Dynamique de succession forestière dans les forêts
feuillues nordiques après coupes partielles réalisées
dans un contexte industriel
Mémoire
Émilie St-Jean
Maîtrise en sciences forestières - avec mémoire
Maître ès sciences (M. Sc.)
Dynamique de succession forestière dans les forêts
feuillues nordiques après coupes partielles réalisées
dans un contexte industriel
Mémoire
Émilie St-Jean
Sous la direction de:
Alexis Achim, directeur de recherche Christian Messier, codirecteur de recherche
Résumé
L’augmentation en dominance du hêtre à grandes feuilles (Fagus grandifolia Ehrh.) jumelée à la diminution de l’érable à sucre (Acer saccharum Marsh.) en régénération menace la viabilité économique des forêts feuillues nordiques aménagées. En se basant sur l’hypothèse que l’aug-mentation de la disponibilité de la lumière pour la régénération favoriserait l’érable à sucre, la coupe progressive irrégulière est passée d’une mise en oeuvre expérimentale à opérationnelle à grande échelle au cours de la dernière décennie. Nous avions comme objectif de vérifier si des variantes de coupes partielles, telles que la coupe de jardinage et la coupe progressive irrégulière, faites dans un contexte industriel, avaient tendance à favoriser la régénération de l’érable à sucre au détriment de celle du hêtre. Nous avons mesuré le coefficient de distri-bution, l’abondance et la croissance de la régénération dans des coupes industrielles âgées de trois à six ans dans la forêt feuillue du Québec, Canada. Le suivi de la régénération a per-mis de démontrer une indéniable dominance du hêtre à grandes feuilles, tout type de coupe confondu, particulièrement chez les grands semis et gaules. Cependant,dans certaines condi-tions, les indices de probabilités de transition (combinant la croissance et l’abondance relative) de l’érable à sucre pouvaient être légèrement favorisés par l’augmentation du prélèvement en surface terrière, dans certaines conditions. En se basant sur le coefficient de distribution et la surface terrière marchande du hêtre avant la coupe, nous avons proposé de nouveaux critères pour aider à décider dans quelles situations une augmentation de l’intensité de la coupe par-tielle permettrait de favoriser significativement la régénération de l’érable à sucre. Au-delà de ces résultats, l’étude met en lumière l’importance de raffiner le processus de diagnostic et les prescriptions sylvicoles de nos forêts feuillues afin de limiter efficacement l’expansion du hêtre à grandes feuilles.
Table des matières
Résumé ii
Table des matières iii
Liste des tableaux iv
Liste des figures v
Remerciements vi
Avant-propos vii
Introduction 1
1 Transition probability modelling of regeneration shows when increased levels of harvest could favour sugar maple over American beech in
northern hardwoods 5
1.1 Abstract . . . 5
1.2 Résumé . . . 6
1.3 Introduction. . . 7
1.4 Material and methods . . . 10
1.5 Results. . . 17
1.6 Discussion . . . 23
1.7 Conclusion . . . 29
Conclusion 30
Liste des tableaux
1.1 Details of the fitted gamlss model. . . 22
1.2 Proposition de traitements sylvicoles à appliquer afin de limiter l’expansion du
Liste des figures
1.1 Study area and location of the 12 harvesting sites . . . 11
1.2 Variable-radius plot design. . . 12
1.3 Attributes related to understory resource availability per treatment. . . 17
1.4 Abundance (mean + SE) and dominance of regeneration. . . 18
1.5 Predicted time to reach two meters in height as a function of measured subplot canopy for five hardwood species and the other species pooled together, for two height classes (small and tall seedlings) and for two subplot locations (skid-trail or strip) . . . 20
1.6 Distribution of transition probability indices per species. . . 21
1.7 Predictions of transition probability indices for AS as a function of plot basal area removal per class of AS pre-treatment occurrence in the advance growth and pre-treatment FG basal area (FG BA). . . 23
1.8 Predictions of transition probability indices for AS as a function of subplot canopy opening per class of AS pre-treatment occurrence in the advance growth and initial FG basal area (FG BA). . . 24
Remerciements
Je tiens d’abord à remercier mon directeur, Alexis Achim. J’ai la chance de compter sur son enthousiasme contagieux et sa générosité depuis cinq ans. Sa confiance et ses encouragements m’ont certainement amenée "à un autre niveau". Il a largement contribué à parsemer mon parcours d’expériences mémorables. La plus précieuse aura été sans aucun doute mon stage à Forest Research, à Édimbourg en Écosse. Un grand merci à Bruce, Juan et Tom pour m’avoir invitée à travailler avec vous à FR.
J’ai également eu la chance de rencontrer Christian Messier et Philippe Nolet, qui m’ont tous deux généreusement accueillie à l’ISFORT. Ils ont contribué à élargir ma compréhension de l’écosystème forestier.
Je remercie tous mes talentueux collègues du labo AA : André, Christine, Claude, David, Guillaume, Iman, Joëlle, Jean-Romain, Lady, Laurence, Thomas et Yann. Mention spéciale à Alex, qui a sacrifié son amour du soleil et de la chaleur pour un stage en Écosse. Merci également à tous mes précieux collègues, amis et partenaires de balon-balai du ABP/GHK et de l’ISFORT. Je n’aurais pas pu accomplir cette maîtrise sans l’aide des multiples auxi-liaires qui m’ont accompagnée durant ses deux dernières années : Oli, Floren(c)e, Raph, Luc, Émile, Myriam et Félix. Merci pour votre bonne humeur inébranlable, votre patience et votre ingéniosité malgré les adversités du terrain.
Merci au Ministère des Forêts, de la Faune et des Parcs du Québec en Outaouais et dans les Laurentides de m’avoir partagé de nombreuses et précieuses données. Un merci particulier à Sébastien Meunier pour m’avoir ouvert les yeux sur les nombreuses problématiques de la forêt feuillue et pour tout le temps donné pour le bon fonctionnement du projet.
Je remercie le Conseil de recherches en sciences naturelles et en génie du Canada et le Fonds de recherche du Québec-Nature et technologies pour le soutien financier. Merci au programme de Bourses canadiennes du jubilé de diamant de la reine Elizabeth II (et à la grande Évelyne Thiffault pour y avoir appliqué).
Finalement, merci à mes amis et à ma famille. Dan et Loulou, votre éternel support m’a permis de me rendre jusqu’ici. Surtout, merci beaucoup à Renaud qui m’a soutenue de près et de loin durant les hauts et les bas de cette belle aventure à Québec, à Gatineau et en Écosse.
Avant-propos
Ce mémoire est constitué d’un article rédigé en anglais, d’une introduction et d’une conclusion générale en français. L’article a été rédigé dans le but d’être publié dans une revue scientifique. Les questions et hypothèses de recherche ont été établies avec l’aide de mon directeur de recherche, Alexis Achim. J’ai fait les analyses statistiques avec les données du Ministère des Forêts, de la Faune et des Parcs du Québec et mes propres données récoltées sur le terrain. J’ai interprété les résultats et rédigé le manuscrit, je suis donc l’auteure principale de l’article. Mon directeur a participé à l’élaboration du projet de recherche, et a fourni d’indispensables conseils tout au long du projet sur la méthodologie, l’analyse des résultats et la rédaction. Mon codirecteur Christian Messier et troisième collaborateur Philippe Nolet, m’ont également supportée lors de la mise en oeuvre de la méthodologie, l’analyse des résultats et la rédaction. Ils sont donc également co-auteurs de l’article scientifique. Finalement, Sébastien Meunier, préalablement maître de stage au projet au Ministère des Forêts, de la Faune et des Parcs, est le cinquième auteur de l’article, pour sa large contribution à l’élaboration de la problématique et des objectifs et ses conseils pour la méthodologie.
Introduction générale
La dynamique de la forêt feuillue du nord-est de l’Amérique du Nord, et par conséquent son aménagement, sont en mutation. L’aménagement de cette forêt doit tenir compte des nombreux services écosystémiques qu’elle fournit à la population (Leak et al., 2014). Au Québec, la forêt feuillue se retrouve au sud, là où la majorité de la population réside. Ses paysages ont donc une grande valeur, esthétique notamment, et jouent un rôle important pour la santé et le bien-être des gens qui la côtoient (Karjalainen et al., 2010). Cette proximité la rend également facile d’accès pour y faire toutes sortes d’activités, de la chasse, à la randonnée, à la production de sirop d’érable. La forêt feuillue du nord-est de l’Amérique du Nord est également l’habitat d’une grande diversité d’espèces fauniques et florales (Leak et al.,2014;
Ordre des ingénieurs forestiers du Québec,2009). Au Québec, la majorité de la forêt feuillue se retrouve dans les domaines bioclimatiques de l’érablière à bouleau jaune et de l’érablière à tilleul. Ces domaines totalisent près de 96 000 km2 du territoire (Saucier et al.,2009). L’érable
à sucre (Acer saccharum), le bouleau jaune (Betula alleghaniensis) et plus marginalement le chêne rouge (Quercus rubra) s’y retrouvant sont parmi les trois espèces de feuillus nobles les plus désirées par l’industrie forestière oeuvrant sur ce territoire actuellement.
Il a été longtemps pris pour acquis que la régénération en érable à sucre s’établissait facilement suite aux coupes partielles dans les forêts tempérées nordiques (Eyre and Zillgitt,1953;Tubbs,
1977, 1968). Pourtant, de plus en plus d’inquiétudes sont soulevées quant à la durabilité de la présence de l’érable à sucre dans les forêts feuillus nordiques aménagées (Bose et al.,2017;
Duchesne et al.,2005;Matonis et al.,2011). Son abondance en déclin jumelée à l’augmentation de la proportion de hêtre à grandes feuilles en régénération sont à la base de ces inquiétudes (Bose et al.,2017;Duchesne et al.,2005;Gauthier et al.,2015).
Ce phénomène est d’autant plus préoccupant puisqu’en parallèle se déploie la maladie corti-cale du hêtre (MCH). Elle résulte de l’infection d’un type de cochenille (Cryptococcus fagisuga Lind.) d’origine exotique, suivie de champignons pathogènes (Neonectria faginata and N. di-tissima)(Cale et al.,2013;Houston,1994). La MCH entraîne une dégradation du bois qui peut limiter le potentiel de valorisation du hêtre (Boulet and Huot,2013). Une grande quantité de drageons de hêtre peuvent émerger des racines de l’arbre mère en réaction à sa dégradation ou à l’ouverture du couvert subséquent (Boulet and Huot,2013;Hane,2003;Houston,1994;Roy
and Nolet,2018). Les drageons de hêtre auraient d’ailleurs de meilleures chances de survie que les semis en plus d’avoir une meilleure croissance (Beaudet et al.,2007;Beaudet and Messier,
2008).
L’érable à sucre préfère les sols riches avec un drainage bon à modéré, mais il a une grande ca-pacité d’adaptation (Boulet and Huot,2013). Depuis quelques décennies cependant, il connaît à certains endroits au Québec une augmentation de mortalité, de déperissement, de mort en cime et donc de diminution en qualité (Côté and Ouimet, 1996). Le hêtre à grandes feuilles pourrait également être une cause du déclin de l’érable à sucre. La diminution de la dispo-nibilité de lumière causée par l’augmentation de l’abondance de gaules de hêtres limite le développement de la banque de semis de l’érable à sucre : sa stratégie de régénération princi-pale (Hane,2003;Marks and Gardescu,1998).
Malgré le rôle écologique important du hêtre, on cherche donc depuis de nombreuses années à refréner le phénomène appelé « l’envahissement du hêtre dans les érablières » (Bose et al.,
2017;Duchesne et al.,2005). À l’échelle provinciale, l’abondance du hêtre à grandes feuilles a considérablement augmenté dans les dernières décennies. Dans la sous-région 3bm, du domaine bioclimatique de l’érablière à bouleau jaune, la densité moyenne aurait augmenté de plus de 600 tiges/ha entre la première (1970-1979) et la 4e campagne d’inventaires (2003-2014), dans les placettes-échantillons permanentes du Ministère des Forêts, de la Faune et des Parcs (MFFP) (Direction de la recherche forestière, 2017). L’augmentation en densité peut être expliquée en partie par le fait que cette espèce a été longtemps délaissée par les compagnies forestières (Boulet and Huot,2013). Les blessures de racines des hêtres résiduels aurait favorisé le développement de drageons (Jones and Raynal,1988). L’érable à sucre est plus brouté que le hêtre par les herbivores comme le cerf de Virgine (Odocoileus virginianus) et l’orignal (Alces alces) (Bose et al., 2018; Matonis et al., 2011). Le phénomène d’envahissement a aussi été associé aux conditions pédologiques (Duchesne and Ouimet,2009), mais des liens de causalité demeurent difficiles à établir (Messier et al.,2011).
La disponibilité de lumière affecte la dynamique de l’érable à sucre et du hêtre. Ces deux espèces sont communément considérés comme très tolérantes à l’ombre et peuvent survivre plusieurs décennies sous couvert (Boulet and Huot, 2013; Burns et al., 1990). En revanche, le hêtre croît en hauteur plus rapidement que l’érable à sucre dans des conditions de faible ouverture de canopée (Canham,1988, 1990). Selon Bose et al. (2017), sans perturbation na-turelle majeure ou sans la création de grandes trouées par la récolte forestière, les forêts du nord-est des États-Unis seront de plus en plus dominées par le hêtre. La coupe de jardinage, un système de régénération créant de petites trouées en prélevant un ou plusieurs tiges uni-formément (communément recommandé pour l’établissement d’espèces tolérantes à la lumière comme celle des forêts feuillues du Québec), aurait avantagé le hêtre au détriment de l’érable à sucre (Bannon et al.,2015;Collin et al.,2017;Larouche et al.,2013;Nolet et al.,2008;Poulson and Platt,1996). Nolet et al. (2008) soutiennent que l’érable serait avantagé par rapport au
hêtre par des perturbations plus sévères.
Plusieurs études ont démontré que la création récurrente de grandes trouées produisant une plus grande disponibilité de lumière favoriserait l’érable à sucre (Canham,1988;Poulson and Platt,1996). Dracup et MacLean (2018) ont suggéré que 45% de prélèvement en surface terrière était suffisant pour limiter les conditions favorables à la régénération en hêtre. Cependant, la majorité des connaissances acquises sur le sujet a été acquise à petite échelle dans des expériences contrôlées, qui peuvent être moins représentatives des conditions créées par des interventions à une échelle industrielle (Puettmann et al., 2009). Des études sur les coupes partielles ont révélées d’importantes différences entre un traitement expérimental et industriel, particulièrement en lien avec l’hétérogénéité spatiale du prélèvement et de la disponibilité de la lumière pour la régénération (Guay-Picard et al., 2015; Guillemette et al., 2013; Moreau et al.,2020).
Un vaste territoire comme la forêt feuillue publique du Québec génère un contexte dans lequel il n’y a pas de solution sylvicole simple pour favoriser l’établissement et le recrutement de l’érable à sucre. L’approche sylvicole ayant actuellement le plus de succès, la création d’exclos et le débroussaillage du hêtre, est coûteuse et n’est peut-être pas nécessaire dans toutes les situations (Bohn and Nyland,2003;Bose et al.,2018;Nyland et al.,2006). Le rétablissement et le recrutement agressif du hêtre ainsi que le déclin subséquent de l’érable à sucre menacent la viabilité économique des forêts feuillues aménagées dans un contexte industriel (Nelson and Wagner, 2014). L’augmentation de la densité du hêtre aux dépens de l’érable à sucre pourrait avoir comme effet de réduire la biodiversité des forêts concernées (Cale et al.,2013). La disponibilité de la lumière est un facteur externe de croissance facile à contrôler avec les opérations forestières : les forestiers peuvent contrôler la largeur d’une trouée (Kern et al.,
2017).
Les aménagistes du Québec prescrivent de moins en moins de coupe de jardinage (Ministère des Forêts, de la Faune et des Parcs,2009,2020). Plusieurs autres raisons expliquent ce chan-gement, mais ils s’appuient entre autres sur les précédents résultats qui soutiennent que le hêtre est présentement favorisé par les faibles taux de prélèvement associés aux coupes de jar-dinage (Direction de la recherche forestière,2017). C’est donc la coupe progressive irrégulière qui est depuis une dizaine d’années le traitement le plus communément appliqué (Ministère des Forêts, de la Faune et des Parcs,2020). Par ses différentes variantes, la coupe progressive irrégulière est un système de régénération flexible permettant de créer des conditions favo-rables à l’établissement d’espèces au besoins différents, mais peut être également définie par un taux de prélèvement plus grand que les coupes de jardinage (Larouche et al., 2013; Ray-mond et al.,2009). C’est la mesure de l’efficacité de ce changement de cap , mis en oeuvre à grande échelle et ayant pour but d’augmenter la disponibilité de lumière pour la régénération, qui constitue la motivation première à la réalisation du présent projet. Selon la Direction de la recherche forestière (2017), les modèles doivent tenir compte des « effets des traitements
sylvicoles sur la régénération des espèces à promouvoir ». Préciser les effets des interventions en forêt permettrait de mieux modéliser la dynamique de succession, et par le même fait la croissance, de la forêt feuillue du nord-est de l’Amérique du Nord des prochaines décennies.
Objectifs
L’objectif principal du projet était d’évaluer les effets des coupes partielles récentes appliquées dans les forêts feuillues du sud-ouest du Québec sur la dynamique de l’érable à sucre et du hêtre à grandes feuilles.
Objectif spécifique et hypothèse :
Plus spécifiquement, notre but était de vérifier si les différentes variantes de coupe partielle produites dans un contexte industriel tendent à favoriser une succession forestière avec davan-tage d’érable à sucre au détriment du hêtre à grandes feuilles.
Nous posons l’hypothèse qu’une augmentation de la disponibilité de la lumière en lien avec une augmentation de l’intensité d’une coupe partielle favoriserait l’établissement et la croissance de la régénération d’érable à sucre aux dépens du hêtre à grandes feuilles.
Chapitre 1
Transition probability modelling of
regeneration shows when increased
levels of harvest could favour sugar
maple over American beech in
northern hardwoods
1.1
Abstract
The increasing dominance of American beech (FG, Fagus grandifolia Ehrh.) and the decreas-ing proportion of sugar maple (AS, Acer saccharum Marsh.) in regeneration challenges the long-term economic viability of commercially managed northern hardwoods in eastern North America. Based on, inter alia, earlier studies showing that more light in the understory favours AS over FG, the irregular shelterwood system has shifted from experimental to operational implementation over the last decade. In this paper, we evaluated the success of such this shift toward irregular shelterwood in promoting AS over FG by measuring the occurrence, abun-dance of AS and FG seedlings and saplings, and growth of seedlings three to six years after logging in northern hardwood forests of western Quebec, Canada. Results showed a domi-nance of FG regeneration regardless of the harvest intensity, particularly among tall seedlings and saplings. However, we found that the transition probability indices (projected relative combined abundance and growth) of AS could be favoured, albeit to a limited extent, by an increased basal area removal, particularly where the initial occurrence of AS in the advance seedling regeneration is lower than 60% and the initial FG basal area is low in the overstory (i.e. 6m2ha-1). Our results highlight the importance of refining our operational management
strategy to effectively limit the increasing abundance of regeneration of American beech over sugar maple in northern hardwood forests
1.2
Résumé
L’augmentation en dominance du hêtre à grandes feuilles (Fagus grandifolia Ehrh.) jume-lée à la diminution de l’érable à sucre (Acer saccharum Marsh.) en régénération menace la viabilité économique des forêts feuillues nordiques aménagées. En se basant sur l’hypothèse que l’augmentation de la disponibilité de la lumière pour la régénération favoriserait l’érable à sucre, la coupe progressive irrégulière est passée d’une mise en oeuvre expérimentale à opé-rationnelle à grande échelle au cours de la dernière décennie. Nous avions comme objectif de vérifier si des variantes de coupes partielles, telles que la coupe de jardinage et la coupe progressive irrégulière, faites dans un contexte industriel, avaient tendance à favoriser la régé-nération de l’érable à sucre au détriment de celle du hêtre. Nous avons mesuré le coefficient de distribution, l’abondance et la croissance de la régénération dans des coupes industrielles âgées de trois à six ans dans la forêt feuillue du Québec, Canada. Le suivi de la régénération a permis de démontrer une indéniable dominance du hêtre à grandes feuilles, tout type de coupe confondu, particulièrement chez les grands semis et gaules. Cependant,dans certaines condi-tions, les indices de probabilités de transition (combinant la croissance et l’abondance relative) de l’érable à sucre pouvaient être légèrement favorisés par l’augmentation du prélèvement en surface terrière, dans certaines conditions. En se basant sur le coefficient de distribution et la surface terrière marchande du hêtre avant la coupe, nous avons proposé de nouveaux critères pour aider à décider dans quelles situations une augmentation de l’intensité de la coupe par-tielle permettrait de favoriser significativement la régénération de l’érable à sucre. Au-delà de ces résultats, l’étude met en lumière l’importance de raffiner le processus de diagnostic et les prescriptions sylvicoles de nos forêts feuillues afin de limiter efficacement l’expansion du hêtre à grandes feuilles.
1.3
Introduction
The increase in abundance of American beech (FG, Fagus grandifolia Ehrh.) over sugar maple (AS, Acer saccharum Marsh.) in the regeneration in northern hardwood forests of eastern North America in the last decades is concerning in light of the sustainable production of good quality sugar maple timber. Not only is FG not normally a desirable species for timber production, it is also severely affected by the exotic beech bark disease, an infection by two fungi(Neonectria faginata and N. ditissima)that can be exacerbated by the presence of an exotic insect (beech scale, Cryptococcus fagisuga Lindinger) (Houston, 1994). The disease causes FG wood degradation and tree death without reducing the overall abundance of the species (Cale et al., 2013; Garnas et al., 2011; Houston, 1994). On the contrary, either as a reaction to the disease or as a consequence of an increased opening in the canopy, a dense thicket of FG suckers and seedlings often develops in affected stands (Giencke et al., 2014;
Houston, 2001; Roy and Nolet, 2018). Due to both its high shade-tolerance and ability to re-sprout from roots as suckers following disturbances, FG tends to be highly competitive in forests affected by small scale disturbances (Beaudet et al.,1999;Roy and Nolet,2018). As a result, a feedforward mechanism is in place in northern hardwood forests whereby the factors negatively affecting the vigor and quality of FG also promote its regeneration, at the expense of local biodiversity (Cale et al.,2013;Giencke et al., 2014) and of the yield of high quality timber (Pothier et al.,2013).
AS is one of the most valuable species from northern hardwood forests and is highly sought after for so-called ‘appearance’ wood products such as furniture, cabinets and flooring as well as for maple syrup production. It is also the main species found in the northern hardwoods of Quebec : in the bioclimatic zone where AS is the dominant species, AS accounts for on average 15% to 25% of the growing stock volume (Boulet and Huot, 2013). AS prefers rich soil with moderate to good drainage, but it is highly adaptable (Boulet and Huot,2013). Over the last few decades in Quebec, the species has experienced increased mortality, defoliation of the crown, and dieback resulting in decreased timber quality in Quebec (Bal et al.,2014;Côté and Ouimet,1996). Although environmental stressors have been identified as probable causes for this AS decline (Bal et al., 2014; Bauce and Allen, 1991), the increase of FG abundance in regeneration has also been considered as a predominant cause of AS regeneration failure (Direction de la recherche forestière,2017). Decreased light availability due to increasing FG sapling abundance limits the development of the AS seedling bank, the main AS regenerative strategy (Hane,2003;Marks and Gardescu,1998).Hane et al. (2003) suggested that negative effects of FG thickets on AS also occur belowground where two species compete for the same resources, and this could be exacerbated by a potential phytotoxic effect of FG on AS. Past forestry practices, soil degradation, FG sprouting and preferential browsing are also re-cognized as contributing to some extent to FG expansion and AS decline. In Quebec, the increasing abundance of FG can also be partially explained by the fact that the species is
of-ten left behind by forest companies (Boulet and Huot,2013;Majcen et al.,2003). In addition, damaged roots of residual FG trees resulting from logging operations may develop root sprouts (Jones and Raynal,1988). The decrease of the soil pH has also been associated with FG ex-pansion (Arii and Lechowicz,2002;Duchesne and Ouimet,2009) although causal relationships remain hard to identify (Bannon et al.,2015;Messier et al.,2011). Preferential browsing also affects northeastern hardwood forest succession as white-tailed deer (Odocoileus virginianus) and moose (Alces alces) were found to prefer AS over FG (Bose et al.,2018;Elenitsky et al.,
2020;Matonis et al.,2011).
In the absence of severe disturbances, the limited availability of light in the understory may also alter the dynamics of AS and FG regeneration. Both AS and FG can survive for several decades in the understory (Canham, 1990; Poulson and Platt, 1996). AS and FG have a similar ecological niche ; however, under a very dense canopy, FG performs better than AS and has greater lateral growth (Beaudet et al.,1999;Burns et al.,1990;Canham,1988;Poulson and Platt, 1996). Repetitive single-tree selection cuts have been recommended for northern hardwoods as they create small gaps to establish shade-tolerant species. This silvicultural approach may thus favour FG regeneration over AS since these gaps create low light availability conditions with limited variability (Bannon et al.,2015;Collin et al.,2017;Nolet et al.,2008;
Poulson and Platt,1996).
Accordingly, several studies have recommended that AS regeneration could be favoured over FG in larger gaps or following a greater removal of the overstory canopy. Nolet et al. (2008 )in-dicated that a partial cut retaining more than 25% of the forest cover was not sufficient to favour AS at the expense of FG although Dracup and MacLean (2018) suggested that removing 45% of basal area is sufficient to limit the conditions that are favourable for FG regeneration. Other studies have also suggested that the repetitive occurrence of large gaps may favour AS over FG by providing higher light availability in the understory (Canham, 1988; Houston,
2001;Poulson and Platt,1996). However, most of these results were obtained from carefully controlled experiments which may not be representative of the conditions created by silvicul-tural interventions conducted at the industrial scale (Puettmann et al.,2009). Previous studies of partial harvesting have revealed important differences between experimental and applied treatment results, especially in terms of the higher spatial heterogeneity of tree removal, and of light availability in the understory, after commercial operations (Guay-Picard et al.,2015;
Guillemette et al.,2013;Moreau et al.,2020).
The vast territory of Quebec’s public forests creates a situation where there is no simple silvi-cultural solution to favour the sustainable establishment and recruitment of AS. The currently recommended approach of mechanical removal of FG and fencing to protect against browsing (Bohn and Nyland,2003;Bose et al.,2018;Nyland et al.,2006) may prove expensive to apply at the regional scale and it may not be necessary in all situations. Applying glyphosate to control FG regeneration is a recognized solution (Bose et al.,2018),but its use is banned in Québec’s
forests. We therefore need to implement other cost-effective, efficient solutions as the vigorous understory re-establishment and recruitment of FG, and concomitant decline of AS, following logging threatens the economic viability of these commercially managed forests(Direction de la recherche forestière,2017;Nelson and Wagner,2014).
Based on the experimental evidence cited above that large openings could create regeneration conditions more favourable to AS than FG, inter alia, practitioners in western Québec have changed their preferred management approach in northern hardwood forests (Ministère des Forêts, de la Faune et des Parcs,2020,2009). Instead of selection cutting, variants of the irre-gular shelterwood system are now commonly applied (Ministère des Forêts, de la Faune et des Parcs,2020). With their different variants (continuous-cover, irregular and expanded-gap), ir-regular shelterwood systems are flexible regeneration systems that facilitate the establishment of species with different light requirements (Raymond et al.,2009). They can also be defined by a higher harvesting rate than selection cutting and more irregular cutting cycles (Larouche et al.,2013). However, no long-term studies have yet confirmed that this shift towards irregular shelterwood systems is efficient in favouring AS regeneration over FG.
To rigorously test the hypothesis that the conditions created by a higher harvesting rate in partial cuts can help favour AS regeneration over FG in commercially harvested forests, we re-sampled 96 plots three to six years after the application of various partial cutting treatments in the northern hardwood forest of Quebec, Canada. Our sampling plots were distributed among 12 harvesting sites where a range of treatments from single-stem selection to irregular shelterwood had been applied. In addition to assessing the current composition of the fo-rest regeneration, we measured the growth of seedlings and projected growth over time using transition probability indices (Guay-Picard et al.,2015;Hill et al.,2005;Kneeshaw and Ber-geron,1998). These indices portray the short-term effect of harvesting by weighting a species abundance with its growth rate.
1.4
Material and methods
1.4.1 Study area
The study area is located in the northern hardwood forest of southwestern Quebec, Canada between longitudes 74◦30’ 0” W and 76° 30’ 0” W and latitudes 45◦47’ 60” N and 46° 36’
0” N. Elevation in the region ranges from 270 to 440 m. The area is within the AS – yellow birch (Betula alleghaniensis Britton) bioclimatic domain of Quebec’s northern hardwood forest type. Mean annual temperatures in this domain vary between 2.5 and 4.0 ◦C (Saucier et al.,
2009). Mean annual precipitation average around 1000 mm (Saucier et al., 2009). Although it is dominated by AS, the main forest canopy in the area is also composed of FG, yellow birch, red maple (Acer rubrum L.) red oak (Quercus rubra L.), balsam fir (Abies balsamea [L.] Miller), eastern hemlock (Tsuga canadensis [L.] Carrière), American hop-hornbeam (Ostrya virginiana [Mill.] K. Koch), and American basswood (Tilia americana L.). Approximately half of the study area experienced different intensities of forest tent caterpillar outbreaks in 2016, 2017 and 2018 (Malacosoma disstria). The insect feeds on codominant and dominant AS tree leaves in northern hardwood forests, but can also eat other deciduous species except red maple (Boulet and Huot,2013;Fortin and Mauffette,2002;Ordre des ingénieurs forestiers du Québec,2009). Most part of the study area has been subjected to two or three commercial harvest over the last century. The study area was located across both the advance front and the killing front of the beech bark disease and damage was mostly prevalent in its southern part (Ministère des Forêts, de la Faune et des Parcs,2019;Roy and Nolet,2018;Shigo,1972). Beech bark disease, or beech scale, was observed in half of our sites. The mean density of white-tailed deer was estimated as ranging from 2.26 to 3.05 per km2 in 2017 for the whole
study area, but it can be expected even lower since our sites are from fields of crops (Ministère des Forêts, de la Faune et des Parcs, personal communication, January 11th, 2020) while mooses are present, but less extensive (∼ 0.24 per km2 in 2002) in the study area (Ministère
des Forêts, de la Faune et des Parcs,2015).
1.4.2 Stand selection
Quebec’s Ministry of Forests, Wildlife and Parks provided a database of available sampling plots established before harvesting. Twelve felling sites were first selected, each consisting of various partial cutting treatments conducted between 2013 and 2016. Throughout each felling site, plots had been established systematically every 150 to 200 meters prior to harvest. We selected plots from the database with at least 25% of the basal area consisting of either AS or FG before treatment, with good or moderate drainage and with at least 50-cm-thick soils of glacial or fluvio-glacial origins, as mapped by the Ministry with photo-interpretation. These soil characteristics are the most common in the study area. We then applied a stratified random sampling method to select eight plots per harvesting site. To ensure variability in basal area removal, plots were selected across a range of partial cutting treatments i.e. single-tree
selection cuts, continuous-cover irregular shelterwood cuts and extended irregular shelterwood cuts. Except when restricted by regulations or where terrain limited access, all plots had been harvested with fully mechanized operations. Harvesting operations were conducted either in the late summer, fall or winter. A total of 96 plots were sampled during the summer of 2019 (Figure 1.1). Québec ! H ! H ! H ! H!H ! H ! H ! H!H !H ! H!H Outaouais Laurentides Ontario -74°0'0" -74°0'0" -75°0'0" -75°0'0" -76°0'0" -76°0'0" -77°0'0" 47 °0 '0 " 47 °0 '0 " 46 °0 '0 " 46 °0 '0 " ! H Sites Studied regions Montréal
Figure 1.1 – Study area and location of the 12 harvesting sites
1.4.3 Data collection
Pre-treatment sampling
The sampling plots database provided by the Ministry contained the pre-treatment data. Variable-radius plots were established with a basal area prism of a factor of 2. All trees with a DBH larger than 9 cm were tallied. Pre-treatment regeneration sampling considered seedlings with minimum heights of either 15 cm or 40 cm, depending on the sites. In the Outaouais region, a cluster of five, fixed-radius subplots was established, one in the center of the plot and four others in the main cardinal directions, 10 meters from the center. Following provincial standards, the subplot radius depended on the species sampled (between 1.13 m and 2.82
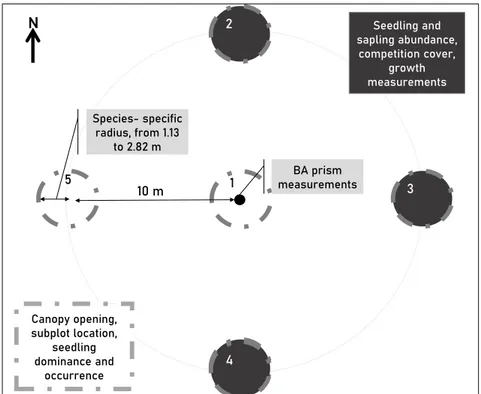
m). The regeneration sampling design in the Laurentides region also consisted of five fixed-radius subplots, using the same species-specific radii. However, in this region the first plot was established 6 meters from the center in the direction of the next plot and the four others were successively established at a 6-meter interval in the same direction. In both regions, the presence of seedlings was recorded for each species to indicate occurrence, but not their abundance. 10 m BA prism measurements 1 2 4 5 N 3 Seedling and sapling abundance, competition cover, growth measurements Canopy opening, subplot location, seedling dominance and occurrence 4 Species- specific radius, from 1.13 to 2.82 m
Figure 1.2 – Variable-radius plot design. In all subplots (grey dotted outline), we noted subplot location and measured canopy opening, seedling dominance and occurrence. In subplot 2, 3 and 4 (black filling), we measured seedling and sapling abundance, competition cover and seedling growth. We measured basal area (BA) from the center with a prism of factor 2.
Post-treatment sampling
Our post-cut sampling was conducted from late May until August 2019. First, we updated the pre-harvest data in the main plot for all trees with a DBH larger than 9 cm, with the identification of live and dead trees as well as stumps to confirm removal and obtain the post-treatment basal area. To assess the regeneration, we reproduced the same regeneration sampling design as in the Outaouais region and maintained the same method of distributing subplots in a cross pattern instead of in a line as in the Laurentides (Figure1.2). In all cases, it was impossible to make sure the regeneration subplots were located at the exact same position as in the pre-harvest assessment. A member of the fieldwork team estimated canopy opening using a forest densiometer calculating an average of five measurements for each subplot, at a height of 1.3 m. Also, we identified whether the subplot was located in a strip or a skid-trail.
. The seedlings were divided into two height classes i.e. small seedlings from 15 to 40 cm in height and tall seedlings from 40 cm in height to a DBH of 1 cm. We also tallied saplings with their DBH (DBH > 1 to 10 cm). The presence of each species for each height class was recorded in each subplot. The species that would most likely dominate the seedling class and the subplot class was noted separately for each subplot. We defined dominance as the tallest vigorous stem in its respective class. If equally dominant, up to three species were identified in this assessment. Finally, we identified the competition cover (0, >0-25%, 25-75 %, 75-100%) for each regeneration height class. We considered as competition vegetation any non-merchantable species that had reached at least 15 cm in height. The competition cover classes were cumulative i.e. the cover of saplings considered as competition (e.g. stripped maple (Acer pensylvanicum)) would affect all height classes including those below. Other examples of competition species were Rubus idaeus, Sambucus pubens, Viburnum lantanoides and Dryopteris carthusiana. In three of the five subplots (north, south and east), we counted non-browsed seedlings per height class and per species. We considered a seedling to be browsed if it had been damaged over the last 3 years, as indicated by the subsequent primary growth of the seedling. In subplots with extreme seedling densities, from one-fourth to three-fourths of the subplot was sampled for efficiency, and estimates were scaled up to that of the entire plot. We measured the annual height growth of seedlings for the previous three growing seasons, using scale scars. For coniferous species, except eastern hemlock and eastern white cedar (Thuja occidentalis), we used the annual whorls of branches. We randomly chose two samples per species per height class and per subplot for time for time efficiency purpose. Again, we did not consider browsed seedlings because we wanted three full years of growth. We randomly chose two samples per species per height class and per subplot to measure the annual height growth. Again, we did not consider browsed seedlings because we wanted three full years of growth. Growth of the current year was not considered since the assessments were made at different times throughout the summer.
1.4.4 Data analysis
Annual height increment
To avoid limiting our assessment to current conditions and to take into account the dynamic nature of regeneration establishment and growth, we used the concept of transition probabi-lities first proposed by Kneeshaw and Bergeron (1998) and later applied by Hill et al. (2005) and Guay-Picard et al. (2015). The first step of this method consists of predicting the height growth of seedlings. For this, we produced an annual height increment linear mixed-effects mo-del for the five most abundant species in our study sites i.e. AS, FG, yellow birch, American hop-hornbeam and red maple, as well as for all the other species pooled together. These groups were included as a six-level categorical variable in the model. Only the non-browsed seedlings were considered. The model was fitted using the nlme package (Pinheiro and Bates,2011) in
the R statistical programming environment (R Core Team, 2018). To limit a potential bias attributable to the climate-induced variability in height growth, our dependent variable was the mean height increment of the last three years. For this, we only used the data from sites that were harvested in 2013 and 2014 (24 subplots from eight felling sites) to average three full years in post-treatment conditions only. Site, plot and subplot levels were considered as nested random effects. A set of seven candidate explanatory variables was defined a priori, na-mely species, seedling height, subplot location (strip or trail-located), subplot canopy opening, subplot competition cover class, subplot sapling basal area and subplot seedling abundance of all species except that of the subject. We also included three possible interactions in the full model (species : seedling initial height, canopy opening : subplot location and canopy opening : competition class). The initial height of the seedling was selected as a covariate for its known effect on seedling growth (Guay-Picard et al.,2015). The other variables were linked to understory resource availability, which can also influence seedling growth (Beaudet et al.,
2007; Canham, 1988; Hannah, 1991). The best model was selected based on the results of likelihood-ratio tests for nested models (Pinheiro and Bates,2011). For all analyses, residuals were analyzed to ensure that statistical assumptions were met. When outliers were identified, the results of the analysis were examined without them to ensure that any outliers were not a large source of bias. All outliers were maintained.
Transition probability indices
Instead of giving a static picture in time, transition probabilities portray regeneration using the current abundance and growth rate of each species (Guay-Picard et al.,2015;Hill et al.,2005;
Kneeshaw and Bergeron, 1998). Transition probabilities are in fact indices of the projected relative abundance, in this case expressed at the subplot level. The method provides indices of forest succession, in this case in the regeneration strata, but does not take into account mortality and recruitment after data sampling. Using height growth predictions from the model described above, we used the relative time to reach 2 m to weight the seedling density in a given height class. For this, we predicted annual height increments for the mean initial height of each height class (27.5 cm for small seedlings and 120 cm for tall seedlings) of each species which was present in any given subplot. We then integrated the annual growth increment function to obtain the predicted time for the seedling to reach 2 m and normalized the data by calculating the ratio of time to reach 2 m for a given species in a given height class to that of the slowest growing species. The reference of 2 m was chosen as an indication of regeneration reaching the sapling stage. Transition probability indices were then calculated using : Pt= P2 i=1(RAi/∆Ti) Pn t=1 P2 i=1(RAi/∆Ti)n (1.1)
where Pt is the transition probability index for a given species t (t = 1,2, . . .,n), RAi is the
relative abundance of species t in height class i(i = 1 (small seedlings), 2 (tall seedlings)) and ∆Ti is the relative time necessary to reach 2 m for species t in height class i. The
sum of transition probability indices within a subplot is 1. For a given species, transition probability indices can therefore vary between 0 and 1 with several occurrences of 0 and 1 values. Accordingly, transition probability indices were modelled using a zero-one inflated beta (BEINF) distribution. We used the gamlss package (Rigby and Stasinopoulos, 2005;
Stasinopoulos et al., 2017) in the R statistical programming environment (R Core Team,
2018) to run the analyses. As explained by Durocher et al. (2019), the BEINF() function defines the beta inflated distribution with a four-parameter distribution where µ describes the vector of location parameter values >0 and <1, σ the vector of scale parameter values >0 and <1, ν the vector of parameter values modelling the probability at zero and τ the vector of parameter values modelling the probability at one.
We first fitted a full model to understand which factors related to silviculture could better explain the transition probability indices obtained for AS. Priority was given to candidate variables that were easily measurable in the field. We selected the pre-treatment occurrence of AS in the advance seedling regeneration (separated into three ordered factors) as a covariate likely related to the transition probability values (Guay-Picard et al., 2015). Since they are manageable as part of commercial forest operations, we added variables linked to light availa-bility, that is, the percentage of basal area removal at the plot level and the canopy opening measured by the densiometer at the subplot level. We then added the pre-treatment basal area per hectare in FG as a candidate variable since mature FG trees tend to produce root suckers and clump shoots when cut (Burns et al.,1990).We added the calendar year of the first growing season after treatment as a candidate variable since it can be related to factors such as climatic conditions, forest tent caterpillar defoliation or seed abundance conditions. Also, because the application of a silvicultural treatment can vary between locations and operators, and because the silvicultural prescription was likely related to the pre-harvest composition and state of the regeneration, we chose not to include the prescribed type of partial cut among the explanatory variables. To reflect the structure of the variance, we aimed to insert nested random effects of the site and the plot to the four components of the model (µ, σ, ν and τ),but the full model had to be simplified to allow fitting. First, only the random effects of site were fitted in the full model for components µ, ν and τ, and then the nested subplot effect was added at the end of the variable selection process. Pearson’s r coefficients between candidate variables were consistently below |0.4|. We based our model selection method on Stasinopou-los et al. (2017) and used a backward elimination by AIC with the gamlss package. We first applied the backward selection to the full model for τ. Then, using the model obtained for τ, we applied the same backward selection process to the ν mode, and successively applied the same process for σ and µ. Statistical assumptions related to residuals for GAMLSS such as homogeneity of variance, normality skewness and kurtosis were checked. Using the final
fitted model, we predicted transition probability indices from two simulated databases. For this, we created classes representative of the observed variability for each variable except basal area removal and canopy opening, which were correlated variables. In theses cases, we created classes for one while using the mean value of the other for each class.
1.5
Results
1.5.1 Pre- and post-harvest attributes
Prior to treatment, the surveyed plots were mainly composed of AS. The mean initial basal areas of AS and FG were 14.9 and 4.2 m2ha-1, respectively. The initial basal area of FG in our
plots varied from no FG stems in the plots to a maximum of 22m2ha-1. The applied partial cuts
reduced stand basal area by 40% on average. Among plots, however, basal area removal varied from 0 to 80%. From an initial mean basal area of 24.8 m2ha-1, the average basal area of the
plots was reduced to 14.7 m2ha-1after treatment. At the time of our sampling, canopy opening
at the subplot scale ranged from 0 to 69% with an average of 12.4%. On average, subplots located in a strip had a canopy opening of 8% (SD=10%) whereas those in a skid-trail had an opening of 22% (SD=15%). There was a high variability among plots and subplots in terms of canopy opening, proportion of basal area removed and final basal area not only between the types of partial cuts, but also between plots with each type of cut (Figure 1.3).
Figure 1.3 – Attributes related to understory resource availability per treatment. STS : Single-tree selection cut, CIS : continuous-cover irregular shelterwood cut, EIS : extended irregular shelterwood cut. For subplot canopy opening, difference between strip (green solid line) and skid-trail (black dotted line) is represented.
1.5.2 Regeneration characterization
Occurrence, abundance and dominance
Prior to harvesting, the mean occurrence of species in the advance seedling regeneration of the surveyed subplots (as provided, combining small and tall seedlings) was 61% (SD= 39%) for AS compared to 40% (SD= 41%) for FG. Three to six years after harvesting, AS had maintained a higher mean occurrence than FG in the small seedling class with 77% (SD = 31%) compared to 56% (SD = 37%), respectively. However, in the tall seedling class the occurrence was similar between species, with 61% (SD = 39%) compared to 64% (SD = 37%) for AS and FG, respectively. The most abundant species within the small seedling class was AS with mean densities of approximately 12 000 stems ha-1(Figure 1.4). The abundance of
AS decreased sharply in the tall seedling class, so that FG became the most abundant species with densities averaging 3200 stems ha-1. FG saplings were also the most abundant. FG was
the most common dominant species in the two seedling classes combined as it dominated 232 of the 480 subplots sampled (48%). FG saplings also dominated 40% of the subplots. By comparison, AS seedlings and saplings dominated only 25% and 19% of the measured subplots, respectively.
Figure 1.4 – Abundance (mean + SE) and dominance of regeneration. Dominance was obser-ved for seedlings (two classes combined) and for saplings and is defined as the tallest vigorous seedling. AR.= Acer rubrum, AS.= Acer saccharum, BA= Betula alleghaniensis, FG. = Fagus grandifolia, OV= Ostrya virginiana. Seedl. =seedlings, Sapl.= Saplings
Growth and relative growth
The final annual growth model included the subplot location (strip or skid-trail), the canopy opening, the species, the seedling height and the interaction of the latter two as explanatory variables. Small AS seedlings located in strips 0% canopy opening were predicted to take the longest time to reach the 2 m threshold (over 36 years).We considered these seedlings as the reference to calculate the relative growth for seedlings in all other conditions. In comparison, the fastest growing seedlings were predicted to be tall yellow-birch seedlings located in a skid-trail with almost 70% of canopy opening that were predicted to reach the 2 m threshold in approximately 1.6 years (Figure 1.5). Moreover, the fastest AS seedling was a tall seedling located in a skid-trail with a subplot canopy opening of just over 50% that was predicted to reach 2 m in 2.7 years. In comparison, the slowest FG seedling was a small seedling in a strip with 0% canopy opening that was predicted to reach 2 m in 31 years. The fastest FG seedling was a tall seedling in a skid-trail with 64% canopy opening for which the anticipated time to reach 2 m was 2.1 years.
Figure 1.5 – Predicted time to reach two meters in height as a function of measured subplot canopy for five hardwood species and the other species pooled together, for two height classes (small and tall seedlings) and for two subplot locations (skid-trail or strip). Tall seedlings are all considered 120 cm high and small seedlings 27.5 cm high. AR.= Acer rubrum, AS.= Acer saccharum, BA= Betula alleghaniensis, FG. = Fagus grandifolia, OV= Ostrya virginiana. Seedl. =seedlings, Sapl.= Saplings
Transition probabilities indices
Despite not being the dominant species in the main canopy, FG had the highest mean transition probability indices after partial harvesting in our study area (Figure 1.6). AS had the second highest mean and all other groups had much lower results with a median index at zero in all cases. However, a high level of variation was observed among the sites, plots and subplots.
Figure 1.6 – Distribution of transition probability indices per species. Transition probability indices combine relative abundance and relative growth of every species in a subplot. The sum of transition probability indices within a subplot is 1. For a given species, the transition probability index can therefore vary between 0 and 1 with several occurrences of 0 and 1 values. AR.= Acer rubrum, AS.= Acer saccharum, BA= Betula alleghaniensis, FG. = Fagus grandifolia, OV= Ostrya virginiana.
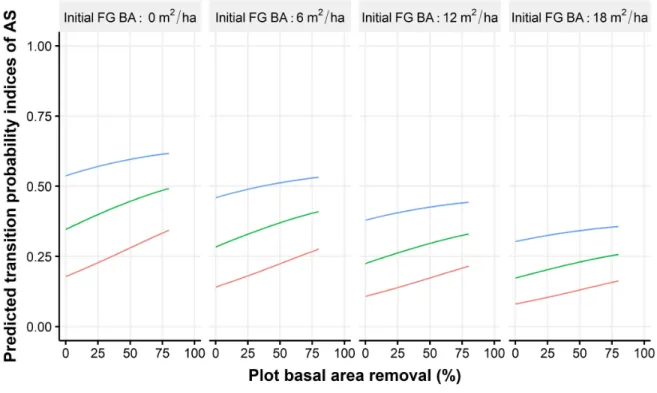
Five explanatory variables and two nested random effects contributed to the most parsimo-nious model for AS transition probability indices (Table 1.1). Based on this model, transition probability indices for AS tended to increase with the pre-treatment occurrence of the species in the advance seedling regeneration, while indices decreased with an increasing initial basal area in FG. An increase in the proportion of the initial basal area removed (intensity harvest) was associated with a lower probability of a ‘zero transition probability’ for AS. Predictions combining the zero and non-zero components of the gamlss fitted model are shown in figures
area removal although the effect was reduced when FG initial basal area was high. An increase in the occurrence of AS among the advance seedling regeneration (i.e., prior to treatment) also favoured AS transition probabilities indices significantly. However, the impact of basal area removal appeared to decrease when the occurrence of AS in the advance seedling regeneration was high. Overall, the difference in transition probability indices for AS between a plot basal area removal of 80% and 25% (maximum and minimum subplot intensity harvest) reached a maximum of around 0.1 when the initial basal area in FG was zero and the occurrence of AS in the advance seedling regeneration was 60% or less. The effect of subplot canopy opening on the transition probability indices of AS was less pronounced than that of basal area removal at the plot level. Moreover, the effect tended to become saturated at a subplot canopy opening of approximately 50%.
Table 1.1 – Details of the fitted gamlss model.Final models for each species group. µ describes the vector of location parameter values >0 and <1, σ the vector of scale parameter values >0 and <1, ν the vector of parameter values modelling the probability at zero and τ the vector of parameter values modelling the probability at one. FG IBA : FG Initial basal area per hectare, FGY : Year of the first growth season after harvest
Species Model DF AIC Component Explanatory variables Full model ∆ AIC
AS 1 117 199.9 µ initial AS occurrence + FG IBA+
Site/Plot nested random effects 116.68
σ FGY + FG IBA
ν Canopy opening+ Proportion of to-tal IBA removed + FG IBA+ Site/Plot nested random effects τ Canopy opening+ Proportion of
Figure 1.7 – Predictions of transition probability indices for AS as a function of plot basal area removal per class of AS pre-treatment occurrence in the advance growth and pre-treatment FG basal area (FG BA).Because we predicted transition probabilities from a simulated database, no confidence intervals are presented. We divided the observed range of basal area removal (0 to 80%) into four sections and calculated the mean canopy opening for each. For a simulated basal area of 0 to 20%, the mean canopy opening was 8%. For a simulated basal area of 20 to 40%, 40 to 60% and 60 to 80%, the mean canopy opening was 10%, 15% and 27%, respectively. For every jump of 1% in basal area removal, we generated all possible combinations with the other variables.
1.6
Discussion
1.6.1 AS-FG regeneration dynamics
Despite the dominance of AS in the pre- and post-harvest overstory and its high abundance of small seedlings, AS seedlings were dominated by FG overall after harvest over the full range of harvesting conditions investigated, and this trend became progressively stronger as regeneration size increased. FG seedlings dominated almost 50% of the sampled subplots and had the highest mean transition probability indices among all species. As observed by Elenitsky et al. (2020) after beech bark disease-motivated harvests in northern Michigan, AS regeneration was less dominant than that found prior to harvest. Such a dynamic occurring in northern hardwood forests has been well documented both for old-growth and managed
Figure 1.8 – Predictions of transition probability indices for AS as a function of subplot canopy opening per class of AS pre-treatment occurrence in the advance growth and initial FG basal area (FG BA). Because we predicted transition probabilities from a simulated database, no confidence intervals are presented. We divided the observed range of subplot canopy opening (0 to 70%) into four sections and calculated the mean basal area removal for each of them. For a simulated subplot canopy opening of 0 to 17.5%, the mean basal area removal used was 35%. For a simulated subplot canopy opening of 17.5 to 35%, 35 to 52.5% and 52.5 to 70%, the mean basal area removal was 45%, 53% and 54%, respectively. For every jump of 1% in subplot canopy opening, we generated all possible combinations with the other variables.
stands (Bose et al.,2017;Duchesne et al.,2005) . Based on this result onlystudy, and as also that observed by Nolet et al. (2008), increasing the proportion of harvest in partial cuts in a commercial context appears to be generally insufficient to favour the regeneration of AS over FG. . While the ecological niche of AS and FG is very similar, FG seems to have better mechanisms to compete with AS, including an exceptional ability to produce root suckers (Beaudet et al.,1999).
However, looking more closely at our results, we found that a higher basal area removal can have a small but significant effect on the likelihood of a transition towards AS-dominated regeneration in certain conditions. Indeed, the observed shift of about 0.1 in predicted tran-sition probability indices for AS between a low (25%) or high (80%) basal area removal is
non-negligible (Figure 1.7), considering that a median value of only 0.25 makes FG the spe-cies with the highest transition probability indices overall (Figure 1.7). Since species-specific transition probability indices are relative to each other in the same subplot (the sum of all species-specific transition probability indices equals one), an increase in the transition proba-bility index for AS necessarily implies a decrease in other species, and vice and versa. We can therefore speculate that FG, the most abundant species, is most disadvantaged by increases in transition probability indices for AS. Moreover, in the range of 0 to 80% of plot basal area removal, AS transition probability indices tended to increase without reaching a plateau. This trend is in line with the observations of Beaudet et al. (2011; 1999) and Poulson and Platt (1996) who suggested that using more severe disturbances or more spatially heterogeneous systems than single-tree selection cuts could help favour AS dominance over FG.
Two factors appear to influence the response of AS to an increase in basal area removal. First, when the initial occurrence of AS in the advance seedling regeneration is high (e.g., occurrence >= 80%), opening the canopy does not have a great impact on AS dominance. In such situa-tions, AS transition probability indices tended to remain high (more than 0.5) regardless of the harvest proportion, especially when the initial FG basal area was low. This supports the finding of Forrester et al. (2014) that when AS advance regeneration is predominant before canopy opening, it is likely to remain so after harvest. Secondly, when the initial FG basal area is high, increasing basal area removal is generally not sufficient to contain the prevalence of FG in the regeneration. The effect of basal area removal (i.e., the slope of the predictions between a basal areal removal of 0% and 80% in Figure1.7) remained low in such conditions regardless of the occurrence of AS in the advance seedling regeneration. The ability of FG to develop fast growing root suckers is a key factor leading to its dominance in regeneration (Forrester et al.,2014;Jones and Raynal,1988). When the initial FG basal area is high, increasing light availability using any variant of a partial cut treatment is clearly not sufficient to reverse the dominance of the species.
Our results showed a more limited effect of subplot canopy opening than of basal area removal, especially when the initial FG basal area was high. This difference might be explained by the resolution of the measurement. Basal area removal was measured in the centre of the plot while canopy opening was measured more locally with assessments made in each subplot. Because of the spatial variation in canopy removal after a partial cut (Forget et al.,2007;Guay-Picard et al.,2015;Moreau et al.,2020), increasing basal area removal will also tend to increase the proportion of subplots in which there is a significant increase in canopy opening. At this more local scale, it is possible that the beneficial effect of the opening for the regeneration success of AS tends to become saturated at high levels of canopy opening. While our results show a saturation of the effect at approximately 50% of canopy opening, it is likely that the post-cut threshold would in fact be higher because of the timing of the measurement. While basal area removal was assessed immediately after treatment, canopy opening measurements were made
only three to six years after treatment. Light availability, especially for seedlings, is known to decrease rapidly after selection cutting in northern hardwood forests (Beaudet et al., 2004). Further research would therefore be required to identify the level of local canopy opening at which the benefits to AS regeneration tend to saturate.
All treatments combined, the mean proportion of basal area removal was 40% compared to a subplot canopy opening of 12%. The two measures portrayed a similar trend for the three variants of partial harvesting. In ascending order, single-tree selection cuts, continuous-cover irregular shelterwood cuts and extended irregular shelterwood cuts generated the expected ascending gradient in light availability. However, we found highly variable light conditions within each type of cut. Forget et al. (2007) also observed high within-stand variability in commercial selection cuts. Part of this variability is attributable to whether a subplot is located in a strip or a skid-trail. Especially when basal area removal is low, as confirmed by Moreau et al. (2020),a commercial cut will tend to concentrate the canopy opening in the skid trails. . A high variability is desirable when the objective is to establish a high diversity of species (Raymond et al.,2018;Raymond and Bédard,2017), but it can be problematic when several subplots are unaltered by the cut. In our study, large untreated areas were observed in low intensity cuts and several subplots had close-to-zero canopy opening. Moreau et al. (2020) found that 42% of the overstory trees remaining after single tree selection cuts did not experience any change in their competitive environment within a 6-meter radius.
In practice, controlling basal area removal is much easier than attempting to control canopy opening to an optimal level throughout the stand. One potential drawback of using high inten-sity partial cuts is that it may favour the proliferation of competition in subplots subjected to almost fully open conditions. Non-tree vegetation competition increases with canopy opening which can, in turn, limit light and other resource availability for regeneration development (Houston, 2001; Matonis et al.,2011). Surprisingly, the AIC selection rejected this factor in our height growth model for seedlings. Estimating competition cover in classes (0, >0-25%, 25-75%, 75-100%) instead of estimating more precisely might have affected its relevance in the analysis.
1.6.2 Management implications
Our results showed that adjusting the prescriptions of commercial partial cuts in selection and irregular shelterwood silvicultural systems could help to favour AS regeneration only in a given range of conditions. Two pre-treatment measurements are needed to help inform this silvicultural decision : 1) the occurrence of AS in the advance seedling regeneration and 2) the pre-treatment basal area of FG. Based on our predicted probability indices model, we recommend an initial FG basal area threshold of ∼ 6m2ha-1,as this was the highest value for
which an increase of basal area removal of up to 80% led to a predicted transition probability index of more than 0.25 for any occurrence level of AS in the advance seedling regeneration.
Below this threshold we observed the strongest effect of basal area removal, particularly when the occurrence of AS in the advance seedling regeneration subplots was below 60%. Thus, where the initial basal area of FG is lower than ∼ 6m2ha-1and the occurrence of AS in the
advance seedling regeneration is below 60%, a higher basal area removal might be considered as an efficient tool to favour AS dominance among seedlings. Prescribing a low mean basal area removal per cutting site (i.e,. less than 25-30%) is unlikely to favour the success of AS regeneration. Considering our canopy opening results, such low intensity cuts will lead to overly large proportions of the cutting site remaining unaltered, and FG will tend to be favoured in such conditions.
Increasing light availability may be achieved by either increasing mean basal area removal or ensuring homogenous harvesting across the site. Due to the reality of mechanized operations, the solution of increasing basal area removal is a simple, more practical way to make sure that an increased proportion of subplots experience a canopy opening that will favour AS. However, a higher removal should ideally not be achieved by creating more skid-trails since machinery passage damages the roots and soil (Hartmann et al., 2009). Bose et al. (2020) observed an increased mortality of advance regeneration after commercial partial harvesting operations in which more than 40% of the basal area was removed. With higher harvesting rates (35% and above), harvester operators have more flexibility to reach stems into the strips. The use of secondary trails, where only harvesters go, were suggested by Moreau et al. (2020) since they cause less damage to the remaining trees and soil. In systems where skid-trails will most likely be reused during the next harvest, such secondary trails might facilitate the protection of AS regeneration. Directional felling and skid trail layouts have also been reported to provide efficient protection measures for advance regeneration, roots and soil (Bose et al.,
2020;Hartmann et al.,2009;Naghdi et al.,2016). In addition, parameters such as trail width and spacing can be planned to monitor the distribution of residual trees (Hartmann et al.,
2009). Improvement could be achieved with greater effort invested in tree marking, creating more heterogeneity in gap size, and creating targeted, larger openings where AS seedlings undergo strong competition with FG.
Where AS occurrence in the advance seedling regeneration is high (more than 80%) and FG basal area is low (less than ∼ 6m2ha-1), there is no need to increase harvest intensity. On
a short-term basis at least, the dominance of AS regeneration and a good transition from seedling to sapling should be observed in such conditions, regardless of basal area removal. Close-to-nature partial cuts with low harvest intensity (Nolet et al., 2014), such as single-tree selection cuts (Lorimer, 1989; Tubbs, 1977), are recommended in this case. Where the initial basal area of FG is high (more than ∼ 6m2ha-1), our results suggest that increasing
the harvest intensity in partial cuts will likely not suffice to favour AS dominance. However, while recognizing the importance of protecting FG trees resistant to the beech bark disease for the species’ future (Houston,2001), our results confirm the need to maintain low levels of FG
basal area. Otherwise, the feedforward mechanism, whereby high FG basal area leads to higher dominance in regeneration, is likely to be maintained (Giencke et al., 2014). As a solution, Nolet et al. (2015), Nyland et al. (2006) and Bose et al. (2018) recommended a combination of harvesting operations to suppress FG advance regeneration. The application of clearcuts was also suggested as a solution to decrease FG dominance (Nolet et al., 2008, 2015). Over the next cutting cycle, the combined effects of a high basal area removal and advance seedling regeneration suppression might then be enough to tip the regeneration dynamics in favour of AS.
In short, efficiently addressing the increase of FG abundance in regeneration will require re-fining and enhancing commercial planning and harvesting in northern hardwoods (Messier et al., 2019; Puettmann et al., 2009). CCurrent knowledge on this issue clearly shows that there is no single, wall-to-wall solution that can be efficiently applied but rather a diversity of targeted measures that can be adapted to local conditions. In general, using only basal area removal to manage light availability for the understory is not sufficient to favour AS. Understory composition most be considered when planning commercial partial cuts and ma-naging this understory is as important. The development of tools for multitreatment planning, such as that proposed by Lussier and Meek (2014), may help implement complex silvicultural treatments within a single stand.
The recommendations made in this study are based on empirical evidence. Caution must be used in applying them in the context of the increasing effects of global changes on forests. Such changing conditions may alter the outcome of silvicultural measures applied to the same forest type in the future. In our analysis, the calendar year of the first growing season after treatment was retained as a variable influencing AS transition probabilities. This implies that results may vary according to changes in yearly conditions that currently remain unquantified. Further research is required to elucidate such effects so that the response of the forest to future partial cuts can be better anticipated. Moreover, increasing AS seedling transition to sapling does not necessarily imply an increase in AS sapling to pole transition. Hence, a meticulous monitoring followed by FG sapling suppression, if required, should be considered to ensure a successful transition from saplings to poles for AS. Also, other issues than FG competition must be considered when making silvicultural prescriptions that promote the future health, quality and resilience of northern hardwood forests. We should not manage forests only for AS, but for a greater species and structural diversity to ensure that forests will maintain their resistance and resilience in a rapidly changing socio-environmental context (Messier et al.,