Optimisation des conditions de fermentation et de
stabilisation pour la production de bio-ingrédients
fonctionnels à base de
Carnobacterium divergens
M35
Mémoire
Laurent Dallaire
Maîtrise en sciences et technologie des aliments - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
Optimisation des conditions de fermentation et de stabilisation pour la
production de bio-ingrédients fonctionnels à base de Carnobacterium
divergens M35
Mémoire
Laurent Dallaire
Sous la direction de :
Ismail Fliss, directeur de recherche
Denis Groleau, codirecteur de recherche
iii
Résumé du mémoire
Lors de travaux antérieurs, un bio-ingrédient permettant la bioconservation du saumon fumé à froid et ayant une forte activité anti-Listeria fut développé et caractérisé. Il consiste en un milieu de culture fermenté par C. divergens M35 et contenant la bactériocine produite par la souche, soit la divergicine M35. Par contre, les conditions de production actuelles ne permettent pas une utilisation efficace et rentable de ce bio-ingrédient. L’objectif de ce travail est de répondre à ces problématiques.
Dans un premier temps, un milieu de culture de grade alimentaire favorisant une forte et rapide croissance de C. divergens M35 et stimulant la production de la divergicine a été développé. Un criblage de différentes sources d’azote, de carbone et de sels a permis de déterminer que la mélasse de canne et le sucre de table (saccharose) sont les sources de carbone de choix, la source d’azote préférentielle reste l’extrait de levure et que l’acétate de sodium stimule la production en bactériocines.
Ce milieu fut testé en fermenteur de 30L afin d’évaluer l’effet de la mise à l’échelle. Le milieu créé permet d’atteindre une biomasse de 9,04 log(UFC/mL) et une activité de 1,3X105 AU/mL en 7h. Il s’agit d’une amélioration significative quant à la performance, mais aussi au coût du milieu de culture (0,89$/L) en comparaison à la référence, le milieu MRS (7$/L).
Dans un deuxième temps, il a été démontré que le séchage par atomisation est bien plus efficace que la lyophilisation afin de produire un bio-ingrédient biologiquement stable. Le séchage par atomisation permet d’obtenir un bio-ingrédient sec possédant une viabilité de 9,85 log(UFC/g) et une activité anti-Listeria de 1,6X106 AU/g. Ce procédé permet d’avoir une production rentable du bio-ingrédient M35.
iv
Table des matières
Résumé du mémoire ... iii
Table des matières ... iv
Liste des tableaux ... vii
Liste des figures ... viii
Remerciements ... ix
Avant-propos ... x
Introduction générale ... 1
Chapitre 1 : Revue de la littérature ... 3
1.1 Conservation de la qualité et de l’innocuité des aliments ... 4
1.1.1 Pertes alimentaires dues aux altérations ... 4
1.1.2 Contaminations microbiologiques ... 4
1.1.3 Méthodes de conservation traditionnelle ... 6
1.2 Nouvelles tendances dans la conservation des aliments ... 11
1.2.1 Bioconservation et cultures protectrices ... 11
1.2.2 Bactériocines ... 12
1.2.3 Les bactériocines comme nouvel additif pour la conservation des aliments .. 13
1.3 Carnobacterium divergens M35 ... 15
1.3.1 Bio-ingrédient anti-Listeria à base de M35 ... 15
1.3.1.1 Milieux de culture ... 17
1.3.1.2 Stabilisation du bio-ingrédient ... 21
1.3.1.3 Mise à l’échelle ... 24
Hypothèse ... 26
Objectifs ... 26
Chapitre 2 : Optimisation d’un milieu de culture et des conditions d’opération pour la croissance de Carnobacterium divergens M35 ... 27
2.1 Résumé ... 28
2.2 Introduction ... 30
2.3 Matériel et Méthodes ... 32
2.3.1 Souches bactériennes et milieux de culture ... 32
v
2.3.3 Mesure de l’activité anti-Listeria ... 32
2.3.4 Criblage des composants du milieu de culture ... 33
2.3.5 Effet des conditions de culture ... 35
2.3.6 Optimisation des concentrations par la MSR ... 35
2.3.7 Effet de la co-culture entre C. divergens M35 et C. maltaromaticum CB1 ... 36
2.3.8 Effet du stress oxydatif sur l’activité de la divergicine M35 ... 37
2.3.9 Fermentations en bioréacteur de 30L ... 38
2.4 Résultats et discussion ... 39
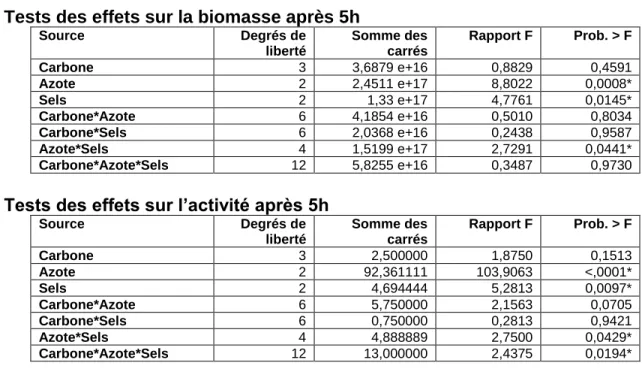
2.4.1 Criblage des composants solubles du milieu de culture ... 39
2.4.2 Criblage des composants non solubles du milieu de culture ... 44
2.4.3 Détermination du milieu de culture ... 49
2.4.4 Effets des conditions de culture ... 54
2.4.5 Optimisation de la concentration des composants ... 58
2.4.6 L’effet de la mélasse, du sucre de table et du milieu minimum ... 61
2.4.7 Co-culture M35 et CB1 ... 66
2.4.8 Résistance à l’oxydation de la divergicine M35 ... 69
2.4.9 Mise à l’échelle du procédé de fermentation ... 71
Chapitre 3 : Stabilisation du bio-ingrédient à base de C. divergens M35 et de sa bactériocine par atomisation ... 76
3.1 Résumé ... 77
3.2 Introduction ... 78
3.3 Matériel et Méthode ... 80
3.3.1 Souches bactériennes et conditions de culture ... 80
3.3.2 Conditions d’utilisation du séchoir atomisateur et MSR ... 80
3.3.3 Calcul des rendements (massique, biomasse et activité) ... 81
3.3.4 Dénombrement de C. divergens ... 82
3.3.5 Mesure de l’activité anti-Listeria ... 82
3.4 Résultats et discussion ... 83
3.4.1 Choix de l’agent aidant au séchage ... 83
3.4.2 Analyse de la variance ... 85
vi
3.4.4 Optimisation des conditions ... 92
3.4.5 Caractéristiques du bio-ingrédient séché ... 94
3.4.6 Application sur le saumon fumé à froid ... 96
Conclusion générale et perspectives ... 97
vii
Liste des tableaux
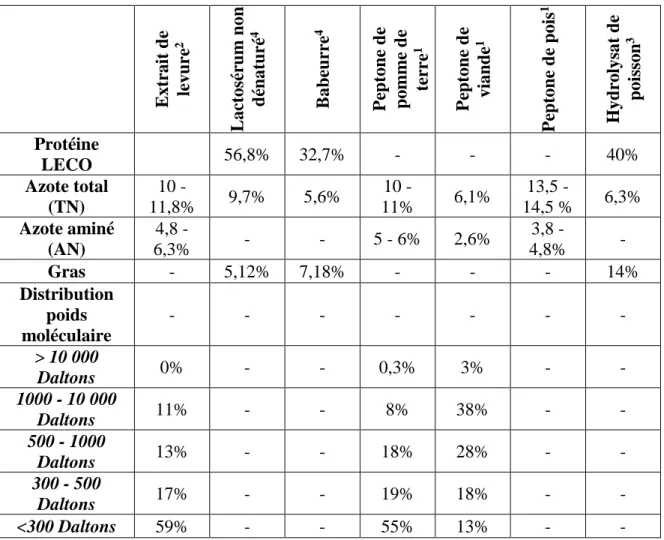
Tableau 1 : Constituant des diverses sources d'azotes ... 20
Tableau 2: Plan factoriel pour le criblage des composants solubles ... 34
Tableau 3: Plan factoriel pour le criblage des composés non solubles ... 34
Tableau 4: Plan expérimental de co-culture ... 37
Tableau 5: Critères de référence pour le criblage des composés solubles... 39
Tableau 6: Effets statistiques des composants solubles sur le µmax, le temps pour l'atteindre et l'activité anti-Listeria ... 41
Tableau 7: Critères de référence pour les composants non solubles ... 45
Tableau 8: Effets statistiques des composants non solubles sur la biomasse et l'activité .... 47
Tableau 9: Comparaison des conditions d'opération des bioréacteurs d'1L ... 57
Tableau 10: Estimations des coefficients triés pour la biomasse ... 59
Tableau 11: Estimations des coefficients triés pour l'activité en microplaque ... 59
Tableau 12: Comparaison économique des différents milieux de culture... 65
Tableau 13: Influence de la co-culture M35 et CB1 sur la biomasse et l'activité obtenue après 18h ... 68
Tableau 14: Évaluation de la stabilité de l'activité de la divergicine M35 lors de l'entreposage ... 70
Tableau 15: Composition finale des milieux STYA et MYA ... 72
Tableau 16: Analyse de la variance de l'optimisation de l'atomisation ... 85
Tableau 17: Analyse de la variance de la température et de la concentration en maltodextrine ... 86
viii
Liste des figures
Figure 1 : Causes microbiologiques de rappels alimentaires en 2016 au Canada et aux
États-Unis [1] ... 5
Figure 2: Principe des barrières en conservation alimentaire (Leistner et al. 1995) ... 8
Figure 3: Croissance des microorganismes sur une matrice de poisson lors de sa conservation (Gram et Huss, 1996) ... 10
Figure 4 : Schéma du séchoir atomisateur ... 23
Figure 5: Plan Box-Behnken pour l'optimisation des concentrations ... 36
Figure 6: Courbe de croissance de M35 en microplaque dans le milieu MRS ... 39
Figure 7: Profileur de prédiction pour le criblage des composants solubles ... 43
Figure 8: Culture statique de M35 dans MRS dans un volume de 200mL ... 45
Figure 9: Profileur de prévision pour le criblage des composants non solubles ... 48
Figure 10: Détermination des composants du milieu de culture STYA ... 51
Figure 11: Coût du milieu de culture en fonction du choix de composants ... 53
Figure 12: Établissement des conditions de culture optimale en fermenteur d'1L ... 56
Figure 13: Profileur de prévisibilité pour les concentrations du milieu de culture ... 60
Figure 14: Courbe de croissance du milieu à base de mélasse (MYA) ... 62
Figure 15: Effet de la mélasse, du sucre de table et du milieu minimum (MM) ... 64
Figure 16: Fermentation des milieux STYA et MYA en fermenteurs de 30L ... 72
Figure 17: Effet de la mise à l'échelle à l'étape de fermentation ... 74
Figure 18: Atomisation sans ajout d'additifs - caramélisation du bio-ingrédient entraînant une forte adhérence sur la paroi du cyclone ... 84
Figure 19: Atomisation avec ajout de maltodextrine - très faible adhérence du bio-ingrédient sur la paroi du cyclone ... 84
Figure 20: Surface de réponse pour le rendement massique ... 88
Figure 21: Surface de réponse du rendement en biomasse ... 91
Figure 22: Surface de réponse du rendement en activité ... 91
ix
Remerciements
J’aimerais d’abord remercier sincèrement mon directeur de recherche, le Dr Ismail Fliss pour l’opportunité qu’il m’a offert de développer mes compétences dans le cadre de la chaire METABIOLAC. Je le remercie pour la confiance qu’il a eu en moi et pour la pleine liberté qu’il m’a donné dans l’orientation de mes travaux. Vous m’avez offert tous les outils afin que je puisse m’épanouir et je suis très heureux d’avoir participé à l’application industrielle de vos nombreuses années de travail.
J’aimerais aussi remercier mon codirecteur de recherche Dr Denis Groleau qui est la référence par excellence dans le domaine des biotechnologies au Québec. Vos nombreux conseils ont toujours permis de bien enligner mes recherches et votre enthousiasme pour la recherche était des plus contagieux.
Je tiens aussi à remercier Dr Allison Vimont et Dr Benoit Fernandez qui ont énormément participé à l’élaboration de ce projet. Les nombreuses séances de remue-méninges que nous avons eu ont permis de faire avancer exponentiellement ce projet. Je vous remercie aussi pour le temps que vous avez dédié à ce projet et à répondre à mes très nombreuses questions!
Je veux aussi remercier les technicien(nes) de laboratoire Marine Béguin et Jean-Nicolas Ouellet de l’Université Laval, ainsi qu’Isabelle Arsenault de l’Université Sherbrooke pour leur soutien durant mes travaux.
Je veux remercier tout le groupe d’étudiants de l’AGIDSA qui ont rendu mon expérience à l’Université Laval incroyablement enrichissant et divertissant.
Je veux finalement remercier ma copine Noémie de m’avoir épaulé depuis le début dans ce projet et d’avoir toujours été présente dans les hauts et les bas de cette maîtrise.
x
Avant-propos
Ce projet s’inscrit à la suite de nombreuses recherches qui ont été faites sur les bactériocines et leur application dans le domaine alimentaire afin de répondre à la demande en nouvelles techniques de conservation biologique. Dans ce cas, ces travaux s’adressent particulièrement à la souche C. divergens M35 dont la caractérisation et les études d’application sur des matrices alimentaires ont été accomplies à l’Université Laval en partenariat spécial avec Fumoir Grizzly de St-Augustin-de-Desmaures, QC, Canada. L’objectif final de ce projet est de faire la démonstration qu’une production abordable en bio-ingrédients à base de bactériocines est possible, et permettre son utilisation dans le domaine alimentaire.
Ces travaux ont été réalisés et financés par la chaire de recherche industrielle du CRSNG sur les activités métaboliques et la fonctionnalité des cultures lactiques bio-protectrices (METABIOLAC). Cette chaire regroupe plusieurs industries et partenaires dont le Consortium de recherche et innovations en bioprocédés industriels au Québec (CRIBIQ), l’Institut sur la nutrition et les aliments fonctionnels (INAF), le Centre de recherche en sciences et technologie du lait (STELA), Cascades, Groupe Sani Marc, Novalait, La Coop, Olymel, Biena et Fumoir Grizzly.
1
Introduction générale
Malgré les progrès dans le domaine de la transformation alimentaire, les contaminations microbiologiques restent choses courantes. Au Québec, les contaminations microbiologiques des produits Maple Leaf® et XL Food Inc. furent très médiatisés. Ajoutant à la difficulté de garantir l’innocuité microbiologique des aliments, les consommateurs sont de plus en plus à la recherche d’aliments sans agents de conservation chimiques. Il y a donc un besoin pour de nouvelles barrières biologiques permettant de répondre à ces nouvelles attentes. Plusieurs alternatives naturelles telles les cultures protectrices, les huiles essentielles et les bactériocines sont présentement étudiées et proposés comme nouvelles barrières de protection microbiologiques des aliments [2]. Les bactériocines sont des peptides (<90kDa) ayant une activité bactéricide ou bactériostatique très spécifique [3].
Les bactéries lactiques (LAB) sont de bons producteurs de bactériocines. Ces bactéries sont généralement reconnues comme sécuritaires (GRAS) par des organismes comme la FDA. Leur approbation devient ainsi plus aisée pour leur application alimentaire [4]. L’ajout de souches productrices de bactériocines dans différents produits alimentaires fermentés permettrait d’ailleurs d’éviter leur contamination par des bactéries d’altération [5]. Actuellement, seules deux bactériocines ont obtenu l’approbation de la FDA, à savoir la nisine (Nisaplin® par Danisco, Chrisin® par Chr. Hansen) pour le contrôle de Clostridium botulinum dans les fromages et la viande notamment [6] et la colicine E1 pour le contrôle d’Escherichia coli dans la viande de bœuf [7].
La divergicine M35 produite par Carnobacterium divergens M35 est une bactériocine qui suscite l’intérêt de l’industrie alimentaire québécoise pour son potentiel comme agent de conservation des aliments carnés et marins. De plus, son utilisation comme additif alimentaire est approuvée auprès de Santé Canada [8]. Malgré l’énorme potentiel que présentent les molécules de ce type, leur utilisation demeure encore très restreinte. Cette situation est attribuable au fait que les bactériocines sont produites en faible quantité (de l’ordre du mg L-1) par les bactéries lactiques. Le milieu de culture MRS est le milieu recommandé afin d’augmenter ces rendements [9], mais il est trop dispendieux pour un usage industriel [10]. Par ailleurs, le séchage par lyophilisation du milieu de culture fermenté
2
contenant la bactérie et sa bactériocine n’est industriellement pas approprié. Tous ces facteurs ne permettent donc pas d’obtenir un procédé de production de la divergicine M35 rentable. Plusieurs travaux ont démontré le potentiel d’optimisation de la production de différentes bactériocines afin d’atteindre le seuil de rentabilité. La première approche étudiée est la réduction du coût du milieu de culture. L’utilisation de sources de carbone et d’azote provenant de déchets de l’industrie alimentaire permettrait de réduire ces coûts [11]. Différents travaux ont d’ailleurs démontré l’obtention de rendements similaires au milieu MRS en utilisant différentes sources d’azote alternatives (ex : hydrolysats de poissons et peptones de patate) et de carbone (ex : mélasse de canne et extrait de malt) [12, 13].
Dans le cas d’utilisations alimentaires, la présence d’allergènes doit toujours être prise en compte [14]. Le deuxième défi rencontré lorsque l’utilisation des bactériocines dans les secteurs alimentaire et vétérinaire est envisagée est le conditionnement et la stabilisation physicochimique et biologique des préparations à base de bactériocines. Le séchage du bio-ingrédient produit et l’ajout d’agents protecteurs est nécessaire afin d’éviter sa dégradation [15]. Enfin, une mise à l’échelle du procédé est nécessaire afin de démontrer la faisabilité industrielle du procédé développé.
3
4
1.1
Conservation de la qualité et de l’innocuité des aliments
1.1.1 Pertes alimentaires dues aux altérationsEn 2010, au Canada seulement, il était estimé que le gaspillage alimentaire dû à la surproduction, aux défauts des aliments et à l’altération de ces aliments équivalait à 27 milliards de dollars canadiens [16]. Ce gaspillage nuit à la demande nutritive exponentielle de la population mondiale causée par l’augmentation de la population. Il est estimé que d’ici 2050, la population aura atteint 9,1 milliards d’êtres humains. Afin de subvenir à cette augmentation, l’offre alimentaire devra augmenter d’au moins 70% en comparaison à la production mondiale offerte en 2005/07 [17]. Afin d’atteindre cette demande, le domaine agroalimentaire devra être plus performant et efficace dans la production des denrées alimentaires, mais aussi réduire les pertes énormes qui nuisent à cette efficacité. Dans bien des cas, ces pertes sont causées par des méthodes de conservation, de transformation et d’entreposage inadéquates, réduisant considérablement la durée de vie de l’aliment [18].
Les trois grands types d’altérations des aliments sont l’altération physique, l’altération chimique et l’altération microbiologique [19]. La première altération est causée par un dommage à la structure de l’aliment qui peut nuire à son esthétisme, mais va aussi favoriser les deux autres types d’altérations. L’altération chimique est due à la modification du contenu protéique, en carbone et lipidique. Ce type d’altération va de pair avec l’activité de l’eau (aw) dans l’aliment qui favorisera l’oxydation des lipides, la réaction de Maillard, stimuler l’activité enzymatique et la croissance de microorganismes. Dans ce dernier cas, l’altération de l’aliment provient de la contamination microbienne de l’aliment. Il serait d’ailleurs estimé que 25% des pertes alimentaires seraient causées par des contaminations microbiologiques comprenant les bactéries, les levures et les moisissures [20].
1.1.2 Contaminations microbiologiques
Malgré les mesures d’hygiène de plus en plus strictes dans le domaine de la transformation alimentaire, les contaminations microbiologiques des aliments sont toujours fréquentes. Au Québec, les récents cas de contamination par Listeria monocytogenes de produits Maple Leaf® et par Escherichia coli dans la viande de bœuf XL Food Inc. démontrent que
5
l’industrie de la transformation alimentaire n’est toujours pas à l’abri des contaminations microbiologiques. Le rappel des produits contaminés est d’ailleurs très coûteux pour l’industrie alimentaire. Une étude conjointe du FMI et du GMA démontrait récemment qu’en moyenne aux États-Unis, le coût d’un rappel dans l’industrie alimentaire était de 10M$ [21]. En plus d’être un choc économique pour la compagnie victime du rappel, ce type de rappel peut même tuer une marque de commerce; le consommateur ayant perdu confiance envers le transformateur alimentaire. En 2016, le principal contaminant microbiologique fut Listeria. Au Canada et aux États-Unis, 196 rappels alimentaires par la FDA, la FSIS ainsi que l’ACIA ont été causés par une contamination à Listeria [1]. Au Canada seulement, il s’agit de 42 produits alimentaires différents qui ont été victime d’un rappel causé par Listeria monocytogenes dans la même année [22].
Mondialement, 600 millions de personnes seront victimes d’une intoxication alimentaire qui est, dans la majorité de cas, causée par une contamination microbienne. Parmi ces personnes, 420 000 vont succomber à la maladie qui peut être causée par Listeria, Campylobacter, E. coli, Salmonella ou par des norovirus [23]. Au Canada, annuellement, les contaminations microbiennes connues des aliments causent 4 millions de maladies, soit un canadien sur huit.
6
Ces maladies requièrent l’hospitalisation de 11 600 de ces malades et entraînent la mort de 238 d’entre eux. Parmi ces contaminations, les norovirus sont les plus communs en causant à eux seuls 1 million de cas rapportés de la maladie, mais Listeria reste la maladie la plus mortelle avec 35 cas de mortalité dû à l’ingestion alimentaire de ce pathogène [24].
Les produits marins sont particulièrement susceptibles à ce type de contamination microbiologique. La première source de contamination sont les eaux d’où proviennent le poisson. En Californie, Listeria est retrouvée dans 81% des eaux fraîches et dans 62% des eaux salées [25]. La transformation de ce type de produit inclut plusieurs étapes qui augmentent le risque de contamination. Que ce soit le filetage, le salage, le séchage, le fumage, le découpage et l’emballage, ce sont toutes des étapes de manipulation rajoutant au risque de contamination. Dans le cas du saumon fumé à froid, en Norvège, il a été déterminé que jusqu’à 10% des saumons commercialisés sont porteurs de Listeria. Cette contamination n’est pas nécessairement critique comme la concentration retrouvée en Listeria n’excède que rarement 100 UFC/g [26].
Cette fréquence de contamination peut être expliquée par le grand nombre de manipulations requises à la transformation, mais aussi par deux autres facteurs. Le premier facteur est la chair du saumon fumé qui est une excellente matrice pour le développement microbiologique. Avec une forte activité de l’eau (aw) entre 0,98 et 0,999, un pH presque neutre, entre 6,1 et 6,3, et un faible taux de sel entre 3 et 2%, le saumon est fortement propice à la croissance de Listeria [27]. Le deuxième facteur est l’absence de traitement thermique. Avant l’arrivée du fumage à froid, le traitement thermique imposé par le fumage à chaud permettait de réduire la charge microbienne de l’aliment. Avec le fumage à froid, il n’y a plus de traitement thermique permettant de réduire cette charge.
1.1.3 Méthodes de conservation traditionnelle
Actuellement, l’industrie de la transformation alimentaire se fie aux barrières physiques et au jumelage de celles-ci afin d’éviter les contaminations microbiennes. Ce concept est illustré par Leitsner et al [28] à la figure 2 qui propose la modulation des traitements de conservation, par la combinaison de plusieurs traitements différents. Ces barrières représentent les
7
différentes techniques de conservation des aliments (pH, O2, température, aw, etc.) qui permettent de réduire la charge bactérienne avec un effet bactéricide ou en limitant la croissance de ces micro-organismes avec un effet bactériostatique. La combinaison de plusieurs barrières permet d’augmenter leur efficacité sans pour autant altérer l’aliment. Par exemple, la stérilisation représente la plus forte barrière permettant d’éliminer tout micro-organisme. Par contre, son application est très limitée dans l’industrie alimentaire. La pasteurisation est aussi un traitement thermique, mais qui représente une barrière moins élevée que la stérilisation. Dans le cas du lait, sa combinaison avec une conservation à 4˚C permet d’avoir deux barrières efficaces et n’altérant pas le produit.
8
Figure 2: Principe des barrières en conservation alimentaire (Leistner et al. 1995)
Légende : F : chaleur, t : froid, aw : faible activité de l’eau, pH : acidification, Eh : faible potentiel redox, pres : agents de conservation, V : vitamines, N : nutriments.
9
Dans le cas du saumon fumé à froid, plusieurs barrières sont en place afin de tenter de limiter la propagation des micro-organismes. Ces barrières comprennent le fumage qui ajoute une multitude de composés ayant des effets antimicrobiens comme des formaldéhydes et des phénols [29]. Elles comprennent aussi l’ajout de sels qui au Canada, est recommandé à une concentration de 1% et permet de réduire l’aw du saumon, réduisant ainsi la propagation des micro-organismes présents sur le saumon. L’ajout de nitrites permet l’inhibition des principaux contaminants du saumon comme Listeria et Clostridium botulinum, mais devient de plus en plus critiqué par les consommateurs [30]. L’emballage permet aussi de contrôler l’atmosphère du produit. Au Canada, cet emballage doit être perméable à l’oxygène à un taux supérieur à 2000 cc/m2/24h afin de limiter la croissance de Clostridium qui est un micro-organisme anaérobique [31]. Finalement, la réfrigération et la congélation permettent de ralentir la croissance de la majorité des contaminants microbiens. Listeria réussit tout de même à croître à 4°C et peut se rendre à des concentrations de 100 UFC/g après 4 semaines d’entreposage [32].
La barrière manquante au procédé de transformation du saumon fumé est le fort traitement thermique qui permettrait de réduire la charge microbienne. Évidemment, cette barrière ne peut être mise en place, car le traitement thermique aurait un effet de cuisson sur la chair du poisson. Il est possible de voir la croissance microbienne lors de la conservation du poisson à la figure 3. En fonction de ces données, la réglementation canadienne limite la conservation des produits marins frais à 14 jours.
10
Figure 3: Croissance des microorganismes sur une matrice de poisson lors de sa conservation (Gram et Huss, 1996)
11
1.2
Nouvelles tendances dans la conservation des aliments
1.2.1 Bioconservation et cultures protectricesLe consommateur étant de plus en plus conscientisé par son alimentation, de nouveaux défis se rajoutent dans le domaine de la conservation. En effet, ce consommateur préfère des aliments ne contenant pas d’agents de conservation chimiques comme les nitrites. Évidemment, ce retrait des agents chimiques de l’alimentation ne doit pas se faire au détriment de son innocuité. Afin de contrôler les flores bactériennes pathogènes altérant les aliments, des bactéries inoffensives et productrices d’agents antimicrobiens sont souvent ajoutées aux aliments. C’est ce qui se nomme la bioconservation des aliments.
La bioconservation inclut aussi l’ajout de métabolites antimicrobiens sans la souche productrice [33]. Les métabolites produits par ces bactéries servent d’agents antimicrobiens permettent l’inhibition de contaminants bactériens. Dans les aliments fermentés, l’acide lactique et acétique, le lactate de sodium, le peroxyde d’hydrogène, le CO2, le diacétyle et l’acétaldéhyde produits par les bactéries lactiques permettent ce pouvoir inhibiteur. Ces composants causent la perméabilisation des membranes bactériennes [34]. La production d’acides organiques, tels l’acide propionique, l’acide sorbique, ainsi que des acides gras sont d’autres exemples d’agents permettant le contrôle de bactéries pathogènes [35]. La bioconservation ne mise pas seulement sur la production d’agents antimicrobiens par les souches protectrices ajoutées, mais aussi sur l’effet de compétition exclusive. Dans bien des produits fermentés comme les saucisses, la flore lactique aura un avantage compétitif sur la flore pathogène et d’altération, ce qui protégera l’aliment d’une potentielle contamination [33].
Certaines bactéries produisent aussi des peptides antimicrobiens (AMB) qui ont un potentiel d’inhibition qui ne se limite pas aux bactéries seulement, mais peuvent aussi cibler des virus et des parasites. Le mécanisme d’inhibition passe majoritairement par la formation de pores dans la membrane cellulaire ou par l’inhibition de leur formation [36]. Ces peptides proviennent aussi bien de procaryotes que d’eucaryotes et peuvent être des métabolites primaires ou secondaires. Chez les animaux, les peptides antimicrobiens sont considérés comme la première ligne de la défense immunitaire [37]. Les peptides antimicrobiens
12
produits chez les plantes comme les cyclotides ont aussi un rôle de défense immunitaire et offrent un potentiel pharmaceutique [38]. Enfin, les AMB produits par les bactéries sont plus communément référés comme des bactériocines.
La bioconservation par l’ajout de cultures protectrices permet d’ajouter une barrière de protection biologique, mais comprend aussi ces défis. En effet, l’ajout d’une flore protectrice qui est régulièrement composés de bactéries lactiques, va acidifier l’aliment. L’acidification et l’activité protéolytique de ces souches peut donc entraîner une altération de la matrice alimentaire. Par exemple, une étude de Leroi et al. [39] démontrait que l’application de plusieurs souches lactiques sur le saumon fumé à froid entraînaient la production d’odeurs non désirables. Il devient donc primordial d’identifier des souches ayant un faible potentiel acidifiant et protéolytique, mais produisant d’autres composés antimicrobiens.
Une autre méthode de bioconservation est l’ajout de bactériophages sur les aliments. Les bactériophages sont des virus qui se servent des bactéries afin de pouvoir se reproduire en utilisant le mécanisme cellulaire de la bactérie infectée. Suite à sa reproduction, le bactériophage entraîne la lyse cellulaire de la bactérie infectée. L’avantage de cette technique est de cibler précisément les pathogènes à éliminer de la matrice alimentaire. Il existe actuellement 5 produits à base de bactériophages permettant le contrôle de Listeria, E. coli, Salmonella, X. campestris et P. syringae (ListexTM P100, ListShieldTM, EcoShieldTM, SalmShieldTM et AgriPhageTM) [33]. Plusieurs défis restent à surmonter avec cette technologie comme la production des phages qui nécessitent la croissance de souches pathogènes et le besoin de créer des mélanges de phages afin de contrer les différents variants de pathogène retrouvés dans l’industrie de la transformation alimentaire [40].
1.2.2 Bactériocines
Les bactériocines font partie de ces peptides antimicrobiens et sont des métabolites bactériens. Ce pouvoir antimicrobien peut être bactéricide ou bactériostatique dépendamment de la bactériocine et de sa concentration. Les bactériocines ont une taille variant entre 30 et 100 acides aminés, et sont synthétisées par voie ribosomale [41]. 99% de l’ensemble des bactéries auraient la capacité de produire au moins une bactériocine [42]. Une minorité des
13
bactériocines est actuellement répertoriée, car le réel intérêt pour ces bactériocines est relativement récent. Toutes les bactériocines connues à ce jour peuvent être retrouvées dans la base de données en ligne appelée BACTIBASE avec le lien suivant :
http://bactibase.hammamilab.org/main.php [43].
À l’inverse des antibiotiques, les bactériocines ont généralement un spectre d’action précis ciblant des souches proches de la souche productrice. De plus, à activité équivalente, une plus grande concentration d’antibiotiques est nécessaire afin d’avoir la même activité que les bactériocines [4]. Plusieurs avenues d’utilisation de ces bactériocines sont présentement à l’étude. Un usage vétérinaire des bactériocines permettrait de remplacer les antibiotiques comme promoteurs de croissance. Les bactériocines peuvent aussi être employées comme agents de conservation dans l’industrie alimentaire, agents désinfectants pour la peau et éventuellement comme médicaments dans le traitement des infections gastro-intestinales [3].
La grande majorité des bactériocines répertoriées et étudiées sont produites par les bactéries Gram+. Les bactéries Gram- sont aussi productrices de bactériocines, mais elles sont moins nombreuses et moins diversifiées [3]. La colicine produite par E. coli (Gram-) est la bactériocine la plus étudiée [42]. Parmi les bactéries Gram+, ce sont surtout les bactéries lactiques (LAB) qui suscitent le plus grand intérêt par leur présence courante dans l’alimentation. Cette présence leur permet d’être généralement reconnues comme sécuritaires (GRAS) par des organismes de santé publique comme la FDA aux États-Unis. Ce n’est pas encore le cas pour toutes les souches de bactéries lactiques. Ailleurs qu’au Canada, ces bactéries sont entre autres fréquemment ajoutées dans les produits laitiers comme souches protectrices [44]. Carnobacterium maltaromaticum CB1 fait partie de ces souches protectrices commercialisées et ayant la reconnaissance GRAS (n° GRAS 305, 2010). La bactérie est commercialisée sous le nom de Micocin® et permet l’inhibition de L. monocytogenes dans une large gamme d’aliments [45].
1.2.3 Les bactériocines comme nouvel additif pour la conservation des aliments
L’utilisation seule des souches productrices de bactériocines dans les aliments ne permet pas de garantir l’élimination complète de pathogènes. L’efficacité des bactériocines comme
14
agents de conservation est limitée par leur spectre d’action spécifique. Leur usage doit donc être fait conjointement à une autre méthode de conservation afin d’assurer la sécurité de l’aliment [46]. L’ajout de souches protectrices produisant des bactériocines permettrait tout de même de remplacer les agents chimiques typiquement ajoutés afin de conserver l’innocuité de l’aliment et prolonger sa durée de vie [47].
Les souches protectrices et leurs bactériocines peuvent être introduites dans les aliments de trois façons. La première consiste à ajouter la bactériocine purifiée. Les désavantages de cette technique sont les coûts supplémentaires afin de purifier la bactériocine et la perte d’activité due aux conditions rencontrées dans l’aliment [6, 47]. La deuxième possibilité consiste à remplacer les souches utilisées lors d’une fermentation par des souches productrices de bactériocines. Par exemple, l’ajout de souches productrices de bactériocines comme Lactobacillus plantarum 423 et Lactobacillus curvatus DF38, comme souches fermentaires, permet d’inhiber la croissance de Listeria innocua [48]. La troisième possibilité est de sécher un milieu de culture fermenté contenant la souche protectrice et ses bactériocines pour en faire un ingrédient actif sous forme de poudre. Cet ingrédient pouvant ensuite être dilué à la concentration désirée et ajouté sur différents aliments afin d’inhiber la croissance de pathogènes. L’activité antimicrobienne est ainsi préservée et la souche ajoutée peut continuer à produire ses bactériocines. Les différents composants du milieu de culture se retrouvant dans cette poudre pourraient néanmoins altérer le goût de l’aliment [47].
L’ajout de cultures protectrices produisant des bactériocines est fréquente dans l’industrie alimentaire. Dans le domaine fromager, il a été remarqué que certaines lactocoques utilisés comme ferments produisaient de la nisine ayant un effet contre Clostridium, Bacillus et Listeria [33]. Dans le domaine des produits carnés, il est fréquent de retrouver des souches productrices de bactériocines dans les saucisses fermentés. La sakacine, leucocine, mésentérocine, l’entérocine et la nisine sont toutes des bactériocines retrouvées dans différentes saucisses fermentées et ayant un effet contre Listeria, Clostridium, Enterococcus, Staphilococcus et Bacillus [33]. Il est aussi possible de retrouver le même genre de souches productrices de bactériocines dans les produits marins, céréaliers, ainsi que la production de vins.
15
Actuellement, seules deux bactériocines purifiées peuvent être ajoutées aux aliments, à savoir la nisine (Nisaplin® par Danisco, Chrisin® par Chr. Hansen) pour le contrôle de Clostridium butyricum dans les fromages et la viande notamment [6] et la colicine E1 pour le contrôle d’Escherichia coli dans la viande de bœuf [7]. Toujours à l’extérieur du Canada, la nisine produite par Lactococcus lactis est fréquemment utilisée dans l’industrie laitière, ainsi que comme désinfectant permettant de soigner les mammites bovines [44]. Son application est limitée par une perte d’activité lorsque le pH augmente, mais surtout par son interaction avec les composants de la viande. Elle ne peut donc pas être utilisée seule comme agent de conservation de produits carnés.
1.3
Carnobacterium divergens M35
Carnobacterium divergens M35 produisant la divergicine M35 présente un intérêt particulier. Les carnobactéries font partie des LAB et sont présentes dans de nombreux produits carnés, marins et laitiers. Cette omniprésence est particulièrement vraie pour Carnobacterium divergens et C. maltaromaticum qui proviennent du microbiote intestinal des poissons. Cette souche fut isolée de moules fumées commerciales lors de travaux antérieurs de l’équipe du Dr Fliss. C. divergens M35 est caractérisée par la production d’une bactériocine, la divergicine M35 [49]. Cette bactériocine ayant un poids moléculaire de 4517,75 Da, possède une forte activité anti-Listeria, mais aussi contre les autres carnobactéries. C’est surtout son activité contre Listeria qui la rend très intéressante dans l’industrie de la transformation des produits de la mer. De plus, sa faible activité protéolytique limite le potentiel de dégradation de la matrice alimentaire si C. divergens M35 y est ajoutée. Récemment, l’équipe du Dr Ismail Fliss a homologué auprès de Santé Canada (réf. ADDFS 11062001) l’application de C. divergens M35 sur les produits marins fumés [50]. La divergicine M35 produite par C. divergens M35 est d’ailleurs protégée par un brevet (US 6013-141).
1.3.1 Bio-ingrédient anti-Listeria à base de M35
Les études d’incorporation de souches de bactéries lactiques sur le saumon fumé à froid afin de lutter contre la croissance de Listeria ont débuté aux débuts des années 2000 avec des travaux comme ceux de Truelstrup Hansen [51]. Depuis, plusieurs travaux ont été effectués
16
avec une autre souche de carnobactéries, soit Carnobacterium divergens V41. Ces travaux ont démontré l’inhibition de Listeria par l’addition de sa bactériocine sur le saumon fumé [52]. Dans la majorité de ces études, la souche protectrice et sa bactériocine avaient été purifiées avant l’application sur la matrice alimentaire. Par contre, le faible rendement de purification des bactériocines, ainsi que la capacité réduite de production de la bactériocine une fois sur la matrice alimentaire diminuent l’efficacité de cette technique. L’équipe du Dr Fliss fut la première à tester l’application du milieu de culture fermenté par C. divergens M35 comprenant aussi sa bactériocine [50].
Cette nouvelle méthode permet de créer un bio-ingrédient ayant deux actions. Dans un premier temps, la bactériocine contenue dans le milieu de culture fermenté agira contre Listeria dès son application sur le saumon fumé à froid. L’application du milieu de culture fermenté ajoutera aussi C. divergens M35 sur le saumon. Cette bactérie continuera donc à produire sa bactériocine sur la matrice alimentaire afin de maintenir l’inhibition contre Listeria. Sa présence en plus forte concentration compétitionnera avec les autres contaminants microbiologiques pour les nutriments présents à la surface du saumon. Cette compétition n’affectant pas la qualité finale de la matrice de saumon. L’application du milieu de culture fermenté sur la matrice alimentaire permet de simplifier le procédé de fabrication, en éliminant les étapes de purification.
Afin de pouvoir offrir ce bio-ingrédient actif à l’échelle industrielle, la production doit être optimisée. En effet, les carnobactéries, ainsi que les bactéries lactiques en général, sont des bactéries nutritionnellement très exigeantes et ont une productivité protéique faible. Les milieux préconisés pour ce type de souches, comme le milieu de Man-Rogosa-Sharpe (MRS), sont très dispendieux et peu adaptés pour les carnobactéries [10, 44]. Un autre facteur affectant le rendement en bactériocines est la méthode de stabilisation. La perte de l’activité antimicrobienne est due à la sensibilité des bactériocines à l’oxydation et aux variations de pH lors de l’entreposage du produit [44]. Il en va de même pour la viabilité de la souche qui est influencée par la méthode de stabilisation choisie.
17 1.3.1.1 Milieux de culture
1.3.1.1.1 Milieu de référence : MRS
Le milieu de culture de référence actuellement pour C. divergens M35 est le MRS. Ce milieu permet une excellente croissance des bactéries lactiques (LAB) et permet de maximiser la production des bactériocines pour une majorité de ces souches. Les LAB sont des souches qui nécessitent des milieux complets et riches. Le MRS répond à ces demandes, mais il est actuellement trop dispendieux pour un usage industriel. Carnobacterium divergens M35, qui est une bactérie lactique, croît jusqu’à une densité de 3×10^8 UFC/mL en 30h, avec une production de divergicine M35 à 2048 AU/mL, dans le milieu MRS [10]. La composition du MRS est (g/L) : Peptone de protéose (10), extrait de bœuf (10), extrait de levure (5), dextrose (20), polysorbate 80 (1), citrate d’ammonium (2), acétate de sodium (5), sulfate de magnésium (0,1), sulfate de manganèse (0,05) et phosphate dipotassique (2). Ensemble, les trois sources d’azote (peptone, extrait de bœuf et extrait de levure) fournissent près de 3,125g/L d’azote [53]. Cette quantité d’azote peut donc servir de référence lors de l’élaboration de milieux alternatifs.
Plusieurs études portent sur l’utilisation de milieux alternatifs et moins coûteux que le MRS, tout en utilisant ce milieu comme contrôle positif. Ce milieu doit être modifié afin d’être adapté aux critères spécifiques de la souche étudiée. Afin de réduire les coûts associés au milieu de culture MRS, les dispendieuses sources de carbone et d’azote sont souvent remplacées par des sous-produits ou déchets de l’industrie alimentaire et de l’agriculture [11].
1.3.1.1.2 Sources de carbone
Dans un milieu de culture, la source de carbone sert de source d’énergie pour la bactérie, mais sert aussi de constituant à la cellule bactérienne. Les sources de carbone ne sont généralement pas les éléments les plus dispendieux d’un milieu de culture, mais elles peuvent le devenir à grande échelle. Diverses sources de carbone provenant de l’industrie alimentaire peuvent remplacer le glucose présent dans le MRS. Le sucrose est métabolisé par les carnobactéries [54] et est la source de carbone dans le milieu CTSI (pour Cresol red Thallium acetate Sucrose Inulin medium) servant à isoler ces carnobactéries [55]. La mélasse de canne contient majoritairement du sucrose et est un sous-produit retrouvé en grande quantité dans
18
l’industrie du sucre. La mélasse blackstrap est le résidu de mélasse suite à l’extraction du sucrose qu’elle contient. Elle contient généralement moins de 46% de sucres totaux, mais contient aussi une large variété de sels et minéraux comme du potassium, du manganèse, du magnésium, etc. [56, 57]. Ce sont d’ailleurs des sels retrouvés dans le milieu MRS. Son abondance et son faible coût favorise son utilisation fréquente dans la fabrication de bioéthanol [58]. L’extrait de malt est une source majoritaire de maltose, mais contient aussi du glucose, du sucrose et d’autres sources variées de carbone. Il s’agit donc d’une source de carbone complexe pouvant aussi contenir des protéines [59, 60]. C’est un ingrédient à faible coût fréquemment utilisé dans l’industrie alimentaire qui pourrait se substituer au glucose dans le MRS.
1.3.1.1.3 Sources d’azote
Les sources d’azote servent à fournir tous les acides aminés nécessaires à la synthèse de protéines par la bactérie. Les sources d’azote sont beaucoup plus dispendieuses et plusieurs études détaillent les diverses sources provenant des sous-produits de l’industrie alimentaire. La croissance des bactéries lactiques est d’ailleurs souvent dictée par leur capacité limitée à synthétiser divers acides aminés. La source d’azote du milieu de culture doit donc être complète et en grande quantité. Les extraits de levure demeurent la source d’azote la plus largement utilisée. En effet, l’extrait de levure est une source complexe d’azote fournissant un mix d’acides aminés, de peptides, de vitamines, mais peut aussi servir de sources de carbone. Cet apport très varié en fait une source d’azote pouvant être utilisé pour la croissance d’une large gamme de micro-organismes [61]. Les autolysats de levures provenant de l’industrie de la bière sont peu coûteux et offriraient les mêmes rendements que les extraits de levure. Par contre, la présence de gluten dans ce produit rend son application difficile dans les différents produits alimentaires. En effet, avec les cas de plus en plus nombreux d’intolérance au gluten, il serait inapproprié d’en ajouter sur un produit qui n’en contient normalement pas. Il est aussi fréquent de voir l’extrait de levure jumelé à une autre source d’azote afin de compléter l’apport en acides aminés.
Ces sources peuvent être des extraits de viande qui contiennent des peptides, des acides aminés, des acides organiques, des minéraux et quelques vitamines. Ce type d’extrait
19
provient de l’extraction liquide de résidus carnés qui est ensuite séché pour obtenir une poudre [61]. Ce type de source d’azote est fréquemment utilisé en microbiologie, mais est hautement supervisé dans l’industrie pharmaceutique afin d’éviter la transmission d’encéphalopathies spongiformes transmissibles (TSE) ou d’encéphalopathies spongiformes bovines (BSE). Les sources d’azote peuvent être également végétales comme les peptones de pomme de terre [13] ou de pois [62]. Elles permettent justement d’éviter la potentielle présence de prions qui pourraient provenir d’une infusion de viande [63]. Les hydrolysats de poissons représentent aussi une source d’azote permettant de valoriser un déchet de l’industrie alimentaire. De plus en plus d’études se penchent sur l’utilisation de cette riche et complexe source d’azote dans le cadre de divers types de fermentation [11, 64]. Récemment, la fermentation d’un milieu à base de peptones de hareng a permis la production de bactériocines avec Pediococcus acidilactici. Cette fermentation permettant d’atteindre une activité anti-Listeria de 32768 AU/mL [64]. Enfin, ces sources peuvent aussi provenir de l’industrie laitière sous la forme de lactosérum et de babeurre [65]. Ce sont deux sous-produits retrouvés en abondance au Québec et ayant tout de même une forte teneur en protéine.
Les sources d’azote pouvant être utilisées dans un milieu de culture sont nombreuses et très variées. Par contre, la disponibilité des acides aminés varie fortement entre ces différentes sources. La taille des peptones va aussi avoir une influence sur la capacité d’une bactérie à avoir accès aux acides aminés. Ainsi, une source d’azote fournissant des peptones de petite taille sera probablement préférentielle pour la croissance de C. divergens M35 et la production en bactériocines.
20
Tableau 1 : Constituant des diverses sources d'azotes
E xtr ait d e levure 2 L ac tosé ru m n on d én aturé 4 B ab eu rr e 4 Pe p tone d e p om m e d e te rr e 1 Pe p tone d e vian d e 1 Pe p tone d e p ois 1 Hyd rolysa t de p oiss on 3 Protéine LECO 56,8% 32,7% - - - 40% Azote total (TN) 10 - 11,8% 9,7% 5,6% 10 - 11% 6,1% 13,5 - 14,5 % 6,3% Azote aminé (AN) 4,8 - 6,3% - - 5 - 6% 2,6% 3,8 - 4,8% - Gras - 5,12% 7,18% - - - 14% Distribution poids moléculaire - - - - > 10 000 Daltons 0% - - 0,3% 3% - - 1000 - 10 000 Daltons 11% - - 8% 38% - - 500 - 1000 Daltons 13% - - 18% 28% - - 300 - 500 Daltons 17% - - 19% 18% - - <300 Daltons 59% - - 55% 13% - -
(1) Organotechnie, La Courneuve, France, (2) Biena, St-Hyacinthe, Canada, (3) Océan NutraSciences, Matane, Canada et (4) Agropur, Granby, Canada
21 1.3.1.1.4 Sources de sels
Finalement, les sels comme le citrate d’ammonium et l’acétate de sodium semblent avoir une influence sur la croissance de C. divergens M35 et sur sa production de divergicine. Ce sont les deux sels en plus grande concentration dans le milieu MRS. Par contre, l’ajout d’acétate de sodium montre des résultats contradictoires tant dans la littérature que suite aux travaux de l’équipe du Dr Ismail Fliss [10, 66]. L’acétate aurait un effet inhibiteur sur la croissance bactérienne tout en favorisant la production de bactériocines. Pour Carnobacterium piscicola A9b, l’acétate serait un inducteur de la production de bactériocines. Sa concentration aurait une influence sur la production de bactériocines [67].
1.3.1.2 Stabilisation du bio-ingrédient
Une fois que la concentration en biomasse et que l’activité antimicrobienne est atteinte, la stabilisation du produit désiré devient essentielle. L’activité des bactériocines produites est fortement vulnérable à l’oxydation et aux variations de pH. Cette stabilisation doit permettre de conserver la viabilité de la souche ainsi que l’activité anti-Listeria de sa bactériocine. Le séchage du milieu de culture fermenté représente la meilleure solution, car elle permet d’abord de réduire la masse du bio-ingrédient en éliminant le contenu en eau. Cette réduction de la masse facilite ainsi son transport et son entreposage. De plus, le séchage permet de réduire l’activité de l’eau du bio-ingrédient. Les phénomènes d’altération du bio-ingrédient comme l’oxydation sont donc fortement réduits, ce qui augmente considérablement la durée de vie du bio-ingrédient. Le séchage permet aussi d’éliminer le risque du développement de bactériophages dans un milieu de culture fermenté et conservé à long terme [68].
Deux méthodes de séchage peuvent être employées dans le cas présent, soit la lyophilisation et l’atomisation. La lyophilisation est une technologie répandue dans l’industrie des biotechnologies, car elle permet généralement de maintenir un taux de survie de la souche bactérienne pouvant aller jusqu’à 100% [69]. Évidemment, ces taux de survie dépendent de la souche bactérienne, mais aussi des agents cryoprotecteurs ajoutés lors du séchage par lyophilisation. En effet, la lyophilisation est considérée comme une méthode de séchage douce, mais la congélation requise avant le séchage peut endommager les membranes cellulaires lors de la formation de cristaux et causer une perte de viabilité [70]. La lyophilisation est aussi limitée par son opération en mode discontinu, ce qui limite sa capacité
22
de séchage. Le séchage peut prendre plusieurs jours en fonction de l’équipement et du volume à sécher. De plus, cette méthode est considérée comme étant la plus dispendieuse au niveau de l’achat de l’équipement, mais aussi lors de l’opération de l’équipement [68, 71]. En effet, pour la même quantité d’eau à retirer, il en coûterait six fois plus cher de le faire par lyophilisation que par atomisation [71]. Dans le cas du séchage de probiotiques, le milieu de culture fermenté est centrifugé afin de recueillir exclusivement la biomasse. Cette biomasse ayant un faible volume, la lyophilisation est appropriée afin de la stabiliser. Dans le cas du bio-ingrédient à base de C. divergens M35, tout le milieu de culture doit être séché. Le volume à traiter est donc considérablement plus élevé et rend l’usage de la lyophilisation économiquement moins intéressant.
Le séchage par atomisation est la méthode d’intérêt dans le cadre de cette étude, car elle permet de traiter de plus larges volumes. À l’inverse de la lyophilisation, l’atomisation est un traitement thermique qui mise sur le séchage très rapide du liquide. Ce liquide est atomisé en une fine bruine dans une chambre chauffée par un jet d’air à co-courant du liquide. La bruine ayant une très grande surface de contact avec l’air chaud, le contenu en eau s’évapore avant le contact avec la paroi de la chambre de séchage [68]. Un cyclone est installé en série après la chambre afin de séparer la matière sèche de l’air humide. La figure 4 permet de visualiser le fonctionnement de cet équipement. Il s’agit d’une méthode très versatile et qui est utilisée dans plusieurs types d’industrie. Les équipements de séchage par atomisation ont une très grande capacité de séchage pouvant même atteindre 100kg/h dans les industries minières [72]. Malgré ses avantages, cette méthode est peu documentée dans le domaine des biotechnologies. Le traitement thermique, bien que court, peut nuire considérablement à la survie de la souche bactérienne. Par contre, des études récentes démontrent la faisabilité technique de cette méthode afin de sécher des cultures bactériennes. Dans la majorité des cas, ces études portent sur le séchage de cultures utilisées dans l’industrie laitière, ce qui inclut souvent les lactobacilles [68, 72, 73]. Une étude démontre même la faisabilité du séchage par atomisation de bactéries produisant des bactériocines [74].
23
24
Un des défis reliés à la méthode de séchage par atomisation est la perte de rendement massique dans le procédé. Le produit devant passer par la chambre de séchage, puis par des canalisations menant au cyclone, le produit peut coller aux parois. Ce problème d’adhérence est particulièrement rencontré dans le domaine du séchage des jus de fruits et légumes. Le sucre qui est en forte concentration dans ces produits va caraméliser lors du séchage et entraîner une forte perte de rendement massique. Ceci est dû à la faible température de transition vitreuse (Tg) de ces sucres [75]. Cette température est de 62°C dans le cas du sucrose. La même problématique s’applique lors du séchage de milieux de culture fermentés qui contiennent encore des sucres. Afin de pallier à cette problématique, des agents aidant au séchage peuvent être ajoutés à la solution, augmentant ainsi la température de transition vitreuse. La maltodextrine et la gomme arabique sont parmi les agents aidant au séchage les plus utilisées dans l’industrie alimentaire [76, 77]. L’isolat de protéines de lactosérum (IPL) en faible concentration (1%) peut aussi être un excellent agent aidant au séchage, mais dans ce cas, il est considéré comme un potentiel allergène [75]. La maltodextrine fut choisie comme agent aidant au séchage dans cette étude, car il s’agit d’un polysaccharide bien connu et qu’il est bien moins dispendieux que la gomme arabique.
1.3.1.3 Mise à l’échelle
Afin de déterminer la viabilité du concept élaboré en laboratoire, il est primordial de faire la mise à l’échelle du procédé de production. Les résultats obtenus en laboratoire peuvent grandement être différents de ceux obtenus à plus grande échelle. Plusieurs variables viennent influencer cette mise à l’échelle, mais la principale est la variation dans le transfert de masse. Ce transfert de masse dicte la présence de gradients en nutriments dans le fermenteur, ce qui pourrait nuire à croissance de C. divergens M35. Le transfert de masse variera en fonction des dimensions du fermenteur, de l’agitation appliquée ainsi que des propriétés physiques du milieu de culture. Il est donc important de s’assurer que le transfert de masse reste optimal lors de la mise à l’échelle du procédé [78]. La distribution de l’oxygène dans le milieu de culture est souvent le facteur le plus influencé par le transfert de masse. Dans le cas de C. divergens M35, ce ne sera pas un facteur majeur puisque cette bactérie est anaérobique. Par contre, la présence de zones mortes, où les nutriments sont en faibles concentrations, vient nuire à la croissance de la souche et à sa production de
25
bactériocines. Le même stress peut être observé dans une zone où les nutriments y sont en trop grandes concentrations [79, 80]. Cette mise à l’échelle est primordiale pour l’industrie afin de démontrer la viabilité industrielle des travaux accomplis en laboratoire.
26
Hypothèse
L’utilisation d’un milieu de fermentation optimal à faible coût jumelé à l’optimisation des conditions de production et de stabilisation permettraient de produire de façon rentable un bio-ingrédient à base de Carnobacterium divergens M35 et de divergicine à forte activité antimicrobienne.
Objectifs
1. Étudier l’effet de la supplémentation du milieu de fermentation par différentes sources de carbone, d’azote et de sels sur la biomasse et la production de bactériocine par Carnobacterium divergens M35;
2. Produire à l’échelle pilote une formule de grade alimentaire stable et biologiquement active à base de divergicine M35;
3. Effectuer la mise à l’échelle du procédé de fermentation et démontrer la faisabilité technique du séchage par atomisation.
27
Chapitre 2 : Optimisation d’un milieu de culture et des
conditions d’opération pour la croissance de
28
2.1
Résumé
Lors de travaux antérieurs, un bio-ingrédient permettant la bio-conservation du saumon fumé à froid et ayant une forte activité anti-Listeria a été développé. Ce bio-ingrédient consiste en un milieu de culture fermenté par Carnobacterium divergens M35 et contenant la bactériocine produite par la souche, soit la divergicine M35. Dans un premier temps, le but de cette étude est de développer un milieu de culture de grade alimentaire favorisant une forte et rapide croissance de C. divergens M35 et stimulant la production de sa bactériocine. Un criblage de différentes sources d’azote, de carbone et de sels a permis de déterminer que les meilleures sources de carbones sont la mélasse de canne et le sucre de table (saccharose), la source d’azote de choix restait l’extrait de levure, que l’acétate de sodium permet d’augmenter la production en bactériocine et que le tween 80 est nécessaire afin d’éviter l’agrégation des bactériocines.
La méthode de surface de réponse fut employée en fermenteurs d’1L afin d’optimiser la concentration de chacun des composants. Ce test a permis d’identifier l’influence majeure de la concentration en extrait de levure sur la croissance de la souche et sur sa production en bactériocine. Deux milieux furent donc créés; le premier, STYA contient 5g/L de sucre de table, 42g/L d’extraits de levure, 5g/L d’acétate de sodium et 1g/L de Tween 80, et le deuxième, MYA, remplace le sucre de table par 30g/L de mélasse de canne. Ces deux milieux furent testés en fermenteurs de 30L afin d’évaluer l’effet de la mise à l’échelle. Le milieu STYA permet d’atteindre une biomasse de 9,04 log(UFC/mL) et une activité de 131 072 AU/mL en 7h, tandis que le milieu MYA permet d’atteindre une biomasse de 9,15 log(UFC/mL) et une activité de 262 144 AU/mL en 9h. Pour ces deux milieux, il s’agit d’une amélioration significative en comparaison avec le milieu MRS qui est normalement utilisé.
Finalement, une co-culture entre C. divergens M35 et Carnobacterium maltaromaticum CB1 fut testée afin de déterminer s’il était possible d’augmenter l’activité produite lors de la fermentation et s’il était possible d’augmenter le spectre d’action du bio-ingrédient. Par contre, la croissance de C. maltaromaticum CB1 fut totalement inhibée par la présence de C. divergens M35. Aucune stimulation de l’activité ne fut possible par la co-culture de ces deux souches. Concernant la stabilisation de la divergicine M35, il a été démontré que la
29
bactériocine n’est pas sensible à l’oxydation, mais que l’activité fut diminuée de moitié si le bio-ingrédient est entreposé pendant 24h à température pièce dans sa forme liquide. Ce résultat démontrant la grande importance d’une méthode subséquente de séchage du bio-ingrédient.
30
2.2
Introduction
Comme le milieu de culture fermenté est directement appliqué sur la matrice alimentaire à protéger, ce milieu ne doit pas contenir d’allergènes ni de peptones animales. Le milieu de référence actuel est le MRS qui permet à C. divergens M35 d’atteindre une concentration de 3×10^8 UFC/mL en 30h, avec une production de divergicine M35 à 2048 AU/mL [10]. Afin de développer un milieu plus performant que le MRS et répondant aux exigences de l’industrie alimentaire, la base de ce milieu fut conservée, mais ses sources d’azote, de carbone et de sels furent modifiées. Quatre sources de carbone furent choisies, soient le glucose et le sucrose qui sont des sources de carbone communes, mais aussi la mélasse et l’extrait de malt qui sont des déchets de l’industrie alimentaire et permettent de réduire le coût de fabrication du milieu [58, 59]. Les sources d’azote sont les composants les plus coûteux d’un milieu de culture et ayant une influence significative sur la croissance de la souche [11]. L’utilisation de sources d’azote provenant de déchets alimentaires comme l’hydrolysat de poisson ou le lactosérum permettent de réduire ces coûts, tout en valorisant une matière organique normalement peu utilisée [64, 65]. Les sources de sels sont aussi à tester afin d’évaluer leurs influences sur l’atteinte d’une forte biomasse ou d’une forte production en bactériocines. La source d’azote pouvant déjà fournir l’apport nécessaire de ces sels. En testant les différentes combinaisons de sources de carbone, d’azote et de sels, il sera possible de créer un milieu plus performant que le MRS et ayant un coût beaucoup plus faible.
Une fois les composants déterminés, l’optimisation de leur concentration respective est nécessaire et permettra l’atteinte d’une croissance optimale de la souche. Une faible concentration des composants permet de réduire les coûts, mais elle peut devenir l’élément limitant la croissance et la production en bactériocines [81]. Cette optimisation par la méthode de surface de réponse (MSR) se fera en fermenteur d’un litre afin de contrôler les conditions de culture comme l’agitation, le contrôle du pH et de la température [10]. Cette étape permettra aussi de faire une première étape de la mise à l’échelle nécessaire à l’obtention de la crédibilité industrielle.
31
D’autres facteurs sont aussi à considérer dans le procédé de fabrication du bio-ingrédient. La stimulation de la production en bactériocines peut être induite dans un milieu de culture défavorable, imposant un stress à la bactérie [82]. Une étude de Sip et al.,1998 [83] a même démontré que la co-culture entre une souche de C. divergens et une souche de C. maltaromaticum permettrait l’obtention d’une activité anti-Listeria 64 fois plus élevée que la culture seule de C. divergens. Un autre facteur à considérer lors du procédé de fabrication du bio-ingrédient est la sensibilité de la bactériocine au stress oxydatif [4]. Cette sensibilité peut compromettre l’entreposage à long terme du bio-ingrédient et force l’ajout d’agents antioxydants si la divergicine M35 démontre cette sensibilité.
La dernière étape de ce chapitre consiste en la deuxième mise à l’échelle du procédé de fermentation en bioréacteur de 30L. La principale difficulté lors de cette étape est la différence dans le transfert de masse entre deux fermenteurs de volumes différents [84]. La qualité de ce transfert de masse va être influencée principalement par la géométrie du fermenteur et la force d’agitation utilisée. Un bon transfert de masse est qualifié par une turbulence uniforme qui réduit la présence de zones mortes dans le fermenteur [79]. Il est donc crucial de s’assurer que la mise à l’échelle du procédé de fermentation n’influence pas à la baisse la culture de C. divergens M35. Cette mise à l’échelle dans un volume pilote permet de démontrer la faisabilité industrielle du procédé de fermentation.
Le premier objectif de ce chapitre est de développer un milieu de fermentation à faible coût, permettant une croissance optimale de C. divergens M35 et favorisant la production de sa bactériocine. L’influence des conditions de culture, de la co-culture et du potentiel oxydatif de la bactériocine seront évalués afin de maximiser le potentiel du bio-ingrédient. Finalement, une mise à l’échelle du procédé de fermentation permettra d’obtenir une évaluation préliminaire de la faisabilité de la production du bio-ingrédient à l’échelle industrielle.
32
2.3
Matériel et Méthodes
2.3.1 Souches bactériennes et milieux de culture
Carnobacterium divergens M35 fut isolée à partir de moules fumées commerciales [49] et est détenue par l’Université Laval. Listeria ivanovii HPB28 provient d’Éric Émond (Santé et Bien-Être Social Canada). Carnobacterium maltaromaticum CB1 provient de CanBiocin, Alberta, Canada. Toutes ces souches furent maintenues à -80°C dans des solutions cryoprotectrices contenant du glycérol à 20%.
La culture de C. divergens M35 et de C. maltaromaticum CB1 s’est fait aérobiquement à 30°C dans le milieu MRS (Difco Laboratories, Sparks, MD, USA). 5% (v/v) d’inoculum fut choisi pour ces deux souches, afin de reproduire les conditions d’opération en industrie. La culture de L. ivanovii HPB28 s’est faite de façon aérobique à 30°C dans le milieu TSB (Difco Laboratories, Sparks, MD, USA) additionné d’extrait de levure à 0,6% (m/v). L’inoculum pour cette souche lors du repiquage était de 1% (v/v).
2.3.2 Dénombrement bactérien
Le dénombrement de C. divergens M35 et de C. maltaromaticum CB1 s’est fait à l’aide de la méthode des gouttelettes sur gélose TSB additionnée d’extraits de levure à 0,6% (m/v), puis incubé à 30°C pendant 48h. Une dilution en série au 1/10 de chacun des échantillons dans l’eau peptonée 0,1% permet ensuite d’évaluer la concentration bactérienne en UFC/mL.
2.3.3 Mesure de l’activité anti-Listeria
Les mêmes échantillons utilisés pour le dénombrement bactérien furent utilisés afin de quantifier l’activité anti-Listeria. L’activité peut être mesurée de deux façons : par diffusion en gélose ou par microtitration. La méthode de diffusion est celle développée par Tagg et al. [85]. L’activité anti-Listeria du bio-ingrédient fut évaluée par le cercle d’inhibition dans une gélose TSBYE inoculée avec Listeria.
Microtitration : Listeria ivanovii HPB28 fut utilisée comme souche sensible. Un millilitre d’échantillon fut prélevé et centrifugé pendant 10 min à 10000 g. Un traitement thermique à
![Figure 1 : Causes microbiologiques de rappels alimentaires en 2016 au Canada et aux États-Unis [1]](https://thumb-eu.123doks.com/thumbv2/123doknet/3209118.91740/16.918.203.704.506.893/figure-causes-microbiologiques-rappels-alimentaires-canada-états-unis.webp)