BRCA1 et réponse aux agents anticancéreux dans les hépatocarcinomes
122
0
0
Texte intégral
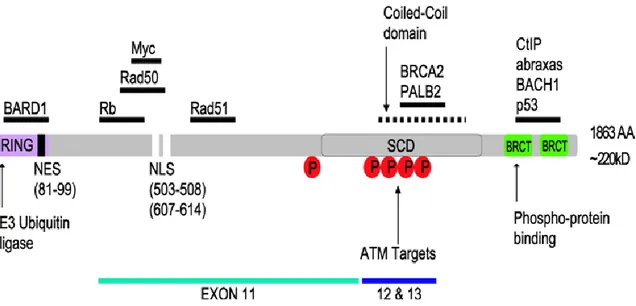
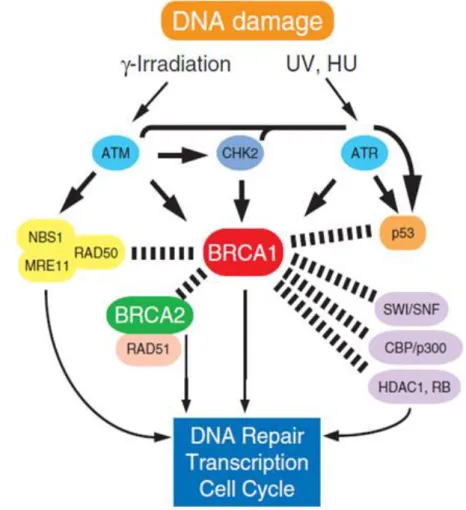
Figure
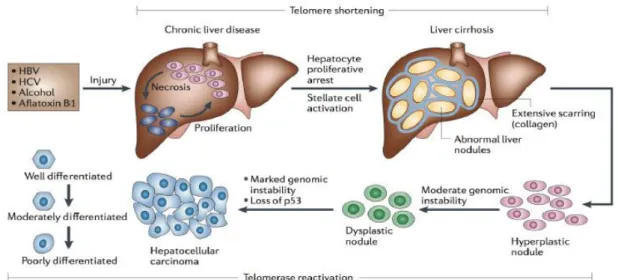
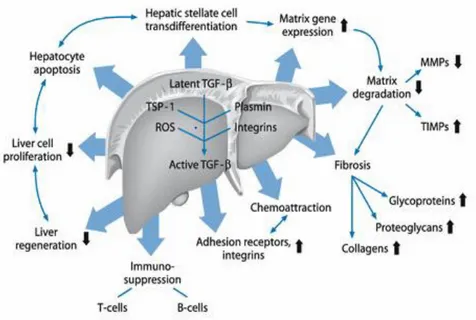
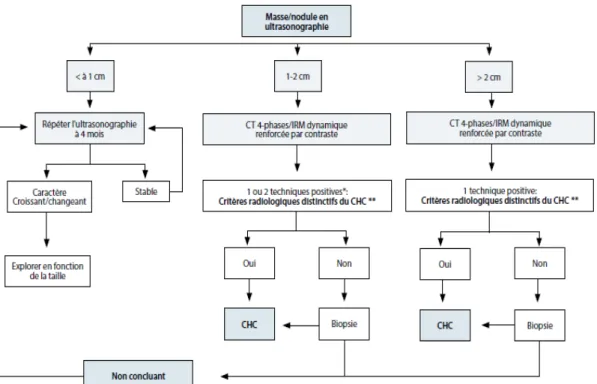
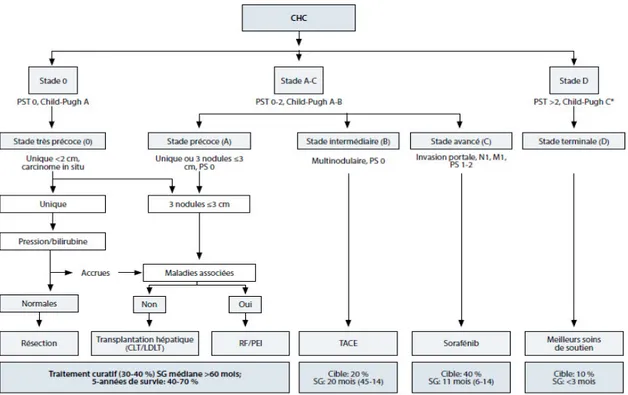
+7
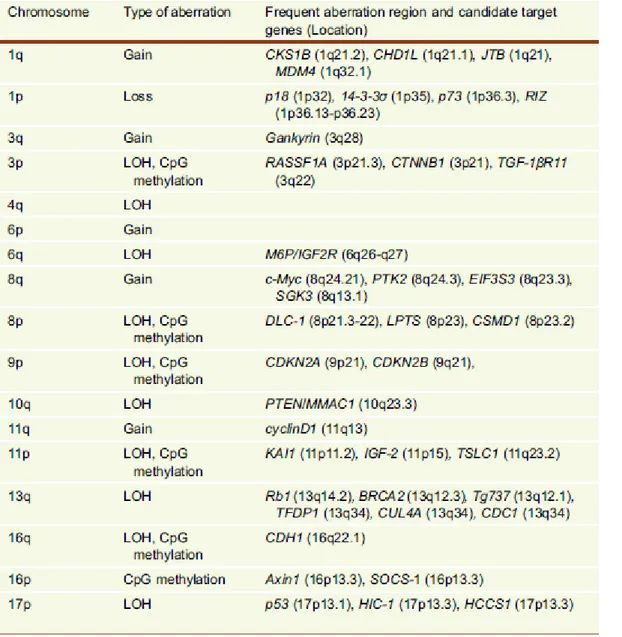
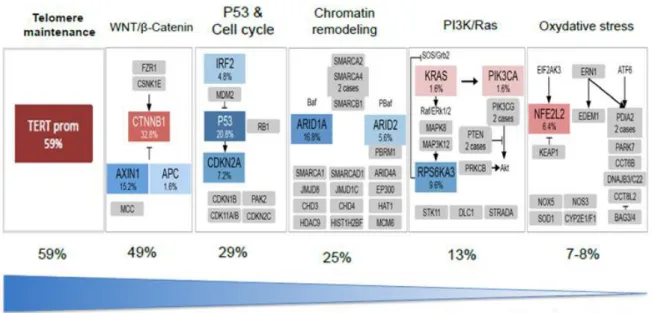
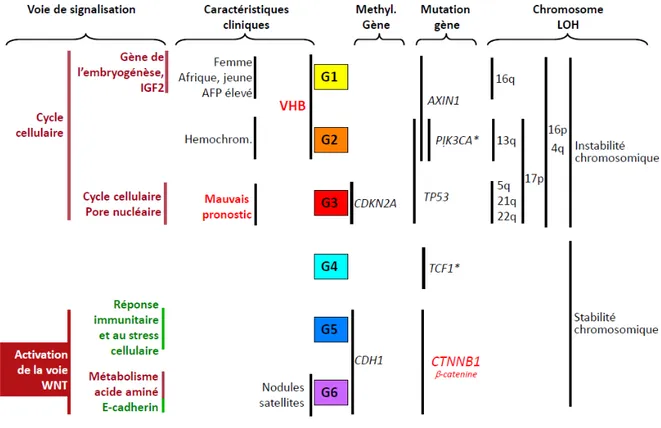
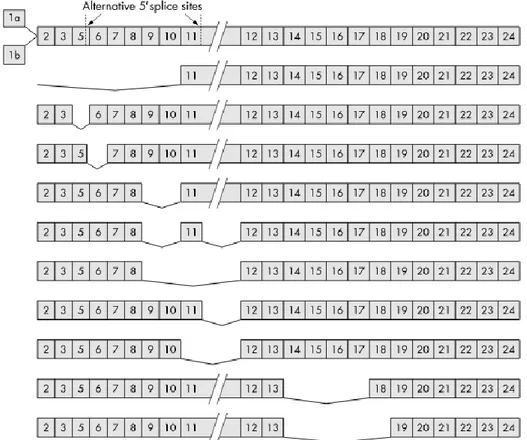
Documents relatifs
