T
T
H
H
È
È
S
S
E
E
En vue de l'obtention du
D
D
O
O
C
C
T
T
O
O
R
R
A
A
T
T
D
D
E
E
L
L
’
’
U
U
N
N
I
I
V
V
E
E
R
R
S
S
I
I
T
T
É
É
D
D
E
E
T
T
O
O
U
U
L
L
O
O
U
U
S
S
E
E
Délivré par l'Université Toulouse III - Paul Sabatier Discipline ou spécialité : Biologie Moléculaire et Cellulaire
JURY
Pr Bernard DUCOMMUN Dr Claude SARDET (Rapporteur)
Dr Bernard LOPEZ (Rapporteur) Dr Didier TROUCHE (Directeur de thèse)
MEMBRE INVITE
Dr Martine CHEVILLARD-BRIET (Co-directrice de thèse)
Ecole doctorale : Biologie, Santé, Biotechnologies Unité de recherche : LBCMCP, CNRS UMR5088
Directeur(s) de Thèse : Dr D. TROUCHE et Dr M. CHEVILLARD-BRIET Rapporteurs : Dr C. SARDET et Dr B. LOPEZ
Présentée et soutenue par Sandrine TYTECA Le 27 Juin 2008
THESE
Auteur :
Sandrine TYTECA
Titre :
Laboratoire de Biologie Cellulaire et Moléculaire du Contrôle de la Prolifération CNRS UMR 5088
Bât 4R3B1, 118 route de Narbonne 31062 TOULOUSE cedex 9
ROLE DE L’HISTONE ACETYLTRANSFERASE TIP60
DANS LA REPONSE AUX DOMMAGES A L’ADN
Remerciements
Merci tout d’abord aux Dr. Claude Sardet et Bernard Lopez, ainsi qu’au Pr. Bernard Ducommun d’avoir accepté de participer à mon jury de thèse.
Merci à Didier et Martine pour m’avoir accompagnée tout au long de ces 5 années de labeur. Didier, pour ton excellence scientifique, ta disponibilité, ton enthousiasme sans faille et ta force de conviction dans les moments de doute ! Martine, pour tes conseils précieux, ton amitié et ton soutien en toutes circonstances.
Merci à tous les « Trouche », passés et présents, du fond du cœur. J’ai passé 5 années plus que sympathiques et enrichissantes en votre compagnie. Je recommande chaleureusement cette équipe à tout étudiant qui souhaiterait faire une thèse ! Merci tout spécialement à Cathy, ma confidente des derniers mois, Yvan pour le projet MRN et son humour si… Yvanesque, Marie et Martine pour des manips magnifiques et déterminantes sur le projet Tip60 et p400, Fabrice et Lise pour votre amitié. Merci également à Olivier, Julie, Imen, Lucas, Gaëlle, Estelle, Céline et Laure, je vous souhaite plein de bonheur et de réussite pour l’avenir !
Merci à mes deux labos d’adoption : le LBME et le LBCMCP. Pardon de renoncer à nommer ici tous les gens qui ont croisé ma route ces dernières années, mais la liste serait vraiment trop longue ! Ceci dit, ils se reconnaîtront sans peine. A tous un immense MERCI !!
Merci à mes potes du LBME. Merci tout particulièrement à Elodie qui a partagé tous les moments importants (et moins importants !) de ces années pour finalement devenir une amie chère. Merci aussi à Isa pour ton amitié, plus « récente » mais non moins précieuse !
Merci enfin à ma famille, toujours présente et qui me soutient dans tous mes choix, même les plus surprenants…
Et bien sûr : merci à David qui me supporte au quotidien.
Avant-propos
L’équipe « Chromatine et Prolifération Cellulaire » s’intéresse aux implications des modifications de la chromatine dans des processus tels que la différenciation, la réponse aux dommages à l’ADN, la tumorigénèse... Au cours de tous ces phénomènes, l’état de prolifération de la cellule est altéré. Nous essayons de comprendre comment la modulation de la structure chromatinienne permet la mise en place de programmes génétiques appropriés ou de signalisations lors de ces changements d’état. Dans ce cadre, nous nous intéressons à des enzymes de modifications de la chromatine : histone acétyltransférases, histone déacétylases, histone déméthylases ; notre but étant de déterminer leurs rôles dans tous ces processus.
Pour ma part, j’ai consacré ces 4 années de recherche à l’étude d’une histone acétyltransférase qui fonctionne sous forme d’un complexe multiprotéique : le complexe Tip60. Je me suis intéressée plus particulièrement à ses fonctions dans la réponse aux dommages à l’ADN. En effet, Tip60 intervient à plusieurs niveaux de la prise en charge des dommages : elle participe à la signalisation des dommages, au déclenchement de l’arrêt du cycle et de l’apoptose mais également à la réparation elle-même.
Après une brève introduction sur la chromatine, son rôle de compaction de l’ADN, sa dynamique et son influence sur les processus cellulaires, je décrirai le complexe Tip60 en terme de composition et de fonction dans des cellules en conditions normales de prolifération. Puis, j’entrerai dans le vif du sujet en présentant l’état des connaissances sur les fonctions connues de Tip60 dans un contexte de réponse aux dommages à l’ADN.
Mon introduction n’a pas la prétention d’être exhaustive. Mon but est simplement d’apporter les éléments suffisants à la compréhension de mes travaux de thèse présentés dans la partie « Résultats ». D’autre part, un troisième chapitre est consacré à une discussion plus approfondie de l’ensemble de mes résultats.
Table des Matières
Remerciements………...2
Avant-propos………..3
Table des matières…..………4
INTRODUCTION………...9
A.
LA CHROMATINE ... 11
I. DEFINITION... 11
II. ROLES DE LA CHROMATINE... 11
1. Compaction de l’ADN et organisation au sein du noyau... 11
a. Premier niveau de compaction : la fibre de 11nm... 11
i. Les histones du cœur ... 13
ii. Le nucléosome... 13
b. Deuxième niveau de compaction : la fibre de 30 nm ... 15
i. L’histone de liaison... 15
c. Niveaux supérieurs d’organisation... 15
2. Régulation des processus cellulaires impliquant l’ADN... 17
a. Influence sur la transcription... 17
b. Influence sur la réplication... 17
c. Influence sur la réparation... 18
3. Stabilité du génome... 18
4. Support de l’information épigénétique... 19
III. DYNAMIQUE DE LA CHROMATINE... 19
1. Déplacements physiques de nucléosomes ... 19
2. Echanges de variants d’histones... 21
a. Variants de l’histone H3 ... 21
b. Variants de l’histone H2A... 21
3. Modifications post-traductionnelles des histones ... 25
a. Acétylation... 25
b. Méthylation... 27
c. Phosphorylation ... 27
d. Autres modifications... 28
e. Le code des histones ... 28
B.
LE COMPLEXE TIP60... 31
I. INTRODUCTION... 31
1. Découverte de la protéine Tip60... 31
2. Purification du complexe Tip60 ... 33
II. L’HISTONE ACETYLTRANSFERASE TIP60 ... 33
1. Isoformes... 33
2. Localisation subcellulaire... 34
3. Expression... 35
4. Domaines fonctionnels... 35
5. Activité HAT et substrats... 37
a. Acétylation des histones... 37
b. Acétylation de facteurs de transcription... 39
c. Acétylation d’autres protéines... 39
6. Modifications et régulations connues de Tip60 ... 39
a. Régulation de la stabilité de Tip60... 40
b. Régulation de l’activité de Tip60... 40
c. Autres modifications... 41
III. FONCTIONS DE TIP60 DANS LA PROLIFERATION CELLULAIRE... 41
1. Tip60 est requise pour la prolifération cellulaire ... 41
2. Tip60 et la régulation par E2F/Rb... 41
3. Tip60 et c-Myc ... 42
4. Tip60 et les récepteurs nucléaires... 43
5. Tip60 et la régulation du gène KAI1, cible de NF-κB... 43
6. Tip60 comme co-répresseur... 45
IV. LES PARTENAIRES DE TIP60 AU SEIN DU COMPLEXE... 45
1. L’ATPase p400... 47
2. Les hélicases Tip49a et Tip49b ... 48
3. TRRAP ... 49 4. Autres sous-unités ... 51 a. BAF53 ... 51 b. ING3 ... 51 c. Mrg15 et MrgX... 52 d. Brd8 ... 52 e. DMAP1... 52
C.
TIP60 ET LA PRISE EN CHARGE DES DOMMAGES A L’ADN .... 53
I. DOMMAGES A L’ADN–GENERALITES... 53
1. Les types de dommages et leurs origines ... 53
a. Dommages d’origine endogène... 53
i. Altérations spontanées de bases... 53
b. Dommages d’origine environnementale ... 55
i. Radiations ionisantes ... 55
ii. Rayonnements ultraviolets... 55
iii. Agents chimiques ... 56
2. Les réponses cellulaires ... 57
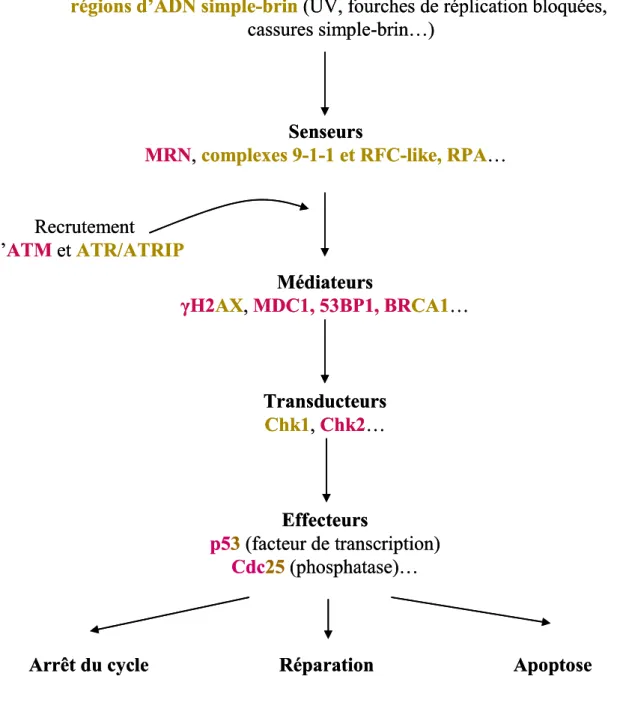
a. Détection des dommages et activation des points de contrôle... 57
i. Les protéines senseurs de dommages... 57
ii. Les kinases ATM et ATR ... 59
iii. Les protéines médiateurs ... 61
iv. Arrêt à la transition G1/S... 61
v. Arrêt à la transition G2/M... 63
b. Réparation... 63
i. Réparation par recombinaison homologue (HR) ... 65
ii. Réparation par ligation des extrémités non homologues (NHEJ) ... 67
c. Apoptose ... 69
d. Le rôle central de p53 ... 71
II. ROLE DE TIP60 DANS LA PRISE EN CHARGE DES DOMMAGES... 73
1. Rôle de Tip60 dans la signalisation ... 73
a. Relation entre Tip60 et p53... 73
b. Rôle de Tip60 dans l’activation d’ATM ... 74
c. Rôle de Tip60 dans la réponse aux dommages induits par les oncogènes... 75
2. Rôle de Tip60 dans l’arrêt du cycle ... 76
3. Rôle de Tip60 dans la réparation... 76
a. Recrutement de Tip60 au niveau des lésions double-brin ... 77
b. Remodelage de la chromatine ... 79
i. Acétylation de l’histone H4 à proximité des lésions... 79
ii. Déplacements de H2AX ... 80
c. Restauration des marques épigénétiques... 80
4. Rôle de Tip60 dans l’apoptose... 82
A.
ROLE DE TIP60 DANS LA VOIE P53 ... 86
I. INTRODUCTION... 86
II. RESUME DES TRAVAUX... 86
III. PUBLICATION :ROLE OF THE HISTONE ACETYL TRANSFERASE TIP60 IN THE P53 PATHWAY (J Biol Chem,2004) ... 87
B.
TIP60 ET P400 SONT REQUISES POUR L’APOPTOSE EN
REPONSE AUX UV MAIS ONT DES ROLES ANTAGONISTES DANS
LA PROGRESSION DU CYCLE CELLULAIRE... 97
I. INTRODUCTION... 97
II. RESUME DES TRAVAUX... 97
III. PUBLICATION :TIP60 AND P400 ARE BOTH REQUIRED FOR UV-INDUCED APOPTOSIS BUT PLAY ANTAGONISTIC ROLES IN CELL CYCLE PROGRESSION (EMBO J,2006) ... 99
C.
UN COMPLEXE TIP60/MRN IMPLIQUE DANS LA REPARATION
DES CASSURES DOUBLE-BRIN PAR RECOMBINAISON
HOMOLOGUE ... 122
I. INTRODUCTION... 122
II. RESUME DES TRAVAUX... 122
III. EN PREPARATION :AMRN/TIP60 COMPLEX INVOLVED IN HOMOLOGY-DIRECTED REPAIR OF DNA DOUBLE-STRAND BREAKS (Soumis,2008) ... 123
A.
TIP60 ET LA SIGNALISATION P53-DEPENDANTE... 152
I. TIP60 ET LA STABILITE DE P53... 152
a. Dégradation par Mdm2 ... 152
b. Dégradation par PIRH2... 153
II. TIP60 ET LA VOIE P53 EN REPONSE AUX DOMMAGES... 153
1. Rôles directs sur p53... 153
a. Interaction Tip60-p53 ... 153
b. Acétylation de p53 ... 154
c. Tip60 : co-activateur de p53 ?... 155
2. Activation d’ATM... 157
B.
TIP60 ET LA REPARATION DES DOMMAGES ... 160
I. IMPLICATION DE TIP60 DANS LES MECANISMES DE REPARATION... 160
II. ROLE DE TIP60 A PROXIMITE DES LESIONS... 161
1. Formation des foyers MRN ... 161
2. Acétylation de l’histone H4... 161
3. Tip60 et H2AX... 162
a. Déplacements de H2AX non phosphorylé ... 162
b. Echange de γH2AX... 163
C.
TIP60 : ONCOGENE OU SUPPRESSEUR DE TUMEUR ?... 165
I. TIP60 EST DEREGULEE DANS DES CANCERS... 165
1. Cas de sous-expression de Tip60 ... 165
2. Cas de surexpression de Tip60 ... 166
II. TIP60 ET LA TUMORIGENESE... 166
1. Tip60 en tant que suppresseur de tumeur ... 167
2. Tip60 en tant qu’oncogène... 168
3. Modèle : 2 seuils critiques de la quantité de Tip60 ... 169
D.
TIP60 ET P400... 171
I. ANTAGONISME FONCTIONNEL ENTRE TIP60 ET P400 ... 171
1. Mise en évidence ... 171
2. Le ratio Tip60/p400 dans les cancers du colon ... 171
II. QUEL MODELE MOLECULAIRE POUR CET ANTAGONISME ?... 172
1. Fonctions indépendantes de Tip60 et p400 ?... 172
2. Antagonisme au sein du complexe Tip60 ? ... 173
3. Compétition entre les complexes Tip60 « classique » et Tip60/MRN ? ... 174
a. Tip60 : une protéine limitante... 174
b. Modèle de travail ... 175
Bibliographie………..183
Annexes………..197
-Figure 1 : Les différents niveaux de compaction de la chromatine
A. L’ADN s’enroule autour d’un octamère d’histones pour former le nucléosome. La
chaîne de nucléosomes constitue le premier niveau de compaction de la chromatine appelé « fibre de 11nm » ou « collier de perles ». Le niveau supérieur est appelé « fibre de 30nm ». La condensation maximale est atteinte avec le chromosome mitotique (modifié de Genetika®). B. Fibres de 11 et 30 nm observées en microscopie électronique (tiré de Molecular Biology of the Cell, 4th edition (2002)).
ADN Nucléosome Fibre de 11nm Fibre de 30nm
B.
A.
11nm 30nm Chromosome mitotiqueA. L
A
C
HROMATINE
I. D
EFINITIONDans la cellule eucaryote, l’ADN n’est pas nu mais associé à de nombreuses protéines selon des structures hautement organisées. C’est ce complexe nucléoprotéique qui occupe le noyau de nos cellules que l’on appelle « chromatine ». Elle fut baptisée ainsi par Walther Flemming en 1882 à cause de son affinité pour des colorants. On sait aujourd’hui que cette structure n’a pas qu’un rôle physique de compaction de l’ADN mais qu’elle a en plus de multiples rôles, tous essentiels. Grâce à une nature très dynamique, elle permet tout d’abord des régulations extrèmement fines de l’expression des gènes ainsi que des autres processus biologiques faisant intervenir l’ADN, notamment par les modifications qui peuvent s’opérer sur les protéines majeures qui constituent la chromatine : les histones. Elle intervient également dans le maintien de l’intégrité du génome et transmet même des informations au-delà de la séquence des gènes : une information épigénétique.
II. R
OLES DE LA CHROMATINE1. Compaction de l’ADN et organisation au sein du noyau
La compaction en chromatine permet en tout premier lieu le confinement des chromosomes dans le volume restreint du noyau. Il existe différents degrés de compaction (Figure 1A).
a. Premier niveau de compaction : la fibre de 11nm
Le premier niveau de compaction, appelé également « fibre de 11nm » ou « collier de perles » correspond à l’enroulement de la molécule d’ADN autour de protéines particulières appelées histones. L’unité de base de cet enroulement, le nucléosome, est répété régulièrement tout au long de la molécule d’ADN. C’est cette succession linéaire de particules, visible en microscopie électronique, qui a conduit ses premiers observateurs à comparer la chromatine à un « collier de perles » (Figure 1B). Un nucléosome correspond à 146 paires de bases d’ADN enroulées autour d’un octamère d’histones.
A.
B.
Figure 2 : Le nucléosome
A. Structure cristallographique du nucléosome à 2,8 Å de résolution (Luger et al., 1997). Bleu = H3, vert = H4, jaune = H2A, rouge = H2B. B. Assemblage de l’octamère d’histones, coeur du nucléosome (tiré du site www.answers.com).
i. Les histones du cœur
Les histones du cœur sont de petites protéines globulaires riches en acides aminés basiques et hautement conservées au cours de l’évolution. On en dénombre quatre : H3, H4, H2A et H2B. Elles constituent le cœur protéique du nucléosome, chaque histone étant présente en 2 exemplaires pour former l’octamère (Khorasanizadeh, 2004). Toutes ces protéines sont essentielles pour les organismes eucaryotes et sont codées par plusieurs gènes regroupés en clusters sur les chromosomes 1 et 6. Leur expressionestmassive en phase S afin de permettre leur incorporation à l’ADN néosynthétisé.
Les quatre histones ont des caractéristiques structurales communes, à savoir une queue N-terminale non structurée de 20 à 30 acides aminés et un domaine globulaire central appelé domaine « histone-fold ». Celui-ci est constitué de trois hélices alpha et permet l’interaction dite en « poignée de main » entre histones ainsi que l’interaction avec l’ADN (Khorasanizadeh, 2004).
ii. Le nucléosome
Le nucléosome est l’unité minimale de la chromatine. Sa structure précise n’est connue que depuis 1997 grâce aux travaux de cristallographie aux rayons X de Luger (Figure 2A) (Luger et al., 1997). Il est constitué d’un octamère d’histones autour duquel s’enroulent 146 paires de base d’ADN. On distingue ainsi l’ADN nucléosomal (autour des histones) et l’ADN internucléosomal qui relie deux nucléosomes (40 à 70 paires de base). L’assemblage d’un nucléosome, au cours de la phase S, se fait en plusieurs étapes (Figure 2B) : il y a tout d’abord formation d’un tétramère central (H3-H4)2 sur lequel se fixent de part et d’autre deux
dimères (H2A-H2B) (Khorasanizadeh, 2004). L’ADN vient ensuite entourer l’octamère en liant les domaines histone-fold, permettant une compaction de l’ADN d’un facteur supérieur à 5. Les interactions entre histones et ADN sont des interactions faibles et par conséquent labiles. D’autre part, elles ne sont pas spécifiques de la séquence d’ADN puisqu’il n’y a aucun contact avec la base. Enfin, une particularité structurale importante du nucléosome est le fait que les queues N-terminales des histones pointent vers l’extérieur de la particule. L’exposition de ces acides aminés à la surface et donc leur accessibilité à des enzymes de modifications est largement utilisée par la cellule à des fins de régulation. Cela sera développé ultérieurement dans ce manuscrit.
A.
B.
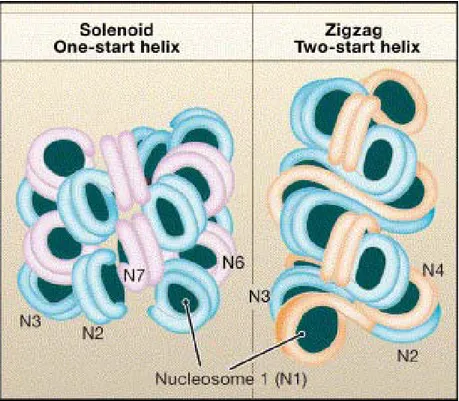
Figure 3 : La fibre de 30nm
A. Il existe à l’heure actuelle deux modèles principaux de repliement pour la fibre de 30nm : le solénoïde et le zigzag (Tremethick et al., 2007). B. Liaison de l’histone H1 sur les nucléosomes et formation de la fibre de 30nm (modifié de http://chemistry.umeche.maine.edu/CHY431/Nucleic4.html.).
Histone H1
b. Deuxième niveau de compaction : la fibre de 30 nm
Afin d’augmenter encore la compaction de l’ADN, la fibre de 11 nm peut à son tour s’enrouler sur elle-même en une super-hélice de 30 nm de diamètre dont chaque tour comprend 6 à 8 nucléosomes (Tremethick, 2007). Il existe plusieurs modèles de repliement (Figure 3A). Dans cette conformation, la séquence primaire de l’ADN est très peu accessible aux différents facteurs protéiques qui doivent s’y lier.
i. L’histone de liaison
La formation de cette super-hélice est possible grâce à l’intervention d’un autre type d’histone : l’histone H1 ou histone de liaison (linker histone). Cette petite protéine est un peu différente des histones du cœur : bien qu’elle possède un domaine globulaire central et une queue N-terminale, elle présente aussi une longue partie C-terminale non structurée. H1 facilite le repliement du collier de perles en interagissant d’une part avec l’ADN internucléosomal et d’autre part avec le cœur du nucléosome (Figure 3B). L’importance de l’histone H1 dans cette structure est clairement établie puisque la déplétion de H1 entraîne un repliement incorrect de la fibre de 30 nm (Thoma et al., 1979). La mise en place de la super-hélice est également facilitée par des interactions entre nucléosomes.
c. Niveaux supérieurs d’organisation
L’état maximal de compaction des chromosomes est atteint lors de la mitose. Au moment de la métaphase, les chromosomes sont arrangés en boucles d’ADN attachées à leur base sur un squelette protéique (Figure 4A). En moyenne, ces boucles font 85 kb, ce qui équivaut à 180 à 300 nucléosomes déjà enroulés en fibre de 30 nm. Atteindre ce niveau de compaction nécessite de l’énergie et notamment l’intervention de deux enzymes qui utilisent l’ATP : la topoisomérase II et le complexe condensin (Swedlow and Hirano, 2003). D’autre part, de nombreuses protéines non histones entrent également dans la composition de la chromatine très condensée.
Notons enfin que la chromatine en interphase est hautement organisée. Non seulement chaque chromosome occupe un espace individuel appelé « territoire chromosomique », mais elle présente également des domaines plus ou moins condensés visibles par des méthodes de coloration de l’ADN (Schneider and Grosschedl, 2007). L’état de compaction de ces domaines est directement lié à leur fonction. En effet, de manière très générale, on distingue
Figure 4 : Niveaux supérieurs d’organisation de l’ADN au sein du noyau
A. Un chromosome mitotique vu en microscopie électronique. Ce chromosome d’insecte à été traité de manière à révéler les boucles d’ADN qui émanent du squelette protéique central (tiré de Molecular Biology of the Cell, 4th edition (2002)). B. Dans un noyau en interphase vu en microscopie électronique, on distingue l’euchromatine (claire, décondensée, active en transcription) et l’hétérochromatine (sombre, condensée, inactive) (tiré de http://academics.hamilton.edu/biology/kbart/image/nucleus.jpg).
B.
A.
euchromatine hétérochromatine boucle d’ADN squelette protéique noyaul’euchromatine, peu condensée et transcriptionnellement active de l’hétérochromatine, condensée et inactive (Figure 4B).
2. Régulation des processus cellulaires impliquant l’ADN
Un des rôles fondamentaux de la chromatine est de réguler l’accès des protéines à la séquence d’ADN par son état plus ou moins compacté. Tous les processus impliquant l’ADN sont soumis de près ou de loin à l’influence de la structure chromatinienne. Voici quelques exemples à l’échelle globale du génome.
a. Influence sur la transcription
La structure de la chromatine permet tout d’abord une régulation fine de l’expression génique. En effet, les zones de chromatine condensée (de manière générale l’hétérochromatine) sont généralement inactives en transcription. Les gènes contenus dans ces régions, déjà peu nombreux, sont donc éteints. Ces différences de compaction permettent la mise en place de patrons d’expression spécifiques et explique comment, au sein d’un individu, des cellules qui possèdent le même patrimoine génétique peuvent être si diverses en morphologies et en fonctions : chaque type cellulaire exprime un ensemble de gènes différents. Cette sélection se fait notamment via une structuration de la chromatine. Au cours du développement par exemple, la différenciation cellulaire nécessite la mise en silence de certains gènes alors que d’autres sont activés. C’est également le cas des programmes de mort cellulaire tels que l’apoptose, de la prolifération, et de toute situation au cours de laquelle la cellule doit répondre à un signal extracellulaire en adaptant son patron d’expression génique.
Chez les mammifères, il existe également un cas particulier de régulation par la structure de la chromatine : la compensation de dose par inactivation du chromosome X. Afin de rééquilibrer l’expression génique entre mâles (1 X) et femelles (2 X), un des deux chromosomes X est mis en silence chez la femelle par formation d’une structure hétérochromatinienne.
b. Influence sur la réplication
Pendant la phase S, la synthèse de l’ADN est initiée en de nombreux points du génome appelés « origines de réplication ». Les origines sont d’une grande diversité de taille, de séquence, d’activité et ne sont pas toutes utilisées lors d’un cycle de réplication. Il a été montré que la structure de la chromatine pouvait influer sur la fréquence d’utilisation d’une origine ou encore la précocité de son déclenchement pendant la phase S. En effet, les origines
de réplication situées dans l’euchromatine se déclenchent plus tôt que celles de l’hétérochromatine (pour revue (McNairn and Gilbert, 2003)). D’autre part, l’acétylation sur les queues N-terminales des histones semble induire une initiation plus précoce des origines de réplication chez la levure Saccharomyces cerevisiae (Zhou et al., 2005).
c. Influence sur la réparation
Il est délicat de déterminer si la structure de la chromatine influence vraiment la réparation de l’ADN. En effet, il faudrait pouvoir connaître la localisation de chaque lésion, qui sont pourtant générées au hasard par les agents génotoxiques. Cependant, des études commencent à émerger qui proposent une influence de la chromatine sur la réparation. Par exemple, la phosphorylation sur la queue N-terminale de l’histone H2AX, qui est une marque indispensable à la signalisation de la lésion, ne semble pas pouvoir se produire dans l’hétérochromatine sauf pendant la réplication (Cowell et al., 2007). Ces résultats suggèrent donc que la chromatine très condensée serait réfractaire à la réparation mais ces données sont encore très préliminaires.
3. Stabilité du génome
C’est principalement le rôle de l’hétérochromatine. En effet, bien qu’elle soit inactive au niveau transcriptionnel, elle possède un rôle essentiel de protection du génome contre la propagation des séquences répétées. En effet, l’insertion d’une de ces séquences dans un gène peut inhiber tout ou partie de son expression et avoir de graves conséquences sur le fonctionnement de la cellule. La compaction de ces transposons (éléments génétiques mobiles) en hétérochromatine empêche la liaison des facteurs protéiques responsables de leur mobilité (Henikoff, 2000).
Le rôle de l’hétérochromatine dans la stabilité du génome passe aussi par la structuration des régions centromériques : des séquences répétées compactées indispensables au fonctionnement des centromères. Cela permet la ségrégation correcte des chromatides sœurs en mitose et une bonne répartition des chromosomes dans les cellules-filles (Henikoff, 2000).
On peut enfin citer le rôle de l’hétérochromatine dans la formation de la structure en boucle des télomères qui protège les extrémités des chromosomes de la dégradation et empêche les translocations. Sans ces structures, les extrémités des chromosomes sont non seulement exposées à des activités exonucléases, mais risquent également d’être reconnues comme des cassures double-brin et « réparées » par jonction de deux chromosomes.
4. Support de l’information épigénétique
Au-delà de la séquence primaire de l’ADN, porteuse de l’information génétique, il existe une information transmissible portée par la chromatine : l’information épigénétique.
Il s’agit notamment de modifications post-traductionelles des queues N-terminales des histones et de méthylation sur les îlots CpG de l’ADN qui sont stables au cours des mitoses successives et même au cours de la méiose, donc des générations.
C’est par exemple le cas du phénomène d’empreinte génomique parentale ou « imprinting » : des marques épigénétiques (méthylations sur l’ADN et les histones) permettent de distinguer les deux allèles d’un même gène (maternel ou paternel). Ceci conduit souvent à la répression de l’allèle méthylé donc à l’expression monoallélique du gène en question. Cette information est transmise à la descendance.
La composition du nucléosome (par exemple en variants d’histones) peut également être transmise à la cellule-fille et peut donc être considérée comme une information épigénétique.
Récemment, il a été proposé l’existence d’un complexe macro-moléculaire impliqué dans la transmission de ces marques épigénétiques au cours de la réplication de l’ADN : le complexe ECREM (Epigenetic Code Replication Machinery), constitué de nombreuses enzymes de modifications de l’ADN et des histones (Bronner et al., 2007).
III.
D
YNAMIQUE DE LA CHROMATINELa chromatine et sa structure jouent donc un rôle primordial dans la régulation de l’accès à l’ADN. Afin que la cellule mette en place la configuration adaptée à chaque situation, il faut que cette structure soit hautement dynamique. Nous verrons ici comment elle peut être modifiée au niveau moléculaire, notamment en jouant sur les histones et le nucléosome. Par ailleurs, les mécanismes décrits ci-dessous sont souvent complémentaires lors d’une réponse cellulaire donnée.
1. Déplacements physiques de nucléosomes
Dans un nucléosome, la liaison entre histones et ADN implique une centaine d’interactions faibles de type liaisons hydrogènes, électrostatiques et hydrophobes. Il existe au sein de la cellule des complexes multiprotéiques capables de détruire une partie de ces interactions et de faire glisser le nucléosome le long de la fibre d’ADN. Ils sont appelés complexes de remodelage de la chromatine ATP-dépendants. Chez l’homme, on en compte 4
ADN
Nucléosome
Complexe de remodelage ATP-dépendant (SWI/SNF)
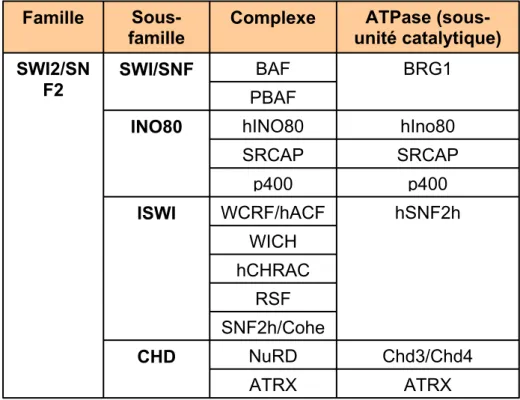
Figure 5 : Remodelage de la chromatine ATP-dépendant
A. Tableau récapitulatif des différents complexes de remodelage ATP-dépendants chez
l’homme. B. Un modèle de translocation de nucléosome. Ici, un complexe SWI/SNF (en vert) interagit avec une large portion d’ADN nucléosomal et détruit les interactions histone/ADN. La grande boucle d’ADN ainsi formée se propage ensuite à la surface du nucléosome. A la fin de la réaction, une nouvelle séquence d’ADN est libre de toute interaction (modifié de Gangaraju and Bartholomew, 2007).
A.
B.
ATRX ATRX Chd3/Chd4 NuRD CHD SNF2h/Cohe RSF hCHRAC WICH hSNF2h WCRF/hACF ISWI p400 p400 SRCAP SRCAP hIno80 hINO80 INO80 PBAF BRG1 BAF SWI/SNF SWI2/SN F2 ATPase (sous-unité catalytique) Complexe Sous-famille Famillefamilles : SWI/SNF, ISWI, CHD et INO80 comprenant un ou plusieurs complexes (Gangaraju and Bartholomew, 2007) (Figure 5A). Ces complexes ont chacun leur spécificité mais on peut cependant dégager un mécanisme commun. Grâce à l’energie libérée par l’hydrolyse de l’ATP, ces complexes parviennent à « détacher » une portion plus ou moins importante d’ADN des histones, créant ainsi une boucle qui se propage le long du nucléosome. A la fin de la réaction, le nucléosome a transloqué d’une distance plus ou moins importante (Figure 5B). Ce remodelage a pour effet de rendre accessibles certaines séquences d’ADN et les complexes cités ci-dessus interviennent dans des processus cellulaires qui nécessitent une modification de la structure chromatienne : transcription, réparation et réplication.
2. Echanges de variants d’histones
Il existe des variants d’histones dont la séquence diffère plus ou moins de celle des histones canoniques (Figure 6). Leur expression n’est pas cantonnée à la phase S mais régulée tout au long du cycle. Certains complexes protéiques sont capables d’échanger une histone canonique au sein du nucléosome contre un de ces variants dans le but de conférer une nouvelle fonction à la chromatine (Jin et al., 2005). Chez l’homme, pour l’instant, aucun variant n’a été identifié pour l’histone H4. Voici l’exemple des principaux variants de H3 et H2A.
a. Variants de l’histone H3
Les deux variants majeurs de H3 sont H3.3 et CenH3 (ou CENP-A). Le variant H3.3 diffère de H3 pour 4 résidus seulement. Il est préférentiellement incorporé au niveau de gènes transcrits avec l’aide de sa chaperone HIRA (Ahmad and Henikoff, 2002; McKittrick et al., 2004; Daury et al., 2006). Le variant CenH3 est quant à lui exclusivement ciblé vers les nucléosomes centromériques grâce à une partie de son domaine histone fold et joue un rôle dans la ségrégation des chromatides sœurs lors de la mitose (Kamakaka and Biggins, 2005).
b. Variants de l’histone H2A
Les variants de H2A sont plus nombreux. Le variant H2AX est connu pour être impliqué dans la signalisation des dommages à l’ADN et la réparation. En effet, il possède une partie C-terminale plus longue que H2A où se trouve notamment un résidu conservé dans l’évolution mais absent de H2A : la sérine 139. Ce site est phosphorylé lors de cassures double-brin de l’ADN à proximité des lésions et participe à la réponse aux dommages
Figure 6 : Les variants des histones H3 et H2A
HFD : Histone Fold Domain. Les séquences conservées sont représentées dans la même couleur pour chaque histone. Variants de H3 : Pour H3.3, les 4 résidus qui diffèrent de H3 sont signalés par des barres jaunes. CENPA présente un N-ter unique et la région en bleu clair est nécessaire au ciblage du variant vers les centromères. Variants de H2A : H2AX est différent par sa partie C-ter qui comporte la sérine 139, phosphorylée après dommage à l’ADN. H2AZ a une partie ter courte contrairement à MacroH2A qui possède une queue C-ter particulièrement longue. H2ABBD est le variant le plus court avec un C-C-ter tronqué et une partie N-ter très courte (modifié de Sarma and Reinberg, 2005)
(Kamakaka and Biggins, 2005). La présence du variant H2AZ dans le nucléosome a quant à elle été corrélée à des effets sur la transcription, soit activateurs, soit répresseurs selon le contexte (Guillemette and Gaudreau, 2006). Le variant MacroH2A est quant à lui spécifique du chromosome X inactif chez les mammifères où il a un rôle répresseur sur la transcription. H2ABbd (Barr body deficient) est au contraire exclu du chromosome X inactif et plutôt associé à une transcription active (Kamakaka and Biggins, 2005).
L’échange de H2A par H2AZ peut être couplé à la transcription. Dans ce cas, c’est la progression de l’ADN polymérase II qui déplace les dimères H2A/H2B, aidée du complexe FACT (Facilitates Chromatin Transcription). Après le passage de l’enzyme, il y a assemblage d’un nouveau nucléosome à partir d’un dimère qui contient H2AZ. Cela nécessite l’intervention de complexes de remodelage ATP-dépendants vus précédemment comme SWI/SNF ou ISWI (Bruno et al., 2003), ainsi que la chaperone NAP-1 qui interagit avec les dimères H2A/H2B. H2AZ peut également être incorporé dans la chromatine indépendamment de la transcription par l’ATPase Swr1 chez la levure (Mizuguchi et al., 2004), ainsi que par ses homologues chez l’homme SRCAP (Ruhl et al., 2006; Wong et al., 2007), et p400 (Gevry et al., 2007). Enfin, chez la drosophile, des données in vitro montrent que le variant H2Aν (équivalent à la fois de H2AZ et H2AX) qui est phosphorylé à proximité des lésions de l’ADN pourrait être échangé contre sa forme non phosphorylée par le complexe dTip60, et plus particulièrement par la sous-unité Domino (p400 chez l’homme).
L’incorporation de variants d’histones est donc un autre moyen de modifier la structure et/ou les propriétés de la chromatine. Certains variants peuvent recruter des facteurs différents et être modifiés sur des résidus spécifiques absents des histones canoniques. Il semble également que MacroH2A joue un rôle dans l’hétérochromatinisation du X inactif grâce à l’encombrement stérique dû à sa partie C-terminale plus longue. A noter que la composition en variants des nucléosomes est encore très peu étudiée. Il semble pour l’instant que les variants d’histones soient plutôt homotypiques : c’est-à-dire que les histones sont les mêmes au sein d’une paire (H2A/H2A, H2AZ/H2AZ, CenH3/CenH3, H3.3/H3.3, …) (Bernstein and Hake, 2006). Aucun cas d’hétérotypie n’a été identifié à ce jour. D’autre part, on ne sait pas si un nucléosome peut contenir des variants de plusieurs histones en même temps. Seulement deux études ont montré l’existence de telles combinaisons. Chez S. cerevisiae, H3.3 peut exister avec H2AX (White et al., 2001) et chez T. brucei, H2AZ co-existe avec H2BV (un variant de H2B chez cet organisme) (Lowell et al., 2005).
Figure 7 : Modifications post-traductionnelles sur les queues N-terminales des histones Représentation schématique des modifications post-traductionnelles sur les queues N-terminales des histones canoniques. Exemples de la phosphorylation, la méthylation et l’acétylation.
P P A R T K Y T A R K S T G G K A P R K Q L A T K A A R K S A P A T G G V K 2 3 4 8 9 10 14 17 18 23 26 27 28 36 Ac Ac Ac Me Me Me MeMe P Ac Me Me Me S G R G K G G K G LG K G G A K R H R K V L R 3 5 1 8 12 16 20 P Me Ac Ac Ac Ac Me S G R F K Q G C K A R A K A 5 1 P Ac P E P A K S A P A P K K G S K K A V T K A 5 12 14 15 20 Ac Ac P Ac Ac
H3
H3
H4
H4
H2A
H2A
H2B
H2B
P Phosphorylation Me Méthylation Ac Acétylation
Figure 7 : Modifications post-traductionnelles sur les queues N-terminales des histones Représentation schématique des modifications post-traductionnelles sur les queues N-terminales des histones canoniques. Exemples de la phosphorylation, la méthylation et l’acétylation.
P P A R T K Y T A R K S T G G K A P R K Q L A T K A A R K S A P A T G G V K 2 3 4 8 9 10 14 17 18 23 26 27 28 36 Ac Ac Ac Me Me Me MeMe P Ac Me Me Me P P A R T K Y T A R K S T G G K A P R K Q L A T K A A R K S A P A T G G V K 2 3 4 8 9 10 14 17 18 23 26 27 28 36 Ac Ac Ac Me Me Me MeMe P Ac Me Me Me S G R G K G G K G LG K G G A K R H R K V L R 3 5 1 8 12 16 20 P Me Ac Ac Ac Ac Me S G R G K G G K G LG K G G A K R H R K V L R 3 5 1 8 12 16 20 P Me Ac Ac Ac Ac Me S G R F K Q G C K A R A K A 5 1 P Ac S G R F K Q G C K A R A K A 5 1 P Ac P E P A K S A P A P K K G S K K A V T K A 5 12 14 15 20 Ac Ac P Ac Ac P E P A K S A P A P K K G S K K A V T K A 5 12 14 15 20 Ac Ac P Ac Ac
H3
H3
H4
H4
H2A
H2A
H2B
H2B
3. Modifications post-traductionnelles des histones
Les queues N-terminales des histones, non structurées et pointant vers l’extérieur du nucléosome, sont le siège de nombreuses modifications post-traductionnelles (Figure 7). Principalement, on décrit deux mécanismes par lesquels ces modifications modulent la dynamique de la chromatine : la destruction de contacts ADN-histones qui « relâchent » la structure de la chromatine et le recrutement de facteurs protéiques sur ces sites modifiés. Nous verrons succintement dans cette partie les principales modifications qui existent et leurs conséquences connues sur les processus cellulaires utilisant l’ADN (Kouzarides, 2007).
a. Acétylation
L’acétylation a lieu sur les résidus lysines (K). C’est une modification réversible. Elle est mise en place par des enzymes appelées Histone Acétyltransférases (HAT). Chez l’homme, il existe plusieurs familles de ces enzymes (Figure 8A) et il existe également des enzymes chargées de la réaction inverse : les Histone Déacétylases (HDAC) réparties en 4 classes en fonction de leur spécificité (Figure 8B). Elles agissent également au sein de complexes multiprotéiques. De nombreux sites sont connus à ce jour pour être acétylés sur les queues N-terminales des histones (Figure 7). Parmi toutes les modifications, c’est probablement la plus susceptible de déstructurer physiquement la chromatine car elle neutralise la charge de la lysine et peut provoquer des changements d’affinité entre ADN et histones. D’autre part, elle est capable d’entraîner le recrutement de facteurs puisque des domaines protéiques tels que le bromodomaine reconnaissent spécifiquement les lysines acétylées.
L’acétylation des histones a été impliquée dans de nombreux processus cellulaires. De manière générale, on associe quasi systématiquement l’acétylation à l’activation de la transcription. Elle est également impliquée dans la réparation : il est montré que l’hyperacétylation de l’histone H4 est nécessaire à la réparation des cassures double-brin (Murr et al., 2006) ; d’autre part, chez Schizosaccharomyces pombe, l’acétylation sur H3K56 (dans le domaine histone fold) persiste en phase G2 lorsque les dommages à l’ADN ne sont pas réparés (Celic et al., 2006; Maas et al., 2006), et Hat1 qui acétyle H4K12 chez la levure est impliquée dans la réparation (Qin and Parthun, 2006). L’histone acétyltransférase HBO1 a également été impliquée dans la réplication de l’ADN : elle interagit avec un complexe ORC (Origin Replication Complex) et plusieurs données montrent qu’elle intervient dans la synthèse de l’ADN et l’assemblage du complexe de pré-réplication (Iizuka et al., 2006).
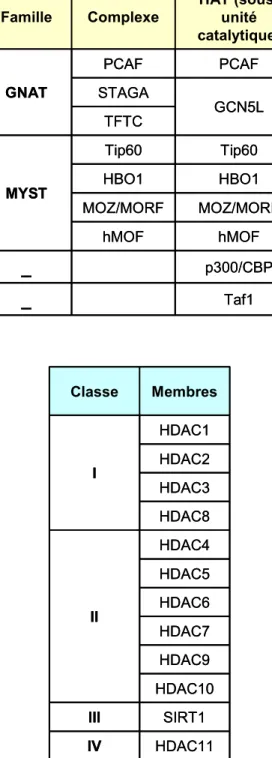
Figure 8 : Familles de complexes HAT et classes de HDAC chez l’homme
A. Tableau récapitulatif des famille de complexes humains contenant une activité HAT. B. Classification des enzymes à activité déacétylase chez l’homme.
A.
HDAC11 IV SIRT1 III HDAC10 HDAC9 HDAC7 HDAC6 HDAC5 HDAC4 II HDAC8 HDAC3 HDAC2 HDAC1 I Membres ClasseB.
Taf1 _ p300/CBP _ hMOF hMOF MOZ/MORF MOZ/MORF HBO1 HBO1 Tip60 Tip60 MYST TFTC GCN5L STAGA PCAF PCAF GNAT HAT (sous-unité catalytique) Complexe FamilleFigure 8 : Familles de complexes HAT et classes de HDAC chez l’homme
A. Tableau récapitulatif des famille de complexes humains contenant une activité HAT. B. Classification des enzymes à activité déacétylase chez l’homme.
A.
HDAC11 IV SIRT1 III HDAC10 HDAC9 HDAC7 HDAC6 HDAC5 HDAC4 II HDAC8 HDAC3 HDAC2 HDAC1 I Membres ClasseB.
Taf1 _ p300/CBP _ hMOF hMOF MOZ/MORF MOZ/MORF HBO1 HBO1 Tip60 Tip60 MYST TFTC GCN5L STAGA PCAF PCAF GNAT HAT (sous-unité catalytique) Complexe Familleb. Méthylation
Les méthylations peuvent avoir lieu sur des arginines (R) ou des lysines (K). Cependant, il exite un niveau supérieur de complexité car plusieurs groupements méthyl peuvent être greffés sur un même résidu. En résumé, il existe des arginines mono- ou di-méthylées (asymétriques ou symétriques) et des lysines mono-, di- ou tri-di-méthylées.
Les enzymes responsables de cette modification, les méthyltransférases, sont très nombreuses et presque toujours spécifiques d’un résidu donné sur une histone donnée. Pendant longtemps, il était admis que la méthylation était une marque irréversible sur les histones, ne pouvant être supprimée que par un remplacement de l’histone modifiée. Cependant, depuis 2004 et la découverte de LSD1 (Lysin Specific Demethylase 1), on connaît l’existence des histones déméthylases (Shi et al., 2004). D’autres enzymes ont été mises en évidence depuis cette date et elles sont toutes spécifiques des lysines. Aucune déméthylase spécifique d’arginine n’est connue à ce jour.
L’effet de la méthylation des histones sur la transcription depend du résidu modifié. Par exemple, la méthylation de H3 sur les lysines 9 et 27 et de H4 sur la lysine 20 répriment la transcription et sont caractéristiques de l’hétérochromatine. La méthylation de H3K9 participe même à sa mise en place en recrutant la protéine HP1. Par contre, la méthylation de H3K4 et H3K36 activent la transcription. Les conséquences des méthylations sur arginines sont moins connues mais il semble également y avoir des effets activateurs ou répresseurs sur la transcription.
Enfin, on peut citer un rôle de la méthylation dans la réparation de l’ADN puisqu’il semble y avoir apparition de méthylation sur H4K20 aux sites de dommages et que cela permette de recruter la protéine Crb2, impliquée dans l’arrêt en G2/M (Sanders et al., 2004).
c. Phosphorylation
Il peut y avoir phosphorylation des histones sur les sérines (S) et les thréonines (T). La phosphorylation de la queue N-terminale de l’histone H3 est globalement associée à l’activation de la transcription (Nowak and Corces, 2000; Dyson et al., 2005). D’autre part, plusieurs phosphorylations sont impliquées dans la réparation de l’ADN. Tout d’abord, la phosphorylation de la sérine 139 de H2AX que nous avons déjà évoquée, mais aussi des phosphorylations sur H4 (sérine 1) et H2A (sérine 129) sont impliquées dans une voie de réparation appélée NHEJ (Non Homologous End Joining). Enfin, des résidus phosphorylés sur H3 (sérine 10 et thréonine 3) sont requis au cours de la mitose pour le déplacement de la protéine HP1 et l’alignement des chromosomes en métaphase.
d. Autres modifications
Il existe de nombreuses autres modifications chimiques sur les histones, pas seulement sur les queues N-terminales, telles que l’ubiquitination, la sumoylation, l’ADP-ribosylation, l’isomérisation des prolines, la déimination des arginines. Ces modifications sont, pour la plupart, moins bien documentées.
En ce qui concerne l’ubiquitination, deux sites jouant un rôle dans l’expression génique ont été mis en évidence : K119 sur H2A et K120 sur H2B, autrement dit, dans la partie C-terminale. Ces modifications ont respectivement des rôles répresseurs et activateurs sur la transcription (Zhu et al., 2005; Wang et al., 2006). Cependant, le mécanisme par lequel l’ubiquitination modifie l’expression génique est mal défini. On pourrait penser que la présence d’une molécule si volumineuse crée un encombrement stérique qui permette à la chromatine de rester ouverte. Elle pourrait également permettre le recrutement de facteurs spécifiques. H3 et H4 sont également ubiquitinylées en réponse aux UV, peut-être pour recruter des protéines de la réparation telles que XPC (Wang et al., 2006).
La sumoylation est également une modification très volumineuse puisque SUMO est très similaire à la molécule d’ubiquitine. Elle survient sur les histones H4, H2A et H2B et entre en compétition avec d’autres modifications qui ont lieu sur les mêmes résidus : acétylation et ubiquitination. Par ce rôle antagoniste, elle permet de réprimer la transcription chez la levure (Nathan et al., 2006).
Le rôle des autres modifications est encore très mal connu et ne sera pas développé ici. Cependant, ce domaine de recherche est en évolution constante et de nouvelles fonctions sont régulièrement découvertes pour ces modifications.
e. Le code des histones
Toutes ces modifications post-traductionelles ne sont pas présentes en même temps sur une même histone. Certaines sont interdépendantes et d’autres mutuellement exclusives. Avant tout, elles sont mises en place en réponse à des stimuli ou bien héritées de la cellule-mère comme marque épigénétique stable. L’idée émise en 2001 par Jenuwein et Allis propose qu’il existe une combinaison de modifications associée à chaque situation : un véritable « code des histones » utilisé par la cellule pour conférer à la chromatine la structure adéquate et/ou une affinité pour des facteurs protéiques, mais aussi pour définir des environnements chromatiniens globaux comme l’hétérochromatine (Jenuwein and Allis, 2001).
Aujourd’hui, il est nécessaire de nuancer la notion de « code des histones » dans la mesure où il est peu probable qu’il existe un code au sens strict du terme qui permette de
prédire précisément la configuration de la chromatine. Ce « code » reflèterait plutôt une tendance à retrouver des marques spécifiques pour une tâche cellulaire donnée, même si plusieurs combinaisons de modifications peuvent amener à la même réponse, et si une combinaison peut servir pour plusieurs processus.
Toutefois, ce modèle est soutenu par l’existence de protéines et de domaines protéiques qui reconnaissent spécifiquement les modifications post-traductionelles sur les histones. La méthylation est reconnue par des domaines « chromo-like » tels que chromo, tudor et MBT, ainsi que par les domaines PHD. L’acétylation est reconnue par les bromodomaines et la phosphorylation, par un domaine de la protéine 14-3-3. Sachant qu’on dénombre des dizaines de protéines contenant ce genre de domaines, cela étend considérablement les possibilités de recrutement à la chromatine. Enfin, la liaison de ces facteurs à la chromatine a très souvent des conséquences sur sa structure car, quand elles ne possèdent pas elles-mêmes une activité de remodelage, elles servent bien souvent à recruter d’autres enzymes qui agissent à leur tour sur la chromatine.
4. Méthylation de l’ADN
La méthylation de l’ADN est également considérée comme une modification de la chromatine jouant un rôle dans sa dynamique. Ici, il ne s’agit plus de modifier les histones ou la structure du nucléosome mais d’ajouter un groupement méthyl directement sur une base de l’ADN, plus précisément sur le carbone 5 du cycle aromatique de la cytosine. Cette modification est mise en place par des enzymes appelées DNA Methyltransférases (DNMT) et elle a lieu spécifiquement sur des cytosines situées dans des régions particulières de l’ADN que l’on nomme « îlots CpG ». Il s’agit de groupes de dinucléotides CG répétés en tandem. A noter que la répartition de ces séquences dans le génome n’est pas uniforme. En effet, la plupart des îlots CpG sont trouvés dans des régions promotrices de gènes.
La méthylation de l’ADN est principalement associée à un état chromatinien répressif (elle participe à la formation de l’hétérochromatine) et à l’inhibition de la transcription (pour revue (Klose and Bird, 2006)). Deux mécanismes sont proposés pour expliquer l’effet sur la transcription. Tout d’abord, la méthylation pourrait empêcher la liaison de facteurs qui reconnaissent la séquence d’ADN (Watt and Molloy, 1988). D’autre part, certaines protéines possèdent un domaine qui leur permet de reconnaître spécifiquement cette modification et de s’y fixer : on les appelle MBPs pour Methyl-CpG Binding Proteins. La plus célèbre d’entre elles est MeCP2. La liaison de ces protéines servirait notamment de plateforme de recrutement pour d’autres enzymes de modifications de la chromatine comme des HDAC
(Jones et al., 1998; Nan et al., 1998) ou des complexes de remodelage de la chromatine (Wade et al., 1999). La coopération de tous ces complexes pourrait expliquer la répression transcriptionnelle sur certains promoteurs.
B. L
E
C
OMPLEXE
T
IP
60
Mes travaux de thèse ont porté sur le rôle d’un complexe à activité histone acétyltransférase, le complexe Tip60. Dans cette partie, je présenterai donc ce complexe en terme de composition, d’activité et de fonctions.
I. I
NTRODUCTION1. Découverte de la protéine Tip60
La protéine Tip60 a été mise en évidence à l’occasion d’un crible double-hybride destiné à rechercher des cofacteurs de la protéine Tat du virus HIV-1 (Kamine et al., 1996). Les auteurs découvrent une nouvelle protéine qui interagit avec Tat in vitro et potentialise son activité transactivatrice lorsqu’on la surexprime. Le cDNA est alors identifié et la protéine est nommée Tip60 pour « Tat interactive protein 60kDa ». Plus tard, une analyse de séquences révèle que deux domaines de Tip60 présentent des homologies avec d’autres protéines (MOF chez la drosophile, MOZ chez l’homme, mais aussi SAS2 et SAS3 chez la levure S. cerevisiae) (Hilfiker et al., 1997). Il s’avère qu’un de ces domaines est un domaine fonctionnel commun à plusieurs histone acétyltransférases permettant la liaison de l’acétyl coenzyme A. Suite à cette observation, les auteurs émettent alors l’hypothèse que Tip60 est peut-être une histone acétyltransférase. L’activité HAT de Tip60 sur un mélange d’histones purifiées est confirmée peu de temps après (Yamamoto and Horikoshi, 1997). Ainsi, Tip60 est une HAT de la famille MYST (pour MOZ, YBF2/SAS3, SAS2, et Tip60). Dès 1998, une histone acétyltransférase de levure présentant des homologies avec la famille MYST, la protéine Esa1, est décrite comme gène essentiel chez S. cerevisiae (Smith et al., 1998). Elle s’avèrera être l’homologue de Tip60 chez la levure. Tip60 est également conservée chez D. melanogaster, D. rerio, M. musculus, A. thaliana et C. elegans (Zhu et al., 2007) avec un taux de similarité allant de 41 à 68% (de D.r à M.m).
Remarque : récemment, le nom HTATIP a été choisi officiellement pour Tip60, nom que l’on retrouve dans la base de données HGNC (HUGO Gene Nomenclature Committee). En 2007, une nouvelle nomenclature pour les enzymes de modification de la chromatine est parue dans le journal Cell (Allis et al., 2007). Tip60 s’appelle à présent KAT5. Par souci de lisibilité et de cohérence avec la bibliographie et mes propres travaux, j’ai conservé l’appellation Tip60 dans mon manuscrit.
Figure 9 : La protéine Tip60
A. Structure du gène tip60 et isoformes connues de la protéine chez l’homme. L’isoforme « canonique », la plus étudiée, est encadrée en rouge. B. Domaines fonctionnels de Tip60. Le domaine MYST, partagé par toutes les HAT de la famille MYST, contient la région responsable de l’activité catalytique (domaine HAT) qui se caractérise notamment par le site de liaison à l’acétylCoA. Tip60 présente également un domaine en doigt de zinc permettant une éventuelle liaison directe à l’ADN, une « NR box », domaine de liaison aux récepteurs nucléaires, ainsi qu’un chromodomaine putatif.
Isoforme 1 : iTip60 (L-Tip60) Isoforme 2 : Tip60 (Tip60α) Isoforme 3 : PLIP (Tip60β, Tip60∆1) 546 aa 513 aa 461 aa
A.
B.
Chromodomaine Doigt de zinc Domaine HAT NR box Domaine MYST Exon 5 Intron 1Figure 9 : La protéine Tip60
A. Structure du gène tip60 et isoformes connues de la protéine chez l’homme. L’isoforme « canonique », la plus étudiée, est encadrée en rouge. B. Domaines fonctionnels de Tip60. Le domaine MYST, partagé par toutes les HAT de la famille MYST, contient la région responsable de l’activité catalytique (domaine HAT) qui se caractérise notamment par le site de liaison à l’acétylCoA. Tip60 présente également un domaine en doigt de zinc permettant une éventuelle liaison directe à l’ADN, une « NR box », domaine de liaison aux récepteurs nucléaires, ainsi qu’un chromodomaine putatif.
Isoforme 1 : iTip60 (L-Tip60) Isoforme 2 : Tip60 (Tip60α) Isoforme 3 : PLIP (Tip60β, Tip60∆1) 546 aa 513 aa 461 aa
A.
B.
Chromodomaine Doigt de zinc Domaine HAT NR box Domaine MYST Chromodomaine Doigt de zinc Domaine HAT NR box Domaine MYST Exon 5 Intron 1 Exon 5 Intron 12. Purification du complexe Tip60
La purification de Tip60 révèle qu’elle fait en réalité partie d’un complexe multiprotéique (Ikura et al., 2000). L’expérience est réalisée à partir de cellules HeLa surexprimant une forme étiquetée HA et FLAG de Tip60. Par deux immunoprécipitations successives utilisant d’abord l’étiquette FLAG, puis HA, les auteurs mettent en évidence au moins 12 protéines co-purifiées avec Tip60. A l’heure actuelle, on en compte environ 18, notamment grâce à l’identification de nouvelles sous-unités en 2003 (Cai et al., 2003). Parmi les protéines identifiées, plusieurs présentent une activité enzymatique. Le « complexe Tip60 » ainsi nommé par les auteurs contient donc au moins les activités suivantes : histone acétyltransférase, ATPase et hélicase. De plus, ils montrent que Tip60 recombinante seule est incapable d’acétyler des histones reconstituées sous forme de nucléosomes. C’est le complexe Tip60 dans son ensemble qui acétyle préférentiellement H4 et H2A au sein de la particule nucléosomale. Ce résultat souligne l’importance du complexe Tip60 en tant qu’entité fonctionnelle. De plus, il est à noter que Tip60 était la première HAT à montrer une spécifité de substrat. Chez la levure, Esa1 fonctionne également sous forme d’un complexe multiprotéique appelé NuA4 et acétyle les mêmes histones. Purifié en 1999 (Allard et al., 1999), le complexe NuA4 est l’orthologue d’une partie du complexe Tip60 chez l’homme (Doyon et al., 2004).
II. L’H
ISTONEA
CETYLTRANSFERASET
IP60
1. Isoformes
Tip60 est codée par un gène situé en position 11q13.1. Le gène s’étend sur environ 7,5kb et se compose de 14 exons. Il existe plusieurs variants d’épissage de Tip60, l’isoforme la plus exprimée étant celle qui correspond à la transcription des 14 exons et que l’on appelle communément Tip60 (Figure 9A).
Un ARNm plus court de 156pb a été mis en évidence par amplification de la séquence de Tip60 à partir d’une banque d’ADNc de cœur humain (Ran and Pereira-Smith, 2000). Les auteurs montrent que cette isoforme est obtenue par exclusion de l’exon 5 au cours de l’épissage et qu’il produit une protéine Tip60 plus courte de 52 acides aminés qu’ils nomment Tip60β. Quelques mois plus tard, deux autres équipes publient également l’existence du même variant d’épissage de Tip60 dépourvu de l’exon 5 (Hlubek et al., 2001; Sheridan et al., 2001). Cependant, les premiers le nomment Tip60∆1 et les autres PLIP pour « cPLA2
phospholipase A2 (cPLA2). Ces mêmes auteurs proposent un rôle de PLIP dans la production
de prostaglandine E2 par cPLA2 et dans l’apoptose. Ce variant existe aussi chez la souris où il
est appelé Tip53 (McAllister et al., 2002).
Au cours du même travail sur des ADNc de cellules Jurkat, Hlubek et al. découvrent une isoforme encore plus courte de Tip60 qu’ils nomment Tip60∆2. Elle correspond à Tip60β/PLIP/Tip60∆1 tronquée de 124 acides aminés dans la partie C-terminale (Hlubek et al., 2001). Jusqu’à présent, aucun autre travail ne fait mention de ce variant.
En 2003, c’est une forme plus longue de Tip60 qui est découverte au laboratoire (Legube and Trouche, 2003). En se basant sur l’étude du gène tip60 de souris qui montre une conservation exceptionnelle de l’intron 1 (99% entre l’homme et la souris, comparé à 20-30% pour les autres introns) (McAllister et al., 2002), les auteurs constatent que dans de nombreux EST et chez plusieurs espèces, l’intron 1 n’est pas épissé. Après analyse des ADNc de cellules Jurkat, HeLa et NIH3T3, ils montrent que cet ARNm plus long est bien exprimé. D’autre part, ils montrent que la transfection d’un plasmide portant la séquence de la forme longue de Tip60 conduit bien à la production d’une protéine Tip60 portant 33 acides aminés supplémentaires.
Une autre isoforme a été récemment découverte chez la souris. Elle est obtenue par rétention d’exons au sein de l’intron 11 (Kim et al., 2006). Cela conduit à la production d’une protéine ayant une partie C-terminale différente des autres Tip60. Les auteurs l’ont baptisée Tip55. Cette isoforme n’a pas été décrite chez l’homme jusqu’à présent.
2. Localisation subcellulaire
Plusieurs études ont montré par immunofluorescence que Tip60 est une protéine nucléaire (Yamamoto and Horikoshi, 1997; Gavaravarapu and Kamine, 2000; Ran and Pereira-Smith, 2000; Cao and Sudhof, 2001). Notons toutefois que ces études ont été réalisées à partir de la surexpression d’une protéine Tip60 étiquetée. Il existe cependant des cas où Tip60 peut être trouvée dans le cytoplasme. Par exemple, elle interagit avec le récepteur membranaire de l’interleukine 9 (IL-9R) lorsqu’il est internalisé dans le cytoplasme (Sliva et al., 1999). D’autres travaux détectent Tip60 dans le cytoplasme de cellules mésangiales rénales de souris (Muckova et al., 2006).D’autre part, peu de données sont disponibles quant à la localisation des autres isoformes, à l’exception des résultats de Ran et Pereira-Smith selon lesquels une forme surexprimée et étiquetée de PLIP/Tip60β est retrouvée autant dans le noyau que dans le cytoplasme (Ran and Pereira-Smith, 2000). Cette différence de localisation
pourrait être une piste pour proposer un rôle spécifique de ce variant d’épissage par rapport à Tip60.
3. Expression
Les taux d’expression de Tip60 et PLIP (dans la plupart des cas il s’agit des ARNm) ont été testés dans quelques tissus humains comme les reins, le foie, la rate, les poumons, le cœur, l’estomac et le cerveau (Ran and Pereira-Smith, 2000; Sheridan et al., 2001). Dans ces études, les deux isoformes ont des profils d’expression comparables et leur expression semble plutôt ubiquitaire, sauf peut-être une expression plus forte dans le cerveau (Ran and Pereira-Smith, 2000). D’autre part, Tip60 est très fortement exprimée dans les testicules chez la souris adulte (McAllister et al., 2002; Thomas et al., 2007).
Enfin, plusieurs études suggèrent un rôle important de Tip60 dans le développement. Dans l’embryon de souris, l’expression de Tip60 est maximale au stade E11 (McAllister et al., 2002). De plus, le knock-out de Tip60 est létal au stade embryonnaire chez les souris homozygotes, ce qui suggère un rôle essentiel de Tip60 dans le développement (Gorrini et al., 2007). Chez la drosophile, l’expression de Tip60 est maximale pendant le stade embryonnaire, puis diminue dans la larve et augmente à nouveau dans l’adulte (Zhu et al., 2007). D’autre part, l’utilisation de siRNA (small interfering RNA) contre Tip60 pendant le développement de l’embryon est létal (Zhu et al., 2007). Chez le poulet, Tip60 est exprimée uniformément au cours du développement à l’exception d’un pic d’expression forte dans le cœur au cours des stades précoces de l’embryogénèse (Lough, 2002).
4. Domaines fonctionnels
La protéine Tip60 comporte plusieurs domaines fonctionnels (Figure 9B). Tout d’abord, elle se caractérise par un domaine catalytique appelé également domaine MYST qui a été déterminé par homologie de séquence avec les autres membres de la famille MYST. Il comprend un domaine en doigt de zinc requis pour des interactions protéines-protéines (Nordentoft and Jorgensen, 2003; Xiao et al., 2003) et le domaine HAT proprement dit, site de liaison de l’acétylCoA et du substrat. Dans la partie N-terminale de Tip60, un chromodomaine a été identifié dont la fonction n’est pas clairement élucidée. Cependant, cela laisse ouverte la possibilité que Tip60 puisse lier des lysines méthylées sur les histones ou sur d’autres protéines. Enfin, une région dans la partie C-terminale appelée « NR box » permet la liaison de Tip60 à des récepteurs nucléaires de type I (Gaughan et al., 2001).
Tip60 H4 K5 K8K12K16 H2A K5 AR UBF c-Myc p53 Rb ATM
Histones Facteurs de transcription Autres protéines
Figure 10 : Les protéines acétylées par Tip60
Tip60 acétyle de nombreuses protéines, classées ici en trois catégories. Tout d’abord les histones H4, H2A et H2AX (les lysines acétylées sont indiquées), mais aussi des facteurs de transcription de plus en plus nombreux ainsi que d’autres protéines cruciales pour la vie cellulaire.
PLAGL2 Ac Ac Ac H2AX K5 Notch1-IC Tip60 H4 K5 K8K12K16 H4 K5 K8K12K16 H2A K5 H2A K5 AR UBF c-Myc p53 Rb ATM
Histones Facteurs de transcription Autres protéines
Figure 10 : Les protéines acétylées par Tip60
Tip60 acétyle de nombreuses protéines, classées ici en trois catégories. Tout d’abord les histones H4, H2A et H2AX (les lysines acétylées sont indiquées), mais aussi des facteurs de transcription de plus en plus nombreux ainsi que d’autres protéines cruciales pour la vie cellulaire.
PLAGL2 Ac Ac Ac H2AX K5 H2AX K5 Notch1-IC
A noter que toutes les isoformes de Tip60 (à l’exception de Tip60∆2) contiennent ces domaines. Il n’est donc pas possible d’imaginer une fonction spécifique à l’un des variants d’épissage sur la simple analyse de sa séquence. En effet, la région absente de PLIP (exon 5) n’a pas, pour l’instant, de fonction déterminée. Il a cependant été proposé qu’elle permette le recrutement de protéines par son caractère riche en proline. A noter également que Tip60 et PLIP semblent former le même complexe dans la cellule (Doyon et al., 2004). La région correspondant à l’intron 1, présente dans la forme longue de Tip60, ne possède pas non plus de fonction connue. En revanche, concernant Tip60∆2, la délétion en C-terminal entraîne la perte du domaine HAT, ce qui devrait lui conférer une fonction très différente de Tip60. Cependant, Hlubek et al. n’ont pas mis en évidence de différence fonctionnelle entre les deux isoformes dans le cas de la régulation de la transcription par Tat, ce qui suggère que, dans leurs conditions expérimentales, cette fonction est indépendante de l’activité HAT de Tip60.
5. Activité HAT et substrats
a. Acétylation des histones
L’activité histone acétyltransférase de Tip60 a été démontrée peu après sa découverte. Les travaux de Horikoshi montrent que la protéine Tip60 recombinante acétyle les histones purifiées H3, H4 et H2A in vitro (Yamamoto and Horikoshi, 1997). Il poursuit ensuite son étude et détermine les sites acétylés sur chacune des histones. Pour cela, il utilise des peptides correspondant aux queues N-terminales : les lysines modifiées sont H2A (K5), H3 (K14) et H4 (K5, K8, K12, K16) (Kimura and Horikoshi, 1998). Toutefois, un résultat important vient nuancer ses conclusions. Des expériences menées sur Tip60 recombinante et des histones assemblées in vitro en nucléosomes montrent que la protéine seule est en réalité incapable de les acétyler au sein de cette structure, ce qui reflète probablement mieux la situation physiologique (Ikura et al., 2000). La présence du reste du complexe Tip60 est indispensable à son activité. De plus, les auteurs s’aperçoivent que dans leurs conditions, H3 n’est pas acétylée par le complexe Tip60. La spécificité de substrat du complexe Tip60 se réduit donc aux histones H2A (K5) et H4 (K5, K8, K12, K16) (Figure 10). Chez la drosophile, Tip60 acétyle également H2Aν sur la lysine 5 (homologue de H2AZ/H2AX) lorsqu’elle est phosphorylée au voisinage de cassures double-brin (Kusch et al., 2004). Enfin, très récemment, il a été montré que Tip60 acétyle le variant d’histone H2AX sur la même lysine en réponse aux cassures double-brins, indépendamment de sa phosphorylation (Ikura et al., 2007).