cortex. These retinal inputs however, account for only ~ 10% of the inputs the dLGN receives. (Litvina, Chinfei Chen and Chen, 2017; Alitto et al., 2019). The remaining 90% of inputs come from other parts of the brain including the visual cortex and brainstem (Kerschensteiner and Guido, 2017). In mice ,the dLGN is segregated into two layers major layers, the ventricular margin of the dLGN also known as the shell, and the inner layer also called the core (Bickford et al., 2015; Seabrook et al., 2017) (Fig.5.A-B). Specific RGC inputs, for example ON-OFF DSGC, project to shell while other RGCs types i.e αRGCs project into the core (Kerschensteiner and Guido, 2017). In humans and non- human primates, the dLGN is arranged in an alternating layer of up to six laminae corresponding to inputs from the ipsilateral and contralateral retinae, termed magnocellular, parvocellular, and koniocellular layers (Rose and Bonhoeffer, 2018) (Fig.5C.). The LGN not only relays the incoming visual signals from the retina to the visual cortex and other nuclei but also modulates attention, facilitates or suppresses binocularity via monocular gain modulation and even selective sensitivity to stimulus orientation (Covington and Al Khalili, 2019).
7
Figure 3. Illustration of the mouse and human dLGN.
A, B. ON-OFF DSGC terminate in the shell of the dLGN while other RGCs terminate in the core of the dLGN in mice.
C. In Humans and non-human primates, inputs from the ipsi and contralateral retinas terminate in successive layers arranged in either parvocellular or magnocellular distinct layers according to their receptive field in the retina. Modified from (Kerschensteiner and Guido, 2017) and entokey.com.
8
1.2.3. Superior colliculus
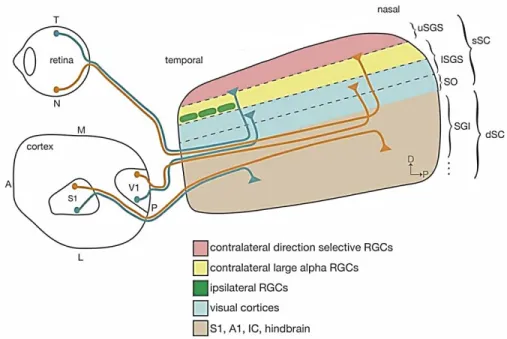
The other major nucleus that receives RGC inputs projections is the superior colliculus (SC) (Fig.4). It is a rostral midbrain integrative sensorimotor structure (one in each hemisphere) receiving inputs from several sensory sources and integrates them to elicit innate functions of an animal (Ito and Feldheim, 2018) (Fig.6.). These include, saccadic motion of the eyes which initiates gaze shifting, in turn eliciting the movement of the neck muscles and the turn of the head and in mice, suspension of locomotion and freeze or flee responses towards looming object (Ito and Feldheim, 2018; Zubricky and M Das, 2020). The SC receives inputs from both the ipsilateral and contralateral RGC projections, the bulk of which comes from the corresponding eye opposite to the position of the SC. in such a way that it retains the retinotopic map thanks to its layered structure that receive inputs from distinct RGC inputs. The most superficial layer is called the stratum griseum superficiale and receives direct RGC input projections from the contralateral retina terminating in different locations in this layer. The lower part of the stratum griseum superficiale receives inputs from the ipsilateral retina, contralateral αRGC from the contralateral eye and from the primary and extra-striate visual cortex (Ito and Feldheim, 2018) (Fig.6.). In mice, these RGC inputs can be about 85% to 90% of all RGC projections while in primates approximately 10% of RGC projections reach the SC pointing to the differing function or importance i.e. event detection in nocturnal animals (Hübener, 2003; Ito and Feldheim, 2018)
9
Figure 4. Illustration of the superior colliculus.
The superior colliculus receives direct retinal inputs, a majority from the contralateral retina to it. Retinal inputs terminate in the stratum griseum superficiale with different sources terminating in different location therein. As an integrative sensorimotor structure, it receives inputs from other sources including the sensory, and auditive cortex. A, anterior; P, posterior; T, temporal; N, nasal; S1, primary somatosensory cortex; V1, primary visual cortex; uSGS, upper stratum griseum superficial; lSGS, lower stratum griseum superficial; SO, stratum
opticum; SGI, stratum griseum intermedium. Modified from (Ito and Feldheim, 2018)
1.2.4. Visual cortex
The visual cortex is the principal cortical area of the brain that receives, integrates, and processes visual information coming from the retinae and relayed through the dLGN (Huff and Dulebohn, 2018)(Fig.4). In higher primates, the visual cortex is located in the occipital lobe of the primary cerebral cortex, the most posterior region of the brain and is divided into five distinct areas (V1-V5) (Huff and Dulebohn, 2018). The V1 also known as the primary visual cortex in both humans and mice, is the main input point of visual information coming from the dLGN and in turn sends its output to the other specialized visual regions and nuclei for further integration (Huff and Dulebohn, 2018; Young et al., 2019) (Fig.7). In mice, these higher visual areas are the lateromedial and anteromedial visual areas, and receive feed forward and giving feedback inputs to the V1 (Young et al., 2019). In both
10
species, the V1 comprises of six layers (layer 1-6) with each receiving distinct inputs in such a way that the V1 retains the contralateral retinotopic map (Hübener, 2003; Gao, DeAngelis and Burkhalter, 2010) (Fig.7.). Likewise, there is a distinct segregation of monocular and binocular regions of the V1. The main entry point for inputs into the V1 is the layer 4, which receives dLGN inputs with other layers receiving a smaller portion of the dLGN output, while feedback inputs from the layer 6 of the V1 project back to the dLGN (Kerschensteiner and Guido, 2017; Huff and Dulebohn, 2018).
Figure 5. Illustration of the visual cortex.
Input from cortical and subcortical areas and outputs to these areas and higher visual areas originate from distinct layers of the visual cortex. Inputs from the dLGN synapse in the layer IV and from layer VI eminates output that go back to the dLGN. Illustration modified from quora.com.
11
2. Reticulon family tree in mammals, characteristics, features, and
function of the family
The reticulons (RTN) are a highly conserved family of proteins that are expressed in all eukaryotes (Yang and Strittmatter, 2007; Sano, Bernardoni and Piacentini, 2012). They are associated with the plasma membrane as well as the endoplasmic reticulum (ER) and the Golgi apparatus. In mammals, this family is comprised of four major groups RTN, RTN-1, RTN-2, RTN-3 and RTN-4/ Nogo, which due to alternative splicing are also further divided into several isoforms (Oertle, Klinger, et al., 2003; Yang and Strittmatter, 2007; Sano, Bernardoni and Piacentini, 2012) (fig1). All the members of this protein family contain a highly conserved domain, the 150 to 200 amino-acid (aa) residues, Reticulon Homology Domain (RHD), which is located on the carboxyl-terminal (C-terminus). This domain is composed of two hydrophobic segments of 28 to 36 aa, a region to span the membrane, a 66 aa hydrophilic loop separating the two and a C-terminus tail of about 50 amino acids long (Yang and Strittmatter, 2007; Sano, Bernardoni and Piacentini, 2012; Chiurchiù, Maccarrone and Orlacchio, 2014). This C-terminus structure has been linked to the proteins conserved function and it is hypothesized that it contributes to the curvature of the ER (Sano, Bernardoni and Piacentini, 2012). In contrast to the C-terminus, the amino terminus (N-terminus) does not exhibit any conservation even within isoforms of the same protein group. This high variability gives to the RTN proteins their ability to interact with different targets in their functions (Iwahashi, Hamada and Watanabe, 2007).
RTNs are named after their localization in the ER membrane thanks to the hydrophobic transmembrane segment (Oertle and Schwab, 2003; Yang and Strittmatter, 2007), although they have been found on the cell membrane as well (Grandpré and Strittmatter, 2001). Due to the hydrophobic and hydrophilic segment there arises different conformations and localizations of the protein to facilitate their function. These conformations are according to the localization of the N and C terminals, whether luminal/extracellular or cytosolic/ intracellular (GrandPré et al., 2000; Yang and Strittmatter, 2007; Chiurchiù, Maccarrone and Orlacchio, 2014).
12
The RTN family are transcribed from 4 different genes in different chromosomes. Alternative splicing gives rise to different isoforms. The C- terminal encodes thereticulon homology domain (RHD) which is highly conserved in all the RTN family and is transcribed from the same exons (4-9) except for RTN-2 (6-11). The N-terminal shows no conservation and encodes for specific isoforms for each RTN family. The nogo-66 loop is located in the RHD of all the RTN family with varying homology even in the same organism, however only in the RTN-4 family has it been shown to have neurite and cell spreading inhibitory effects. Data from (Oertle and Schwab, 2003), (Yan et al., 2006).
Reticulon 1 (RTN-1)
previously known as neuroendocrine-specific protein (Roebroek et al., 1993) has three isoforms, RTN-1A, RTN-1B and RTN-1C all of which are mostly expressed in neuroendocrine tissue, especially in dendrites and in Purkinje neurons (Steiner et al., 2004; Shi et al., 2017). In humans the RTN-1 gene is located in chromosome 14, while in mice it is located in chromosome 12 (Yan et al., 2006). The RHD segment of this family has a 188 aa chain that is important for its localization and function. Indeed, the RTN-1 family is mostly found in the ER and has been implicated in the structural conformation of the ER (De Craene et al., 2006; Iwahashi, Hamada and Watanabe, 2007). RTN-1A is the longest in the family consisting of 776 amino acids spliced from exon one, two and three (Yang and Strittmatter, 2007). RTN-1A and RTN-1B share a similar 168 aa section that is spliced from the third exon that gives the two proteins their similar functions and that differ from those of RTN-1C (Yang and Strittmatter, 2007) (Fig.1). RTN-1A and B are thought to be involved in endocytosis by their binding to the adaptor-related protein-50 (AP-50) component of the adaptor complex formed by the transcription factor AP-2 (Iwahashi and Hamada, 2003). Studies also show that members of the RTN are associated with pathologies like Alzheimer’s disease (AD) through their interaction with proteins associated with the disease (He et al., 2004). RTN-1 has been shown to have the least effects on the activities of BACE1, an enzyme implicated in the cleavage of amyloid precursor protein to form amyloid β peptides (APP), which limits BACE1 enzymatic activities (Shi etal., 2017). RTN-1C is the shortest of the three isoforms of RTN-1 and Differs from the other
two isoforms in its (Yang and Strittmatter, 2007; Sano, Bernardoni and Piacentini, 2012)(Fig.1). Similar to the rest of the family RTN-1C has been linked to AD, its expression has been shown to be reduced in patient suffering from this pathology (Tagami et al., 2000). RTN-1C has also been observed to interact with proteins associated with apoptosis, Bcl-2 and Bcl-xl, to reduce their anti-apoptotic activity by binding to them and re localizing them from the mitochondria to the ER (Tagami et al., 2000).
13
Reticulon 2 (RTN-2)
is the least studied of the four reticulon proteins. In humans, RTN-2 is comprised of three isoforms, RTN-2A, B and C with RTN-2A and B sharing more homology than with RTN-2C that has only the RHD segment in common (Roebroek et al., 1998). In humans the RTN-2 gene is located on the chromosome 19, while in mice it is found on the chromosome 7 (Yan et al., 2006). The RHD of the RTN-2 differs from the rest of the reticulon family as it is coded in exon 6-11 unlike the 4-9 of the rest, pointing out a potential difference in terms of its functions and or expression of RTN-2 (Yan et al., 2006)(Fig.1). Indeed, RTN-2C is highly expressed in skeletal muscles unlike the rest that are highly expressed in both the central nervous system and peripheral nervous system tissue, nevertheless RTN-2 is expressed in the nervous system to a certain extent (Roebroek et al., 1998). RTN-2B has been shown to be involved in the trafficking of glutamate transporter EAAC1 from the ER to the cell surface thus increasing its expression (Liu et al., 2008).Reticulon 3 (RTN-3)
the second most studied reticulon, is comprised of two isoforms, RTN-3-A and B (Yang and Strittmatter, 2007). In humans the RTN-3 gene is located on the chromosome 11, while in mice it is located on the chromosome 19 (Yan et al., 2006). RTN-3A is the longest with 1032 amino acids and shares the exon 1 with RTN-3B which has only 236 amino acids (Yang and Strittmatter, 2007)(Fig.1). RTN-3 is highly expressed in the central nervous system especially in pyramidal neurons in the cortex, the hippocampus and the cerebellum (Di Scala et al., 2005). RTN-3 plays a broad range of functions from intracellular trafficking to apoptosis (Yang and Strittmatter, 2007), (Prior etal., 2010), (Wakana et al., 2005). Indeed, Wakana and colleagues observed that RTN-3 was
most likely implicated in the retrograde transport of proteins like BACE1 and Bcl-2 from ER Golgi intermediate compartment and cis Golgi back to the ER (Wakana et al., 2005). RTN-3 have also been co-immunoprecipitated with Synaptic adhesion-like molecules (SALMs), a family of adhesion molecules highly expressed in the CNS and involved in synaptic formation and neurite growth, by interacting with the SALMs through their leucine rich repeat domain (Chang et al., 2010). Furthermore, RTN-3 promotes the binding of Bcl-2 and Beclin-1, proteins involved in both autophagy and apoptosis, to regulate autophagy in cells after ER stress (Chen et al., 2011). RTN-3 has also been linked to Alzheimer’s disease through its interaction with (Kume et al., 2009; Prior et al., 2010).
14
The reticulon 4 (Nogo-A)
is the most studied member of the reticulon family. It is made up of three isoforms, Nogo-A, B and C and like all reticulons they share a similar RHD and differ only on their N-terminus side. In humans the RTN-4 gene is located in chromosome 2, while in mice it is located in chromosome 11 (Yan et al., 2006). Functionally however, the 66 amino acids loop in the RHD of the other family member has no similar function to Nogo (Yang and Strittmatter, 2007). Nogo-A is the longest of the three with approximatively 1192 aa chain, containing a Nogo-A specific region, a N terminal region common with Nogo-B and the RHD domain. Nogo-B is about 373 aa chain and shares a protein region encoded in exon 1 of the Nogo gene with Nogo-A (GrandPré et al., 2000; Oertle and Schwab, 2003; Yang and Strittmatter, 2007). Nogo-C is the shortest of the three isoforms containing only the RHD and a very short N terminal chain forming a 199 aa chain (Oertle and Schwab, 2003; Yang and Strittmatter, 2007)(Fig.1).Like the other RTNs, Nogo family members expression varies in diverse tissues with Nogo-A being principally expressed in the CNS tissues but is also expressed in the testis and heart tissue and at a lower levels in the skeletal muscles (Oertle and Schwab, 2003). Of the three Nogo family members, Nogo-B is the most widely expressed as it is found in nearly all tissue in the body indicating a possible housekeeping function of the protein (Oertle and Schwab, 2003; Oertle, Huber, et al., 2003). Nogo-C is highly expressed in skeletal muscle tissue but also to some extent in the CNS and other organ tissue such as the heart (GrandPré
et al., 2000; Oertle and Schwab, 2003; Oertle, Huber, et al., 2003). In all these tissues the
Nogo proteins are localized on the membrane of the ER, Golgi and on the endoplasmic membrane of the cells. On the cell surface, Nogo-A exerts its inhibitory effects on the nearby cells (GrandPré et al., 2000; Dodd et al., 2005; Yang and Strittmatter, 2007) .
Nogo proteins have diverse functions arising for their structural and expression differences. Nogo-A has two major active sites that confer its functions. The first active site, Δ20, is localized on the Nogo-A specific region, the second, nogo-66, is on the 66 aa loop in the RHD (Oertle, Van der Haar, et al., 2003; Schwab, 2010). The most widely known function for Nogo-A is it inhibitory effect on cellular growth of both CNS (Oertle, Van der Haar, et al., 2003) and non-CNS cells (Chen et al., 2000; Wälchli et al., 2013; Joly et al., 2018) however, it also possess protein trafficking functions (Yang, Harel and Strittmatter,
15
2009), modulation of synaptic plasticity including long-term potentiation and experience dependent plasticity (McGee et al., 2005; Jitsuki et al., 2016), and the maintenance of the ER morphology (Rämö et al., 2016), modulation of inflammation (Fang et al., 2015). On the other hand, Nogo-B has two active domains, one on the N-terminal region and another on the RHD, the 66 aa loop. By virtue of being expressed in many cell type and having many interaction partners Nogo-B is involved in a myriad of functions including modulation of apoptosis (Tagami et al., 2000; Kawaguchi et al., 2018), APP synthesis (He et al., 2004), ER morphological maintenance (Rämö et al., 2016), vascular remodeling (Acevedo et al., 2004) and protein trafficking (Harrison et al., 2009). Even though Nogo-A and B share an active domain and the RHD, Nogo-B does not have neuronal inhibitory functions (Ng and Tang, 2002). Nogo-C is the least studied member of the Nogo family and has largely unknown functions in the CNS (Schwab, 2010). However it has been shown to have similar effects like Nogo-A and B in inhibition of cellular growth in the Peripheral nervous system (PNS) (Kim
et al., 2003). In non CNS studies, Nogo-C has been shown to be involved in apoptosis in
human embryonic kidney cell line HEK293 cell culture (Chen et al., 2006). Through its interaction with ER calcium channels, it also regulates post myocardial infarction fibrosis (Weng et al., 2018).
3. Nogo-A
In this thesis, we will be focusing on Nogo-A and its effects on the visual system especially in the retina and a little on the visual cortex (V1) after retinal injury. We will start with further characterizing Nogo-A, its receptors and signaling pathways. We will then look at its functions in the CNS in general and in the visual system during development and in mature animals. Lastly, in chapter 1 and 2, we shall focus on what we observed in the retina and the visual cortex, in normal conditions and after retinal injury in WT and KO mice and after treatment with Nogo-A function blocking antibodies.
16
3.1 Nogo-A Receptors and signaling pathway
3.1.1. NGR1, PIRB, S1PRs
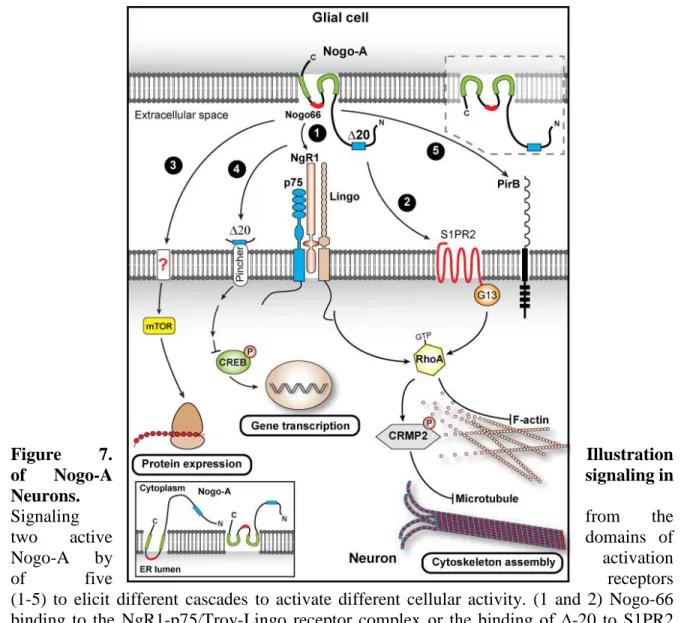
As we have seen above, Nogo-A has two major active domains, one on the Nogo-A specific region, and the other one being the 66 aa loop in the RHD commonly defined as nogo-66 (Oertle, Van der Haar, et al., 2003; Schwab, 2010). Nogo Receptors 1 (NgR1) was the first receptor to be characterized and is a glycosylphosphatidylinositol-linked leucine rich repeat protein (Fournier, GrandPre and Strittmatter, 2001; Schwab, 2010). NgR1 localized on the cell membrane binds to nogo-66 active site to initiate downstream pathways and trigger inhibition of cellular growth and spread in CNS cells including neurons and oligodendrocytes (Fournier, GrandPre and Strittmatter, 2001; Hu et al., 2005). The second receptor recognized as a binding partner of nogo-66 is the Paired immunoglobulin-like
receptor B (PirB) that like NgR1 is involved in cortical plasticity, although it is not clear
what downstream partners are in the inhibitory function of Nogo-A after PirB activation (Filbin, 2008; Djurisic et al., 2013). For ∆-20 on the other hand, its binding receptor was discovered much later. The G-protein coupled receptors of the Sphingolipid 1 phosphate
receptors family (S1PR) member, S1PR2, had an affinity to ∆-20 which upon binding
triggered downstream effectors leading to the inhibitory effects of Nogo-A (Kempf et al., 2014).
3.1.2. LINGO1, p75, TROY
One of the downstream signaling cascade that is activated by nogo-66 binding to NgR1 is the RhoA-ROCK signaling cascade (Fournier, Takizawa and Strittmatter, 2003). This cascade is composed of RhoA, a member of the Rho family of small GTPases that are responsible for several cellular processes including migration, proliferation and morphology, and its downstream effector ROCK, which is a serine-threonine kinase (Leung et al., 1995; Fournier, Takizawa and Strittmatter, 2003; Schwab, 2010). Given that NgR1 does not have a transmembrane domain, it is thought that it requires to form complexes with signal transducing receptors like the Leucine rich repeat and Immunoglobin-like domain-containing
17
(TROY) or p75 neurotrophin receptor (p75) (Barrette et al., 2007; Llorens et al., 2008; Schwab, 2010). Upon activation of the complex, p75 is cleaved by alpha and gamma secretase leading to the release of an intracellular signaling peptide that activates the Rho GTPase pathway (Saha, Kolev and Nikolov, 2014) (Fig.2). Then, the activated RhoA-ROCK signaling cascade leads to the destabilization of actin filaments thus cytoskeleton remodeling (Fournier, Takizawa and Strittmatter, 2003; Schwab, 2010). It is not known if TROY can be cleaved in the same manner as p75to activate its complex (Saha, Kolev and Nikolov, 2014). Likewise, ∆-20 activates the RhoA through its receptor S1PR2 which activates other downstream effectors like the G13, to induce cytoskeletal reorganization (Vogt et al., 2003; Kempf et al., 2014). The binding of the ∆-20 segment and S1PR2 activates the RhoA cascade after being internalized into a signaling endosome in a Pincher-dependent endocytosis (Fig.2). These endosomes are then retrogradely transported back to the soma from the growth cone, where they promote the activation of Rho-GTPase and reduce the levels of
18
phosphorylated cAMP response element binding protein (CREB). A decrease in CREB promotes the inhibitory effects of Nogo-A (Joset et al., 2010).
Figure 7. Illustration
of Nogo-A signaling in
Neurons.
Signaling from the
two active domains of
Nogo-A by activation
of five receptors
(1-5) to elicit different cascades to activate different cellular activity. (1 and 2) Nogo-66 binding to the NgR1-p75/Troy-Lingo receptor complex or the binding of ∆-20 to S1PR2 activates the RhoA pathway to lead to cytoskeleton rearrangement. (3 and 4) Nogo-66 and ∆-20 are also involved in protein and gene transcription through their action on mTOR and CREB. (5) Nogo-66 also binds to the receptor PirB. Different Nogo-a conformations are shown in the insets. Modified from (Pernet, 2017)
19
3.2. The role of Nogo-A in in the development of the central nervous
system in mammals
During the development of the CNS, there are several undergoing cellular processes ranging from cellular differentiation to migration, refinement, and maturation of the different neuronal circuits. Several proteins are expressed in different quantities, times and locations in order to aid these processes by being permissive or inhibitory, attractive or repulsive, or growth promoting or growth hindering in order to have a successfully developed CNS (Petrinovic et al., 2010). Nogo-A is one of these critical proteins, highly expressed in myelin forming oligodendrocytes and also expressed in neurons and other CNS cells in varying amounts (Petrinovic et al., 2010; Pernet, 2017). During development, Nogo-A is highly expressed in neurons, as observed in mice, and its expression decreases as the animal matures (Buss et al., 2005). This points to the possibility that Nogo-A contributes to cellular differentiation, proliferation, and cellular migrations (Seiler, Di Santo and Widmer, 2015). Indeed, using mouse models, Ramasamy and colleagues observed that the administration of nogo-66 substrates in cellular cultures increased differentiation of neural progenitor cells into an astro-glial lineage (Ramasamy et al., 2014). They also observed that the stimulation of cortical neural stem cells with Nogo-66 promotes their survival and proliferation through interaction with the NgR1 complex or PirB complex (Ramasamy et al., 2014) (Fig.2). Furthermore, it has been observed that Nogo-A, through the NgR1/LINGO-1 complex, influenced the radial migration of murine cortical neuronal precursor cells (Mathis et al., 2010), the tangential migration of cortical GABAergic interneurons (Mingorance-Le Meur
et al., 2007), and the migration of murine neuroblasts to the olfactory bulb through the ∆-20
domain (Rolando et al., 2012). It can also be hypothesized that Nogo-A works in the same way in humans, as the expression of Nogo-A in the developing murine CNS is similar to that in human CNS (Seiler, Di Santo and Widmer, 2015).
3.3. Nogo-A implications in neuronal functions
In the mature brain, Nogo-A also has diverse expression patterns and functions throughout the CNS.
20
3.3.1. Neuronal cytoskeletal modelling
The most known is its inhibitory effects on neurite outgrowth and growth cone collapse. This collapse is due to the reassembly of microtubule and actin filaments. This reassembly is done through the RhoA-ROCK pathway. The binding of Nogo-66 to its respective receptor complexes activates this pathway leading to a cascade of processes. RhoA activates its major downstream effector ROCK which has several effects on the cytoskeletal proteins associated to actin and tubulin (Mulherkar and Tolias, 2020). One of these associated proteins is cofilin, a member of the actin depolymerizing factor /cofilin family which is an actin-binding protein responsible for actin turnover. The RhoA-ROCK pathway induces LIM kinase-dependent phosphorylation and inactivation of cofilin, thus destabilizing the actin filaments dynamics (Maekawa et al., 1999; Kanellos and Frame, 2016). Another of these associated proteins is collapsin response mediator protein-2, a microtubule-binding protein. Activation of the RhoA-ROCK pathway leads to the phosphorylation of this protein preventing its binding to tubulin and thus collapsing of the microtubule scaffolding of the neuron at the growth cone (Arimura et al., 2005).
3.3.2. Neuronal gene and protein expression
Growth cone collapse mediated by Nogo-A can be dependent on the concentration of associated proteins transcription and synthesis (Manns et al., 2014). For example, Nogo-A has been associated with neuronal gene and protein expression in the CNS due to its ∆-20 domain’s link to transcription factors like CREB, and protein synthesis pathways like the
mammalian target of rapamycin pathway (mTOR), a major regulator of translational
initiation (Manns et al., 2014). Indeed, the retrograde transport of the internalized ∆-20 domain in endosomes has been shown to lead to a downregulation of phosphorylated CREB (Joset et al., 2010) in turn leading to a sustained effect of Nogo-A (Pernet, 2017). Likewise, the ∆-20 domain of Nogo-A increased growth cone phosphorylation of eukaryotic initiation factor 4E binding protein 1, a key downstream target of mTOR complex 1 (Manns et al., 2014). Furthermore the association of Nogo-A and the mTOR pathway also affects the post translational regulation of gamma-aminobutyric acids B receptor (GABAB), the synaptic