Variabilité du chant de la paruline à gorge grise (Oporornis agilis)
Texte intégral
Figure

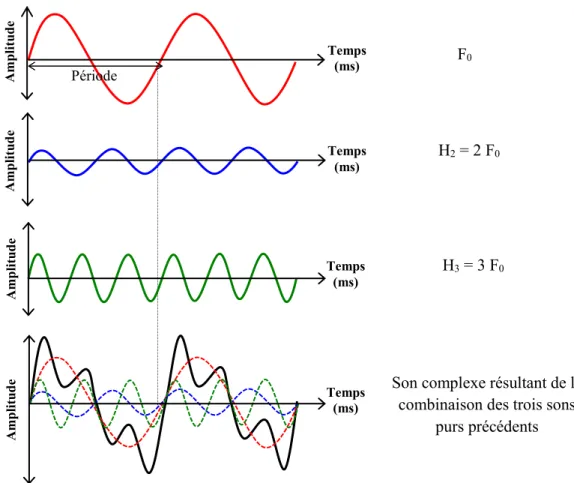
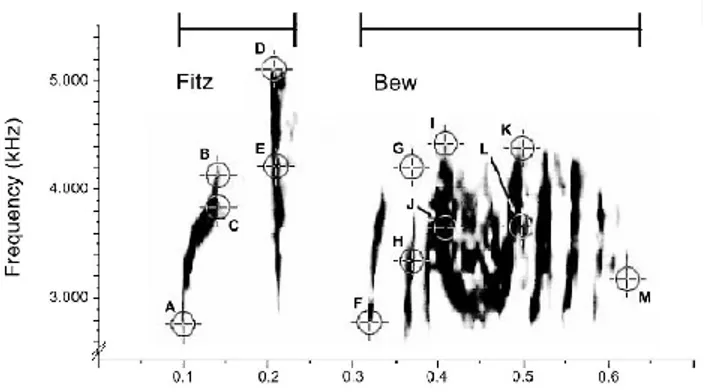


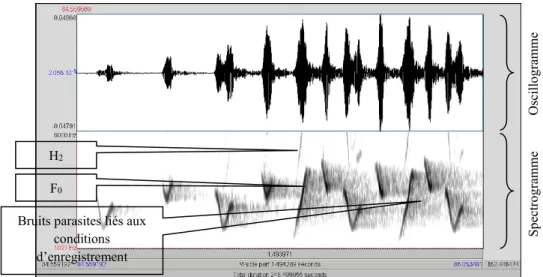




Documents relatifs
Les résultats de cette étude confirment ceux de l’étude Cochrane, c’est à dire une réduction du risque relatif de mortalité toutes causes confondues pour
Dans cette étude, dont les résultats sont présen- tés dans le tableau X, l’auteur retrouve moins de troubles de la voix chez les enseignants pratiquant le chant et le théâtre. Il
Les résultats de cette étude confirment la présence, dans le secteur sud du bassin volcano-sédimentaire de la Pako, de la majorité des lithofaciès (schistes, amphibolites,
Si les résultats de cette étude confirment une préférence des filles pour le français et des garçons pour les mathématiques, avec globalement une plus grande
Les résultats de notre étude confirment la faisabilité, l’efficacité et l’intérêt de la voie laparoscopique dans le traitement des prolapsus génitaux par promontofixation,
Regard sur la pratique du chant choral chez les personnes âgées : place dans les maisons de retraite, effets sur l’évolution de la voix.. Médecine humaine
Nous nous appuyons sur les résultats de cette étude pour montrer les références qui sont mobilisées dans les interactions verbales et le support qu’elles prêtent
Dans notre étude, chez les 137 patients traités, parmi les 11 résultats divergents, la non- prise en compte des valeurs élevées d’automesure chez 4 patients ayant une
