ROLE DE LA MAPKKK DLK DANS L'ADIPOGENESE
par
Jean-Philippe Couture
these presentee au Departement de Biologie en vue de I'obtention du grade de docteur es sciences (Ph.D.)
FACULTE DES SCIENCES UNIVERSITE DE SHERBROOKE
Ill
Library and Archives Canada Published Heritage Branch 395 Wellington Street Ottawa ON K1A 0N4 Canada Bibliotheque et Archives Canada Direction du Patrimoine de I'edition 395, rue Wellington Ottawa ON K1A 0N4 CanadaYour file Votre reference ISBN: 978-0-494-83307-0 Our file Notre reference ISBN: 978-0-494-83307-0
NOTICE:
The author has granted a
non-exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
telecommunication or on the Internet, loan, distrbute and sell theses
worldwide, for commercial or non-commercial purposes, in microform, paper, electronic and/or any other formats.
AVIS:
L'auteur a accorde une licence non exclusive permettant a la Bibliotheque et Archives Canada de reproduire, publier, archiver, sauvegarder, conserver, transmettre au public par telecommunication ou par I'lnternet, preter, distribuer et vendre des theses partout dans le monde, a des fins commerciales ou autres, sur support microforme, papier, electronique et/ou autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur conserve la propriete du droit d'auteur et des droits moraux qui protege cette these. Ni la these ni des extraits substantiels de celle-ci ne doivent etre imprimes ou autrement
reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
While these forms may be included in the document page count, their removal does not represent any loss of content from the thesis.
Conformement a la loi canadienne sur la protection de la vie privee, quelques formulaires secondares ont ete enleves de cette these.
Bien que ces formulaires aient inclus dans la pagination, il n'y aura aucun contenu manquant.
Le 21 juillet 2011
lejury a accepte la these de Monsieur Jean-Philippe Couture
dans sa version finale.
Membres du jury
Professeur Richard Blouin
Directeur de recherche
Departement de biologie
Professeur Benoit Leblanc
Membre
Departement de biologie
Monsieur Andre Tchernof
Membre externe
Universite Laval
Professeur Luc R. Gaudreau
President rapporteur
Departement de biologie
SOMMAIRE
Depuis plusieurs annees, le tissu adipeux est reconnu comme un organe complexe et extremement important au maintien de Phomeostasie chez les mammiferes. En effet, une deregulation de sa taille et/ou de ses fonctions endocrines peut mener a 1'apparition de nombreuses pathologies potentiellement mortelles, telles que le diabete de type II, 1'hypertension ou encore des troubles cardiovasculaires. Ainsi, il est d'une importance capitale de bien comprendre les phenomenes moleculaires qui entourent la differenciation des cellules graisseuses afin de mieux reagir en clinique lors de l'apparition de telles conditions.
Dans cette these, j ' a i etudie le role d'une proteine kinase appelee dual leucine zipper-bearing kinase (DLK) dans la formation du tissu graisseux en utilisant un modele cellulaire murin a differenciation inductible, les 3T3-L1. A I'aide d'approches pharmacologiques et d'ARN interference, j ' a i pu demontrer que cette proteine est essentielle a Padipogenese. En effet, une diminution de Pexpression de DLK dans les cellules 3T3-L1 bloque la differenciation a un stade precoce, i.e. entre la liaison du facteur de transcription C/EBP/3 a PADN et Pexpression de ses genes cibles, tels que C/EBPa et PPARy. L'etude plus poussee des voies de signalisation des MAPKs a I'aide d'inhibiteurs pharmacologiques dans des cellules ou Pexpression de DLK a ete diminuee a revele que Paction benefique de DLK lors de la differenciation se situe au niveau de la voie ERK. En effet, I'induction de DLK au cours de Padipogenese resulte en une diminution de Passociation des proteines KSR-1, Mek et ERK, ce qui mene a une diminution temporaire de Pactivite de ERK et de la phosphorylation inhibitrice sur la serine 82 du regulateur central de la differenciation des adipocytes, PPARy. Finalement, nous avons pu determiner que I'induction de DLK au cours de la differenciation des cellules graisseuses est modulee par PPARy.
REMERCIEMENTS
Mes premiers remerciements s'adressent a mon directeur de these, le Dr. Richard Blouin, pour m'avoir accueilli dans son laboratoire, moi, petit biologiste inexperimente du Saguenay. Sa confiance, sa patience, et son empathie dans les situations difficiles ont ete maintes fois des bouees sans lesquelles la reussite de ce doctorat aurait ete compromise. Merci Richard!
Merci egalement a mes conseillers, les Drs. Benoit Leblanc et Luc Gaudreau, sans qui nous aurions ete confrontes a plusieurs impasses experimentales et theoriques, au niveau de la transcription. Leur soutien moral fut egalement tres apprecie! Un merci special au Dr. Andre Tchernof, de PUniversite Laval, qui a accepte d'evaluer ce travail a titre de membre externe. Merci aux organismes subventionnaires suivants pour avoir supporte nos recherches : CRSNG et IRSC.
Merci aux fous du laboratoire Lafontaine. Jeff, Benny, Simon, Renaud, Jeje, Anne-Marie, Pooht et meme Daniel pour leur soutien, encouragements et les excellents moments que nous avons passes ensemble. Que ce soit chez Jeje ou encore a PUniversite, vous avez tous facilite mon passage a Sherbrooke. Merci!!
Merci a mes anciens collegues de travail, Alex, Gabou et Karine, avec qui j ' a i cultive mon amour pour cette proteine capricieuse qu'est DLK. Que ce soit par votre bonne humeur ou vos expertises, vous avez positivement marque mon experience dans le labo!
Finalement, un immense merci a ma fille, a ma famille, et a Nancy, sans qui j e n'aurais jamais eu la force de continuer. Votre support inconditionnel et vos encouragements
perpetuels ont certainement aide a me garder concentre sur mes objectifs. Je sais que je n'ai pas toujours ete positif, mais c'est certainement grace a vous que je conserve un optimisme pour Pavenir.
TABLE DES MATIERES
SOMMAIRE iii REMERCIEMENTS iv
TABLE DES MATIERES v LISTE DES ABREVIATIONS viii
LISTE DES FIGURES x
1.1 Le tissu adipeux 1 1.1.1 La physiologie du tissu adipeux 2
1.1.1.1 Un reservoir energetique 2 1.1.1.2 Les fonctions endocrines du tissu adipeux 3
i ) Leptine 3 ii) Adiponectine 4 iii) Resistine, TNF-aet IL-6 5
1.1.2 Les pathologies du tissu adipeux 6 1.2 La differenciation des cellules adipeuses 8
1.2.1 : L'engagement des cellules dans la voie adipocytaire 10
1.2.2 La differenciation terminale 11 1.2.2.1 Arret de croissance et expansion clonale 11
1.2.2.2 Differenciation terminale 12
i ) C/EBPpet C/EBPS 12 ii) ADD 1/SREBP-lc 13 iii) CHOP-10 13 iv)C/EBPa 14 v) PPARy 14 1.3 La signalisation cellulaire par les MAPK 17
1.3.1 La voie p38 19 1.3.2 La voie ERK 21 1.3.3 La voie JNK 22 1.3.4 La MAPKKK DLK 24 1.4 Hypotheses et objectifs du projet de recherche 31
1.5 Presentation du modele utilise 32
CHAPITRE1 34
La proteine DLK est un regulateur central de Padipogenese 34 The mixed-lineage kinase DLK is a key regulator of 3T3-L1 adipocyte differentiation 35
ABSTRACT 36 INTRODUCTION 37
RESULTS 38 DISCUSSION 48 MATERIALS AND METHODS 52
ACKNOWLEDGMENTS 57
REFERENCES 57
CHAPITRE2 62
L'expression du gene codant pour DLK est controlee par PPARy 62
The DLK gene is a transcriptional target of PPARy 63
ABSTRACT 64 INTRODUCTION 64 MATERIALS AND METHODS 66
RESULTS 71 DISCUSSION 80 FUNDING 83 ACKNOWLEDGMENTS 83 REFERENCES 83 CHAPITRE3 88
DLK promeut Padipogenese en diminuant Pactivite de la voie ERK 88 The MAPKKK DLK promotes adipogenesis via downregulation of the ERK pathway in
3T3-L1 preadipocytes 89 ABSTRACT 90 INTRODUCTION 90 MATERIALS AND METHODS 92
RESULTS 95 DISCUSSION 101 AKNOWLEDGEMENTS 104
REFERENCES 104
DISCUSSION GENERALE 109
Implications de la MAPKKK DLK dans Padipogenese 109
Regulation de l'expression de DLK par PPARy 114 Modele propose pour le role de la MAPKKK DLK dans Padipogenese 118
CONCLUSION ET PERSPECTIVES 120
ANNEXE 1 122
DLK : The signaling gateway (Nature Publishing Group) 122
ABSTRACT 123 Protein function 123 Regulation of activity 125 Interactions with ligands and other proteins 126
Regulation of concentration 126 Subcellular localisation 127 Major sites of expression 128
Phenotypes 128 Splice variants 129 Antibodies 129 References 129 ANNEXE II 135
DLK : Encyclopedia of Signaling Molecules (Springer) 135
DLK (dual leucine zipper-bearing kinase) 136
Historical background 137 Structure, expression and subcellular localization 137
Signaling properties and regulation 138
Biological functions 140
Summary 141 Acknowledgements 142
References 142
LISTE DES ABREVIATIONS
ADN : Acide desoxyribonucleique AMP : Adenosine monophosphate ARN : Acide ribonucleique ARNm : ARN messager ATP : Adenosine triphosphate BMP : Bone morphogenetic protein C/EBP : CCAAT enhancer binding protein CREB : cyclic-AMP response element-binding ChIP : Chromatin immunoprecipitation DEX : Dexamethasone
DLK : dual leucine zipper-bearing kinase DMSO : Dimethylsulfoxyde
EMSA : Electrophoretic mobility shift assay ERK : Extracellular signal-regulated kinase FAS : Fatty acid synthase
IBMX: 3-isobutyl-l-methylxanthine JNK : c-jun N-terminal kinase kb : kilobases
kDA: Kilodalton
MAPK : Mitogen-activated protein kinase MAPKK : MAPK kinase
MAPKKK : MAPK kinase kinase MCE : Mitotic clonal expansion MEF : Mouse embryonic fibroblasts miRNA : Micro ribonucleic acid MLK: Mixed-lineage kinase PPRE : PPAR response element
rt-qPCR : Reverse transcription quantitative polymerase chain reaction Rosi: Rosiglitazone
shRNA : Small hairpin ribonucleic acid
SREBP : Sterol regulatory element binding protein WB : Western blot
LISTE DES FIGURES
INTRODUCTION
Figure 1.1 Exemple d'implications d'une surcharge ponderale et de ses effets sur la
production d'adipocytokines et sur le metabolisme general 7
Figure 1.2 Engagement des cellules dans la voie des adipocytes et differenciation
terminale 9
Figure 1.3 Schema des interactions existantes entre les principaux facteurs de
transcription regulant Padipogenese 17
Figure 1.4 Fonctionnement des voies de signalisation de ERK, p38 et JNK 19 Figure 1.5 Representation schematique de la structure primaire des formes murines et
humaines de DLK 24
Figure 1.6 Modele d'activation de la MAPKKK DLK 27
CHAPITRE 1
Figure 1. 1 Up-regulation of DLK expression and activity during 3T3-L1 adipocyte
differentiation 40
Figure 1.2 DLK depletion in 3T3-L1 impairs lipid accumulation in response to
differentiation inducers 42
Figure 1.3 Loss of DLK in 3T3-L1 cells prevents expression of C/EBPa, PPARy,
adiponectin and fatty acid synthase proteins but not that of C/EBP/S 43
Figure 1. 4 Depletion of DLK in 3T3-L1 cells blocks expression of PPARy and
C/EBPa at the mRNA levels 45
Figure 1. 5 Knockdown of DLK does not interfere with the binding of C/EBP|8 to the
cebpa and pparyl promoters 46
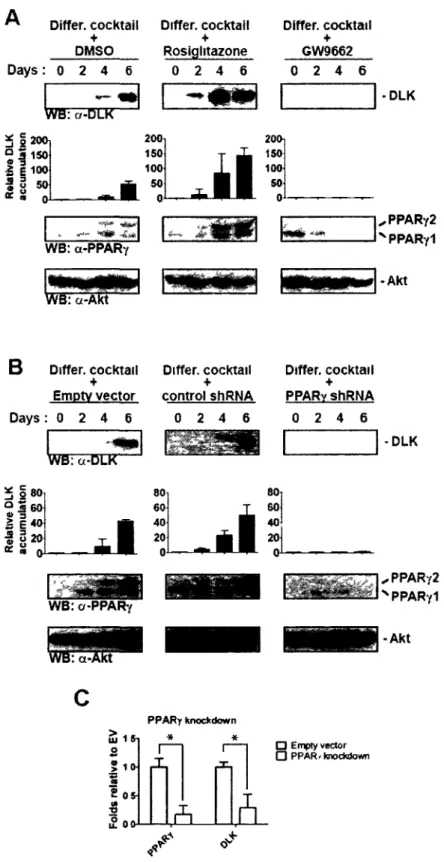
Figure 1.6 (continued) PPARy activation by rosiglitazone rescues adipocyte
differentiation in DLK-depleted 3T3-L1 cells 48
CHAPITRE 2
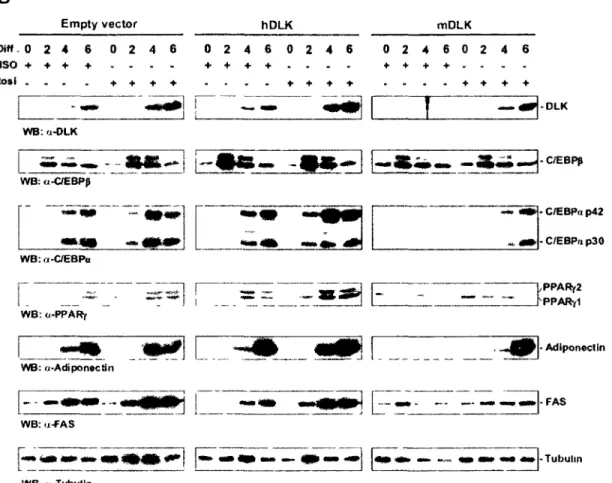
Figure 2. 1 DLK expression requires PPARy activation in differentiating adipocytes.. 72
Figure 2. 2 Rosiglitazone treatment increases DLK expression in vivo 74
Figure 2. 3 Identification of two putative PPREs in the proximal promoter of DLK... 76
Figure 2. 4 PPARy, RXRa and RNA polymerase II bind to the promoter of DLK in
differentiating adipocytes 79
Figure 2. 5 PPARy is required for DLK expression in neuronal cells 80
CHAPITRE 3
Figure 3. 1 ERK inhibition restores lipid accumulation and adiponectin expression in
DLK knockdown 3T3-L1 96
Figure 3. 2 DLK knockdown stimulates phosphorylation of ERK upstream kinase Mek
and ERK downstream substrates C/EBPP and PPARy in differentiating 3T3-L1 cells 99
Figure 3. 3 DLK associates with KSR-1 to downregulate ERK activity 100
ANNEXE 2
Figure A2. 1 Primary structures of mouse and human DLK 137
Figure A2. 2 Schematic representation of the DLK-JNK signaling pathway 139
INTRODUCTION
1.1 Le tissu adipeux
Longtemps relegue au rang de simple reservoir energetique, le tissu adipeux est aujourd'hui reconnu par les scientifiques comme etant un organe complexe jouant un role central dans une panoplie de processus metaboliques. En effet, le nombre impressionnant d'etudes effectuees depuis le milieu des annees 90 sur le sujet ont permis de mettre en lumiere une implication des cellules graisseuses, aussi appelees adipocytes, dans des processus aussi varies que la regulation du sentiment de satiete et la thermogenese, en plus d'agir comme facteur central dans la regulation de la glycemie et du controle des reserves energetiques, pour ne nommer que ceux la. Ainsi, une mauvaise regulation de la taille et des fonctions de ce tissu amenent non seulement un probleme de poids, mais aussi une deregulation systemique importante dont les repercussions peuvent s'averer mortelles, dans les cas les plus graves.
Chez les mammiferes, le tissu adipeux est letrouve sous deux formes, soient le tissu adipeux blanc et le tissu adipeux brun. Bien qu'etant tous deux des adipocytes, ces deux types cellulaires different sur plusieurs points, autant au niveau microscopique que moleculaire. La tres grande concentration en mitochondries des adipocytes bruns leur confere, par exemple, une couleur brunatre caracteristique qui les differencie aisement des adipocytes blancs. De plus, les adipocytes bruns possedent Petonnante capacite de reguler la temperature corporelle et de bruler une partie de Penergie excedentaire grace a une proteine appelee uncoupling protein 1 (UCP-1). Une fois activee, cette proteine exclusivement exprimee chez les adipocytes bruns bloque la respiration aerobique en perturbant le gradient de proton de part et d'autre de la membrane mitochondriale (Cinti, 2005). Le flux anormal d'ions ainsi cree rends plus difficile la synthese d'ATP et libere de la chaleur, ce qui permet d'augmenter localement la temperature du corps. Comme elles ne sont presentes qu'en tres petite quantite, ces cellules sont surtout retrouvees dans un depot interscapulaire et en peripheric de grandes arteres et de quelques organes chez Phomme (Cinti, 2005).
Les adipocytes blancs forment quant a eux la presque totalite des cellules graisseuses chez les mammiferes et sout responsables, a ce titre, de la majorite des phenomenes physiologiques attribues aux cellules graisseuses. Ces cellules sont aisement identifiables de par leur couleur blanchatre et sont regroupees en plusieurs depots differents chez les mammiferes, generalement classifies en depots sous-cutanes ou visceraux. Afin d'alleger le texte, le tissu adipeux blanc sera simplement appele « tissu adipeux » dans les sections a suivre.
1.1.1 La physiologie du tissu adipeux
1.1.1.1 Un reservoir energetique
Afin de permettre aux nombreuses reactions biochimiques qui assurcnt leur survie d'avoir lieu, les cellules eucaryotes doivcnt pouvoir compter sur un apport continu en energie. Pour plusieurs eucaryotes superieurs, cet apport est assure par P alimentation, a partir de laquelle de nombreux composes chimiques, dont le glucose, sont extraits et assimiles par le systeme digestif avant d'etre distribues aux cellules via le systeme circulatoire. Cependant, comme Pingestion d'aliments ne se fait pas de facon continue, une partie de Penergie fournie par Palimentation doit pouvoir etre emmagasinee et relachee de maniere controlee par Porganisme. Chez les mammiferes, ce role de « reservoir energetique » est assure par le tissu adipeux. En effet, les adipocytes sont a merae de recueillir une partie du glucose circulant dans le sang et de Pemmagasiner sous forme de lipides, suite a une stimulation adequate (Badman et Flier, 2007; Lafontan, 2008; Rosen et Spiegelman, 2006; Sethi et Vidal-Puig, 2007). Plus precisement, Pincorporation du glucose sanguin a Pinterieur des adipocytes est controlee par une hormone, Pinsuline, secretee par les cellules /3 du pancreas apres une augmentation de la concentration sanguine de glucose (glycemie) qui suit normalement les repas (Ohneda et al., 2000). La liaison de Pinsuline sur son recepteur au niveau des adipocytes declenchera une cascade de signalisation qui aura pour effet de favoriser la localisation membranaire plutot que cytoplasmique de Glut4, une proteine specialisee dans Pincorporation du glucose a Pinterieur de la cellule (Kahn, 1996). Ainsi, un relachement d'insuline dans le sang entrainera une diminution la glycemie, en plus de
favoriser le stockage d'energie sous forme de triglycerides (lipides) au niveau des adipocytes. En periode de carence energetique, ces triglycerides pourront etre hydrolyses en glycerol et en acide gras afin d'etre relaches dans la circulation et d'etre utilises comme source d'energie par Porganisme. Mentionnons que les acides gras provenant des triacylsglycerols et/ou de 1'alimentation sont egalement metabolises par la cellule grasseuse, par un processus dependant de Pinsuline. Ainsi, ces molecules pourront aussi servir de substrat pour la production d'energie par un processus de beta oxydation.
1.1.1.2 Les fonctions endocrines du tissu adipeux
Faisant partie integrante du processus de maintien de Phomeostasie generale en sa qualite de reservoir energetique, le tissu adipeux se doit de savoir reconnaitre les signaux chimiques envoyes par Porganisme et d'y repondre de facon appropriee. A Pinverse, les adipocytes doivent egalement pouvoir communiquer avec le rcste de Porganisme afin de moduler adequatement les differents aspects du metabolisme energetique qui lui sont rattaches. Ce dialogue moleculaire entre les divers organes du corps humain est en partie assure par des hormones qui, une fois secretees par differentes glandes dont le tissu adipeux, seront acheminees vers leurs cibles par le systeme circulatoire ou elles pourront induire une reponse biologique. Dans cette section, les fonctions des principales hormones produites par les adipocytes, egalement appelees adipocytokines, seront brievement relatees.
i ) Leptine
De toutes les adipocytokines identifiees a ce jour, la leptine est probablement celle qui fut la plus etudiee. Son implication dans le metabolisme energetique a ete soupconnee pour la premiere fois en 1950, alors qu'une etude rapportait qu'une simple mutation dans un gene appele ob (pour « obese ») chez la souris cause une hyperphagie severe qui resulte en un enorme surplus de poids et provoque une resistance a Pinsuline semblable a celle remarquee dans les cas de diabete de type II (Ingalls et al., 1950). Ce n'est toutefois qu'en
1994 que le groupe de Friedman a identifie la leptine comme etant le produit du gene ob (Zhang et al., 1994), et done une composante essentielle au maintien de Phomeostasie energetique. Au cours d'un cycle de 24 heures, les concentrations seriques de leptine
vanent enormement, atteignant un maximum entre 23h00 et lhOO et diminuant graduellement par la suite pour attemdre un minimum en debut d'apres-midi (Sinha et a l ,
1996) La prise de repas amene egalement une augmentation de la production et de la secretion de la leptine de maniere transitoire (Saladin et a l , 1995)
Une fois relachee dans le sang, la leptine pourra her son recepteur et induire une reponse biologique qui sera specifique au tissu ou cette liaison aura lieu Bien qu'il soit majoritairement retrouve au mveau de Phypothalamus, le recepteur de la leptine (Ob-R) est egalement present au niveau des adipocytes, des ovaires, des testicules, du placenta, du foie, des cellules p du pancreas, de la medulla de la glande surrenale, des poumons, de certaines cellules sanguines, des chondrocytes, du coeur et des muscles squelettiques (Gahc et a l , 2010) Au niveau du cerveau, la liaison de la leptine sur son recepteur provoquera une activation de la voie de signalisation Jak/Stat qui fera augmenter le sentiment de satiete (Friedman et Halaas, 1998). Comme ii existe une forte correlation entre la taille du tissu adipeux et la production de leptine, un relachement de cette adipocytokme dans le sang permet de controler Padiposite en limitant P ingestion de nournture chez les organismes ayant un surplus de poids Au niveau des muscles squelettiques, la cascade de signalisation induite par la liaison de la leptine sur son recepteur permettra plutot une plus forte oxydation des acides gras, ce qui amehorera la sensibihte a Pinsuline (Minokoshi et Kahn, 2003) II est a noter que de fortes concentrations de leptine ont toutefois ete associees a une progression de maladies lnflammatoires et/ou auto-immunes en favonsant la production de certaines cytokines pro-inflammatoires et en agissant comme un inducteur des cellules T (Stofkova, 2009), en plus d'etre associee a la progression tumorale (Percik et Stumvoll, 2009). Un dosage precis de la secretion de leptine est done necessaire afin d'eviter une surproduction et les effets nefastes qui s'y rattachent, tout en favonsant ses actions benefiques sur le metabolisme energetique
ii) Adiponectine
Contrairement a la leptine, de hauts niveaux d'adiponectine dans le sang sont associes au maintien d'une bonne signalisation par Pinsuline Cette adipocytokme clonee au milieu des annees 1990 partage certaines caractenstiques structurales avec la famille du complement lq (Clq), dont un domaine collagene et un domaine globulaire (Hu et a l , 1996b, Scherer
et al, 1995) Des ponts disulfurcs au niveau de la cysteine 39 dans le domaine collagene permettent d'ailleurs a Padiponectine de circuler sous forme d'homomultimeres composes de 3, 6 ou 18 sous-unites, alors que la forme monomenque semble confinee au niveau des adipocytes (Tilg et Moschen, 2006) Une fois liee a son recepteur, Padiponectine favonsera Pactivation de PAMPK (5'-AMP-activated protein kinase), qui elle-meme causera un blocage de la gluconeogenese au niveau du foie et une augmentation de Poxydation des acides gras au niveau des muscles squelettiques et du foie (Stofkova, 2009) Une activation des facteurs de transcnption PPARa et PPARy par Padiponectine favonse egalement une augmentation de Poxydation des acides gras et une diminution du contenu musculaire en tnglycendes (Kadowaki et Yamauchi, 2005) Ainsi, comme la sensibilite a Pinsuline est dimmuee par une trop forte concentration senque ou cellulaire d'acide gras et de tnglycendes, Padiponectine agit comme un regulateur positif de la signalisation par Pinsuline et de la regulation de la glycemie en permettant une degradation/utilisation de ces molecules nefastes a forte concentration
in ) Resistine, TNF-a et IL-6
A Pinverse de la leptine et de Padiponectine, de forts taux senques de resistine favonsent la gluconeogenese hepatique, ce qui resulte en une diminution de la reponse des cellules du foie a Pinsuline et mene a une augmentation de la glycemie (Banerjee et a l , 2004, Qi et a l , 2006) Malgre le tres grand interet du monde scientifique et medical pour cette molecule (Steppan et al., 2001), tres peu d'informations ont ete rapportees quant a la signalisation intracellulaire stimulee par la resistine De fait, le seul effet constant attnbuable a la resistine observe par plusieurs groupes, dans divers modeles, serait une diminution indirecte de Pactivite de PAMPK (Banerjee et a l , 2004, Qi et a l , 2006, Satoh et a l , 2004) De plus, ii semble que les actions biologiques de cette molecule ne soient pas les memes dans les modeles munns et chez Phomme, ce qui comphque encore d'avantage sa comprehension (Barnes et Miner, 2009, Steppan et Lazar, 2004) Finalement, P identification du recepteur de la resistine pourra certainement favonser une meilleure comprehension de la signalisation mduite par cette hormone et contnbuer a mieux comprendre les voies metaboliques touchees par sa production Comme pour la resistine (Steppan et a l , 2001), de forts taux senques de tumor necrosis factor alpha (TNF-a) et d*interleukme 6 (IL-6) sont associes a une mauvaise signalisation par Pinsuline et a une
activation du systeme immunitaire (Galic et a l , 2010). Bien que pouvant etre secretees par les adipocytes, ces deux molecules sont surtout produites par les macrophages ou par des cellules stromales presentes dans le tissu adipeux et interferent directement dans la cascade de signalisation par Pinsuline en favorisant une phosphorylation inhibitrice de la proteine IRS-1 par la MAPK JNK (Galic et al., 2010; Weisbcrg et al., 2003). Ainsi, une trop grande production de ces molecules aura comme effet de bloquer P incorporation du glucose a Pinterieur des cellules, ce qui resultera en une augmentation de la glycemie et des problemes qui y sont rattaches.
1.1.2 Les pathologies du tissu adipeux
Comme le tissu adipeux joue un role dans un grand nombre de processus biologiques, un dereglement dans Pune ou I'autre de ses fonctions peut avoir d'importantes repercussions sur Porganisme, tels qu'illustre a la figure 1.1. Ainsi, les consequences d'une prise de poids trop importante depassent de loin la simple esthetique. Les risques de maladies cardiovasculaires, d'hypertension et de diabete de type II sont effectivement tous augmentes par une surcharge ponderale, ce qui fait de Pobesite une condition tres particuliere a traiter en clinique. Cette affirmation est d'autant plus vraie que lorsque des changements dans les habitudes de vie tels que les dietes et une augmentation de la frequence de Pactivite physique sont utilises comme seul traitement, une reprise de poids ulterieure au traitement est rencontree dans la presque totalite des cas.
Surcharge ponderale
1
| Acides gras libres | Resistine t TNF-a, IL-6 t Leptine I Adiponectine t Glycemie t Apoptose cellules (J
I Secretion et signalisation par Pinsuline
Diabetc de type II Inflammation chronique Figure 1.1 Atheroscerose Accidents vasculaires Risques cardiaqucs Cancer Hypertension
Exemple d'implications d'une surcharge ponderale et de ses effets sur la production d'adipocytokines et sur le metabolisme general.
Du point de vue biologique, le developpement de Pobesite s'explique par un apport energetique superieur a la depense. Le tissu adipeux blanc sous-cutane, lieu de stockage energetique par excellence, peut evidemment agir comme tampon et absorber une grande quantite de Penergie excedentaire sous forme de triglyceride, avant d'atteindre sa capacite de stockage limite (Ibrahim, 2010). Par contre, lorsque cette limite est atteinte et que les muscles squelettiques n'arrivent plus a utiliser le glucose superflu en reponse a Pinsuline, Penergie excedentaire commence a s'accumuler massivement a des endroits atypiques tels que le tissu adipeux visceral, les muscles squelettiques et le foie (Hotamisligil, 2006;
Ibrahim, 2010; Muoio et Newgard, 2006). En plus de compromettre les fonctions normales de ces tissus, une saturation du tissu adipeux sous-cutane amene une augmentation des concentrations d'acides gras et de triglycerides dans le sang, ce qui interfere avec la signalisation a Pinsuline en bloquant la production d'adipocytokines hypoglycemiantes telles que Padiponectine (Laclaustra et al., 2007). Ainsi, un apport energetique trop important durant une periode prolongee compromet les fonctions de plusieurs organes, en plus de favoriser Petablissement d'une resistance systemique a Pinsuline et une augmentation chronique de la glycemie, qui peuvent mener a un diabete de type II.
Une inflammation chronique au niveau du tissu adipeux a egalement ete montree comme etant responsable d'un bon nombre de phenomenes moleculaires observes chez les patients souffrant d'obesite. Par exemple, une forte augmentation de l'expression de molecules reliees a Pinflammation, telles que la leptine (Galic et al., 2010), le TNF-a (Cawthorn et Sethi, 2008) et 1TL-6 (Rotter et al., 2003) a ete remarquee au niveau du tissu adipeux de personnes obeses. Ainsi, comme ces molecules ont la capacite d'interferer directement avec la signalisation a Pinsuline et de diminuer la production d'adiponectine (Bniun et al., 2003), leur augmentation pourrait contribuer a Papparition de la resistance a Pinsuline associee a Pobesite. Finalement, Pingestion massive de nourriture chez les patients souffrant d'obesite a ete liee a une augmentation du risque de developper un cancer du colon, de Poesophage, du sein, de Pendometre, du foie, du rein, de la vesicule biliaire et du pancreas (Percik et Stumvoll, 2009), soulignant encore d'avantage Pimportance d'une bonne regulation du tissu adipeux.
1.2 La differenciation des cellules adipeuses
La formation du tissu adipeux est un processus complexe et hautement regule qui interesse fortement la communaute scientifique. En effet, Petude des mecanismes moleculaires qui orchestrent la differenciation adipocytaire, ou adipogenese, peut apporter plusieurs explications sur les phenomenes qui entourent Papparition de pathologies comme Pobesite ou la lipodystrophie. En comprenant mieux la formation du tissu adipeux, les scientifiques esperent trouver des indices sur les mecanismes qui regissent Papparition de Phyper ou de Phypoproliferation des adipocytes et ainsi mieux comprendre les causes qui menent au
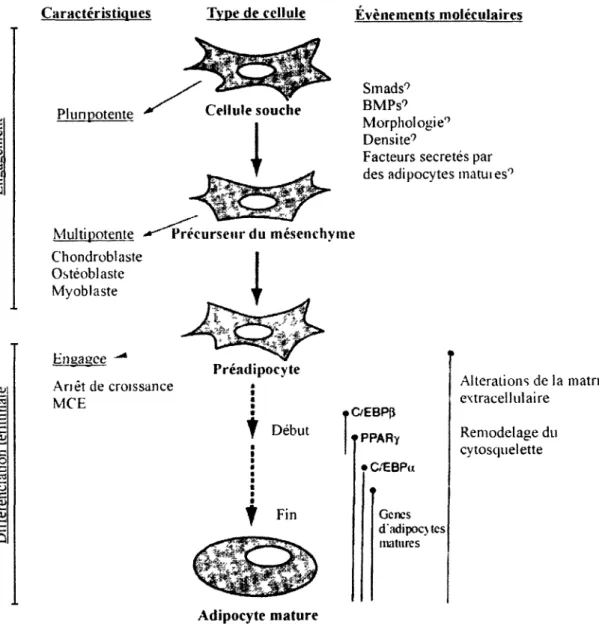
dereglement des fonctions endocrines du tissu adipeux. Tels que schematises dans la Figure 1.2, les processus moleculaires entourant les deux grandes etapes de Padipogenese, soient l'engagement des cellules totipotentes dans la voie adipocytaire et la differenciation terminale des adipocytes, seront abordes dans la prochaine section.
Caracteristiques Type de cellule
Plunpotente Multipotente Chondroblaste Osteoblaste Myoblaste Engagce -* Anet de croissance MCE Preeursetirdu mesenchyme Evenements moleculaires Smads9 BMPs9 Morphologie9 Densite9
Facteurs secretes par des adipocytes matuies9
OEBP(J f PPARY • C/EBPu
f Fin
Genes d'adipoc>tes matures Adipocyte matureFigure 1.2 Engagement des cellules dans la voie des adipocytes et differenciation terminale. (Adapte
de Gregoire et al. 1998)
Alterations de la matricc extracellulaire
Remodelage du cytosquelette
1.2.1 : L'engagement des cellules dans la voie adipocytaire
Bien qu'il ait ete montre depuis plusieurs annees que la proliferation des preadipocytes est stimulee par un ou plusieurs facteurs inconnus presents dans le sumageant d'une culture d'adipocytes matures (Considine et al., 1996; Lau et al., 1990; Marques et al., 1998), la premiere molecule identifiee comme pouvant potentiellement reguler l'engagement des cellules souches dans la voie adipocytaire a ete la proteine BMP-4 (bone morphogenic protein 4), en 2004. En effet, le traitement de cellules souches C3H10T1/2 au BMP-4 amene plus de 90% des cellules a acquerir les caracteristiques des preadipocytes et a se differencier en adipocyte mature, lorsque subsequemment stimulees avec un cocktail de differenciation classique (Tang et al., 2004). II est a noter que les cellules C3H10T1/2 non-traitees au BMP4 ou non-traitees seulement avec le cocktail de differenciation n'accumulent pas de lipides et n'expriment aucun des marqueurs de differenciation testes. Ayant ete largement etudiees pour leur capacite a faire s'engager dans la voie des cellules osseuses les cellules souches, les BMP sont des proteines de la famille du TGF/3 connues pour se Her a un recepteur membranaire et activer la voie de signalisation impliquant les proteines Smad (Shi et Massague, 2003). Dans leur etude de 2004, l'equipe du Dr. Lane a d'ailleurs montre que la proteine Smadl est bel et bien phosphorylee suite a un traitement des cellules C3H10T1/2 au BMP-4 (Tang et al., 2004). Afin de confirmer de maniere plus physiologique les resultats obtenus avec les traitements au BMP4 recombinant, la meme equipe a isole une lignee cellulaire possedant toutes les caracteristiques des preadipocytes, les A33, a partir de cellules C3H10T1/2 traitees au 5-azacytidine, un agent inhibiteur de la methylation de PADN reconnu pour induire un engagement spontane des cellules dans plusieurs voies de differenciation cellulaire. Une augmentation de I'ARN messager de BMP-4 d'un facteur de 10 ainsi qu'une plus grande activation de la voie des Smad ont ete observees dans les cellules A33 lorsque comparees a la lignee parentale C3H10T1/2, confirmant d'avantage un role potentiel de BMP4 dans l'engagement des cellules souches dans la voie adipocytaire (Bowers et al., 2006).
La morphologie des cellules souches ainsi que leur densite semblent egalement pouvoir influencer leur engagement dans une voie de differenciation. Une equipe a effectivement demontre dans une elegante etude parue en 2004 que des cellules souches humaines
avaient une tendance a se differencier en adipocytes lorsqu'elles sont ensemencees a forte densite et sont maintenues sous une forme ronde et compacte, alors qu'une plus faible densite et une morphologie plate et etendue favorise plutot la differenciation des cellules en osteoblastes (McBeath et al., 2004). Ainsi, l'engagement des cellules souches dans la voie adipocytaire semble reposer a la fois sur la secretion par les adipocytes matures d'un ou de plusieurs signaux proteiques, tels que le BMP-4, ainsi que sur l'environnement physique dans lequel elles se trouvent.
1.2.2 La differenciation terminale
Si peu d'informations sont connues sur le processus d'engagement des cellules dans la voie adipocytaire, ce n'est certainement pas le cas du programme de differenciation terminale des adipocytes. En effet, le developpement de modeles cellulaires preadipocytaires tels que les cellules 3T3-L1 (Green et Kehinde, 1975) a permis aux scientifiques d'explorer en profondeur les mecanismes moleculaires lies a la differenciation. L'acquisition du phenotype final d'adipocyte mature peut etre separe en trois grandes etapes successives, soient Parret de croissance, Pexpansion clonale, ainsi que la differenciation terminale.
1.2.2.1 Arret de croissance et expansion clonale
Au tout debut du processus de differenciation, les preadipocytes en culture subissent un arret de croissance obligatoire au point de controle Go/Gi du cycle cellulaire par inhibition de contact (Gregoire et al., 1998; Pairault et Green, 1979). Suite a une stimulation hormonale adequate, les cellules entreprendront de facon synchrone deux rondes de mitose, un phenomene appele « expansion clonale », ou MCE pour « mitotic clonal expansion» (MacDougald et Lane, 1995). Malgre quelques contradictions dans la litterature, une majorite d'etudes s'entendent sur la necessite des cellules a passer par cette etape afin d'acquerir un phenotype d'adipocyte mature. Plusieurs equipes ont effectivement demontre que le blocage du cycle cellulaire de differentes facons resulte en une inhibition de la differenciation (Reichert et Eick, 1999; Tang et al., 2003; Yeh et al., 1995). D'un point de vue moleculaire, les cascades de signalisation activees lors de la MCE permettent de preparer les cellules a la differenciation terminale en augmentant
l'expression et Pactivite de certains facteurs de transcription, tel que C/EBP/3 (Tang et al., 2005). Les genes cibles de ces facteurs de transcription pourront par la suite bloquer de maniere definitive le cycle cellulaire (Altiok et al., 1997; Johnson, 2005) et induire la differenciation terminale des adipocytes.
1.2.2.2 Differenciation terminate
Comme le sont la majorite des processus cellulaires, la differenciation terminale des adipocytes est un processus hautement regule dont le bon deroulement est assure par une apparition et une activation successive de plusieurs facteurs de transcription. Les roles, modes d'actions et interactions des principaux regulateurs de la differenciation, soient C/EBP|8, C/EBP5, ADD 1/SREBP-lc, CHOP10, C/EBPa et PPARy seront discutes dans cette section.
i)C/EBPJ3etC/EBP5
Les facteurs de transcription C/EBP/3 et C/EBPS peuvent etre consideres comme les regulateurs de premiere ligne de Padipogenese, puisque leur expression atteint leur maximum seulement 4 heures apres I'induction de la differenciation (Tang et Lane, 1999). La transcription du gene codant pour C/EBP/3 est regulee par la proteine CREB, un facteur de transcription active par phosphorylation suite a un traitement des cellules a Pinsuline (Klemm et al., 1998) et/ou a une augmentation de la concentration intracellulaire d'AMP cyclique (Gonzalez et Montminy, 1989). Deux elements de reponse a PAMP cyclique sont d'ailleurs retrouves dans le promoteur proximal du gene codant pour C/EBP/3 et sont necessaires au maintien de Pactivation transcriptionnelle du gene codant pour C/EBP/3 par CREB (Zhang et al., 2004b). La transcription du gene codant pour C/EBPS est quant a elle assuree par Pactivation du recepteur des glucocorticoi'des et sa liaison subsequente a un element de reponse aux glucocorticoi'des, au niveau du promoteur de C/EBP5 (MacDougald et al., 1994). Malgre une induction rapide, C/EBP/3 et C/EBP5 n'ont pas la capacite de se Her a PADN, et done d'induire la transcription de leurs genes cibles immediatement (Otto et Lane, 2005). Ce n'est qu'apres avoir ete phosphorylee de facon sequentielle par ERK (extracellular signal-regulated kinase) sur le residu threonine 188 environ 4 heures apres I'induction de la differenciation et par GSK.3/3 sur les residus
threonine 179 et serine 184, environ 12 heures post-induction que C/EBP/3 acquiert la capacite de Her PADN (Otto et Lane, 2005; Tang et al., 2005) et d'induire l'expression de certains de ses genes cibles, dont PPARy et C/EBPa. Finalement, il a ete rapporte que C/EBP/3 aurait un role a jouer dans la production d'un ligand endogene de PPARy (Figure 1.3), puisqu'une absence de C/EBP/3 rend des preadipocytes dependants a un ligand exogene pour se differencier correctement (Hamm et al., 2001).
ii) ADDl/SREBP-lc
La proteine ADDl/SREBP-lc (adipocyte determination et differentiation-dependent factorl/sterol regulatory element binding protein-lc) fait partie de la famille des «sterol element binding proteins» (SREBPs), qui regroupe des proteines jouant un role important au niveau du metabolisme du cholesterol et des acides gras (Horton et al., 2002). Au niveau des adipocytes, ADDl/SREBP-lc est responsable de la transcription de plusieurs genes codant pour des enzymes responsables de la synthese des acides gras et de la synthese des triglycerides (Kim et Spiegelman, 1996). L'ARN messager de ADDl/SREBP-lc augmente dans les 24 heures apres I'induction de la differenciation, et sa transcription semble etre regulee par Pinsuline (Kim et al., 1998). De plus, deux etudes de l'equipe du Dr. Spiegelman ont tisse un lien entre ADDl/SREBP-lc et la production d'un ligand endogene de PPARy (Kim et al., 1998; Kim et al., 1998). Cependant, la tenue de ces etudes n'a pas permis Pidentification du ligand en question.
iii) CHOP-10
CHOP-10, aussi appelee C/EBP^, est une autre proteine faisant partie de la grande famille des C/EBPs. Elle differe toutefois de ses consoeurs par le fait que des residus glycine et proline presents dans son domaine de liaison a PADN rendent impossible sa liaison aux elements de reponses aux C/EBP, sur PADN (Ron et Habener, 1992; Tang et Lane, 2000). Par contre, comme elle possede toujours la possibility de former des dimeres avec d'autres C/EBP via un domaine « leucine-zipper », elle agit comme dominant-negatif en empechant la liaison de C/EBP/3 aux «C/EBP regulatory elements» (Figure 1.3) (Tang et Lane, 2000). Ce phenomene pourrait etre un mecanisme supplemental assurant a la cellule que C/EBP/3 ne gagne sa capacite de Her PADN qu'au moment opportun, puisqu'une diminution de Pexpression de CHOP-10 est observee environ au meme moment ou
C/EBP/3 acquiert la capacite de Her PADN (Tang et Lane, 2000). Une surexpression de CHOP-10 dans le modele cellulaire 3T3-L1 confirme d'ailleurs cette hypothese, puisqu'elle resulte en un blocage de la liaison de C/EBP/3 a PADN, ce qui inhibe la MCE, et Padipogenese (Batchvarova et al., 1995).
iv) C/EBPa
L'initiation de la transcription du gene codant pour C/EBPa est controlee en partie par la liaison de C/EBP/3 et C/EBP5 a un element de reponse aux C/EBP, au niveau du promoteur proximal du gene, plusieurs heures apres I'induction de la differenciation (Rosen et MacDougald, 2006). L'activite anti-mitotique de C/EBPa (Umek et al., 1991) pourrait expliquer le delai d'activation de C/EBP/3 et C/EBP5 en debut de differenciation, puisque les cellules semblent devoir passer par la MCE pour acquerir le phenotype d'adipocyte mature (Tang et al., 2003). L'expression de C/EBPa amene done la fin de la MCE et induit Pexpression de plusieurs genes codant pour des proteines associees aux adipocytes telles que aP2, GLUT4 (Otto et Lane, 2005), leptine (MacDougald et Lane, 1995), et adiponectine (Qiao et al., 2005), en plus de favoriser sa propre expression (Christy et al.,
1991) et celle de PPARy (Wu et al., 1999). C/EBPa forme avec PPARy le duo de facteurs de transcription central orchestrant la differenciation terminale des adipocytes (Figure 1.3) (Rosen et MacDougald, 2006). En effet, la surexpression de C/EBPa dans un modele cellulaire de preadipocyte stimule Padipogenese sans que les cellules n'aient ete induites a I'aide d'un cocktail de differenciation (Lin et Lane, 1994). A Pinverse, la diminution de l'expression de C/EBPa a I'aide d'un ARN antisens inhibe Padipogenese dans le meme modele cellulaire (Lin et Lane, 1992) et les souris C/EBPa -/- presentent une anomalie au niveau de 1'accumulation de lipides (Wang et al., 1995).
v) PPARy
La proteine PPARy est un membre de la grande famille des recepteurs hormonaux nucleaires. Afin de permettre la transcription de ses genes cibles, PPARy doit Her son ligand et former un heterodimere avec le recepteur X de Pacide retinoi'que (RXR), egalement lie a son ligand (Brown et Plutzky, 2007). Bien que plusieurs ligands artificiels aient ete identifies et soient meme utilises comme agents therapeutiques dans certains cas de diabete de type II (Higgins et Depaoli, 2010), aucun ligand endogene de PPARy
directement lie a la differenciation terminale des adipocytes n'a encore ete identifie (Otto et Lane, 2005; Rosen et MacDougald, 2006).
PPARy existe sous deux isoformes, obtenues par epissage alternatif et usage different de promoteur, appelees PPARyl et PPARy2 (Zhu et al., 1995). L'isoforme 2, arborant 30 acides amines supplementaires a Pextremite amino-terminale est exprimee presque exclusivement au niveau des adipocytes, alors que l'isoforme 1 est retrouvee dans plusieurs autres tissus. L'expression dominante de PPARy2 au niveau des adipocytes a d'ailleurs laisse croire aux scientifiques que les deux isoformes n'auraient pas les memes roles a jouer lors de Padipogenese, et que PPARy2 serait d'avantage important que PPARyl pour la differenciation des adipocytes (Tontonoz et al., 1994a). Afin de verifier ces hypotheses, plusieurs etudes sur le sujet ont ete effectuees et ont mene a des conclusions legerement differentes. En effet, le groupe de Ren et collegues ont demontre que seule Pisoforme 2 pouvait retablir Padipogenese dans le modele cellulaire 3T3-L1 (Ren et al., 2002), alors le groupe de Mueller et collegues ont rapporte que les deux isoformes pouvaient retablir Padipogenese dans un modele de MEFs PPARy-/-, en mentionnant tout de meme que Pisoforme 2 semblait plus efficace (Mueller et al., 2002). La generation de modeles de souris PPARy2"/_ a egalement permis de demontrer
P importance de cette isoforme dans la formation du tissu adipeux et dans la sensibilite a Pinsuline (Medina-Gomez et al., 2005; Zhang et al., 2004a). Une analyse fonctionnelle de l'activite transcriptionnelle des deux isoformes de PPARy a quant a elle permis de demontrer que les 30 acides amines supplementaires de Pisoforme 2 lui conferent une activite transcriptionnelle independante du ligand plus grande que celle de l'isoforme 1 (Werman et al., 1997), mais que cette difference est perdue en condition ou le ligand est en exces (Mueller et al., 2002).
Malgre une importance non negligeable de C/EBPa dans Pacquisition du phenotype d'adipocyte mature, tous s'entendent sur Pabsolue necessite pour les cellules d'exprimer PPARy afin de proceder a la differenciation terminale (Rosen et MacDougald, 2006). Comme pour C/EBPa, la surexpression de PPARy dans des fibroblastes embryonnaires est suffisante pour induire Padipogenese (Tontonoz et al., 1994b). De plus, il semblerait qu'aucun facteur ne soit capable d'induire Padipogenese en absence de PPARy. En fait, la
plupart des facteurs qui promeuvent Padipogenese le font, en partie du moins, en favorisant Pexpression ou Pactivation de PPARy (Rosen et Spiegelman, 2006), tel que demontre par les interactions entre PPARy, C/EBPa et C/EBP/3. L'induction de l'expression de PPARy, comme celle de C/EBPa, est controlee en partie par C/EBP/3 (Wu et al., 1999). Par contre, I'induction de l'expression de C/EBPa par C/EBP/3 necessite absolument la presence et Pactivation de PPARy, a defaut de quoi un complexe forme de Phistone deacetylase HDAC1 et de mSin3A reste accroche au promoteur proximal de C/EBPa et en inhibe la transcription (Rosen et al., 2002; Zuo et al., 2006). Ainsi, apres remodelage de la chromatine en debut de differenciation au promoteur du gene codant pour PPARy, C/EBP/3 promeut l'expression et Pactivite de PPARy, ce qui permet d'activer Pexpression de C/EBPa, et done Padipogenese. L'importance centrale de PPARy dans la differenciation terminale des adipocytes est egalement supportee par la demonstration qu'une surexpression de C/EBPa dans des MEF PPARy-/- ne suffit pas a retablir Padipogenese (Rosen et al., 2002). L'inverse est toutefois possible, puisqu'une surexpression de PPARy dans une lignee de fibroblastes C/EBPa-/- resulte en une induction de Padipogenese, sans toutefois que les cellules ne soient sensibles a Pinsuline (Wu et al., 1999). II semblerait done que le role de C/EBPa soit de soutenir l'expression de PPARy, en plus de favoriser l'expression de certains genes responsables de la sensibilite a Pinsuline. En retour, PPARy permet l'expression de C/EBPa et celle d'autres genes necessaires a la differenciation terminale, ce qui fait de ces deux facteurs de transcription les maitres regulateurs de Padipogenese (Figure 1.3).
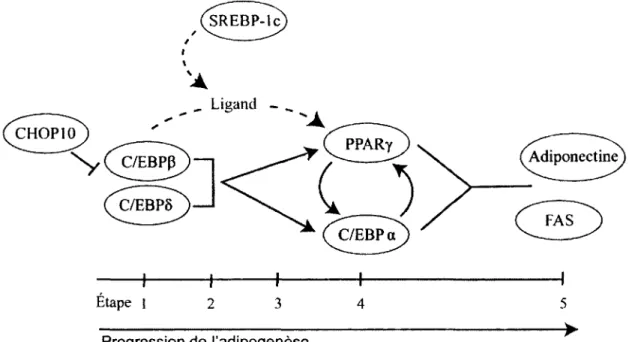
CHOP10
N^^C/EBPjT)
h
Etape 1
Progression de I'adipogenese
Figure 1.3 Schema des interactions existantes entre les principaux facteurs de transcription regulant
I'adipogenese. Etape 1 : Suite a la stimulation hormonale, C/EBP/3 et 5 sont exprimees, mais incapables de lier V ADN. Etape 2 : La MCE amene la phosphorylation de C/EBP/3 et la degradation de CHOP-10, ce qui permet a C/EBP/3 et 6 de lier l'ADN. SREBP-lc est induite et commence a favonser l'expression de ses genes cibles et d'un ligand de PPARy.
Etape 3 : Le remodelage de la chromatine permet l'expression de PPARy ct de C/EBPa. Etape 4 : PPARy et C/EBPa commencent a s'accumuler, favorisant leur expression
respective et celle de leurs genes cibles. Etape 5 : Expression de genes de differenciation terminale. Les fleches pleines indiquent un effet sur l'expression du gene, alors que les fleches pointillees indiquent un effet sur l'activite de la proteine. FAS et adiponectine represented des genes exprimes dans des cellules adipeuses matures.
1.3 La signalisation cellulaire par les MAPK
Afin de reagir correctement aux nombreux signaux chimiques qui lui sont envoyes, une cellule doit pouvoir percevoir et reconnaitre ces signaux afin de transmettre adequatement Finformation de la membrane plasmique jusqu'a Peffecteur biologique vise par le messager chimique. Cette transmission de Finformation a travers le cytoplasme est assuree par diverses proteines qui, en interagissant entres-elles, forment differentes voies de signalisation qui pourront repondre a differents stimuli. Chez les mammiferes, Pune des cascades de signalisation les plus etudiees est celle des mitogen-activated protein kinases, ou MAPK. De maniere gencralc, la transmission de Pinfomiation par les MAPK se fait par
un assemblage de plusieurs proteines en un seul complexe a I'aide d'une proteine echafaud (Bost et al., 2005). Par la suite, une proteine designee comme la MAPKKK (MAPK kinase kinase), prealablement activee par un stimulus tel qu'un facteur de croissance ou une cytokine, pourra phosphoryler et activer une MAPKK (MAPK kinase), qui pourra a son tour phosphoryler et activer la MAPK (Figure 1.4). Une fois activee, la MAPK pourra egalement phosphoryler et/ou interagir avec differentes proteines effectrices, qui meneront a l'etablissement d'une reponse biologique appropriee au stimulus de depart.
La famille des MAPK englobe plusieurs voies de signalisation differentes, dont les trois principales sont les voies de p38, ERK et JNK (nominees d'apres la MAPK de la cascade). Bien que fonctionnant sur le meme patron, ces voies de signalisation ne repondent pas exactement aux memes stimuli et n'engendrent pas les memes reponses biologiques. Comme le present projet implique plusieurs proteines de ces voies de signalisation, le detail du mode de fonctionnement de chacune de ces trois voies et leur implication dans la differenciation des adipocytes seront presentes ci-apres.
Stimuli Facteurs de croissance, cytokines Facteurs dc croissance, stress, cytokines Facteurs de croissance, stress, cytokines
M A P K K K Raf ASK 1. TAK1, MLKs
• T T T
M A P K K MEK 1/2 MKK3/6 MKK4/7
MAPK
Substrats
ERK 1/2
I i
p38a, p\
yet 8
i
T JNK1/2/3
l
Facteurs de transcription, Mks
F i g u r e 1.4 Schema de fonctionnement des voies de signalisation de ERK, p38 et JNK. Le mode de fonctionnement est schematise a gauche, alors que le detail des trois voies est indique a droite. Ce schema ne presente que certaines des proteines participant a la transmission d'un signal par les MAPK. (Mks : MAPK-activated kinases)
1.3.1 La voie p38
La proteine p38, aussi appelee SAPK2 (Stress-activated protein kinase 2), existe sous quatre isoformes et fut identifiee en 1994 comme une proteine devenant phosphorylee sur tyrosine suite a un choc osmotique et a Padministration d'une endotoxine chez les mammiferes (Han et al., 1994). Depuis, il a ete demontre que comme les autres MAPKs, p38 doit etre phosphorylee a la fois sur un residu tyrosine et un residu threonine pour etre activee (Han et al.. 1994; Kyriakis et Avruch, 2001). Les isoformes p38or et p38/3 sont
exprimees de facons ubiquitaire, alors que l'expression de p38y et de p385 est limitee a quelques tissus (Bost et al., 2005). La phosphorylation et Pactivation de la MAPK p38 est assuree principalement par deux MAPKK, appelees MKK3 et MKK6 (Enslen et al., 1998). Ces deux MAPKKs peuvent quant a elles etre activees par differentes MAPKKKs, dont ASK1, TAK1 et certaines proteines de la famille des MLK (Mixed-lineage kinase) (Bost et al., 2005; Zarubin et Han, 2005). L'activation de p38 par phosphorylation sur les residus threonine 180 et tyrosine 182 amene sa dimerisation et sa translocation au noyau, ou elle pourra phosphoryler une proteine effectrice dont Pidentite sera liee au stimulus de depart (Bost et al., 2005; Zarubin et Han, 2005). De maniere generate, la voie des p38 peut moduler une grande variete de processus, passant de Porganisation du cytosquelette au controle de la transcription de genes et/ou de la traduction d'ARN messagers, en plus d'agir sur le remodelage de la chromatine (Kyriakis et Avruch, 2001).
Le role de la voie de signalisation de p38 dans Padipogenese n'est pas clairement defini. Des resultats contradictoires ont en effet ete publies, quelques etudes suggerant un role positif de la voie de p38 dans Padipogenese, alors que d'autres plutot ont rapportes un role negatif. En effet, les travaux du groupe de Scherer ont demontre que le blocage de p38 a I'aide d'un inhibiteur pharmacologique menait a une diminution de la phosphorylation de C/EBP/3 et de Padipogenese (Engelman et al., 1998). Le meme groupe a egalement montre qu'une activation de p38 suite a l'expression d'une forme constitutivement active de MKK6, Pune des MAPKK connues pour activer p38, dans des fibroblastes non-differencies resulte en une differenciation spontanee des cellules en adipocytes (Engelman et al., 1999). D'autres ont plutot fait la demonstration que p38 joue un role positif sur l'activite transcriptionnelle de PPARy dans des cellules C3H10T1/2 traitees au BMP2 (Hata et al., 2003). Inversement, la phosphorylation de CHOP, un facteur de transcription possedant un effet dominant negatif sur les C/EBP, par p38 semble augmenter sa capacite a inhiber Padipogenese (Wang et Ron, 1996). De plus, une activation de p38 par une forme constitutivement active de MKK6 dans des adipocytes matures resulte en une augmentation de Papoptose, et une diminution constante de l'activation de p38 lors de la differenciation des 3T3-L1 a ete rapportee (Engelman et al., 1999). D'autres etudes devront done etre effectuees afin de determiner le role exact de p38 dans la differenciation adipocytaire.
1.3.2 La voie E R K
Etant Pune des voies de signalisation les mieux caracterisees chez les MAPK, la voie de ERK (Extracellular signal-regulated kinase) peut etre divisee en deux sous-groupes principaux: ERKl/2 et ERK5. Beaucoup moins etudiee que la voie ERKl/2, la voie ERK5 semble jouer un role dans le developpement cardiovasculaire (Hayashi et Lee, 2004). Les fonctions remplies par la voie ERKl/2 sont quant a elles beaucoup plus larges, et beaucoup mieux documentees. En effet, cette voie est impliquee dans plusieurs processus cellulaires, tels que la proliferation et la survie cellulaire, la regulation du cycle cellulaire et la differenciation. Les proprietes anti-apoptotiques de la voie ERK 1/2 font d'ailleurs de cette voie une cible de choix pour Papparition de mutations menant a une transformation cellulaire et a Petablissement de plusieurs types de cancers. L'activation de la voie de ERK est regulee par une pleiade de facteurs de croissance, tels que le PDGF et le EGF, en plus de repondre a certaines hormones et cytokines (Roskoski, 2010). Les MAPKK responsables de la phosphorylation de ERKl/2 sont appelees MEKl/2 et sont elles memes phosphorylees et activees, entre-autres, par la MAPKKK Raf (Bost et al., 2005). Cette derniere peut quant a elle etre activee par la petite GTPase Ras en reponse a l'activation de recepteurs membranaires a activite tyrosine kinase (Roskoski, 2010). Au niveau structural, Passemblage du complexe proteique forme de Raf, Mekl/2 et Erkl/2 est assure par la proteine echafaud KSR1, qui permet de bien positionner les kinases Pune par rapport a I'autre et favorise leur phosphorylation (McKay et al., 2009). Comme pour p38, la phosphorylation de ERK sur des residus threonine et tyrosine specifiques resulte en une activation et une dimerisation de la proteine, qui pourra par la suite effectuer Paction appropriee au niveau du cytoplasme, de la membrane cellulaire ou du noyau, selon ce que dicte le stimulus.
Plusieurs etudes sur les implications de la voie de ERKl/2 dans Padipogenese ont ete effectuees, sans toutefois que le role exact que remplit cette voie ne soit trouve. Dans les 3T3-L1, la surexpression de la forme active de Ras, une GTPase connue pour activer la voie ERKl/2, amene Parret de croissance et la differenciation terminale en adipocyte (Benito et al., 1991). Dans le meme ordre d'idee, Putilisation de I'ARN interference contre
ERKl/2 bloque Padipogenese, ce qui suggere un role positif de ERKl/2 dans ce processus de differenciation (Sale et al., 1995). Les travaux d'un autre groupe ont par contre demontre que Pactivation de ERKl/2 amene la phosphorylation de PPARy sur la serine
112, resultant en une diminution de son activite transcriptionnelle (Camp et Tafuri, 1997). De plus, ERKl/2 ont la capacite de reduire la sensibilite des cellules a Pinsuline, en phosphorylant IRS-1 sur un residu serine (Taniguchi et al., 2006), ce qui bloque la transmission du signal et empeche Pincorporation de glucose a Pinterieur de la cellule. D'autres resultats montrent par contre que l'activite de ERKl/2 n'est presente qu'au tout debut de la differenciation, jouant un role critique dans la MCE en phosphorylant et en activant C/EBP/3 (Tang et al., 2005). De plus, une activation constitutive de ERK par une surexpression de la proteine echafaud KSR1 resulte en une inhibition de Padipogenese, tout comme le fait une inhibition totale de la voie par une depletion de KSR1 (Kortum et al., 2005). Ces resultats suggerent done que le niveau d'activation de ERK dans les adipocytes doit etre regule de maniere tres precise, autant au niveau de la force d'activation qu'au niveau temporel, afin de permettre a la differenciation d'avoir lieu.
1.3.3 L a voie J N K
La voie de signalisation de JNK (c-jun N-terminal kinase), egalement appelee SAPK (stress-activated protein kinase), peut etre activee par divers stress subis par la cellule, allant du choc osmotique aux rayons ultraviolets, en plus de repondre a certains facteurs de croissance et cytokines (Kyriakis et Avruch, 2001). Les trois isoformes connues de cette proteine, JNKl, JNK2 et JNK3, existent chacune sous une forme courte et une forme longue, de 46 et 54 kDa, respectivement (Johnson et Nakamura, 2007). Les isoformes JNKl et JNK2 sont exprimees de facon ubiquitaire, alors que Pexpression de l'isoforme 3 est restreinte au niveau du cerveau, du coeur et des testicules (Johnson et Nakamura, 2007). Chacune des trois isoformes semble posseder des capacites de liaison distinctes a differents facteurs de transcription et ne sont pas activees par les memes stimuli, ce qui amene une tres grande diversite dans les reponses activees par cette voie de signalisation (Gupta et al., 1996). Les differentes isoformes de JNK peuvent etre phosphorylees par deux MAPKK, appelees MKK4 et MKK7 (Johnson et Nakamura, 2007). Comme pour p38 et ERK, les isoformes de JNK doivent etre phosphorylees sur un residu tyrosine et sur un residu
threonine par les MAPKK pour etre actives. Au moins 14 MAPKKK sont connues pour activer la voie MKK4/7-JNK, dont DLK (Dual Leucine zipper-bearing Kinase), la proteine sur laquelle les travaux de la presente these ont ete effectues. Comme les deux autres voies des MAPK, l'activation de la voie de JNK passe par Passemblage d'un complexe proteique a I'aide de JIP (JNK interacting protein), Pune des proteines echafaud associees a cette voie (Bost et al., 2005). Etant une voie de signalisation pro-apoptotique, une suractivation de JNK est impliquee dans plusieurs pathologies. En effet, si le bon fonctionnement de la voie est necessaire a bien des egards, une activation abusive de cette voie peut mener a des inflammations chroniques (comme remarque dans les cas d'obesite), a la surdite ou a Petablissement de maladies neurodegeneratives (Johnson et Nakamura, 2007). Une inhibition complete de la voie n'est guere plus recommandable, puisque Pexpression d'une forme inactive de MKK4 dans des cellules souches embryonnaires resulte en une transformation cellulaire, caracterisee par une forte diminution de Pactivation de JNK et de p38 ainsi qu'une activation constitutive de ERK (Cazillis et al., 2004).
A ce jour, aucune etude n'a pu lier de facon mecanistique la voie de signalisation de JNK a Padipogenese. Malgre le fait que comme la voie de ERK, la voie de JNK soit activee par Pinsuline (Taniguchi et al., 2006), aucun apport positif a Padipogenese n'a ete trouve pour JNK. Fait interessant par contre, il a ete montre que cette voie de signalisation joue un role dans le developpement de Pobesite. Des souris JNKl-/- (mais non JNK2-/-), ont demontre une resistance au developpement de Pobesite induite par une diete grasse et avaient une meilleure sensibilite a Pinsuline que les souris sauvages (Hirosumi et al., 2002). De plus, des souris JIP 1 -/-, Pune des proteine echafaud jouant un role dans la regulation de la voie JNK, montrent un phenotype semblable, soit une diminution du developpement de Pobesite induite par une diete grasse et une meilleure sensibilite a Pinsuline (Jaeschke et al., 2004). Ainsi, considerant que Pactivation de JNK par Pinsuline amene une diminution de l'efficacite de la signalisation par Pinsuline par une boucle de retro inhibition (Taniguchi et al., 2006) et que JNK semble etre impliquee dans Petablissement de Pinflammation chronique lors de Pobesite (Hirosumi et al., 2002), Pinhibition de cette voie pourrait s'averer interessante comme traitement contre Pobesite.
1.3.4 La MAPKKK DLK
Clonee pour la premiere fois en 1994 et initialement nominee ZPK (zipper protein kinase), la MAPKKK DLK a ete identifiee dans le cadre d'une etude visant a isoler diverses proteines kinases dont l'expression varie suite a la differenciation de teratocarcinomes humains en neurones (Reddy et Pleasure, 1994). Par la suite, les homologues de ZPK chez la souris et le rat ont rapidement ete identifies et nommes DLK et MAPK upstream kinase (MUK), respectivement (Blouin et al., 1996; Hirai et al., 1996; Holzman et al., 1994). Tout comme les autres membres de la famille des MLKs dont elle fait partie, DLK possede des caracteristiques structurales uniques chez les proteines kinases. En effet, elle presente un domaine catalytique hybride entre ceux retrouves chez les serines/threonines kinases et les tyrosines kinases, en plus d'arborer deux domaines « leucine zipper» essentiels a sa dimerisation et a son activation (Nihalani et al., 2000). Deux domaines riches en proline, potentiellement importants pour la formation d'interactions proteine-proteine et/ou pour sa localisation cellulaire, ont egalement ete identifies a chaque extremite de la proteine (Figure 1.5). T ~ DLK murin DLK humain «r I O O X O i n f-ooo> GP O C T T - (0<O i i i DK LZ GSP mctf © « N GP DK LZ GSP
Figure 1.5 Representation schematique de la structure primaire des formes murines et humaines de
DLK. (GP : Domaine riche en glycine/proline, DK : Domaine kinase, LZ : Leucine zipper, GSP : Domaine riche en glycine/serine/proline)
Une analyse transcriptionnelle de divers organes humains ou murins a revele que DLK est majoritairement exprimee au niveau des reins et du cerveau (Blouin et al., 1996; Holzman et al., 1994; Nadeau et al., 1997; Reddy et Pleasure, 1994). De maniere plus precise, son expression a ete localisee dans les zones subventnculaire et intermediaire de cerveaux de souris embryonnaires, preferentiellement au niveau des axones ou elle est fortement
associee aux microtubules (Hirai et al., 2002). Cependant, DLK a egalement ete retrouvee au niveau de Pappareil de Golgi dans des cellules corticales embryonnaires en culture (Hirai et al., 2002) et des NIH 3T3 (Douziech et al., 1999), en plus d'apparaitre pendant une courte periode au noyau, suite a un dommage cellulaire dans des cerveaux de rats (Merritt et al., 1999). D'autres equipes ont aussi rapporte l'expression de DLK au niveau de cellules /3 du pancreas (Plaumann et al., 2008), du foie (Douziech et al., 1998) et des keratinocytes (Germain et al., 2000; Robitaille et al., 2005).
Comme cite plus haut, DLK agit principalement a titre de MAPKKK dans la voie de JNK (Fan et al., 1996; Hirai et al., 1996; Robitaille et a l , 2005; Robitaille et al., 2008), potentiellement par une phosphorylation directe de MKK4 et/ou MKK7, deux MAPKK capables d'activer directement JNK (Hirai et al., 1996; Merritt et al., 1999). Cependant, seule une interaction avec MKK7 a ete identifiee in vitro (Merritt et al., 1999). Un role pour DLK dans Pactivation d'autres voies de signalisation, dont celles de ERK et de p38, a egalement propose (Daviau et al., 2009; Fan et al., 1996; Nakata et al., 2005; Robitaille et al., 2010). Bien que certaines des cibles de DLK soient bien identifiees (Merritt et al., 1999), peu d'informations sur les mecanismes d'activation ou de suppression du signal par DLK ont ete rapportees a ce jour. Les donnees actuelles suggerent que la dimerisation ou Foligomerisation de DLK est un pre requis a son autophosphorylation, activation et a la stimulation de la voie de JNK (Nihalani et al., 2000; Robitaille et al., 2008). Cette idee est d'ailleurs supportee par la demonstration qu'une oligomerisation forcee par la formation de liens covalents entre deux ou plusieurs unites de DLK par la tissue transglutaminase (tTG) suite a un traitement de fibroblastes avec un agent pro apoptotique (Hebert et al., 2000) resulte en une forte augmentation de l'activite catalytique de DLK et mene a l'activation de JNK (Robitaille et al., 2004). En plus d'une autophosphorylation, la phosphorylation de DLK par d'autres kinases semble egalement etre d'une grande importance pour la regulation de son activite. II a effectivement ete rapporte qu'une fraction du pool de DLK cytoplasmique est phosphoryle a I'etat basal in vivo, et que cet etat de phosphorylation augmente dramatiquement suite a un traitement a Pacide okadaique, un inhibiteur des serine/threonine phosphatases PP1 et PP2A (Mata et al., 1996). Cette etude demontre egalement que la dephosphorylation de DLK par la PP2B survient in vivo suite a une depolarisation membranaire de neurones en culmre (Mata et al., 1996). De plus, des
travaux recents dans notre laboratoire ont demontre qu'un traitement au PDGF et au sodium orthovanadate, un inhibiteur des tyrosines phosphatases, dans la lignee cellulaire NIH-3T3 amene la phosphorylation sur tyrosine de DLK par Src (Daviau et al., 2009). Finalement, il a ete rapporte que l'activite de DLK pourrait egalement etre modulee par la calcineurine, une serine/threonine phosphatase activee par une depolarisation membranaire au niveau des cellules /3 du pancreas, bien qu'aucune donnee ne confirme d'interaction directe entre les deux proteines (Plaumann et al., 2008). L'ensemble de ces resultats suggere done que l'activite de DLK pourrait etre regulee par une phosphorylation a la fois sur des residus serine/threonine et sur des residus tyrosine, dont Pidentite reste a etre determinee.
D'autres interactions proteiques pouvant reguler l'activite de DLK ont egalement ete rapportees par plusieurs groupes de recherche. En effet, la liaison de DLK a la proteine echafaud JIP ou a MUK inhibitory protein (MBIP) en condition basale semble jouer un role important dans sa regulation en bloquant sa dimerisation et son activation (Fukuyama et al., 2000; Nihalani et al., 2001). Cette sequestration a I'etat monomerique permet de prevenir une activation constitutive de la voie de JNK et des autres cibles de DLK en absence de signal. Toutefois, suite a une stimulation adequate, JNK pourra se lier a JIP et provoquer le relachement de DLK (Nihalani et al., 2001). Une fois libre, DLK pourra former un dimere, s'autophosphoryler et contribuer d'avantage a l'activation de la voie de JNK (Figure 1.6).
En plus des interactions transitoires inhibitrices entre DLK et JIP/MBIP, la diminution des niveaux intracellulaires de DLK est egalement fortement utilisee afin de moduler a la baisse son activite. Une etude menee dans des cellules embryonnaires de souris a effectivement revele qu'une forme active de DLK pouvait interagir avec Hsp70 et sa co-chaperone CHIP, une E3 ubiquitine ligase, ce qui promeut sa poly-ubiquitinylation et sa degradation subsequente via le proteasome (Daviau et al., 2006). La regulation des niveaux de DLK via sa poly-ubiquitinylation semble etre un mecanisme hautement efficace, puisque ses orthologues chez la mouche du vinaigre {Drosophila melanogaster) et le ver Caenorhabditis elegans sont egalement regules a la baisse par une E3 ubiquitine ligase,
appelees Highwire et RPM-1, respectivement (Collins et al., 2006; Nakata et al., 2005; Wu et al., 2007). L'orthologue de Highwire/RPM-1 chez les mammifere, appelee Phr-1, a egalement ete identifie comme pouvant potentiellement reguler les niveaux cellulaires de DLK dans les neurones (Lewcock et al., 2007). Cependant, comme aucune augmentation des niveaux de DLK n'a ete remarquee dans les neurones de souris depletees en Phr-1, davantage d'evidences experimentales seront necessaires afin de Her Phr-1 a la regulation des niveaux de DLK (Bloom et al., 2007). L'ensemble de ces donnees suggere que le choix entre une degradation de DLK ou une simple inhibition de son activite sera fait en fonction de la nature et de la duree du stimulus, du type cellulaire et de Pidentite des proteines avec lesquelles elle interagit.
A B C
Figure 1.6 Modele d'activation de la MAPKKK DLK. A) DLK est maintenue sous une forme
monomerique inactive par la proteine echafaud JIP. B) Suite a une stimulation, JNK se lie a JIP, ce qui amene une diminution de l'affinite de DLK pour JIP et son relachement. C) DLK peut alors d'homodimeriser, s'autophosphoryler et activer la signalisation par la voie JNK.
A ce jour, les mecanismes de controle transcriptionnel ou traductionnel du gene codant pour DLK sont encore mal compris. En fait, mis a part les resultats presentes dans cette these, seules deux etudes se sont penchees sur la question et ont contribue a faire avancer les connaissances sur le domaine. D'une part, il a ete rapporte en 2004 que l'expression de ZPK, la forme humaine de DLK, pouvait etre regulee positivement par le facteur de transcription SP3, mais non par les autres facteurs de la meme famille, tels que SP1, SP2 et SP4 (Itoh et al., 2004). Cependant, les evidences rapportees dans ce manuscrit ne demontraient pas hors de tout doute que cette regulation s'effectue ventablement in vivo,