Le point sur la maladie du hêtre en Wallonie
Texte intégral
Figure




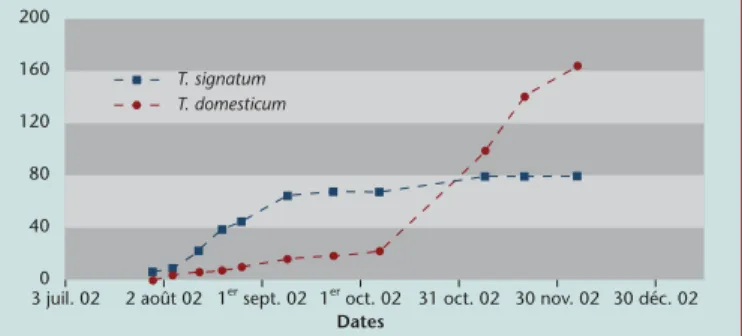

Documents relatifs
Au 4 ème tour après les déclarations de Sébastien, Damien et Victor, on constate que sur les 18 triplets possibles, Damien sait répondre avec cette fois-ci une seule valeur
Sur le bureau, dans le dossier «photos allemand», choisis la photo de départ et glisse-la sur le cliché4. Sandra van
Evaluation - Vérification des acquis de la séquence.. Projet: 03 – Rédiger un manuel technique expliquant le procédé de fabrication de différents produits afin de le
kch individual teat viII be cleilcibed below under the"MadIa. At other locations of the sereen, .Indo.s centaining Infor.ation .ay ~!lppear. One ts 'to report
the given instruction.· If the masked half -word matches this instructi0r exac'tly, ita location and entire contents are typed out. This mask indicates that the
SPOROZOAIRES D'ARTHROPODES D’IRAN 27 Nous avions déjà signalé un cas tératologique analogue chez une autre Grégarine : Didymophyes guttiformis Cordua (Théodoridès
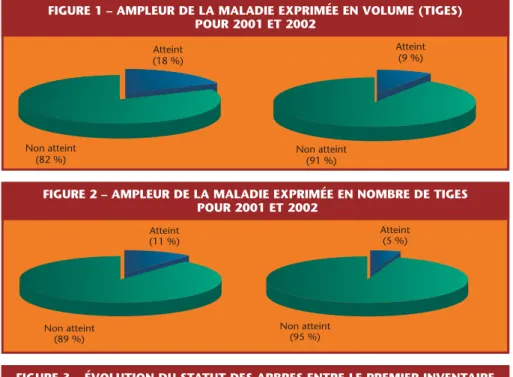
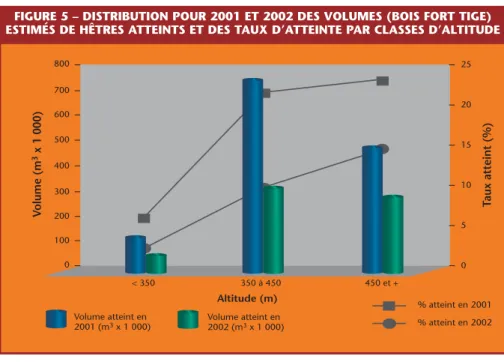
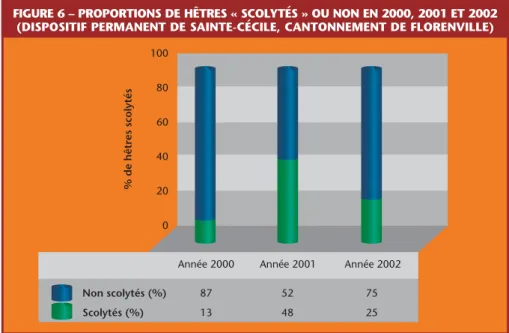
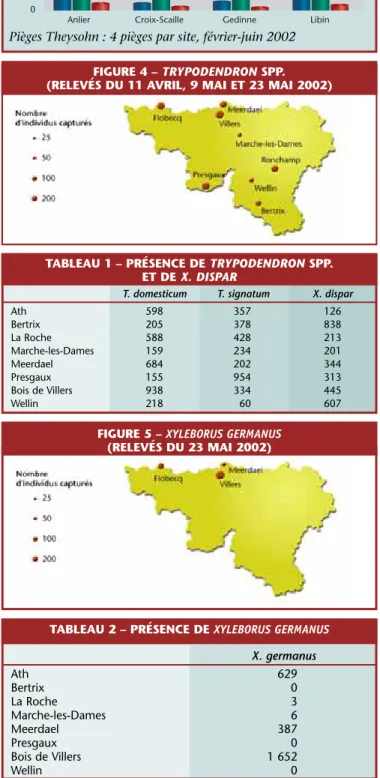
Le dépérissement du Hêtre est attribué à l’action conjuguée d’une cochenille (Cryptococcus fagi) et d’un champignon (Nectria coccinea)!. On se reportera à
— Les notations restant les mêmes qu'au théorème VIII, si une même valeur de uy 1 '^ 1 ^ est la partie principale de plus d'une racine de f(x,y)==0, et de même pour u'y 1 ^, ...,
