Département de Génie des Procédés
THÈSE
Présentée par
Amel DJEHAL
Pour l’obtention du grade de
Docteur en sciences
Option: GENIE DES PROCEDES PHARMACEUTIQUES
THEME
Synthèse de ligands promélanogéniques des prohibitines
Soutenue le, 03 Mars 2019 devant le jury:
BAITICHE Milad Professeur Univ.Setif 1 Présidente
BENTOUHAMI Embarek Professeur Univ.Setif 1 Directeur de thèse DESAUBRY Laurent Dir. Recherche CNRS Univ.Strasbourg Co-Directeur BELHAMEL Kamel Professeur Univ.Bedjaia Examinateur SOBHI Widad MCA Univ.Setif 1 Examinatrice BENMEKHBI Lotfi MCA Univ.Constantine 3 Examinateur GALONS Hervé Professeur Univ.Paris Descartes Membre
invité
leur soutien, leurs encouragements pendant ces longues années d’études. Grace à
leur présence, leur grande patience, leur compréhension, leur générosité, j’ai pu
aller au bout de mes rêves.
Il me sera impossible de rendre tout ce qu’ils m’ont offert. Rien n’aurait été
possible sans eux. Cette réussite est donc la leur avant qu’elle soit la mienne.
Aucun remerciement ne pourrait être suffisant.
A mes nièces, Emilie et Elena. Que ce modeste travail soit l’expression des vœux
que vous n’aviez cessé de formuler dans vos prières. Que Dieu vous protège et
préserve santé et longue vie.
A mes sœurs Nardjes, Lilia et Amira pour l'amour, l'aide et le soutien qu'elles m'ont
toujours apporté aussi bien dans les moments difficiles que radieux.
direction du Professeur Embarek BENTOUHAMI et du Laboratoire d’Innovation Thérapeutique de la Faculté de Pharmacie de Strasbourg sous la direction du Docteur Laurent DESAUBRY.
Mes remerciements s’adressent au Professeur Embarek BENTOUHAMI, mon directeur de thèse, pour m’avoir encadré, dirigé, orienté durant toutes ces années où j’ai travaillé avec lui. Merci de m'avoir donné l'opportunité de travailler sur un sujet d’intérêt majeur pour le laboratoire, et d’avoir été un deuxième père pour moi. Merci pour son aide précieuse, son soutien, sa disponibilité et ses encouragements qui m’ont fait apprécier le monde de la recherche.
Je souhaite exprimer ma profonde reconnaissance envers le Docteur Laurent DESAUBRY, Directeur de recherches au CNRS Strasbourg et chef d’équipe au Laboratoire d’Innovation Thérapeutique à la Faculté de pharmacie de d’Illkirch. Merci de m’avoir accueilli dans son laboratoire pendant ces années, de m’avoir fait confiance,et de m’avoir encadré. Je le remercie également pour ses connaissances, ses conseils, sa grande disponibilité, pour tous les moments passés à discuter du projet et pour la valorisation de mon travail. Merci aussi pour la sympathie qu’il m’a témoignée au cours de toutes ces années de thèse.
Mes remerciements s’adressent également au professeur Milad BAITICHE, du département de Génie des procédés de la faculté de technologie de l’Université Ferhat Abbas Sétif 1, d’avoir accepté de présider ce jury. Qu’elle trouve ici la preuve de ma reconnaissance.
Merci au Professeur Widad SOBHI de la faculté de Biologie de l’Université Ferhat Abbas Sétif 1 et au Professeur Kamel BELHAMEL de l’Université Abderahmane Mira de Bedjaia d’avoir bien voulu évaluer ce manuscrit de thèse et l’enrichir par leurs recommanadations pertinentes.
Toute ma gratitude au Professeur Hervé GALONS, de l’Université Paris Descartes d’avoir accepté d’évaluer mes travaux de recherche et d’apporter les corrections nécessaires.
Je voudrais enfin exprimer ma reconnaissance au Docteur Lotfi BELMOKHBI, du Département de génie des procédés de l’Université Constantine 3 pour avoir eu l’amabilité de consacrer son temps à l’examen de mes travaux de thèse.
Je remercie chaleureusement les thésards, les stagiaires et post-doc côtoyés pendant ces années, compagnons de fortune mais aussi d’infortune dans notre quotidien de « chercheur » : Sabira, Redouane, Hussein, Laura, Fabrice, Hassan, Jordan, Fun. Merci pour votre compagnie au laboratoire. Merci pour toutes ces discussions, plus ou moins chimiques selon les intervenants et les circonstances…
Merci aussi à tous les thésards et membres des autres laboratoires que j’ai eu l’occasion de rencontrer et avec qui j’ai eu le plaisir de passer ces années.
J’adresse mes sincères remerciements au Service Commun d’Analyse de la Faculté de Pharmacie d’Illkirch.
Mes remerciements s’adressent aussi à mes amis : Jean Luc, Barbara, Akiko pour leur soutien permanent.
Je tiens à remercier le Professeur Eric RAYMOND (Chef de service en Oncologie Medicale chez Groupe Hospitalier Paris Saint Joseph, Paris), le Professeur Ghanem Elias GHANEM (Directeur du Laboratoire d'Oncologie et de Chirurgie Expérimentale du centre oncologique Jules Bordet et Professeur de Radiopharmacologie et de Médecine Nucléaire au Centre Hospitalier Universitaire de Bruxelles), le docteur Mohammad KRAYEM de l’équipe du professeur Ghanem GHANEM d’avoir testé mes produits.
Je tiens à exprimer mes sincères remenrciments au professeur Thierry CRISTEIL (IPST, Faculté de pharmacie, Université Paris Sud), le docteur Canan G.NEBIGIL (UMR7242, CNRS/Université de Strasbourg, Ecole Supérieure de Biotechnologie de Strasbourg, laboratoire de Biomolecules, Université de Sorbne, Paris), le professeur Dong WANG et le professeur Peng YU (Sno-French Joint Lab of Food Nutrition /Safty and médicinal Chemistry. College de Biotechnologie, Université de science et de téchnologie de Tianjin, Chine).
Finalement, je remercie mon ancien directeur le Professeur BOUSSEBOUA, mon nouveau directeur le Professeur KHLIFI (Directeur de L’ENSB Constantine), Monsieur Reda BENMOURALLAH (Directeur du CACQE de Constantine), et tous mes anciens collègues du CACQE, pour leur sympathie et leurs encouragements.
Å Angström
Ac Acétyle
AcOEt Acétate d’éthyle
ADN Acide Désoxyribonucléique ARN Acide Ribonucléique AIF Apoptosis Inducing Factor
Ar Aryle
DIPEA N,N-diisopropyl éthylamine
Bn Benzyle
n-BuLi n-butyllithium
DBU 1,8-diazabicyclo[5.4.0] undec-7-ène
DCC N,N’-dicyclohexyl
carbodiimide
DDQ 2,3-dichloro-5,6-dicyano-1,4-benzoquinone
DIAD Diisopropyl azodicarboxylate DIBAL-H Hydrure de diisobutylaluminium DMDO diméthyldioxirane DME 1,2-diméthoxyéthane EDCI 1-éthyl-3-(3-diméthyl aminopropyl) carbodiimide EDTA Ethylène diamine tétra
acétate
eIF4 Eukaryotic initiation factors
eq. équivalent
ESIPT Transfert de proton à l’état excité
ESI-TOF Electron spray ionisation-time of flight
Et éthyle
EtOM éthoxyméthyle FL Flavagline
FT-IR Infrarouge à transformée de Fourier
HOBt Hydroxybenzotriazole HMPA Hexaméthylphosphoramide HRMS Spectre de masse
haute-résolution
Hsp Heat shock protein Im Imidazole
i.p. Intrapéritonéale IR Infrarouge i.v. Intraveineuse
LDA Diisopropylamidure de lithium mCPBA Acide 3-chloropéroxybenzoique
Me Méthyle
NBS N-bromosuccinimide
NI pas d’inhibition à la
PhMe Toluène PMBCl Chlorure de para-méthoxybenzyle Piv Pivaloyle Rdt Rendement RMN Résonance magnétique nucléaire
ROS Espèces oxygénées réactives SiRNA Petit ARN d’interférence TA Température ambiante
TFA Acide trifluoroacétique TFE Trifluoroéthanol THF Tétrahydrofurane TMS Triméthylsilyle
TMSNCO Isocyanate de triméthylsilyle Ts Tosyle
Tableau 1. Directives recommandées pour les propriétés des sondes chimiques……….5
Tableau 2. Cytotoxicité dans les cellules HCT15 (IC50, nM)……….7
Tableau 3. Relation structure-activité de la mélanogénine et de ses analogues………..8
Tableau 4. Essais d’oxydation du composé 9……….35
Tableau 5. Les analogues 16a synthétisés………39
Tableau 6. Les analogues 16b synthétisés………..40
Tableau 7. Les analogues 16c synthètisés………41
Tableau 8. Les analogues 16d synthétisés………..42
Tableau 9. Les analogues 16e synthètisés………44
Tableau10. Pourcentage d'induction de la mélanogenèse dans la lignée de cellules de mélanome humain HBL………..……….47
Tableau 11. Pourcentage d’inhibition de la croissance cellulaire par la mélanogénine à 10-5 et 10-6M sur differentes lignées cellulaires cancereuses dans le DMSO, mesuré après 72h d’exposition………..62
Tableau 12. Inhibition de de la croissance cellulaire par la mélanogénine, le Mel9, le Mel41 rapport aux cellules sur les cellules KB et HL60, mesuré après 72h d’exposition……….………….63
Figure 4. Requis structuraux des flavaglines pour la cytotoxicité dans les cellules cancéreuses
(FL3: R1=R2=OMe, R3=OH, R4= R5= R6= R7= R9=H, R8=Br)……….………..7
Figure 5. Taux de survie des souris traitées avec la doxorubicine(31%) ou doxorubicine + 5 injections de FL3 (0,1 mg / kg i.p.)……….………….8
Figure 6. Mécanisme cardioprotecteur de FL3 proposé par le Dr Désaubry et al………..9
Figure 7.Exigences structurelles pour la radioprotection et la cytotoxicité des cellules cancéreuses………..9
Figure 8. Structure et neurotoxicité de l’Aftine-4……….10
Figure 9. Structure de la fluorizoline………10
Figure 10. La mélanogenèse ou la synthèse de la mélanine est impliquée dans des voies réactionnelles enzymatiques et biochimiques complexes appelées voie de Raper-Mason………11
Figure 11. Principales voies régulatrices de la mélanogenèse, cette figure a été adaptée de Pillaiyar et al et légèrement modifiée……….………..13
Figure 12. Mode d’action de LC3 lié à la PHB2 lors de la mitophagie………..16
Figure 13. Structure de la mélanogénine………....17
Figure 14. Mécanisme proposé de l’action de la mélanogénine………....17
Figure 15. Découverte de la mélanogénine………..19
Figure 16. Identification d'un agent améliorant la pigmentation qui se lie à la prohibitine……….20
Figure 17. Les analogues 16a……….…39
Figure 18. Les analogues 16b ...39
Figure 19. Les analogues 16c……….42
Figure 20. Les analogues 16d………43
Figure 21. Les analogues 16 e………44
Figure 22. Effet sur la pigmentation cellulaire des analogues de la mélanogénine………53
Figure 23. Effet de Mel9 et Mel 41 sur la pigmentation cellulaire dans les lignées cellulaires MM162 et MM043………..54
phosphorylation d'Akt pour induire l'apoptose dans les cellules du mélanome ..………..….60 Figure 28. Le ligand de PHB Mel9 régule à la baisse le niveau de protéine des PHB et la
phosphorylation de Akt dans les cellules de mélanome MM043 et MM162……….61 Figure 29. . Induction de la dégradation de PHB1 et l’activation de la signalisation de MITF dans les cellules de mélanome HBL par le MEL9, le MEL41 et la mélanogénine……….. ..62 Figure 30. Dégradation induite de la PHB1 et inhibition de singalisation AKT dans les cellules HL60 par le Mel9 et Mel41.………...………63 Figure 31. Modèle proposé d'induction de la mélanogénèse par les dérivés de la mélanogénine ………..……….65
Schéma 4. Condensation du pentafluorophénol 4 sur la dichlorotriazine 3 ...33
Schéma 5. Double substitution nucléophile d’une benzylthiol sur la trichlorotriazine selon les travaux de T. Masquelin et al. ………...34
Schéma 6. Essai de développement d’une méthode de synthèse de triazines substituées par un groupement pentafluorophenoxy faisant appel à des intermédiaires thioétehers et sulfoxydes………..34
Schéma 7. Troisième approche pour la mise au point de méthode de synthèse de la mélanogenine et de ses analogues………36
Schéma 8. Travaux de Katharina Reichenbacher et al……….………36
Schéma 9. Approche finale permettant d’accéder à la mélanogénine et à ses dérivés………..37
Schéma 10. Synthèse des amines Y et Z non-commerciales………..38
Schéma 11. Synthèse de la mélanogénine………..45
Schéma 12. Synthèse de l’analogue de la mélanogénine couplé au groupement espaceur et à l’affigel10……….46
1.1 Les prohibitines ... 1
1.1.1 Signalisation et rôles physiologiques ... 1
1.1.2 Importance actuelle des sondes chimiques en biologie cellulaire ... 4
1.1.3 Ligands des prohibitines ... 6
1.1.3.1 Les flavaglines ... 6
1.1.3.2 L’Aftine-4 ... 10
1.1.3.3 La fluorizoline ... 10
1.2 Découverte et propriétés de la mélanogénine ... 11
1.2.1 Mélanine et mélanogenèse ... 11
1.2.2 Protéines régulant la mélanogénèse : ... 14
1.2.2.1 MITF : Microphthalmia-associated transcription factor ... 14
1.2.2.2 ERK : Généralités et relations avec le MITF et la mélanogenèse ... 15
1.2.2.3 LC3 : Généralités et relations avec la mélanogenèse, la PHB2 et ERK ... 15
1.2.3 Découverte et effets pharmacologiques ... 17
1.2.4 Relations structure-activité ... 19
1.3 Chimie des triazines ... 29
2 Objectifs ... 32
4 Résultats biologiques ... 47
4.1 Stimulation de la pigmentation cellulaire ... 47
4.2 Mel55 interagit avec PHB1 ... 55
4.3 Mel9 et le Mel41 régulent à la hausse le MITF et la tyrosinase grâce à l'activation de LC3 ……….55
4.4 Mel9 et le Mel41 régulent à la baisse les PHB et AKT pour induire l'apoptose dans les cellules de mélanome HBL ... 59
4.5 Effet de la mélanogénine dans les cellules de mélanome HBL ... 61
4.6 Effet sur la croissance des cellules cancéreuses ... 62
4.7 Discussion ... 64
5 Conclusion générale ... 68
6 Partie expérimentale : ... 70
1 La Prohibitine-1 (PHB1) et son homologue PHB2 sont des protéines d'échafaudage agissant comme un hub pour de nombreuses voies de signalisation activées par des facteurs de croissance, de réponse immunitaire et d’hormones stéroïdes, pour réguler le métabolisme, la dynamique et la fonction mitochondriale, la prolifération cellulaire et la survie.1 En raison de leur effet régulateur sur diverses voies de signalisation cellulaire, les PHB sont impliquées dans l'étiologie des cancers, des maladies inflammatoires, cardiovasculaires, neurodégénératives, du diabète, de l'obésité et dans la réponse immunitaire.2,3 De nombreuses modifications post-traductionnelles des PHB modulent leur activité et également leur distribution intracellulaire, dans les mitochondries, le noyau, le réticulum endoplasmique et la membrane plasmique. Les fonctions spécifiques de chacune de ces différentes formes de PHB ne peuvent pas être examinées par l'utilisation de méthodes de délétion de gènes, car elles induisent l'apoptose, en raison de leur rôle essentiel dans le maintien de l'intégrité mitochondriale. Par conséquent, l'utilisation d'agents ciblant les PHB peut être très utile pour déchiffrer leurs rôles dans divers contextes cellulaires.
1.1.1 Signalisation et rôles physiologiques
Les PHBs sont régulées par plusieurs phosphorylations de la tyrosine et de la sérine, des modifications de O-GlcNAc, des palmitoylations, des transamidations, des nitrosylations de la tyrosine, ... A titre d'exemple, la phosphorylation d'Akt, de PHB1 à Thr258, qui bloque son interaction avec Shp1 / 2 et facilite la signalisation d’Akt (Figure 1). La PHB1 est également phosphorylée par PKC-δ, pour bloquer l'apoptose. L'activité de PHB2 est régulée par la phosphorylation de la serine et de la thréonine, en particulier à Ser91 et Tyr248. La protéine kinase IV dépendant du calcium / calmoduline (CaMKIV) phosphoryle la PHB2 chez Ser91, ce qui
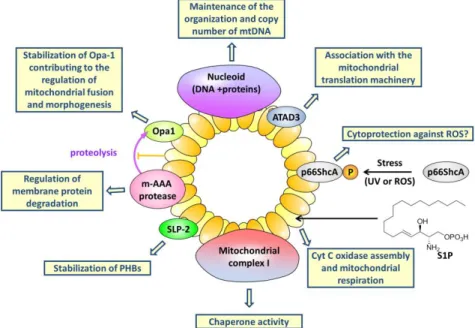
2 Dans la membrane interne mitochondriale, la PHB1 et la PHB2 interagissent enssemble pour former des oligodimères qui sont organisés dans des structures de type anneau avec un diamètre de 20-25nm (Figure 2). Ces supers complexes maintiennent la structure des mitochondries et régulent leurs fonctions.2 Ils agissent aussi dans la régulation du cycle cellulaire, la régulation de la transcription génique
Dans le noyau, les PHB régulent l'activité de nombreux facteurs de transcription, co-activateurs de transcription et co-répresseurs, tels que le p53, le récepteur d'œstrogène, E2Fs, STAT3, PGC. De plus, PHB1 contrôle l'activité des protéines MCM pour réguler la synthèse de l'ADN, Elles protègent les protéines nouvellement importées de la dégradation, par la protéase m-AAA, favorisent la synthèse des protéines mitochondriales, maintiennent l'organisation, copient le nombre d'ADN mitochondrial et agissent comme chaperons pour les protéines nouvellement synthétisées, protéines du complexe mitochondrial I et de l'atrophie optique GTPase 1 (Opa1) lors de la fission mitochondriale et de la morphogenèse.
3 Figure 1. Signalisations des PHB1
(Copiée de la référence Thuaud, F.; Ribeiro, N.; Nebigil, C. G.; Desaubry, L., Prohibitin ligands in cell death and survival: mode of action and therapeutic potential. Chemistry & biology 2013, 20 (3), 316-31).
4 CaMK et PKC) pour orchestrer divers aspects de la physiologie cellulaire, y compris le métabolisme, la transcription, l'apoptose, la survie, la réorganisation du cytosquelette et la différenciation.
Figure 2. Fonction de la PHB1 et de la PHB2 dans les mitochondries.1
(Copiée de la référence Thuaud, F.; Ribeiro, N.; Nebigil, C. G.; Desaubry, L., Prohibitin ligands in cell death and survival: mode of action and therapeutic potential. Chemistry & biology 2013, 20 (3), 316-31).
1.1.2 Importance actuelle des sondes chimiques en biologie cellulaire
Le séquençage du génome humain en 2001 a contribué à l'émergence de la génétique chimique qui vise à identifier des sondes chimiques pour les produits de chaque gène humain.4 En effet, les petites molécules peuvent servir de sonde pour explorer divers processus biologiques.
Les sondes chimiques sont des acteurs majeurs pour valider de nouvelles cibles moléculaires. En biologie cellulaire, elles sont très complémentaires à l'utilisation de l'interférence ARN (ARNi), notamment en étant capables d'inhiber une fonction spécifique de la protéine cible plutôt que d'éliminer la protéine entière, évitant ainsi des problèmes de fonction
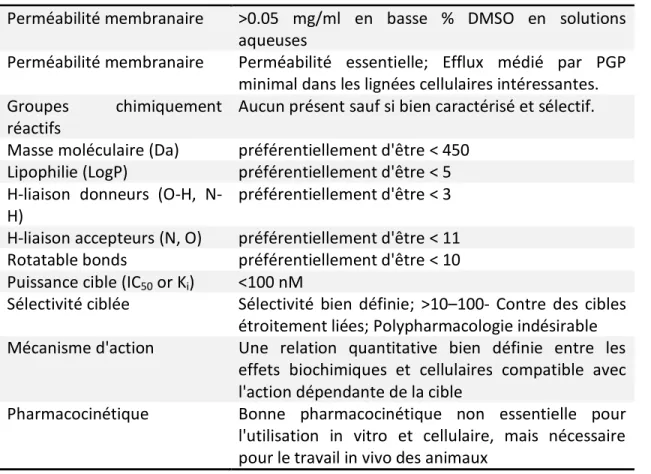
5 Pour être utiles, les sondes chimiques doivent satisfaire à certains critères, tels que la perméabilité (se rendre au site d'action dans la cellule), la puissance et la sélectivité. Paul Workman et Ian Collins ont proposé quelques critères pour évaluer la qualité des sondes chimiques (Tableau 1).3
En ce qui concerne leur puissance, elle devrait être submicromolaire pour les études biochimiques et dans la gamme micromolaire pour les études cellulaires. Pour les études in vivo, elle doit présenter des propriétés pharmacocinétiques acceptables. Fait important, les effets biochimiques et cellulaires doivent être corrélés dans une série d'analogues.
Tableau 1. Directives recommandées pour les propriétés des sondes chimiques.3
Perméabilité membranaire >0.05 mg/ml en basse % DMSO en solutions aqueuses
Perméabilité membranaire Perméabilité essentielle; Efflux médié par PGP minimal dans les lignées cellulaires intéressantes. Groupes chimiquement
réactifs
Aucun présent sauf si bien caractérisé et sélectif. Masse moléculaire (Da) préférentiellement d'être < 450
Lipophilie (LogP) préférentiellement d'être < 5 H-liaison donneurs (O-H,
N-H)
préférentiellement d'être < 3 H-liaison accepteurs (N, O) préférentiellement d'être < 11 Rotatable bonds préférentiellement d'être < 10 Puissance cible (IC50 or Ki) <100 nM
Sélectivité ciblée Sélectivité bien définie; >10–100- Contre des cibles étroitement liées; Polypharmacologie indésirable Mécanisme d'action Une relation quantitative bien définie entre les
effets biochimiques et cellulaires compatible avec l'action dépendante de la cible
Pharmacocinétique Bonne pharmacocinétique non essentielle pour l'utilisation in vitro et cellulaire, mais nécessaire pour le travail in vivo des animaux
6 Les activités des PHB sont affectées par les produits naturels appelés «flavaglines» et de petites molécules entièrement synthétiques telles que l'Aftine-4 ou la mélanogénine.1
1.1.3.1 Les flavaglines
Jusqu'à présent, les ligands de PHB les plus étudiés étaient des flavaglines (Figure 3). Ces composés représentent une famille de plus de 100 cyclopenta [b] benzofuranes trouvés dans l'arbre Chu-lan (Mǐ Zaǐ Lán, 米 仔 兰) utilisés dans la MTC. Les flavaglines présentent de puissantes activités insecticides, antifongiques, anti-inflammatoires, neuroprotectrices, cardioprotectrices et surtout anticancéreuses.6,7
La caractéristique la plus interessante est la sélectivité de leur cytotoxicité vis-à-vis des cellules cancéreuses par rapport aux saines. En effet, il a été observé que lors des cultures in-vitro, toutes les lignées cellulaires cancéreuses et celles transformées sont sensibles à cette cytotoxicité, alors que les cellules non cancéreuses n’ont pas affectées.
Figure 3. Exemples représentatifs de flavaglines naturelles (rocaglamide et silvestrol) et synthétiques (FL3).
7 les PHB d'interagir avec la kinase C-RAF et inhibent par conséquent la signalisation Ras-C-RAF-MEK-Erk, qui est essentielle à la survie de nombreux types de cellules cancéreuses.9
Activités anticancéreuses
De nombreux dérivés de flavaglines ont été synthétisés pour déterminer les exigences structurales de leur cytotoxicité dans les cellules cancéreuses (Figure 4) .1,10
Figure 4. Requis structuraux des flavaglines pour la cytotoxicité dans les cellules cancéreuses (FL3: R1=R2=OMe, R3=OH, R4= R5= R6= R7= R9=H, R8=Br).
Tableau 2. Cytotoxicité dans les cellules HCT15 (IC50, nM).
Anticancer agents IC50 (nM) FL3 2 Doxorubicine 81 Docetaxel 13 Cisplatin 2900 Camptothecine 3 Vinblastine 10
8 cytotoxicité supérieure à celle des flavaglines naturelles (Figure 4, Tableau 2). Une flavagline a été conjuguée à un lieur est utilisé pour identifier les prohibitines qui sont leur cible et ce par chromatographie d'affinité. 9
En 2005, les chercheurs de Bayer Healthcare ont rapporté que les flavaglines présentaient des activités anti-inflammatoires et neuroprotectrices puissantes dans des modèles animaux de la maladie de Parkinson.11
Sur la base de cette observation, il a été démontré que ces composés protègent les neurones et les cardiomyocytes contre l'apoptose induite par les médicaments anticancéreux (cisplatine et doxorubicine), et qu’en particulier le FL3 protège les souris in vivo de la toxicité induite par la doxorubicine. Ces données indiquent également que le FL3 présente une biodisponibilité appropriée. Aucun signe de toxicité ou de perte de poids chez des souris traitées uniquement avec FL3 n’a été détecté (Figure 5).
Figure 5. Taux de survie des souris traitées avec la doxorubicine(31%) ou doxorubicine + 5 injections de FL3 (0,1 mg / kg i.p.).12
Il a été également montré que le FL3 induit des propriétés cardioprotectrices via la signalisation par la PHB1 (Figure 6).13 En effet, la doxorubicine induit la translocation de PHB1 et de STAT3 phosphorylatée dans le noyau des cardiomyocytes pour réaliser l'apoptose. Au contraire, FL3 induit la translocation de ces protéines de signalisation dans les mitochondries pour protéger la cellule contre les effets indésirables de la doxorubicine.
9 Figure 6. Mécanisme cardioprotecteur de FL3 proposé par le Dr Désaubry et al.
Cette activation du complexe STAT3 / PHB1 mitochondrial par des ligands de PHB représente une nouvelle stratégie pour protéger l'organisme contre la cardiotoxicité induite par la doxorubicine et éventuellement d'autres problèmes cardiaques.
Les relations structure-activité de la cardioprotection sont identiques à celles de la neuroprotection et similaires pour la cytotoxicité dans les cellules cancéreuses (Figure 7).14
Figure 7. Exigences structurelles pour la cardioprotection et la cytotoxicité des cellules cancéreuses.
En plus des flavaglines, plusieurs ligands PHB ont été identifiés, tels que la mélanogénine, l’Aftine-4, l’aurilide, l’adipotide, la Fluorizoline et les sulfonylamidines.
10 Figure 8. Structure et neurotoxicité de l’Aftine-4
Lors de l’étude qui déterminait comment le peptide amyloïde-b42 (Ab42) toxique est produit dans la maladie d'Alzheimer, le Professeur Hervé Galons, a observé que le dérivé d'adénine Aftine-4 interagit avec trois protéines: la PHB1, le canal VDAC1 et la mitofiline.15 Au niveau cellulaire, il favorise la production de neurotosines dans les neurones.
1.1.3.3 La fluorizoline
La fluorizoline est un nouvel agent anticancéreux qui cible les PHBs.16 Les requis structuraux de ce composé pour son activité cytotoxique demeurent peu décrits.17
11 1.2.1 Mélanine et mélanogenèse
La mélanine est le pigment noir qui protège la peau des radiations ultraviolettes, fournit les différentes décorations de la peau, des yeux et des fourrures d’animaux.
La couleur de la peau, des yeux et des cheveux sont en grande partie le résultat de la synthèse de la mélanine par les mélanocytes dans une organelle spécialisée appelée le mélanosome.18 Les mélanocytes responsables de la pigmentation protègent des dommages induits par les UV sur la peau.19 Un grand nombre de maladies de la peau, qui comprennent l’hyperpigmentation acquise, tels que le melasma, la mélanodermie postinflammatoire et lentigos solaires, sont dûs à l’augmentation de la production et de l'accumulation de la mélanine.20 A l’inverse, l’albinisme ou le vitiligo résultent d’un défaut de pigmentation.
Les agents éclaircissant sont devenus de plus en plus importants dans le domaine cosmétique. Plus de 100 gènes sont impliqués dans le processus de la mélanogénèse.21 Les
altérations dans la transcription, la traduction, le traitement ou le trafic intracellulaire d'une de ces protéines peut affecter la synthèse de la mélanine. Les mélanocytes répondent aussi à divers facteurs produits par leur environnement ou par les cellules voisines de la peau.22,23
Le transfert des mélanosomes des mélanocytes aux kératinocytes et leur répartition dans les kératinocytes sont également des procédés qui jouent des rôles importants dans la régulation de la pigmentation de la peau et des cheveux.24,25
12 Figure 10. La mélanogenèse ou la synthèse de la mélanine est impliquée dans des voies réactionnelles enzymatiques et biochimiques complexes appelées voie de Raper-Mason5,27
Cette figure a été adaptée d’un article de T.Pillayiyar et al légèrement modifié. (Tyr, tyrosinase, DQ, dopaquinone, L-Dopa, L-3,4-dihydroxyphénylalanine, DHICA, acide 5,6-dihydroxyindole-2-carboxylique;DHI, 5,6-dihydroxyindole; ICAQ, acide indole-2-carboxylique-5,6-quinone; IQ, indole-5,6-quinone; HBTA, 5-hydroxy-1,4-benzothiazinylalanine).26
13 Figure 11. Principales voies régulatrices de la mélanogenèse, cette figure a été adaptée de Pillaiyar et al et légèrement modifiée.26
La régulation de la mélanogenèse dans les mélanocytes est un phénomène complexe régulé par de nombreuses voies de signalisation, notamment celles de l’AMP cyclique, du GMP cyclique de la β-caténine et des MAPS kinases. Ces voies convergent vers l’activation du MITF (microphthalmia-associated transcription factor) qui active la transcription d’enzymes-clés de la mélanogenèse, et en particulier la tyrosinase.26
14 Parmi les différentes protéines régulant la mélanogénèse, nous allons présenter en particulier celles qui sont affectées par les composés que nous avons synthétisés : le MITF, pERK et LC3.
1.2.2.1 MITF : Microphthalmia-associated transcription factor
Le MITF est un facteur de transcription qui contrôle la mélanogenèse. Cette protéine joue un rôle dans le développement, la survie et la fonction de certains types de cellules.27, 28
Le MITF contrôle le développement et la fonction des mélanocytes. Au sein de ces cellules, cette protéine contrôle la production du pigment mélanine. Les mélanocytes sont également présents dans l'oreille interne et jouent un rôle important dans l'audition. De plus, le MITF régule le développement des cellules épithéliales du pigment rétinien. Ces cellules nourrissent la rétine.
Le MITF régule également le développement des cellules qui décomposent et éliminent les os (ostéoclastes) et les cellules qui jouent un rôle dans les réactions allergiques (mastocytes).29
Le MITF régule l'expression du récepteur de Kit29 ainsi que celle des enzymes telle que la tyrosinase et la protéine 1 liée à la tyrosinase (TRP1),30 qui sont essentiels pour la production de la mélanine. La tyrosinase est l'enzyme limitante de la biosynthèse de la mélanine, catalysant la conversion de la tyrosine en 3,4-dihydroxyphenylalanine (DOPA) et ensuite en DOPA quinone.31 La TRP1 catalyse la conversion de l’acide 5,6-quinonecarboxyindole-2-carboxylique (DHICA) en l’acide indole-5,6-quinonecarboxylique.32
15 kinases activées par des phosphorylations au niveau d’une tyrosine et d’une thréonine.
ERK1 et ERK2 phosphorylent aussi des protéines sur des résidus tyrosine, thréonine ou sérine pour activer à leur tour d’autres protéines. ERK1/2 font partie de la famille des MAP kinases, mitogen-activated protein kinases.33
L’ extracellulaire signal-regulated kinase (ERK) est une clé médiatrice de signalisation de la prolifération et de la différenciation cellulaire,34,35 et l'inhibition de la voie ERK induit une hyperpigmentation, suggérant ainsi que ERK est également impliqué dans la mélanogenèse.36
En outre, l'activation d'ERK conduit à la phosphorylation de MITF à la serine 73, qui sera suivie par l'ubiquitination de MITF et sa dégradation.37 L’activation d’ERK par le sphingosine-1-phosphate ou la céramide régule la mélanogenèse.38
L’activation de ERK a également un effet inhibiteur sur l'activité de la tyrosinase et la transcription.39,40
1.2.2.3 LC3 : généralités et relations avec la mélanogenèse, la PHB2 et ER
La chaîne légère 3β associée aux microtubules (ci-après dénommées LC3) est une protéine qui chez les humains est codée par le gène MAP1LC3B.
LC3 est une protéine centrale dans la voie d'autophagie où elle fonctionne dans la sélection de substrat d’autophagie et la biogenèse d’autophagosomes. LC3 est le marqueur des autophagosomes le plus largement utilisé.24
La LC3 a été identifiée à l'origine comme une protéine associée aux microtubules dans le cerveau du rat.41
Cependant, il a été constaté plus tard que la fonction primaire de LC3 est dans l'autophagie, un processus qui implique la dégradation en bloc des composants cytoplasmiques.
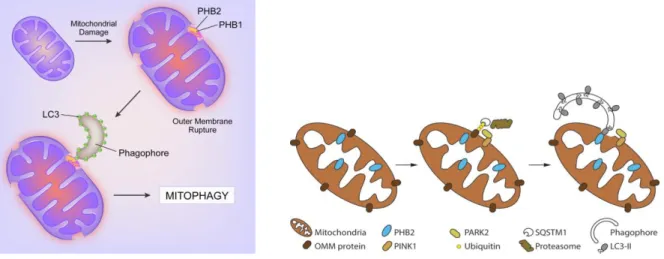
16 d'ERK, Beth Levine et al ont récemment montré que la PHB2 interagit physiquement avec la chaîne légère 3β (LC3) de la protéine 1 associée aux microtubules pour promouvoir la mitophagie en réponse au dysfonctionnement mitochondrial des cellules HeLa, et que pendant la mitophagie, la protéine associée à l'autophagosome LC3-II se lie directement à la protéine mitochondriale interne PHB2 et se lie indirectement à l’hétérodimère PHB2 et son partenaire de liaison PHB.43
Figure 12. Mode d’action de LC3 lié à la PHB2 lors de la mitophagie. 43,44
L'interaction entre PHB2 et LC3 nécessite une rupture de la membrane mitochondriale externe. PHB2 est requis pour la mitophagie à médiation par Parkin.44 Figure copiée de Wei, Y.; Chiang, W.-C.; Sumpter, R.; Mishra, P.; Levine, B., Prohibitin 2 is an inner mitochondrial membrane mitophagy receptor. Cell 2017,
168 (1), 224-238. e10. Et de Bang, S.; Won, K. H.; Moon, H. R.; Yoo, H.; Hong, A.; Song, Y.; Chang, S. E.,
Novel regulation of melanogenesis by adiponectin via the AMPK/CRTC pathway. Pigment cell &
melanoma research 2017, 30 (6), 553-557.
Les mitochondries dépolarisées expriment PINK1 sur la membrane externe, entraînant le recrutement de PARK2 et l'ubiquitination et la dégradation subséquente des protéines de la membrane externe mitochondriale. La rupture de la membrane externe qui en résulte permet à LC3-II sur un phagophore de se lier à la protéine membranaire interne mitochondriale PHB2, conduisant à la mitophagie.43
17 induit la formation de mélanine en inhibant la translocation de la prohibitine-1 (PHB1) des mitochondries vers le noyau des mélanocytes.14 Il a été proposé que la PHB1 bloque la transcription d’un facteur contrôlant la transcription de la tyrosinase. L’enzyme qui catalyse l’étape limitante de la mélanogénèse est que la mélanogénine en se liant à la PHB1 lèverait cette inhibition (Figure 14).
Figure 13. Structure de la mélanogénine.
Figure 14. Mécanisme proposé de l’action de la mélanogénine.1
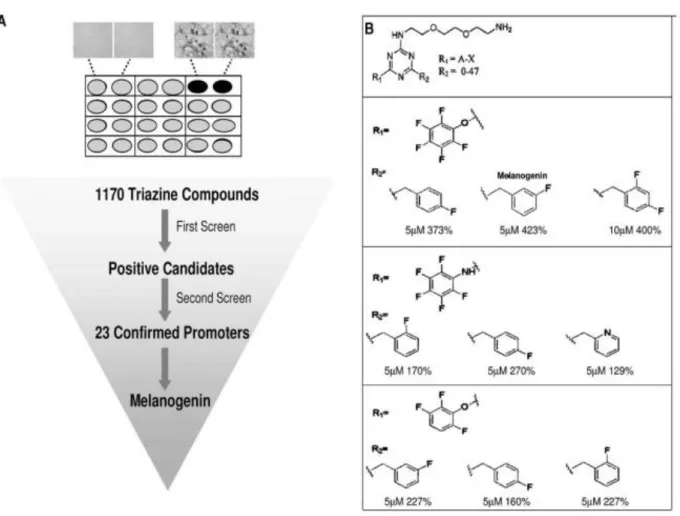
Pour identifier des composés qui modulent la pigmentation de la peau, Snyder et al ont criblé une chimiotèque constituée de 1170 triazines sur des mélanocytes non pigmentés.14 Ils ont identifié la mélanogénine comme inducteur de pigmentation avec une EC50 de 2,5 mM. Ce composé augmente la tyrosinase, l'enzyme limitant la vitesse dans la biosynthèse de la mélanine. Ces auteurs ont conjugué la mélanogénine à un support d'agarose, ce qui leur a
18 recherches biologiques ont confirmé que la PHB1 est responsable de l'induction de la pigmentation. Les études de microscopie ont montré que la PHB1 était localisée uniquement dans les mitochondries, ce qui a conduit ces auteurs à suggérer que la liaison de la mélanogénine au PHB1 pourrait perturber l'interaction entre PHB1 et un facteur de transcription, provoquant ainsi sa translocation dans le noyau et l'induction de l'expression de la tyrosinase, l'enzyme limitant le taux de mélanogenèse. Cette étude a été la première à démêler l'implication de la PHB1 dans la régulation de la pigmentation des mammifères.
19 structurelles ont été produites en modifiant la structure chimique de la triazine aux positions R1 et R2. L'activité chimique est exprimée par le pourcentage d'augmentation de la pigmentation au-dessus des mélanocytes non traités.14
1.2.4 Relations structure-activité
Identification de la Prohibitine comme Protéine cible de la Mélanogénine
Pour déterminer si la prohibitine a un rôle fonctionnel dans l'induction de pigmentation, la technologie ARNi a été utilisée pour réduire l'expression de la prohibitingene de manière sélective (Figure 9c). Ils ont démontré une atténuation significative de la réponse cellulaire à la mélanogenine suivant l'inactivation du gène de la prohibitine spécifique (Figure 16e).
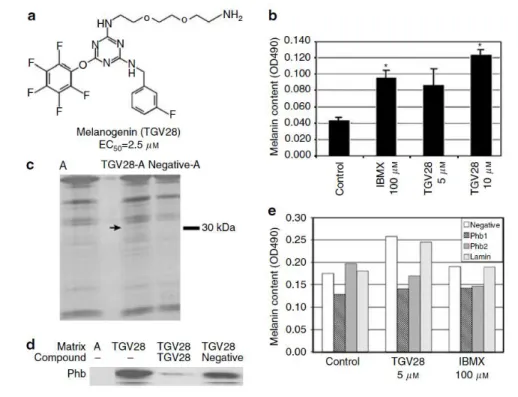
Figure 16. Identification d'un agent améliorant la pigmentation qui se lie à la prohibitine.45 (Copiée de Ni-Komatsu, L.; Orlow, S. J., Identification of novel pigmentation modulators by chemical genetic screening. The Journal of investigative dermatology 2007, 127 (7), 1585-92).
20 mélanocytes murins immortalisés (mélan-a cells). La teneur en mélanine est normalisée par rapport à la quantité totale de protéines dans les lysats cellulaires.
c) La mélanogénine se lie spécifiquement à la prohibitine. Le gel SDS-PAGE coloré à l'argent à une concentration de 5-15% illustre la spécificité de la liaison mélanogénine-protéine. A est la matrice de billes d'agarose non conjuguée, TGV28-A est la matrice d'affinité de la mélanogénine, et négatif-A est la matrice d'affinité conjuguée à témoin négatif. La flèche indique la bande protéique qui se lie spécifiquement à la matrice conjuguée à la mélanogénine.
d) Le Western blot du lysat de mélanocyte incubé avec des billes d'agarose conjuguées à la mélanogénine (TGV28) ou des billes d'agarose seules utilisant un anticorps contre la PHB (A). La liaison mélanogénine-prohibitine a été supprimée lors de la pré-incubation du lysat cellulaire avec la mélanogénine libre mais pas avec le composé témoin inactif (négatif).
e) L’inhibition du gène de la prohibitine atténue efficacement la réponse cellulaire à la mélanogénine. Dosage de la mélanine des cellules de mélanome B16-F10 transfecté avec deux séquences d'siRNA interditines distinctes (PHB1 et PHB2), siRNA contrôle négatif, ou siRNA Lamin A / C non pertinent suivi par aucun traitement (témoin), mélanogénine 5 mM, ou 100 mM traitement à l'isobutylméthylxanthine.45
Relation structure-activité de la mélanogénine et de ses analogues
Orlow et al. ont déposé un brevet couvrant la structure de plusieurs dizaines de triazines. Ces données nous fournissent des premières informations sur le développement de nouveaux analogues de la mélanogénine.46
Tableau 3. Relation structure-activité de la mélanogénine et de ses analogues
Entrées Structures des analogues de la mélanogénine Concentrations (µM) % d’induction de la pigmentation comparé aux cellules non traitées 1 5 423
21 2 5 373 3 5 537 4 10 400 5 10 3 360 6 10 360 7 10 353 8 5 270
22 9 5 227 10 1 167 11 5 72 12 5 109 13 5 103 14 5 340 15 5 270
23 16 5 227 17 10 102 18 N N N O HN O O H N N H F F F F F F O 5 493 19 5 182 20 5 197
24 21 5 537 22 5 277 23 5 170 24 10 87 25 5 160 26 5 537
25 27 10 400 28 10 3 360 29 10 360 30 10 353 31 5 270 32 5 227 33 1 167
26 34 5 72 35 5 109 36 5 103 37 5 340 38 5 270 39 5 227
27 40 10 102 41 5 493 42 5 182 43 5 197 44 5 537
28 Le remplacement du bout de la chaine dans la mélanogénine par d’autres substituants augmente considérablement leurs activités.
Pour le R1 : les composés fluorés sont ceux qui augmentent l’activité des analogues.
Pour le R2 : le pentafluorophénoxy est celui qui apporte la meilleure activité. On remarque que plus il y a du fluor plus c’est actif, et que l’oxygène est plus actif que l’azote et doit être directement lié au cycle aromatique fluoré.
29 substitution séquentielle de la trichlorotriazine 1 (Schéma 1). La première substitution se produit à basse température alors que les deuxième et troisième réactions nécessitent des températures plus élevées.
Schéma 1. Voie de synthèse générale des triamino-triazines.47
La mélanogénine a été préparée par Y-T. Chang et al. selon l’approche ci-dessus.14 Les détails de cette synthèse ne figurent ni dans l’article décrivant ce composé, ni dans le brevet correspondant.46 En effet, les triazines décrites dans cette étude ont été préparées par une synthèse sur surface solide où le groupement espaceur est conjugué par amination réductrice à des résines fonctionalisées par du di-méthoxybenzaldéhyde (Schéma 2).
30 Schéma 2. Synthèse supportée des triazines fonctionnalisées développée par Chang et al.46
Une première amine est condensée à la trichlorotriazine pour former un premier intermédiaire C, qui est ensuite combiné à la résine B pour produire la triazine trisubstituée D. Cette approche a permis de préparer des chimiothèques comportant des milliers de triazines substituées par deux amines différentes et par un groupement espaceur avec des quantités d'environ 1 à 2 mg. C’est par cette approche décrite par Chang et al. qu’a été synthétisé la chimiothèque incluant la mélanogénine, cependant ni les conditions expérimentales, ni les rendements des différentes étapes, n’ont été décrites dans le cas spécifique de la mélanogénine. Cette méthode a montré son efficacité pour préparer des triaminotriazines. En revanche, elle ne peut pas être efficacement appliquée à la synthèse de diaminotriazines substituées par un groupement pentafluorophénoxy. En effet la pentafluorophénol n’est pas suffisamment nucléophile pour réagir avec les triazines immobilisées D ou E.46
T. Masquelin et al. ainsi que J. T. Brock et al en s’associant avec Y-T. Chang ont également développé une synthèse de triazines sur surface solide qui présente comme particularité de faire appel à une amino-chlorotriazine reliée à une résine par un thioéther comme intermédiaire clé (Schéma 3). Après une première addition d’amine, le thioéther est
31 Schéma 3. Synthèse des triazines fonctionnalisées développée par Masquelin et al.48
32 préparer des analogues de la mélanogénine qui sont des composés originaux ayant potentiellement de meilleures activités biologiques, puis d’examiner les requis structuraux pour leurs activités promélanogéniques.
33
3.1 Mise au point d'une synthèse sélective d'analogues de la mélanogénine
La première tâche de mon projet fût de mettre au point une synthèse pratique et efficace de la mélanogénine et de ses analogues. Une première condensation à -20°C de la benzylamine 2 sur la trichlorotriazine 1 a donné le monoadduit 3 avec un rendement de 75 %. La condensation ultérieure avec le pentafluorophénol 4 à 0°C conduit à un mélange de rendement global de 72% dont les produits finaux sont le monoadduit 5 et le diadduit 6 dans un rapport respectif de 74 :26. Cette réaction n’étant pas sélective, nous avons cherché à examiner une autre stratégie (Schéma 4).
Schéma 4. Condensation du pentafluorophénol 4 sur la dichlorotriazine 3.
Notre deuxième démarche a été inspirée des travaux de T. Masquelin et al,48 où la
trichlorotriazine 1 est mise en réaction avec du benzylthiol 7 en présence de DIPEA pour donner un mélange de rendement 64% composé de monoadduit 8 et de di-adduit 9 dans un rapport 87:13 (Schéma 5).
Schéma 5. Double substitution nucléophile d’une benzylthiol sur la trichlorotriazine Selon les travaux de T. Masquelin et al.
34 sur la trichlorotriazine, suivie d’une 3ème substitution par du pentafluorophénol et d’une oxydation des thioéthers en sulfones ou sulfoxides pour faciliter leurs déplacements décrite par A. Vallribera et al. J’ai essayé deux oxydants (oxone et mCPBA) dans de multiples conditions (Schéma 6, Tableau 4).
Schéma 6. Essai de développement d’une méthode de synthèse de triazines substituées par un groupement pentafluorophenoxy faisant appel à des intermédiaires thioétehers et sulfoxydes.
35 a Oxone(5 éq)/THF/H2O ; 0°C→RT Principalement produit de départ.
b Oxone (5 éq)/THF/H2O (quantité de
solvant doublée) ; 0°C→RT
Principalement produit de départ
c Oxone (5 éq)/Acétone/H2O: 4/1; RT Mélange non séparable 13 (n=1 ou 2) +
monosulfoxyde 14 dans un ratio 80%/20%.
d Oxone (6 éq)/DMF, 0°C →RT Mélange de disulfoxyde/sulfone, sulfoxyde dans un ratio de 95%/5%.
e mCPBA (2.1 éq)/CH2Cl2 ;0°C→RT Mélange de disulfoxyde/disulfoxyde.dans
un ratio de : 75%/25%
Le traitement des produits issus des conditions c et d par des amines a donné des mélanges, ce qui nous a incité à mettre en place une nouvelle approche. Lors de nos expériences nous avons constaté que le groupement pentafluorophénoxy se déplace aussi facilement que les groupements sulfones ou sulfoxydes.
36 Schéma 7. Troisième approche pour la mise au point de méthode de synthèse de la
mélanogenine et de ses analogues
Contrairement à la substitution par la plupart des nucléophiles, le remplacement d’un chlorure par un groupement pentafluorophénoxy ne perturbe guère le caractère électrophile des triazines, ce qui fait que les SNAr faisant intervenir le pentafluorophénol conduisent à un mélange de mono- et di-adduits. Nous avons donc cherché à développer une méthode pour nous affranchir de ce manque de sélectivité.
Notre nouvelle stratégie s’est basée sur les travaux de Katharina Reichenbacher et al. qui a décrit une triple addition du pentafluorophénol sur la trichlorotriazine.50
Schéma 8. Travaux de Katharina Reichenbacher et al.50
Nous nous sommes inspirés de ces travaux et avons effectué dans un premier temps une di substitution nucléophile de la tricholorotriazine avec le pentafluorophénol avec 90% de rendement.
37 permet d’obtenir les produits finaux désirés. La structure des composés synthétisés avec les rendements de la dernière étape sont indiqués dans les tableaux 5-9.
Schéma 9. Approche finale permettant d’accéder à la mélanogénine et à ses dérivés.
Certaines amines utilisées dans la 3ème étape du schéma 17 n’étaient pas disponibles, ce qui nous a ramené à les synthétiser. Ainsi L’amine Y a été préparée par la méthode de Ghosh
et al. par amination réductrice de la cétone X en utilisant du cyanoborohydrure de sodium.51
38
Schéma 10. Synthèse des amines Y et Z non-commerciales.
39 Figure 17. Les analogues 16a
Tableau 5. Les analogues 16a synthétisés
NR1R2 Rdt NR1R2 Rdt
Mel 1 NH2 57% Mel 14 81%
Mel 2 NHMe 52% Mel 15 60%
Mel 3 NMe2 99% Mel 16 57%
Mel 4 32% Mel 17 99%
Mel 5 82% Mel 18 88%
Mel 6 99% Mel 19 27%
40 Mel 9 65% Mel 22 89% Mel 10 41% Mel 23 99% Mel11 62% Mel 24 94% Mel 12 96% Mel 25 94% Mel 13 55% Mel26 95%
41 Figure 18. Les analogues 16b.
Tableau 6. Les analogues 16b synthétisés.
NR3R4 Rdt NR3R4 Rdt Mel27 81% Mel33 91% Mel 28 84% Mel 34 98% Mel 29 81% MEL 49 80% Mel 30 34% Mel 50 99% Mel 31 99% Mel 51 81% Mel 32 83%
42 Figure 19. Les analogues 16c.
Tableau 7. Les analogues 16c synthètisés.
NR3R4 Rdt NR3R4 Rdt Mel35 99% Mel42 47% Mel37 91% Mel43 83% Mel38 95% Mel52 98% Mel39 13% Mel53 90% Mel40 65% Mel54 97% Mel41 86%
43 Figure 20. Les analogues 16d
Tableau 8. Les analogues 16d synthétisés.
NR3R4 Rdt NR3R4 Rdt
Mel 36 90% Mel 46 50%
Mel 44 92% Mel 47 92%
44 Figure 21. Les analogues 16e.
Tableau 9. Les analogues 16e synthètisés.
NR3R4 Rdt NR3R4 Rdt NR3R4 Rdt
45 en utilisant la diamine monoprotégée A pour obtenir l’adduit A’ qui a été déprotégé ensuite par hydrogénation. Il s’agit là de la première synthèse efficace de la mélanogénine.
46 Pour les tests biologiques notamment celles du pull down, (visant à prouver que les analogues de la mélanogénine synthétisés se lient aux prohibitines), nous avons dû conjuguer le Mel 41 à un groupement espaceur (linker) susceptible d’être immobilisé sur l’affigel10 (agarose activée).
La synthèse de ce bras espaceur s’est faite à partir d’une animation réductrice, en faisant réagir l’anime A et la N-Boc- piperidin-4-one B en utilisant le triacetoxyborohydride de sodium comme agent réducteur à 0°C avec un rendement de 82%, suivi d’une déprotection du groupement Boc par l’acide trifluoroacétique pour le coupler avec l’analogue 16e obtenu avec un rendement de 88%, puis le déprotéger du groupement Cbz par hydrogénation suivie d’un couplage avec l’Affigel10.
47
4.1 Stimulation de la pigmentation cellulaire
L’équipe du professeur Ghanem à l’institut Jules Bordet (Bruxelles) a examiné si ou non nos nouveaux analogues de la mélanogénine favorisent la mélanogénèse dans les cellules de mélanome HBL en mesurant l'ampleur de la formation du pigment (Figure 22, Tableau 10).52 Tableau10. Pourcentage d'induction de la mélanogenèse dans la lignée de cellules de mélanome humain HBL. NR1R2 NR3R4 Induction de la mélanogènesea 1 μM 10 μM Vehicule 1.00 1.00 Fluorizoline 1.51 1.37 Mélano- génine 1.05 1.25 Mel 1 NH2 0.92 0.91 Mel 2 NHMe 0.89 0.84 Mel 3 NMe2 1.04 1.05 Mel 4 0.99 1.08 Mel 5 0.85 0.93
48 Mel 7 0.70 0.89 Mel 9 1.15 2.01 Mel 10 0.99 1.04 Mel 11 0.91 1.00 Mel 12 0.89 0.84 Mel 13 0.80 0.76 Mel 14 1.06 1.39 Me l15 1.09 1.25 Mel 16 1.07 0.95 Mel 17 1.02 1.01 Mel18 0.92 0.94
49 Mel 20 0.97 1.09 Mel 21 0.89 1.10 Mel 22 0.90 1.11 Mel 23 1.07 1.07 Mel 24 0.90 1.00 Mel 26 0.80 0.61 Mel 27 NHMe 0.90 0.69 Mel 28 NHMe 1.16 0.89 Mel 31 NHMe 0.93 0.98 Mel 32 NHMe 0.84 0.87 Mel 33 NHMe 0.80 0.86 Mel 34 NHMe 0.84 0.78
50 Mel 36 0.77 0.81 Mel 38 0.77 0.43 Mel 39 0.86 0.55 Mel 40 0.78 0.61 Mel 41 0.89 2.23 Mel 42 1.15 1.97 Mel 43 0.94 0.71 Mel 44 0.78 0.68 Mel 45 0.95 0.74
51 Mel47 0.94 0.83 Mel48 0.96 0.89 Mel49 NHMe 1.01 0.84 Mel50 NHMe 0.88 0.94 Mel51 NHMe 1.01 0.86 Mel52 1.25 1.53 Mel53 0.98 0.99 Mel54 1.11 1.43 Mel55 0.87 1.79 Mel56 0.83 1.51
52 Contrairement aux cellules (mom) utilisées à l'origine pour découvrir la mélanogénine, les cellules HBL sont moins sensibles aux signaux d’induction de la mélanogenèse, ce qui permet de distinguer les composés puissants des composés peu actifs et d'éviter les faux positifs. Dans les cellules HBL, la mélanogénine a stimulé la formation de pigment 1,25 fois par rapport au véhicule, alors que la plupart des composés criblés n'ont pas activé la mélanogénèse. Les agents les plus actifs comprenaient une fraction N,N-diméthyl-éthylènediamine, avec une forte préférence pour une tétrahydrobenzazépine (Mel41). Ce composé ayant augmenté de 2,23 fois la pigmentation (Figure 22A).
53 Figure 22. Effet sur la pigmentation cellulaire des analogues de la mélanogénine.
54 B) Sructures des composés actifs représentatifs.
C) Résumé des requis structuraux pour l'activité promélanogénique.
Fait intéressant, un autre ligand du PHB, la fluorizoline,53,54 favorise significativement la mélanogénèse dans les cellules HBL, confirmant que cette induction de la pigmentation est en effet médiée par une action sur les PHB. Tous les composés actifs ont été substitués par un substituant diaminé, avec une légère préférence pour la N,N-diméthyl-éthylènediamine sur la pipéridinopipéridine (Mel55), la 4-diméthylaminopipéridine (Mel56) ou la
N,N-diméthylpropylènediamine (Mel57). L'induction de la mélanogenèse a également été favorisée par la N-méthylation, la N-éthylation et la C-méthylation en position benzylique avec une configuration R, ou par l'introduction d'un fluor en méta sur le phényle.
Pour examiner si les effets promélanogéniques sont spécifiques à ce type cellulaire, l'activité de Mel9 et Mel41 dans deux autres lignées cellulaires a été testée. Le Mel9 et le Mel41 induisaient significativement la mélanogenèse également dans les cellules de mélanome MM043 et MM162 ce qui est en accord avec les résultats précédents (Figure 23).
Figure 23. Effet de Mel9 et Mel 41 sur la pigmentation cellulaire dans les lignées cellulaires MM162 et MM043.
55
4.2 Le Mel55 interagit avec PHB1
Pour vérifier que les nouveaux dérivés de la mélanogénine se lient bien à la PHB1, le Mel55 a été conjugué à des billes d'Affigel-10 à travers un bras éspaceur et un pull-down a été effectué en utilisant des lysats de cellules entières issus des cellules de mélanome HBL (Figure 24). Le Mel55 a été choisi parce qu'il combine les caractéristiques structurales de Mel9 et Mel41 et est par conséquent un représentant typique de cette classe de composés. L’Affigel-10 conjugué à Mel55 a pu se lier à la PHB1, ce que le témoin Affigel-10 n’a pas pu faire.
Figure 24: Mel55 interagit avec PHB1.
A) Structure de l'Affigel-10 conjugué à Mel55 ou au linker uniquement (témoin négatif, NC).
B) Analyse par transfert de Western de protéines arrachées à partir des billes Affigel-10 couplées au Mel55 ou au négatif, en utilisant un lysat de cellules de mélanome HBL.
4.3 Le Mel9 et le Mel41 régulent à la hausse le MITF et la tyrosinase grâce à l'activation de LC3
Ayant identifié des sondes avec des activités promélanogéniques améliorées, les effets de Mel9 et Mel41 ont été ensuite éxaminé sur les protéines clés de signalisation qui régulent la mélanogenèse dans les cellules du mélanome HBL (Figure 25). Le facteur de transcription
56 particulier, MITF augmente la transcription de la tyrosinase, l'enzyme limitant la vitesse dans la biosynthèse de la mélanine. Ainsi, nous avons examiné le Mel9 et le Mel41 pour la régulation du MITF et l'expression de la tyrosinase. Conformément à la stimulation de la mélanogenèse, l'expression du MITF et de la tyrosinase a augmenté considérablement lors du traitement avec le Mel9 et le Mel41. L'augmentation de l'expression de MITF était plus faible à 24h qu’à 48h, mais l'expression de la tyrosinase était légèrement plus élevée après 24 heures de traitement.
La chaîne légère 3β de la protéine 1 associée aux microtubules (LC3) est impliquée à la fois dans la régulation du MITF et dans la mitophagie.42,43 En effet, lors de l'induction de la mélanogenèse, la forme cytosolique de LC3 (LC3-I) est conjuguée à la phosphatidyléthanolamine pour former le conjugué LC3-phosphatidyléthanolamine (LC3-II), ce qui favorise l'expression du MITF.42 De plus, PHB2 interagit avec LC3-II pour induire la dégradation des mitochondries endommagées.43 Ainsi, nous avons supposé que le Mel9 et le Mel41 pourraient induire une mélanogenèse par activation de LC3 ce qui est effectivement le cas. En conséquence, le Mel9 et le Mel41 ont induit une accumulation de LC3-II d'une manière dépendante de la dose, indiquant que le Mel9 et le Mel41 agissent sur les PHB pour promouvoir la mélanogenèse par la voie LC3-II-MITF. Cet effet était moins prononcé après 48h qu’après 24h.
57 Figure 25. Activation de MITF via l’activation de LC3-II par les ligands PHB le Mel9 et le Mel41. A) Les Western Blots représentatifs illustrant l'effet du Mel9 et du Mel41 (0, 0,1, 1, 10 μM) pendant 24 et 48 heures dans des cellules HBL. Des quantités égales de protéines extraites (35 μg) ont été analysées par la méthode de transfert de Western avec des anticorps contre le MITF, la tyrosinase, le LC3 et le phospho-ERK.
B) Les analyses densitométriques des bandes immunoréactives. Les niveaux de phosphorylation de ERK ont été corrigés avec l'expression de la protéine totale ERK, les taux d'expression de MITF, de tyrosinase et de LC3 ont été corrigés avec l'expression de la ß-actine. Les résultats représentent le moyen de deux expériences indépendantes.
Pour corroborer l'effet de nos ligands PHB sur l'activation de LC3, nous avons examiné si l'activité promélanogénique peut être modulée par un inhibiteur (PIK-III) ou un activateur (HD1) de l'autophagie et l'activation de LC3. Comme prévu, le PIK-III a effectivement inhibé l'activité promélanogénique du Mel9, tandis que HD1 a amélioré cette activité (Figure 26A). En conséquence, PIK-III a réprimé l'expression de la conversion MITF et LC3 induite par Mel9, tandis que HD1 a favorisé ces deux événements (Figures 26B et 26C).
58 Figure 26. Inhibition et amélioration de l’activité promélanogénique du Mel9 respectivement par l’inhibiteur (PIK-III), l’activateur de l'autophagie(HD1) et l'activation de LC3.
A) Modulation de l'activité promélanogénique du Mel9 par PIK-III et HD1. Les cellules HBL ont été traitées par du Mel9 (10 μM) pendant dix jours avec et sans le PIK-III (10 μM) et le HD1 (10 μM), la teneur en mélanine exprimée par ces cellules est relativement supérieure à celle à celle traitées avec le véhicule.
B) Modulation de l'expression MITF induite par du Mel9 et l'activation du LC3 par le PIK-III et le HD1. Les cellules HBL ont été traitées par du Mel9 (10 μM) pendant 24 heures avec et sans le PIK-III (10 μM) et le HD1 (10 μM). Des quantités égales de protéines extraites (35 μg) ont été analysées par Western blot avec des anticorps contre le MITF, le LC3 et la ß-actine. Le PIK-III bloque l'activation de LC3 et l'autophagie en inhibant l'activité enzymatique de VPS34. L’HD1 a l'effet opposé en activant l'AMPK.
En plus du LC3-II, le MITF est également régulée par d'autres facteurs, y compris la kinase ERK. Plus spécifiquement, le ERK phosphoryle le MITF pour favoriser simultanément son activité transcriptionnelle et sa dégradation dépendante de l'ubiquitine comme mécanisme de rétroaction.56 Nous avons observé un effet biphasique: après un traitement de 24h, le Mel9 et le Mel41 ont induit une phosphorylation de l’ERK, ce qui est en accord avec l'induction de l'expression et de l'activité du MITF, conduisant à la mélanogenèse. Après un traitement de 48h, le Mel9 et le Mel41 inhibaient la phosphorylation d’ERK, suggérant l'existence d'un mécanisme de retroaction retardée.
Ces données mettent en évidence le lien entre l'autophagie et la mélanogénèse, ainsi que la régulation de ces événements par les PHB. Ainsi, il est probable que les analogues de la mélanogénine se lient aux PHB pour réguler l'activité du LC3 et du MITF.
59 De manière intéressante, le Mel9 et en particulier le Mel41 régulent à la baisse les niveaux de protéines PHB (Figure 27) après 24 et 48 h dans les cellules de mélanome HBL. Considérant qu'une régulation négative des PHB est connue pour promouvoir l'apoptose,1 nous avons examiné si le Mel9, le Mel41 et leurs analogues le Mel42, le Mel55, le Mel56 et le Mel57 induisent l'apoptose dans trois cellules de mélanome différentes (Figure 27C). Ces composés, en particulier le Mel9 et le Mel 56, ont significativement induit l'apoptose après un traitement de 48h dans les cellules HBL et MM043, et également, dans une moindre mesure, dans les cellules MM162.
Nous avons également observé que le Mel9 et le Mel41 inhibaient la phosphorylation de le AKT. En effet, l'activation de le AKT favorise non seulement la survie cellulaire, mais empêche également la mélanogénèse.57
60 Figure 27. Ligands de PHB régulent à la baisse le niveau de protéine des PHB et la
phosphorylation d'Akt pour induire l'apoptose dans les cellules du mélanome
En A et B), les cellules HBL ont été traitées par le Mel9 et le Mel41 (0 ; 0,1 ; 1 et 10 μM) pendant 24 et 48 heures. Des quantités égales de protéines extraites (35 μg) ont été analysées par Western blot avec des anticorps dirigés contre le PHB1, le PHB2 et le phospho-AKT. Pour garantir un chargement égal, les transferts ont été évalués avec un anticorps contre la ß-actine (rangée du bas). Les graphiques représentent la moyenne des 2 expériences indépendantes de l'intensité des bandes dans les mêmes conditions de traitement comparées au véhicule.
En C) Les cellules HBL, MM043 et MM162 ont été traitées par le Mel9, le Mel41, le Mel42, le Mel55, le Mel56 et le Mel57 (0, 1 ou 10 μM) pendant 48 heures et le pourcentage des cellules apoptotiques a été déterminé par coloration à l'annexine V (* P ≤ 0,05 , ** P ≤ 0,01, *** P ≤ 0,01).
61 (MM043 et MM162, Figure 28). Ainsi, l'inhibition de l'activité de l'AKT par Mel9 et Mel41 peut contribuer à la fois aux effets inducteurs promélanogéniques et à l'apoptose.
Figure 28. Mel9 régule à la baisse le niveau de protéine des PHB et la phosphorylation d’Akt dans les cellules de mélanome MM043 et MM162.
En A et B), les cellules HBL ont été traitées par le Mel9 (0 ; 0,1 ; 1 et 10 μM) pendant 24 heures. Des quantités égales de protéines extraites (35 μg) ont été analysées par analyse Western blot avec des anticorps dirigés contre le PHB1, le PHB2 et le phospho-AKT. Pour garantir un chargement égal, les transferts ont été évalués avec un anticorps contre la ß-actine (rangée du bas).
4.5 Effet de la mélanogénine dans les cellules de mélanome HBL
Une comparaison des effets de la mélanogénine sur les voies de signalisation dans les cellules HBL, avec ceux des Mel9 et Mel41 a été effectuée. Il a été ainsi observé que la mélanogénine modulait la même signalisation que le Mel9 et le Mel41, mais à une concentration plus élevée et avec une intensité plus faible, ce qui confirme l'activité accrue de ces derniers (Figure 29).
62 Figure 29. Induction de la dégradation de PHB1 et l’activation de la signalisation de MITF dans
les cellules de mélanome HBL par le MEL9, le MEL41 et la mélanogénine.
Des cellules de mélanome humain HBL ont été traitées avec de la mélanogénine (0 ; 0,1 ; 1 et 10 μM) pendant 24 heures. Des quantités égales de protéines extraites (35 μg) ont été analysées par Western blot avec des anticorps contre le MITF, la tyrosinase, le LC3, le phospho-ERK, le PHB1 et le phospho-AKT. Les graphes représentent la moyenne des 2 expériences indépendantes de l'intensité des bandes dans les mêmes conditions de traitement comprées au véhicule.
4.6 Effet sur la croissance des cellules cancéreuses
Puisque les PHB sont des cibles émergentes en oncologie,1,58,59 l’équipe du professeur Thierry Cristeil à la Faculté de Pharmacie de Paris Sud à Chatenay-Malabry a examiné si la mélanogénine est cytotoxique sur diverses lignées de cellules cancéreuses humaines par le dosage MTS. Comme le montre le tableau 12, la mélanogénine réduit significativement la prolifération cellulaire et la viabilité à 10 μM. La cytotoxicité de ce composé n'a pas été précédemment rapportée. Ils ont également trouvé que le Mel9, et en particulier le Mel41, sont plus cytotoxiques que la mélanogénine dans les cellules KB et HL60 (Tableau 11).
Tableau 11. Pourcentage d’inhibition de la croissance cellulaire par la mélanogénine
à 10-5 et 10-6M sur differentes lignées cellulaires cancereuses dans le DMSO, mesuré après 72h d’exposition. Cell line 1 μM 10 μM PC3 15±5 52±5 LNCaP 11±3 32±1 EPC 27±6 58±2 HT29 12±7 20±10 HCT116 10±1 36±7 MCF7 11±9 27±5 MRC5 12±9 61±2 HBL 9+3 18+5