1
T
T
H
H
È
È
S
S
E
E
En vue de l’obtention duD
D
O
O
C
C
T
T
O
O
R
R
A
A
T
T
D
D
E
E
L
L
’
’
U
U
N
N
I
I
V
V
E
E
R
R
S
S
I
I
T
T
E
E
D
D
E
E
T
T
O
O
U
U
L
L
O
O
U
U
S
S
E
E
Délivré par l'Université Toulouse III - Paul Sabatier
Discipline ou spécialité : Biologie Cellulaire et Moléculaire
Présentée et soutenue par Claire Leduc
Le 16 Novembre 2009
Titre : Impact de la délétion de TRRAP ou MSH2 sur le développement des cellules B et la commutation isotypique
JURY
Michèle Goodhardt Directrice de recherche Pierre Ferrier Directeur de recherche Ahmad Oulmouden Professeur d'université Catherine Muller Professeur d'université
Michel Cogné Professeur d’université Amine Ahmed Khamlichi Chargé de recherche
Ecole Doctorale : Biologie Santé Biotechnologie Toulouse
Unité de recherche : CNRS UMR 5089
Directeur(s) de thèse : Amine Ahmed Khamlichi
3
Remerciements
Je souhaite en premier lieu remercier le Docteur Ahmed Amine Khamlichi pour m’avoir accueillie dans son laboratoire, pour m’avoir encadrée et conseillée tout au long de mon doctorat.
Je tiens également à remercier le Professeur Catherine Muller pour avoir accepté de présider le jury au cours de ma soutenance, ainsi que les Docteurs Michèle Goodhardt, Pierre Ferrier et les Professeurs Ahmad Oulmouden et Michel Cogné pour avoir accepté de juger mon travail.
Je remercie également les membres de l’équipe, Nadine pour ses précieux conseils, Dania pour m’avoir épaulée, et Nathalie pour sa bonne humeur ainsi que pour s’être occupée avec soin de mes souris.
Je remercie l’équipe de l’animalerie de l’IPBS: Laurence, Magalie J, Pascale, Dimitri, Magalie P, Jean-Michel, Serge, Olivier, Jean-Pierre, qui ont également pris soin de mes souris.
Je souhaite remercier Zdenko Herceg pour nous avoir permis d’initier notre travail sur TRRAP en nous faisant don de ses souris.
Je remercie Fatima et Camille, de la plateforme de tri de Purpan, pour leur sympathie ainsi que pour avoir passé de longues heures afin de me permettre de réaliser mes tris cellulaires.
Je remercie le Professeur Christophe Cazaux qui a été le premier à m’accueillir à l’IPBS au cours de mon M2R et qui a soutenu mon choix de rejoindre l’équipe du Docteur Khamlichi.
Je souhaite remercier tout particulièrement Fanny, Oriane et Corinne pour m’avoir aidée et soutenue tout au long de ces trois années et pour tous les bons moments que nous avons passés enssemble. Ainsi que tous les collègues de l’IPBS, Denis, Marie-Jeanne, Valérie, Stéphanie, Pierre, Camille, Sophie, Leyre, Lara, Ludivine, Marielle, Luc, Sébastien, Giada, Eddy, Frédérique, Anne, Sandrine, Mathieu, Emilyne, Rémy, Romain, Laurie… pour tous les bons
4 moments que nous avons partagés et toutes les discussions enrichissantes scientifiques et moins scientifiques que nous avons eues.
Je remercie ma famille pour leur soutien inconditionnel tout au long de ma vie et particulièrement durant ces années de doctorat, ainsi que pour m’avoir permis de poursuivre mes études aussi loin que je souhaitais aller.
Enfin, merci à Sébastien qui m’a toujours soutenue pendant ces années de thèse et qui a su me donner des conditions de travail hors paire avec entre autres ses fabuleux petits plats.
5
Sommaire
I.
Structure et fonction des Immunoglobulines ... 9
A.
La chaîne lourde ... 10
B.
La chaîne légère ... 10
II.
Développement du Lymphocyte B ... 11
A.
Engagement dans la voie B : Pré-pro-B ... 12
B.
Pro-B ... 12
C.
Pré-B ... 14
D.
B immature ... 14
E.
B transitionnel ... 15
F.
B mature ... 15
G.
Plasmocyte ... 17
III.
Modifications de la chromatine ... 18
A.
Modifications de la chromatine liées à la transcription... 18
1. Modifications post-traductionnelles des histones ... 18
2. Les facteurs de remodelage ... 21
B.
Modifications de la chromatine liées aux cassures double brin ... 22
1. Détection et signalisation de la cassure ... 22
2. Facteurs de remodelage ... 24
IV.
Réparation des cassures double brin de l’ADN par NHEJ ... 26
A.
Détection et signalisation des cassures ... 26
B.
Maintien et protection des extrémités ... 28
1. Maturation des extrémités ... 29
6
V.
Mécanismes de diversification des immunoglobulines ... 31
A.
Recombinaison V(D)J ... 31
1. Mécanisme ... 31 2. Régulation de la recombinaison V(D)J ... 34B.
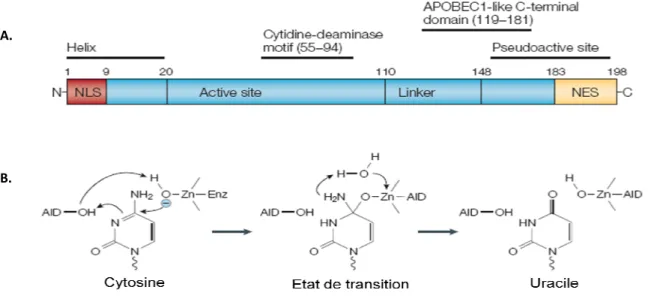
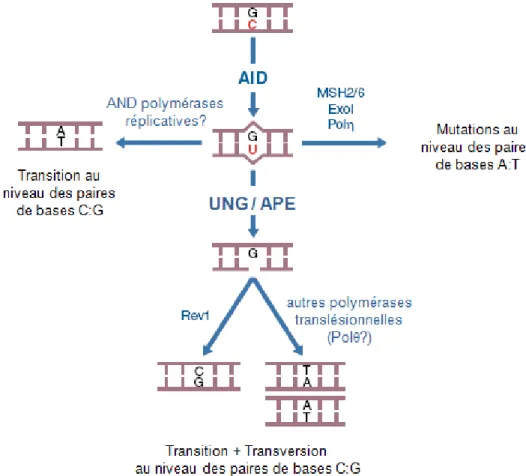
L’enzyme AID ... 38
1. Mode d’action ... 39 2. Ciblage ... 40 3. Régulation ... 41C.
L’hypermutation somatique (SHM) ... 44
1. Mécanisme ... 44 2. Ciblage et régulation ... 46D.
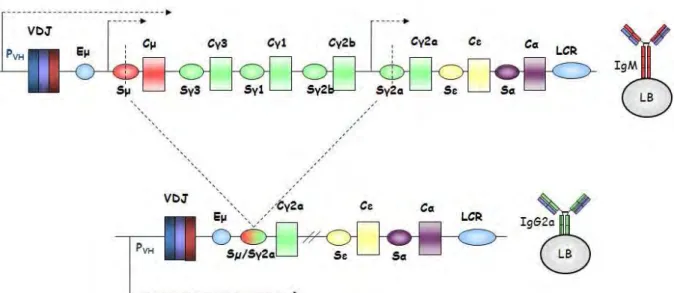
La commutation isotypique ... 47
1. Fonctions des différents isotypes ... 48
2. Les séquences Switch ... 48
3. La transcription germinale ... 50
4. Modifications de la chromatine liées au Switch ... 51
5. Induction de cassures double brin et Jonction ... 52
VI.
Rôle du MMR dans les mécanismes de diversification ... 58
A.
Mécanisme du MMR ... 58
B.
MMR et hypermutation somatique ... 59
C.
MMR et commutation isotypique ... 60
1. Impact de la délétion des protéines du MMR sur le Switch ... 60
2. MMR dans les jonctions des séquences S... 61
3. Rôle du MMR dans le ciblage de Sµ ... 61
VII.
Les éléments régulateurs du locus IgH ... 63
A.
L’enhancer intronique Eµ ... 63
7
C.
La région régulatrice en 3’RR ... 65
VIII.
TRRAP ... 67
A.
Structure ... 67
B.
TRRAP et les HATs ... 68
C.
Rôle dans le développement ... 69
D.
Rôle dans différents processus cellulaires ... 70
1. Transcription ... 71 2. Réparation ... 71 3. Réplication ... 73
Manuscrit 1 ... 75
Manuscrit 2 ... 103
Discussion et perspectives ... 122
Délétion de la protéine TRRAP dans les cellules B ... 122
Délétion de la protéine MSH2 en l’absence de la région Sµ ... 125
8
Introduction
Après un bref rappel des étapes du développement du lymphocyte B au sein de l’organisme, je présenterai différentes modifications de la chromatine liées à la transcription et aux cassures double brin (cdb) de l’ADN pouvant intervenir au sein des loci des immunoglobulines. Je présenterai plus en détail la réparation des cdb notamment par le mécanisme de NHEJ (Non Homologous End Joining). Les trois processus essentiels à la production et la diversification des immunoglobulines, la recombinaison V(D)J, l’hypermutation somatique et la commutation isotypique, seront également abordés dans cette introduction et plus particulièrement le rôle du mécanisme de réparation des mésappariements (MMR). Enfin j’orienterai mon introduction sur la protéine TRRAP impliquée à la fois dans la transcription et la réparation des cdb.
Afin de contrer les différents pathogènes (bactéries, virus, parasites) ou certaines cellules malignes, l’organisme s’est doté d’un système immunitaire extrêmement développé qui se décompose en deux parties. La réponse immunitaire innée faisant intervenir les phagocytes (monocytes, macrophages, neutrophiles, cellules NK), et la réponse immunitaire adaptative dont les acteurs sont les Lymphocytes T (réponse à médiation cellulaire) et les Lymphocytes B (réponse à médiation humorale). Les lymphocytes B expriment à leur surface un récepteur appelé immunoglobuline (Ig), anticorps (Ac) ou BCR (B Cell Receptor) qui leur permet dans un premier temps de reconnaître spécifiquement un antigène et dans un deuxième temps de s’activer en cellules sécrétrices d’anticorps qui vont alors lier l’antigène et entraîner son élimination ou se différencier en cellule B mémoires et assurer une défense continue de l’organisme.
10
A.
La chaîne lourde
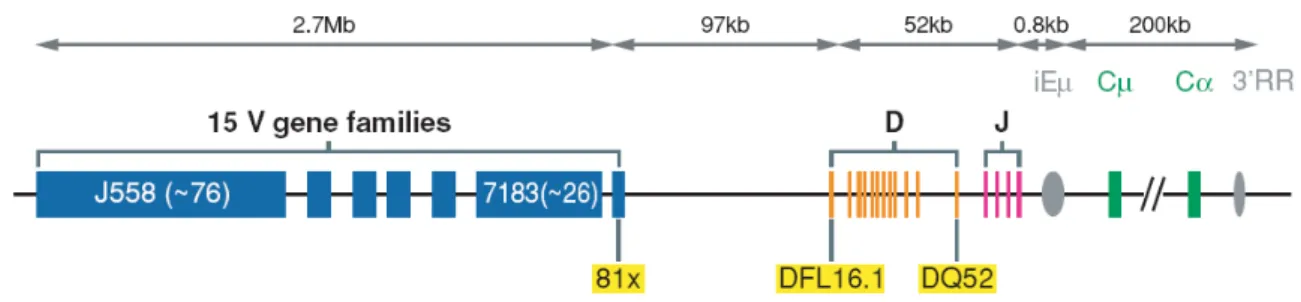
Chez la souris, le locus codant la chaîne lourde (IgH) est localisé sur le chromosome 12 (chromosome 14 chez l’homme) et s’étend sur environ 3Mb. Il est constitué de plusieurs centaines des segments VH (Variable) regroupés en 15 familles qui s’étendent sur 2,7Mb, d’une douzaine de segments DH (Diversity, propres au locus IgH) et de quatre segments JH (Joining) suivis de huit gènes codant les régions constantes. Les différentes régions constantes CH sont µ, δ, γ3, γ1, γ2b, γ2a, ε et α, chacune comporte les différents exons codant pour les domaines structuraux de chaque chaîne lourde (figure 2).
Figure 2: Organisation génomique du locus IgH
Les distances représentées correspondent à la lignée murine 129 et peuvent varier d’une lignée à l’autre (Jung et al, 2006)
B.
La chaîne légère
Chez la souris, il existe deux types de chaînes légères définies par leur région constante : κ et λ. Les loci codant les chaînes légères sont situés sur le chromosome 6 pour Igκ (chromosome 2 chez l’homme) et sur le chromosome 16 pour Igλ (chromosome 22 chez l’homme).
Le locus κ comporte 140 segments Vκ et 5 segments Jκ qui constituent, après réarrangement, la région variable de la chaîne légère, suivie d’un unique gène constant Cκ. Les gènes λ sont répartis en quatre familles comprenant une paire de segments Jλ et Cλ numérotés de 1 à 4. Ce locus ne contient que 3 segments Vλ et n’est utilisé pour constituer une Ig que dans 5% des cas chez la souris.
II.
Développement du Lymphocyte B
Au sein de toutes les cellules de l’organisme les gènes des
configuration germinale. Ce n’est qu’au sein des lymphocytes B que ces gènes subisse différents réarrangements qui permettent
Le développement des cellules B a lieu dans la moelle osseuse et se poursuit dans les organes lymphoïdes secondaires (rate, ganglions lymphatiques, amygdales, plaques de Peyer). Il se décompose en deux grandes phases
moelle osseuse, au cours de laquelle le lymphocyte B naissant constitue une première phase dépendante de l’antigène, dans les organes lymphoïdes secondaire d’élaborer des Ig toujours plus performantes (figure 3).
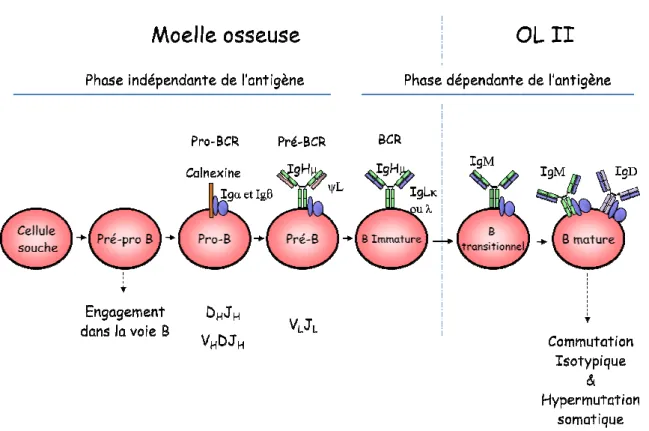
Figure 3: Développement des lymphocytes B
Représentation des différents stades de développement des cellules B avec les différents réarrangements ayant lieu à chaque stade et l’expression des pro
Développement du Lymphocyte B
Au sein de toutes les cellules de l’organisme les gènes des Ig se trouvent dans leur configuration germinale. Ce n’est qu’au sein des lymphocytes B que ces gènes subisse différents réarrangements qui permettent l’expression du BCR.
Le développement des cellules B a lieu dans la moelle osseuse et se poursuit dans les organes lymphoïdes secondaires (rate, ganglions lymphatiques, amygdales, plaques de Peyer). Il
mpose en deux grandes phases : une phase indépendante de l’antigène au cours de laquelle le lymphocyte B naissant constitue une première dépendante de l’antigène, dans les organes lymphoïdes secondaire
toujours plus performantes (figure 3).
: Développement des lymphocytes B
Représentation des différents stades de développement des cellules B avec les différents réarrangements ayant stade et l’expression des pro-BCR, pré-BCR et BCR (adapté de Meffre et al, 2000).
11 se trouvent dans leur configuration germinale. Ce n’est qu’au sein des lymphocytes B que ces gènes subissent
Le développement des cellules B a lieu dans la moelle osseuse et se poursuit dans les organes lymphoïdes secondaires (rate, ganglions lymphatiques, amygdales, plaques de Peyer). Il : une phase indépendante de l’antigène au sein de la au cours de laquelle le lymphocyte B naissant constitue une première Ig, et une dépendante de l’antigène, dans les organes lymphoïdes secondaires, qui permet
Représentation des différents stades de développement des cellules B avec les différents réarrangements ayant de Meffre et al, 2000).
12
A.
Engagement dans la voie B : Pré-pro-B
Dans la moelle osseuse, les cellules souches débutant leur différenciation en lymphocytes B expriment à leur surface les marqueurs B220 (CD45R), ce marqueur est spécifique des B et sera exprimé tout au long de la vie de la cellule. Les pré-pro-B expriment également c-kit (CD117) et CD43 (Hardy and Hayakawa 1991; Rolink et al. 1996). A ce stade les gènes des Ig se présentent encore sous leur configuration germinale. Les gènes Rag-1 et Rag-2, permettant l’expression de l’endonucléase nécessaire aux réarrangements, sont très faiblement exprimés (Schatz et al. 1989; Oettinger et al. 1990).
B.
Pro-B
Le stade pro-B se caractérise tout d’abord par l’apparition de CD19 à la surface des cellules, l’expression de ce marqueur augmentera tout au long du développement (Zhou et al. 1991; Krop et al. 1996). L’expression du complexe RAG et de la Terminal-désoxynucléotidyl-Transférase (TdT) est plus importante à ce stade. Les premiers réarrangements ont lieu au sein des gènes des Ig, entre les segments DH et JH du locus des chaînes lourdes.
A ce stade l’hétérodimère Igα/Igβ se trouve à la surface des cellules en association avec la calnexine, c’est le pro-BCR (Nagata et al. 1997). Il arrive qu’une protéine µ tronquée appelée Dµ, dépourvue de région variable, soit exprimée (Reth and Alt 1984). Dans ces conditions le développement du lymphocyte est bloqué (Gong and Nussenzweig 1996) par un signal négatif en provenance d’Igα/Igβ via leurs motifs ITAM (Immunoreceptor Tyrosine-based Activation Motif).
A la suite de la recombinaison donnant naissance à un segment DJH, les segments VH deviennent accessibles pour la recombinaison. Le réarrangement VH à DJH a lieu, permettant d’obtenir la région variable complète VDJH de la chaîne lourde. C’est également à ce stade qu’à lieu l’exclusion allélique : le phénomène permettant à un lymphocyte B d’exprimer à sa surface un seul type d’Ig capable de reconnaître un seul et unique antigène. En effet, dans 99% des cellules B la chaîne lourde est codée par un seul des deux allèles du locus IgH et la chaîne légère
13 par un seul allèle des différents loci IgL (Pernis et al. 1965; Cebra et al. 1966). Toutefois le mécanisme de l’exclusion reste encore incertain.
Deux modèles sont toujours d’actualité, le modèle non régulé ou stochastique et le modèle régulé (Gorman and Alt 1998). Un autre modèle avait été proposé selon lequel l’expression de deux chaînes lourdes issues des deux allèles serait toxique pour la cellule. Des expériences menées sur la souris, au sein de laquelle les deux allèles du locus IgH ont été modifiés pour exprimer deux chaînes lourdes différentes, viennent contredire ce modèle. Les lymphocytes B de ces souris expriment les deux types de chaînes lourdes (Sonoda et al. 1997).
Le modèle stochastique stipule que le réarrangement productif des deux allèles d’un locus est possible mais que la probabilité que cet évènement se produise est extrêmement faible (Coleclough et al. 1981; Liang et al. 2004). Le choix du réarrangement d’un des deux allèles se ferait au hasard. Par la suite, le réarrangement d’un allèle pourrait recruter et titrer les protéines impliquées dans le processus et empêcher le remaniement du second allèle (Liang et al. 2004). Dans le cas du locus IgH, le réarrangement DH à JH a lieu sur les deux allèles, en revanche le réarrangement de VH à DHJH se ferait de façon moins efficace et ne ciblerait le plus souvent que l’un des deux allèles.
Le modèle régulé semble le plus évident, soutenu par les études montrant que l’expression de transgènes de chaînes lourdes ou légères inhibe les réarrangements endogènes (Ritchie et al. 1984; Rusconi and Kohler 1985; Weaver et al. 1985; Nussenzweig et al. 1987; Manz et al. 1988). Dans le cas de la chaîne lourde, une étude démontre que la délétion du domaine transmembranaire du gène µ entraîne les réarrangements productifs des deux allèles (Kitamura et al. 1991; Kitamura and Rajewsky 1992). Le modèle régulé implique donc que le réarrangement productif d’un allèle et son expression à la surface de la cellule entraîne l’inhibition du réarrangement du second allèle par rétrocontrôle négatif (Alt et al. 1980; Alt et al. 1984). Si le premier réarrangement n’est pas productif alors le réarrangement du second allèle a lieu. C’est le réarrangement de VH à DJH qui semble être l’étape régulée au sein du locus IgH, le réarrangement DH à JH se faisant sur les deux allèles. Deux types de B coexistent alors dans l’organisme, certains avec un seul allèle réarrangé en totalité (~60%) et d’autres avec deux allèles réarrangés dont un seul est productif (~40%) (Alt et al. 1986; Mostoslavsky et al. 2004).
14
C.
Pré-B
A ce stade l’expression de CD43 diminue, celle de c-kit disparaît tandis que celle de CD19 augmente et celle de CD25 apparaît (Rolink et al. 1994). La transcription de la chaîne lourde, devenue possible après réarrangement, permet au pré-B d’exprimer à sa surface le pré-BCR constitué de deux chaînes lourdes µ en association avec Igα/Igβ et deux pseudo-chaînes légères (V-Pre-B ou λ5) (Karasuyama et al. 1990; Tsubata and Reth 1990).
L’expression du pré-BCR entraîne l’expansion clonale des pré-B avant la recombinaison entre les segments VL et JL dans les gènes des chaînes légères. Les pseudo-chaînes légères sont alors remplacées par Igκ ou Igλ.
D.
B immature
Le lymphocyte B immature exprime donc à sa surface une IgM complète et n’exprime désormais plus CD43. C’est à ce stade que se fait la sélection positive et négative de l’anticorps par l’antigène. En effet, un lymphocyte auto-réactif, c'est-à-dire capable de reconnaître un antigène du soi via son BCR, est sujet à l’édition du récepteur (Tiegs et al. 1993; Hertz and Nemazee 1997; Melamed and Nemazee 1997; Pelanda et al. 1997). Au cours de ce processus, la poursuite de l’expression de RAG entraîne des réarrangements secondaires qui modifient les régions variables au niveau des loci Igκ en priorité puis Igλ ou plus rarement IgH (Nemazee 2000; Nemazee and Weigert 2000). Dans le cas des lymphocytes auto-réactifs l’édition aurait lieu après réarrangement de la chaîne légère (Chen et al. 1995). Ce mécanisme a été mis en évidence grâce à l’étude de souris transgéniques dont les cellules B expriment un anticorps auto-réactif (Gay et al. 1993; Tiegs et al. 1993). Chez ces souris, le BCR est modifié afin d’induire une tolérance au soi.
Si l’édition ne permet pas de corriger l’auto-réactivité, alors le lymphocyte B est éliminé par apoptose (Hartley et al. 1993; Norvell et al. 1995) ou subit l’anergie (Pike et al. 1982; Goodnow et al. 1988). L’anergie est un phénomène physiologique par lequel la cellule B n’est pas éliminée mais inactivée, les cellules anergiques présentent une faible espérance de vie (Bell et al. 1994; Fulcher and Basten 1994).
E.
B transitionnel
Une fois la sélection terminée les lymphocytes B quittent la moelle osseuse, ces cellules sont alors qualifiées de B transitionnel
plus faible expression de B220 et IgD des ARN codant RAG (Monroe et al.
Elles vont transiter dans l’organisme avant de s’accumuler dans les organes lymphoïdes périphériques pour devenir des lymphocytes B matures.
F.
B mature
La rate constitue un des principaux organes lymphoïdes seco
en différents compartiments permettant une rencontre efficace de l’antigène et la mise en place de la réponse immunitaire
La rate est extrêmement vascularisée pour permettre
Cette vascularisation est notamment localisée dans la pulpe rouge qui contient également des cellules présentatrices d’antigènes (cellules dendritiques et macrophages). Les follicules sont enrichis en cellules B (IgMlow
B transitionnel
Une fois la sélection terminée les lymphocytes B quittent la moelle osseuse, ces cellules B transitionnelles. Ils se différencient des cellules B matures par une plus faible expression de B220 et IgD (Carsetti et al. 1995) et par une persistance de l’expression
(Monroe et al. 1999; Yu et al. 1999; Meffre et al. 2000)
Elles vont transiter dans l’organisme avant de s’accumuler dans les organes lymphoïdes périphériques pour devenir des lymphocytes B matures.
La rate constitue un des principaux organes lymphoïdes secondaires,
s compartiments permettant une rencontre efficace de l’antigène et la mise en place de la réponse immunitaire (pour revue Batista and Harwood 2009) (figure 4).
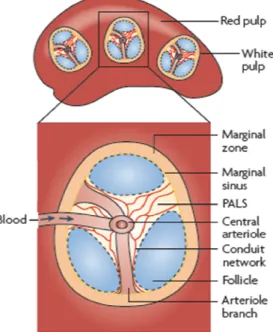
Figure 4 : Organisation de la rate
La rate s’organise autour d’
une gaine lymphoïde périatériolaire (PALS) correspondant à la pulpe blanche,
follicules entourés de la zone marginale trouvant au centre de la pulpe rouge. Harwood 2009)
La rate est extrêmement vascularisée pour permettre l’apport d’antigènes exogènes vascularisation est notamment localisée dans la pulpe rouge qui contient également des cellules présentatrices d’antigènes (cellules dendritiques et macrophages). Les follicules sont
low
IgDHICD23Hi CD21low) et en cellules folliculaires dendritiques. Les
15 Une fois la sélection terminée les lymphocytes B quittent la moelle osseuse, ces cellules Ils se différencient des cellules B matures par une et par une persistance de l’expression 1999; Yu et al. 1999; Meffre et al. 2000).
Elles vont transiter dans l’organisme avant de s’accumuler dans les organes lymphoïdes
ndaires, elle est organisée s compartiments permettant une rencontre efficace de l’antigène et la mise en
(figure 4). : Organisation de la rate
La rate s’organise autour d’une artériole et contient une gaine lymphoïde périatériolaire (PALS) rrespondant à la pulpe blanche, plusieurs icules entourés de la zone marginale le tout ce la pulpe rouge. (Batista and
l’apport d’antigènes exogènes. vascularisation est notamment localisée dans la pulpe rouge qui contient également des cellules présentatrices d’antigènes (cellules dendritiques et macrophages). Les follicules sont ) et en cellules folliculaires dendritiques. Les
16 lymphocytes T sont localisés dans la gaine périartériolaire qui contient de nombreuses cellules présentatrices d’antigènes et un réseau de fibres de collagène permettant le passage de petites molécules comme les chimiokines. La zone marginale, fortement vascularisée, contient également des cellules présentatrices d’antigènes et d’une population importante de cellules B (IgMhi IgDlowCD23low CD21hi).
Dans un premier temps la cellule B est qualifiée de naïve lorsqu’elle n’a pas encore rencontré d’antigène exogène. Les lymphocytes B peuvent entrer directement en contact avec les antigènes solubles de petite taille, mais les antigènes de taille plus importante doivent être présentés aux lymphocytes B par les macrophages ou les cellules dendritiques. Lorsque les cellules B ne reçoivent pas l’aide de cellules T (réponse T-indépendante) elles se différencient en plasmocytes extra-folliculaires et produisent rapidement les anticorps dirigés contre l’antigène. Les lymphocytes B de la zone marginale semblent particulièrement impliqués dans cette réponse T-indépendante (Lopes-Carvalho et al. 2005). D’une autre façon, les cellules B activées peuvent interagir avec les lymphocytes T (réponse T-dépendante). Les cellules B migrent alors au centre des follicules, entrent dans une nouvelle phase d’expansion clonale et forment les centres germinatifs (CG) (Allen et al. 2007).
Brièvement, les CG, découverts en 1884 par Walter Flemming, contiennent de nombreuses cellules dendritiques présentatrices d’antigènes. Ils se décomposent en deux zones : une « zone claire » et une « zone sombre » du fait de leur apparence histologique. Les lymphocytes B de la zone sombre sont les centroblastes, ceux de la zone claire sont les centrocytes. Les cellules dendritiques sont localisées principalement au niveau de la zone claire afin de capter plus facilement les antigènes tandis que la zone sombre est constituée en grande majorité de lymphocytes empaquetés. Les lymphocytes B sont très mobiles au sein de ces structures, les cellules de la zone sombre migrent vers la zone claire et vise versa (Allen et al. 2007; Hauser et al. 2007; Schwickert et al. 2007). Bien que la zone sombre ait été définie comme la zone de prolifération (Liu et al. 1991; MacLennan 1994), où les centroblastes seraient les précurseurs des centrocytes (Hollowood and Goodlad 1998; Haberman and Shlomchik 2003; McHeyzer-Williams 2003), des études ont démontré que les cellules B prolifèrent également dans la zone claire (Rahman et al. 2003; Wang and Carter 2005). Les CG sont le siège de
17 l’hypermutation somatique et de la commutation isotypique. L’hypermutation cible les centroblastes au sein de la zone sombre tandis que la commutation a lieu dans les centrocytes au sein de la zone claire (pour revue (Rajewsky 1996).
Au cours du processus de commutation la région Cµ est remplacée par une région constante située en aval. Le BCR est alors constitué de chaînes lourdes possédant une nouvelle région constante lui conférant de nouvelles propriétés effectrices. Le mécanisme d’Hypermutation Somatique permet d’introduire des mutations dans les régions variables et modifie l’affinité du BCR pour l’antigène. Par la suite, la sélection des cellules en fonction de l’affinité de leur BCR se fait dans la zone claire, où les cellules interagissent notamment avec les cellules dendritiques (MacLennan 1994). Les lymphocytes qui ne sont plus capables de reconnaître l’antigène ou dont l’affinité a diminué sont éliminés.
Après la réponse immunitaire les CG disparaissent, les LB de haute affinité produits au sein de ces structures sont conservés dans l’organisme et possèdent une durée de vie extrêmement longue, ce sont les Lymphocytes B « mémoires ». Lors d’une prochaine rencontre avec l’antigène, ces cellules permettront d’effectuer une réponse immunitaire bien plus rapide et plus efficace que la première.
G.
Plasmocyte
Les plasmocytes ont deux origines différentes : il y a tout d’abord les LB activés n’ayant pas initié la formation de CG et certains LB possédant une forte affinité issus des CG, les premiers ayant une durée de vie plus courte que les seconds. C’est le stade final du développement de la cellule B, ces cellules sont des « usines à anticorps ». En effet les Ig produites en masse par les plasmocytes sont sécrétées dans l’organisme où elles se lient à l’antigène pour faciliter son élimination par les phagocytes. Les plasmocytes se caractérisent également par leur incapacité à proliférer. A la suite de la réponse immunitaire les cellules à longue durée de vie migrent vers la moelle osseuse où elles peuvent survivre pendant plusieurs mois permettant ainsi une défense continue de l’organisme (McHeyzer-Williams 2003).
18
III.
Modifications de la chromatine
L’information génétique, au sein des cellules, est compactée sous forme de chromatine, qui est constituée d’une succession de nucléosomes. Un nucléosome contient un octamère d’histones : H2A, H2B, H3 et H4 (deux molécules de chaque), entouré par la molécule d’ADN et de l’histone H1 qui stabilise l’assemblage. La chromatine se présente sous deux formes, une forme ouverte et accessible aux protéines, l’euchromatine, et une forme hyper-compactée et moins accessible, l’hétérochromatine. La dynamique de la chromatine est essentielle dans les évènements cellulaires de réplication, réparation, recombinaison, transcription et progression dans le cycle cellulaire (Groth et al. 2007; Kouzarides 2007). Le passage de la chromatine d’une conformation à l’autre se fait grâce à des modifications post-traductionnelles des histones.
Les mécanismes permettant l’élaboration des Ig font intervenir différents processus qui seront détaillés par la suite. Ces processus nécessitent une transcription active et la création de cdb qui requièrent un remodelage important de la chromatine. Les modifications de la chromatine associées à ces processus sont trop nombreuses pour être présentées dans cette introduction. Je présenterai donc les principales modifications qui ont été étudiées dans le locus IgH.
A.
Modifications de la chromatine liées à la transcription
1. Modifications post-traductionnelles des histones
La transcription est associée à diverses modifications post-traductionnelles des histones (tableau ci-dessous).
Modifications de la chromatine associées à la transcription.
PTMs (post-translational modifications), § concerne la levure
¶ concerne les mammifères (Berger 2007)
19 a) Méthylation des histones et de l’ADN
La méthylation de l’ADN est généralement associée à une inactivation des gènes. La méthylation de l’ADN a lieu au niveau de dinucléotides, dits CpG, regroupés en îlots, régulièrement retrouvés au niveau des promoteurs. La répression de la transcription par la méthylation de l’ADN s’opère en empêchant l’interaction de la machinerie transcriptionnelle ou les facteurs de transcription avec les cytosines méthylées. Cette modification de l’ADN joue un rôle important dans l’activation et la répression des gènes de façon tissu-spécifique (Bird and Wolffe 1999). Ainsi, elle collabore avec les modifications des histones pour moduler la transcription.
En règle générale, la méthylation des lysines 4, 36 et 79 de l’histone H3 est associée à l’activation de la transcription. A l’inverse, la méthylation des lysines 9 et 27 de H3 ainsi que de la lysine 20 de H4 corrèle davantage avec la répression transcriptionnelle (Jenuwein and Allis 2001) La méthylation est ajoutée sur les lysines par les histone-méthyltransférases (HMTs), qui peuvent produire différents états de méthylation (mono, di ou tri), chaque état pouvant avoir une fonction différente (Schotta et al. 2008). La méthylation ne modifie pas la charge des protéines mais augmente progressivement leur masse et leur hydrophobicité occasionnant ainsi l’altération des liaisons hydrogènes intra ou inter-moléculaires. Ce phénomène pourrait alors créer de nouveaux sites d’interaction au sein de l’histone méthylée, inhiber les interactions protéiques ou encore modifier directement la structure des nucléosomes, modulant ainsi leur accessibilité (Lee et al. 2005)
b) Acétylation des histones
L’acétylation des histones H3 et H4 est un interrupteur central dans l’activation et la répression transcriptionnelle de la chromatine. L’acétylation des histones est importante dans les mécanismes qui affectent les loci Ig. Elle a, dès le départ, été liée à la transcription (Allfrey et al. 1964; Pogo et al. 1966; Brownell and Allis 1996; Struhl 1998) mais ces dernières années son implication a également été évoquée dans la réparation, l’épissage et la progression dans le cycle cellulaire (Carrozza et al. 2003a). Cette modification est catalysée par les histone-acétyltransférases (HATs) qui transfèrent un groupement acétyle sur les lysines contenues dans
20 les queues N-terminales des histones (Grunstein 1997) (Figure 5). L’addition d’un groupement acétyle au sein d’une lysine neutralise la charge positive de celle-ci, diminuant ainsi son interaction avec l’ADN. La structure de la chromatine se trouve alors relaxée et devient accessible à la machinerie de transcription (Workman and Kingston 1998). D’une part, l’acétylation des histones pourrait faciliter le recrutement de régulateurs transcriptionnels (Dhalluin et al. 1999; Owen et al. 2000; Hassan et al. 2002; de la Cruz et al. 2005), d’autre part, elle pourrait s’inscrire dans le contexte de « code des histones », en association avec d’autres modifications, pour permettre un signal bien particulier (Strahl and Allis 2000; Jenuwein and Allis 2001).
Transcription OFF Transcription ON
Figure 5 : Passage de la chromatine fermée à la chromatine ouverte après acétylation
Les lysines sont acétylées par les HATs à partir de l’acétylCoA, le retour à une chromatine fermée se fait après désacétylation par les HDAC.
(Adapté de Verdone et al, 2005)
Il existe plusieurs superfamilles d’HATs, les deux familles le plus étudiées sont la famille des GNAT (gcn5 related N-acétyltransférases) et la famille des MYST (MOZ, Ybf2/Sas3, Sas2, et Tip60) (Marmorstein 2001). La plupart de ces enzymes se présentent sous forme de complexes multiprotéiques, composés de sous-unités catalytiques et de sous-unités régulatrices qui assureraient le ciblage et la régulation de l’activité de l’enzyme.
Ces complexes étant impliqués dans de nombreux mécanismes cellulaires, ils peuvent participer au processus de tumorigénèse et constituent alors d’éventuelles cibles médicamenteuses (Dekker and Haisma 2009).
21
2. Les facteurs de remodelage
Les facteurs de remodelage de la chromatine sont des complexes multiprotéiques contenant une sous-unité catalytique appartenant à la superfamille des ATPases SWI2/SNF2. Cette superfamille comporte quatre classes de facteurs de remodelage : SWI/SNF, ISWI, CHD et INO80 (van Attikum and Gasser 2005a). Les facteurs SWI/SNF contiennent un bromodomaine reconnaissant les histones acétylées. Ils sont capables d’induire un espacement entre deux nucléosomes, mais également de séparer les nucléosomes de l’ADN entraînant leur désorganisation (Lorch et al. 1999; Saha et al. 2006). Ces facteurs sont recrutés au niveau des promoteurs en cours d’activation, via leur interaction avec les histones acétylées, probablement pour parachever l’initiation de la transcription. Les facteurs ISWI possèdent un domaine de liaison à l’ADN et réorganisent les nucléosomes au sein des régions inactives en transcription (Ito et al. 1997; Langst et al. 1999) en insérant un espacement entre deux nucléosomes (Kagalwala et al. 2004; Saha et al. 2006). Les membres de la famille CHD se lient à l’ADN et reconnaissent les histones méthylées via leur chromodomaine. Ces facteurs sont ainsi recrutés au niveau de promoteurs actifs ou inactifs et permettent le remodelage de la chromatine (Hall and Georgel 2007). Les facteurs INO80 contiennent une insertion séparant en deux la région ATPase (van Attikum and Gasser 2009). De la même façon que les facteurs de la famille ISWI, ils font glisser les nucléosomes et rendent ainsi l’ADN accessible (Morrison and Shen 2009). Des études plus récentes démontrent que, le facteur SWR1, appartenant à la famille INO80 est capable de promouvoir des échanges de dimères d’histones déstabilisant ainsi la composition des nucléosomes (Mizuguchi et al. 2004).
Le remodelage de la chromatine se fait au niveau de l’organisation des nucléosomes et débute systématiquement par une altération du contact entre les histones et l’ADN. Les facteurs de remodelage utilisent l’énergie de l’hydrolyse de l’ATP pour provoquer cette altération conduisant à la réorganisation et à la mobilisation des octamères d’histones en cis (sur l’ADN adjacent) et en trans (d’une molécule d’ADN à une autre). Globalement, les facteurs de remodelage sont recrutés au niveau des promoteurs et facilitent ainsi l’accès à la machinerie de transcription.
22
B.
Modifications de la chromatine liées aux cassures double brin
Les processus de recombinaison V(D)J et de commutation isotypique nécessitent la formation de cdb. Ces cassures sont associées à des modifications intenses de la chromatine qui permettent leur signalisation et leur réparation (Downs 2007).
1. Détection et signalisation de la cassure
La détection de la cassure est une étape essentielle dans la réparation. Le complexe MRN, présenté plus en détail par la suite, assume ce rôle de reconnaissance de façon très précoce (Lee and Paull 2005). Plusieurs protéines s’assemblent ensuite au niveau de la cassure et altèrent la structure de la chromatine.
a) Phosphorylation
Les histones H1 et H2AX sont phosphorylées en réponse aux cdb. La plus étudiée est le variant de H2A : H2AX. H2AX peut être phosphorylée au niveau de la sérine 139 (γH2AX). C’est l’une des premières cibles de la sérine/thréonine kinase ATM (ataxia-telangiectasia mutated), responsable de la reconnaissance précoce des cdb (Redon et al. 2002). γH2AX intervient quelques minutes après apparition de la cdb et s’étend jusqu’à 2Mb autour de la cassure (Rogakou et al. 1999). Ainsi, γH2AX agirait comme un signal capable d’induire le recrutement des protéines de recombinaison et de réparation. Pour exemple la protéine MDC1, définie comme un régulateur important dans la restructuration de la chromatine, se lie au niveau des cdb de façon γH2AX-dépendante (Lukas et al. 2004). Toutefois, bien que γH2AX occupe une position centrale dans le bon déroulement de la réparation des cdb elle ne participe pas à la reconnaissance initiale de la cassure (Celeste et al. 2003).
L’histone H2B est rapidement phosphorylée au niveau de la sérine 14 en réponse aux cdb. Elle agit ainsi de concert avec γH2AX pour signaler la cassure et recruter les protéines de la réparation (Fernandez-Capetillo et al. 2004).
L’histone H4 est également phosphorylée, au niveau de la sérine 1, en réponse aux cdb. H4 est modifiée par la Caséine kinase 2, cependant, cette modification empêche son acétylation et pourrait davantage intervenir après réparation pour rétablir la structure initiale de la chromatine (Cheung et al. 2005; Utley et al. 2005).
23 b) Acétylation
Les HATs jouent un rôle essentiel dans la réparation des cdb en participant au remodelage de la chromatine facilitant la réparation. En l’occurrence, TIP60 acétyle les histones H2A, H3 et H4 (Yamamoto and Horikoshi 1997). L’histone H4 a particulièrement été étudiée et différents travaux démontrent son importance dans le maintien de l’intégrité du génome, les mutations affectant sa partie N-Terminale entraînent de l’instabilité génétique et un arrêt du cycle cellulaire (Megee et al. 1990; Durrin et al. 1991; Morgan et al. 1991; Megee et al. 1995). L’acétylation permet de relaxer la chromatine créant ainsi une plateforme d’accès et de recrutement des facteurs de remodelage de la chromatine (Pandita and Richardson 2009).
c) Méthylation
La diméthylation de la lysine 20 de l’histone H4 (H4K20me2) permet également le recrutement des protéines de la réparation au niveau des cdb. En effet, 53BP1 présente un domaine interagissant de façon spécifique avec cette modification (Botuyan et al. 2006). De même, la méthylation de la lysine 79 de l’histone H3 (H3K79me) pourrait participer à l’interaction avec 53BP1 mais ce rôle reste encore incertain (Huyen et al. 2004).
d) Ubiquitination
L’ubiquitination est une des dernières modifications d’histone identifiées en rapport avec les cdb et se fait au niveau des histones H2AX, H2A et H2B par l’ubiquitine ligase RNF8 (Ma et al. 2002a; Huen et al. 2007). RNF8 est recrutée sur le site de la cassure et s’accumule via son interaction avec la protéine MDC1 (Huen et al. 2007; Kolas et al. 2007; Wu et al. 2008a). Une seconde ubiquitine ligase, UBC13, a été identifiée comme étant impliquée dans l’ubiquitination des histones H2A et H2AX (Zhao et al. 2007). Toutefois UBC13 semble davantage impliquée dans la réparation par recombinaison homologue que par NHEJ.
Le rôle de l’ubiquitination des histones serait, encore une fois, de faciliter le recrutement et le maintien des protéines de la réparation au niveau des cdb, en particulier BRCA1 et 53BP1 (Kolas et al. 2007; Mailand et al. 2007; Wang et al. 2007). Ces deux protéines, pourtant liées à la réparation par recombinaison homologue, ont été définies ces dernières années comme étant
24 impliquées dans le mécanisme de NHEJ (Dimitrova et al. 2008; Wei et al. 2008; Poplawski et al. 2009).
Pour conclure, l’organisation nucléaire de la chromatine joue un rôle clé dans la réparation des cdb et pourrait déterminer le « choix » du mode de réparation. Dans le cas des mécanismes de recombinaison V(D)J et de la commutation isotypique c’est la voie NHEJ qui est systématiquement adoptée.
2. Facteurs de remodelage
Les facteurs de remodelage agissent de concert avec les HATs pour ouvrir la chromatine et permettre l’accès des protéines de réparation à l’ADN (Osley et al. 2007) (Figure 6). Au niveau des cassures, les facteurs de remodelage les plus étudiés sont INO80, SWR1 et SWI/SNF et RSC. Le complexe RSC appartenant à la famille SWI/SNF interagit avec le complexe MRN chez la levure et serait donc impliqué dans la relaxation de la chromatine dès le début de la réparation (Shim et al. 2005). Le complexe SWI/SNF est également impliqué au début du processus de réparation et permettrait de phosphoryler efficacement H2AX (Park et al. 2006). INO80 modifie la structure de la chromatine grâce à son activité hélicase. Ce complexe est recruté au niveau des cdb grâce à son interaction avec γH2AX (Downs et al. 2004; van Attikum et al. 2004). INO80 pourrait faciliter l’accès au niveau de la cassure aux protéines impliquées dans la maturation des extrémités, comme MRN et Ku80 (Morrison et al. 2004; van Attikum et al. 2007). Pour permettre l’accès aux protéines, INO80 retire γH2AX, ce qui pourrait également faciliter le retour à la normale après réparation. Enfin, SWR1 affecte la chromatine via le remplacement d’H2A par H2AZ (Mizuguchi et al. 2004). Le rôle de ce variant reste encore inconnu mais semble important pour le bon déroulement de la réparation. En son absence, les cellules sont plus sensibles aux génotoxiques (Kobor et al. 2004).
26
IV.
Réparation des cassures double brin de l’ADN par NHEJ
Deux processus majeurs sont responsables de la réparation des cdb chez les eucaryotes (Jackson 2002; Friedberg 2003). La réparation par jonction des extrémités non homologues (NHEJ) prédomine au cours de la phase G1 du cycle cellulaire, bien qu’elle ait également lieu au cours des autres phases du cycle. La NHEJ répare les cdb indépendamment des homologies entre les séquences, pouvant occasionner une réparation infidèle des cassures. A l’inverse, la réparation par recombinaison homologue (RH) permet une réparation fidèle des cdb car elle utilise la chromatide sœur comme modèle. Pour cette raison la RH a lieu pendant les phases S et G2 au cours desquelles la chromatide sœur est disponible.
Au cours des processus affectant les loci Ig, la réparation par NHEJ semble majoritairement utilisée (figure 9 et 15), c’est pourquoi je décrirai principalement les évènements concernant ce mode de réparation.
A.
Détection et signalisation des cassures
La réponse cellulaire face aux cdb est initiée par les protéines ATM et NBS1 (Nijmegen Breakage Syndrome) qui agissent en coopération.
ATM fait partie de la famille des PI3 kinases et représente l’une des protéines indispensables au maintien de la stabilité du génome et à l’activation des points de contrôle du cycle cellulaire (Shiloh 2003). Après activation, ATM est capable de phosphoryler et d’activer de nombreuses protéines de la réparation dont NBS1 (Zhou et al. 2006; Matsuoka et al. 2007).
De même qu’ATM, la protéine NBS1 est fortement impliquée dans la stabilité du génome et l’activation des points de contrôle du cycle (Iijima et al. 2004). NBS1 forme un complexe avec les protéines MRE11 et RAD50, le complexe MRN, recruté au sein des cdb dès leur intervention. Par la suite il est maintenu en place via l’interaction de NBS1 avec γH2AX (Kobayashi et al. 2002). Ce complexe est nécessaire au recrutement d’ATM et à la phosphorylation efficace de ses substrats (Lee and Paull 2005; Lee and Paull 2007). MRE11 possède une activité 3’-5’exonucléase (Paull and Gellert 1998), cette activité est particulièrement accrue lorsque MRE11 participe au complexe MRN (Trujillo et al. 1998; D'Amours and Jackson 2002). De plus MRE11 possède une activité endonucléase à la fois simple brin et double brin (Paull and Gellert 1999).
27 RAD50 est un membre de la famille des protéines SMC (Structural Maintenance of Chromosomes), impliquées dans la condensation des chromosomes et la cohésion entre les chromatides sœurs. RAD50 possède un motif en crochet (zinc-hook) qui lui permet de se dimériser et présente une activité ATPase indispensable à cette dimérisation (Paull and Gellert 1999). La délétion de l’une des protéines du complexe MRN conduit à une létalité embryonnaire (Xiao and Weaver 1997; Luo et al. 1999; Zhu et al. 2001).
Après apparition de la cdb, le complexe MRN se lie aux extrémités et recrute la protéine ATM, qui elle-même active le complexe (Lee and Paull 2005). MRN initie les premiers remodelages de la chromatine pour permettre l’accès à l’ADN. MRN maintiendrait également les deux extrémités d’ADN en juxtaposition pour permettre la réparation. En effet, la dimérisation de RAD50 entraîne une interaction entre deux complexes MRN (Hopfner et al. 2000; de Jager et al. 2001). Par la suite ATM activée phosphoryle de nombreuses protéines impliquées dans différents processus comme le remodelage de la chromatine ou l’arrêt du cycle cellulaire : H2AX (Burma et al. 2001), MDC1 (Goldberg et al. 2003), CHK2 (Matsuoka et al. 2000), p53 (Banin et al. 1998), 53BP1 (Anderson et al. 2001), BRCA1 (Cortez et al. 1999). Dans le cas de la réparation par NHEJ, elle phosphoryle Artémis (Chen et al. 2005), DNA-PKcs (Chen et al. 2007), APLF (Macrae et al. 2008) et XLF (Cernunnos) (Yu et al. 2008). Ainsi, le rôle d’ATM est crucial dans l’orchestration de la réparation en permettant la mise en place d’un environnement idéal.
Chez les mammifères, la voie NHEJ classique requiert différentes protéines : l’hétérodimère Ku70/80, DNA-PKcs, XRCC4, DNA ligase IV, Artémis et XLF. D’autres protéines semblent également participer à ce mécanisme dans une voie NHEJ alternative. Plusieurs expériences menées in vitro, sur différents extraits cellulaires, et in vivo ont permis mettre en évidence que la réparation des cdb pouvait avoir lieu indépendamment de la voie impliquant la DNA-PKcs (North et al. 1990; Johnson and Fairman 1997; Cheong et al. 1999; DiBiase et al. 2000; Wang et al. 2003; Perrault et al. 2004). Au cours de ces voies alternatives, les ADN polymérases µ et λ, PNK (polynucléotide kinase) et l’hélicase WRN pourraient être impliquées dans la maturation des extrémités avant l’étape de ligation. La protéine PARP1 (Poly
ADP-28 ribosepolymérase 1), normalement impliquée dans la réparation par BER, a été proposée pour intervenir dans la voie alternative avec XRCC1 et la ligase III au cours de l’étape de jonction.
La réparation par NHEJ semble se dérouler en trois phases. Tout d’abord une phase de reconnaissance, de maintien et de protection des extrémités, suivie de la maturation de ces extrémités pour permettre enfin leur jonction.
B.
Maintien et protection des extrémités
Au cours de la NHEJ, Ku est la première protéine qui se lie à l’ADN (Walker et al. 2001). Comme évoqué précédemment, Ku est composée de deux sous-unités, Ku70 et Ku80. Dans sa partie N-terminale, Ku70 contient un domaine phosphorylé par DNA-PKcs in vitro (Chan et al. 1999) tandis que sa partie C-terminale présenterait un domaine de liaison à l’ADN (Lees-Miller and Meek 2003). Ku80 possède un long bras dans sa partie C-terminale, probablement impliqué dans les interactions protéiques (Zhang et al. 2004). En effet, cette région est nécessaire à l’interaction avec la DNA-PKcs (Singleton et al. 1999). Une fois liée aux extrémités des cdb, Ku se déplace sur l’ADN, permettant ainsi l’accès à d’autres protéines, en premier lieu à la DNA-PKcs (Yoo and Dynan 1999). Par ailleurs, la liaison de Ku se traduit par une modification de la conformation des régions C-terminales de Ku70 et Ku80 qui faciliterait leur interaction avec les autres protéines impliquées dans le processus (Lehman et al. 2008) telles que XRCC4-Ligase IV (Nick McElhinny et al. 2000; Mari et al. 2006; Costantini et al. 2007), XLF (Yano et al. 2008), ADNpol µ et λ (Mahajan et al. 2002; Ma et al. 2004). Ainsi Ku représente le point d’ancrage des protéines de la NHEJ et sa liaison à l’ADN constituerait donc un pré-requis au bon déroulement du processus.
La DNA-PKcs est donc recrutée au niveau des cdb par son interaction avec Ku (Gottlieb and Jackson 1993). Elle appartient à la famille des PI3 kinase, au même titre qu’ATM, et contiendrait un domaine de liaison à l’ADN dans sa partie N-terminale (Gupta and Meek 2005). L’interaction de la DNA-PKcs avec Ku permet, dans un premier temps, de maintenir les deux extrémités de la cassure en juxtaposition. Cette interaction semble également protéger les extrémités contre l’action de nucléases et stimule l’activité kinase de la DNA-PKcs (Gottlieb and Jackson 1993; DeFazio et al. 2002). Ainsi la DNA-PKcs devient capable de phosphoryler les différents acteurs de la NHEJ. Ses substrats sont Ku70/80, XRCC4, XLF, Artémis, WRN et la
29 Ligase IV, qui sont phosphorylés in vitro et/ou in vivo (Karmakar et al. 2002; Yu et al. 2003b; Wang et al. 2004; Douglas et al. 2005; Goodarzi et al. 2006; Yu et al. 2008). La DNA-PKcs est également capable de s’autophosphoryler et régulerait ainsi sa liaison à Ku et son activité au sein des cdb (Chan and Lees-Miller 1996; Merkle et al. 2002).
1. Maturation des extrémités
Après la détection des cassures, les extrémités seraient soumises à des remaniements permettant de retirer les séquences ne pouvant être liguées ou les éventuelles autres lésions. Cette étape est en partie régulée par la DNA-PKcs (Ding et al. 2003; Budman and Chu 2005). La maturation des extrémités peut varier selon la nature de la cassure (Mahaney et al. 2009). Les protéines susceptibles d’être impliquées sont Artémis, les ADNpol µ et λ, PNK, APLF et WNR.
Artémis possède une activité 5’-3’ exonucléase et, en présence de la DNA-PKcs et d’ATP, une activité endonucléase simple et double-brin (Ma et al. 2002b; Ma et al. 2005). Elle est recrutée au sein des cdb via son interaction avec la DNA-PKcs (Ma et al. 2002b). Toutefois la délétion d’Artémis n’entraîne pas de défaut majeur de réparation. Ainsi, elle serait impliquée dans la réparation d’une certaine catégorie de cassures in vivo (Wang et al. 2005).
La maturation des extrémités peut conduire à l’apparition de brèches, qui requièrent l’action des ADN polymérases pour permettre la réparation. Les ADNpol µ et λ ainsi que la TdT sont impliquées dans la réparation par NHEJ (Nick McElhinny and Ramsden 2004), bien que la TdT ne soit exprimée que dans les lymphocytes (Gilfillan et al. 1995). Toutes les trois sont recrutées au niveau de la cdb via leur interaction avec Ku.
PNK possède les activités 3’-phosphatase et 5’-kinase. Au cours de la maturation des extrémités, ses activités sont particulièrement importantes pour permettre la jonction (Bernstein et al. 2008). PNK serait recrutée au sein de la cassure via sont interaction avec XRCC4 (Koch et al. 2004).
Tout comme Artémis, la protéine APLF possède les deux activités exonucléase et endonucléase. Elle constitue l’un des derniers acteurs potentiels identifiés dans la réparation par NHEJ. De la même façon que PNK, elle interagit avec XRCC4, mais également avec Ku (Kanno et al. 2007).
30 Enfin la protéine WRN serait impliquée dans la maturation grâce à ses activités 3’-5’ hélicase et 3’-5’ exonucléase (Brosh and Bohr 2007). WRN interagit avec Ku et cette interaction stimule son activité exonucléase (Cooper et al. 2000; Li and Comai 2001).
Ainsi la maturation des extrémités des cdb, au cours de la NHEJ, se fait au sein d’un complexe multiprotéique. La composition de ce complexe peut varier en fonction de la nature de la cassure (Lieber et al. 2008).
2. Jonctions des extrémités
Une fois que les extrémités ont subi la maturation nécessaire à leur ligation, leur jonction est assurée par la ligase IV en complexe avec XRCC4, avec la participation de la protéine XLF.
XRCC4 ne possède pas d’activité enzymatique connue mais agirait davantage comme protéine « plateforme », en facilitant le recrutement des autres protéines impliquées dans la NHEJ. Au cours de la ligation, XRCC4 stabilise la Ligase IV et stimule son activité (Grawunder et al. 1997; Gu et al. 2007a).
XLF possède une structure semblable à celle de XRCC4 (Andres et al. 2007; Li et al. 2008b). De la même façon, elle semble capable de stimuler l’activité de la Ligase IV au cours de la ligation d’extrémités non compatibles. Ceci suggère que XLF ne régulerait l’activité du complexe XRCC4-Ligase IV que dans certaines conditions (Gu et al. 2007b; Lu et al. 2007; Tsai et al. 2007).
Au cours de la voie alternative PARP1 se fixe aux extrémités des cdb et serait suffisante pour les maintenir à proximité l’une de l’autre. La jonction serait ensuite assurée par le complexe XRCC1-Ligase III (Audebert et al. 2004).
Comme évoqué précédemment, chez l’homme et la souris, trois évènements moléculaires sont à l’origine de la production d’une grande diversité d’
V(D)J, l’hypermutation somatique et la commutation isotypique.
V.
Mécanismes de diversification des immunoglobulines
A.
Recombinaison V(D)J
Ce mécanisme entraîne l’assemblage des régions variables au cours du développement des lymphocytes B dans la moelle osseuse, pe
d’anticorps (figure 7).
Figure 7: Représentation de la Recombinaison V(D)J
Représentation simplifiée de la recombinaison ayant lieu au niveau du locus IgH et aboutissant à la production premiers anticorps (IgM) (adapté de Chaudhuri et Alt, 2004).
1. Mécanisme
La recombinaison entre les segments V(D)J se fait au niveau de sites spécifiques and Alt 2004). Les gènes Rag
cours des stades de développement précoces des cellules B, essentielles du complexe multimérique RAG. Cette
mécanisme de recombinaison
cassures double-brin de façon ciblée au niveau des séquences signal de recombinaison ( Ces séquences sont composées d’un heptamère (CACAGTG) et d’un nonamère (ACAAAAAC tous deux extrêmement conservés et séparés de 12 ou 23 paires de bases (pb)
Comme évoqué précédemment, chez l’homme et la souris, trois évènements moléculaires sont à l’origine de la production d’une grande diversité d’
l’hypermutation somatique et la commutation isotypique.
de diversification des immunoglobulines
Recombinaison V(D)J
Ce mécanisme entraîne l’assemblage des régions variables au cours du développement des lymphocytes B dans la moelle osseuse, permettant de constituer un répertoire initial
: Représentation de la Recombinaison V(D)J
Représentation simplifiée de la recombinaison ayant lieu au niveau du locus IgH et aboutissant à la production de Chaudhuri et Alt, 2004).
Mécanisme
entre les segments V(D)J se fait au niveau de sites spécifiques . Les gènes Rag-1 et Rag-2 (Recombination-activating genes 1 et 2)
cours des stades de développement précoces des cellules B, codent les deux p es du complexe multimérique RAG. Cette endonucléase
mécanisme de recombinaison (Mombaerts et al. 1992; Shinkai et al. 1992)
brin de façon ciblée au niveau des séquences signal de recombinaison ( Ces séquences sont composées d’un heptamère (CACAGTG) et d’un nonamère (ACAAAAAC
eux extrêmement conservés et séparés de 12 ou 23 paires de bases (pb)
31 Comme évoqué précédemment, chez l’homme et la souris, trois évènements moléculaires sont à l’origine de la production d’une grande diversité d’Ig : la recombinaison
de diversification des immunoglobulines
Ce mécanisme entraîne l’assemblage des régions variables au cours du développement rmettant de constituer un répertoire initial
Représentation simplifiée de la recombinaison ayant lieu au niveau du locus IgH et aboutissant à la production des
entre les segments V(D)J se fait au niveau de sites spécifiques (Jung activating genes 1 et 2), exprimés au codent les deux protéines est indispensable au (Mombaerts et al. 1992; Shinkai et al. 1992). RAG introduit des brin de façon ciblée au niveau des séquences signal de recombinaison (RSSs). Ces séquences sont composées d’un heptamère (CACAGTG) et d’un nonamère (ACAAAAACC) eux extrêmement conservés et séparés de 12 ou 23 paires de bases (pb) (Max et al. 1979;
32 Sakano et al. 1979; Tonegawa 1983). Dans le locus IgH, les RSSs sont placées en aval des segments V, de part et d’autre des segments D et en amont des segments J. La recombinaison V(D)J ne se fait qu’entre RSSs possédant un séparateur de 12pb d’un côté et de 23pb de l’autre selon la règle 12/23 (Sawchuk et al. 1997; Steen et al. 1997; Swanson 2002) (Figure 8).
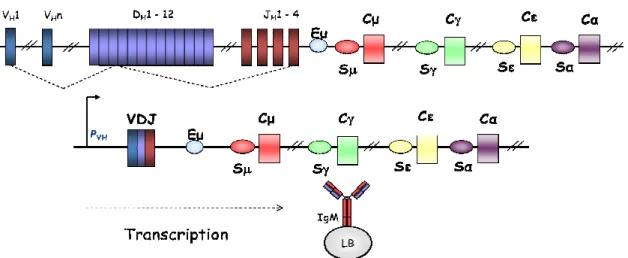
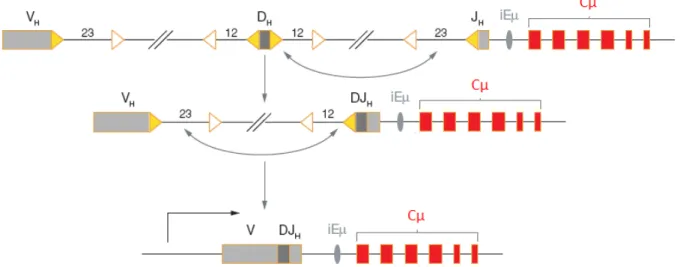
Figure 8: Assemblage et expression du locus IgH
Les segments VH, DH et JH sont schématisés par des rectangles gris, entourés de leurs séquences RSSs respectives les
heptamères sont représenté par des triangles jaunes, les nonamères par des triangles blancs et la taille des séparateurs est indiquée au dessus de chaque RSSs. La séquence réarrangée est ensuite transcrite à partir du promoteur situé en amont de VH et se poursuit sur le gène constant Cµ dont les exons sont représentés en rouge
(adapté de Jung et al, 2006).
Les cassures sont prises en charge par le mécanisme ubiquitaire de NHEJ (Bassing and Alt 2004; Rooney et al. 2004) qui permet alors la juxtaposition et la jonction des segments V(D)J et par conséquent l’expression du locus.
En effet, après action de RAG, quatre extrémités sont générées : deux extrémités dites « codantes » en épingle à cheveux et deux extrémités dites « signal » phosphorylées en 5’. Ceci entraîne l’activation d’ATM qui active à son tour les protéines de la réparation (Perkins et al. 2002; Bredemeyer et al. 2006; Huang et al. 2007a; Bredemeyer et al. 2008a; Bredemeyer et al. 2008b). Le complexe MRN serait activé pour stabiliser les extrémités codantes (Helmink et al. 2009).
Par la suite les extrémités signal sont liguées par XRCC4/Ligase IV tandis que les extrémités codantes nécessitent une maturation avant ligation : Ku70/Ku80 se lient aux extrémités et recrutent la DNA-PKcs en complexe avec Artémis qui clive les extrémités en
épingle à cheveu (Ma et al. 2002b; Bredemeyer et al. 2008b) complexe XRCC4/Ligase IV qui relie alors les extrémités.
La jonction entre les extrémités codantes est imprécise et peut comporter de courtes délétions, l’addition de duplications palindromiques (appelées éléments P) ou de courtes insertions de nucléotides appelés N ajouté
Alt 2004). (Figure 9)
Figure 9: Rôle du NHEJ dans la recombinaison V(D)J
Les extrémités « codantes » et « signal
par XRCC4/Ligase IV alors que les extrémités « DNA-PKcs et Artémis avant ligation (
(Ma et al. 2002b; Bredemeyer et al. 2008b). Cette maturation Ligase IV qui relie alors les extrémités.
La jonction entre les extrémités codantes est imprécise et peut comporter de courtes duplications palindromiques (appelées éléments P) ou de courtes insertions de nucléotides appelés N ajoutés par la TdT avant la ligation des extrémités
: Rôle du NHEJ dans la recombinaison V(D)J
signal » sont fixées par Ku70/80, les extrémités « signal
par XRCC4/Ligase IV alors que les extrémités « codantes » en épingle à cheveu nécessitent une maturation par la et Artémis avant ligation (adapté de Jung et Alt, 2004).
33 . Cette maturation facilite l’accès au
La jonction entre les extrémités codantes est imprécise et peut comporter de courtes duplications palindromiques (appelées éléments P) ou de courtes avant la ligation des extrémités (Jung and
signal » sont simplement liguées » en épingle à cheveu nécessitent une maturation par la
34
2. Régulation de la recombinaison V(D)J
L’expression de RAG ne permet la recombinaison V(D)J que dans les lymphocytes B et T en cours de développement. Cependant l’expression spécifique de RAG ne suffit pas à expliquer l’ordre des réarrangements et leur intervention dépendante du stade de développement. Ainsi une hypothèse, basée sur une accessibilité différentielle du locus IgH, a été proposée pour expliquer ces phénomènes (Yancopoulos and Alt 1985). La transcription, la conformation de la chromatine et l’organisation spatiale des loci Ig au sein du noyau, semblent impliquées dans le contrôle de l’accessibilité des différents segments V(D)J et permettent ou non leurs réarrangements.
a) La transcription
Au cours de la recombinaison, les segments VH DH et JH sont activement transcrits à partir des promoteurs présents en amont des segments VH et DH (Hesslein and Schatz 2001). Le profil de la transcription au cours des différents stades de développement, corrèle avec l’accessibilité des différents segments (Yancopoulos and Alt 1985). Cependant plusieurs études suggèrent que la transcription n’est pas suffisante pour initier la recombinaison (Angelin-Duclos and Calame 1998; Sikes et al. 2002). La transcription ne permettrait donc pas directement le ciblage de la recombinaison, mais induirait les modifications de la chromatine nécessaires pour rendre les séquences transcrites accessibles à RAG (Krangel 2003). Toutefois, il est également envisageable que la transcription ne soit pas la cause, mais la conséquence des modifications de la chromatine. Son rôle précis au cours de la recombinaison reste donc encore à élucider.
Au sein du locus IgH, la transcription est également initiée à partir du promoteur de DQ52, le dernier des segments DH, produisant le transcrit µ0 qui recouvre les segments JH et se termine après Cµ. La transcription du segment DQ52 serait une étape nécessaire à l’accessibilité des segments DH et JH à la recombinaison (Thompson et al. 1995). A la suite des réarrangements DJH, les éléments recombinés sont transcrits (Reth and Alt 1984; Alessandrini and Desiderio 1991) et simultanément, la transcription des segments VH non réarrangés est initiée à partir de leurs promoteurs respectifs. La transcription semble s’éteindre lorsqu’un réarrangement productif a lieu (Yancopoulos and Alt 1985; Corcoran 2005).
35 Par ailleurs, plusieurs transcrits anti-sens ont été identifiés recouvrant les segments VH (Bolland et al. 2004), DH (Bolland et al. 2007; Chakraborty et al. 2007) et JH (Bolland et al. 2007; Perlot et al. 2008). Leur initiation et leur fonction dans la recombinaison V(D)J restent encore à démontrer. Cependant, ces transcrits pourraient également augmenter l’accessibilité du locus à la recombinaison. Toutefois une étude récente suggère que l’observation de transcrits anti-sens pourrait être artéfactuelle et que ces transcrits sont absents au sein des cellules B humaines (Zhao et al. 2009).
Un nombre important de régions cis-régulatrices ont été identifiées tout au long du locus IgH : les enhancers situés entre le dernier segment JH et la région Cµ, et en 3’ du locus, les promoteurs localisés en amont des segments VH et DH. Ces éléments ne sont pas seulement responsables de l’expression du locus IgH mais jouent également un rôle crucial dans le contrôle de l’accessibilité permettant la réalisation des processus de diversification. L’implication de ces séquences régulatrices sera détaillée par la suite.
b) Modifications de la chromatine liées à la recombinaison V(D)J Comme évoqué précédemment, l’état de méthylation de l’ADN joue un rôle dans la régulation de la transcription. Par conséquent, elle pourrait également être impliquée dans la recombinaison V(D)J. En effet, il a été obsevé que la méthylation d’une séquence RSS inhibe le clivage de RAG au niveau de cette RSS (Whitehurst et al. 2000). Par ailleurs, la déméthylation de l’ADN est nécessaire mais insuffisante pour permettre la recombinaison V(D)J (Engler and Storb 1999; Maes et al. 2001)
Plusieurs études ont démontré in vitro qu’une forte condensation de la chromatine inhibe le clivage initial des RSSs par RAG (Kwon et al. 1998; Ng and Bird 2000). La transcription et diverses modifications de la chromatine permettent à la machinerie de recombinaison de cibler les séquences RSSs.
La modification la plus étudiée est l’acétylation des histones H3 et H4 (Nightingale et al. 2007). L’acétylation n’est pas retrouvée de façon homogène sur la région variable mais uniquement au niveau des segments en cours de recombinaison. La région DH-Cµ se trouve acétylée avant l’initiation des premiers réarrangements, les segments VH sont, quant à eux,