Transposable elements and human cancer: A causal relationship?
Texte intégral
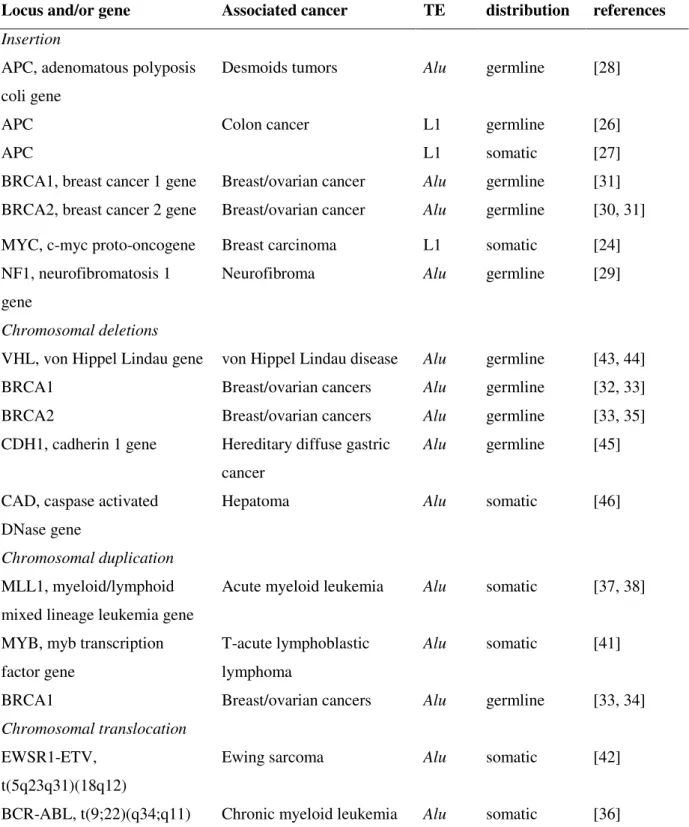
Figure
Documents relatifs
Modify pharmacokinetics; bypass [88,89] drug resistance; target AML cells In vitro evaluation [118] Ex vivo, in vitro, and Modify administration route [112–114] in vitro
sFLc (expressed in pg/mL) was assayed from thawed plasma samples by ELISA (DY308, R&D Systems, Minneapolis, MN). All patients provided informed consent. The trial was registered
An Architecture for Describing SNMP Management Frameworks http://www.ietf.org/ internet-drafts/draft-ietf-snmpv3-arch-01.txt SNMPv3 Applications
In a family of proteins encoded by multicopy genes, names will, there- fore, commonly be used to specify a class of related orthologs and paralogs rather than a specific protein
The analysis of the islands shape reveals that the electric current biases significantly the kinetic rate of mass transfers at step edges modifying the overall island shape..
Interne Forces : • Taille, 6 villages-stations • Conseil d’administration représentatif des principaux acteurs de la destination • Certification « Valais Excellence » •
Our method for proving Theorem 1.2 (i) is rather general and only requires some simple hypothesis on the maximal degree and the diameter of the graph, which is satisfied in
The present study was an evaluation of proliferation, cytotoxicity, differentiation and secretion of AML cell lines (U937, NB4, HL-60, MonoMac-6) in response to treatment with
