Effet d’une supplémentation en acide folique et
vitamine B
12et de deux niveaux de glucides chez des
vaches de boucherie sur les performances de leurs
veaux.
Mémoire
JOSÉE MERCIER
Maîtrise en Sciences Animales Maître ès Sciences (M.Sc.)
Québec, Canada
iii
Résumé
Le but de l’étude était de déterminer l’impact d’un supplément d’acide folique et de vitamine B12 (v+ ou v-) à la vache et de diètes contenant différentes teneurs en hydrates de carbone non structuraux sur la production laitière, le gain des veaux, le rendement de carcasse et la qualité de la viande des veaux. Trente-deux couples vache-veau ont été répartis en blocs selon la parité de la mère et la date de vêlage (8 blocs de 4 paires) selon un plan en blocs complets aléatoires. Dans chaque bloc, 2 vaches ont reçu une diète pauvre en hydrates de carbone non-structuraux (PHC) et 2 vaches ont reçu une diète riche en hydrates de carbone non-structuraux (RHC). À l’intérieur de chaque diète, les vaches ont reçu un supplément d’acide folique et de vitamine B12 ou aucun supplément. Durant les 100 premiers jours suivant le vêlage, les vaches ont reçu de l’ensilage avec (RHC) ou sans (PHC) 1 kg de mélasse sèche. Entre 100 et 200 jours, les paires vaches-veau ont reçu de nouvelles parcelles d’ensilage quotidiennement à 6h30 (PHC) ou 18h30 (PHC). Entre 200 et 300 jours, les paires vache-veau ont reçu de l’ensilage fauché à 6h00 (PHC) ou 18h00 (RHC). Les veaux ont été abattus à 305 ±12 jours. La production laitière et le gain des veaux n’ont pas été influencés par les traitements (P>0.05). La qualité de la viande n’a pas été influencée par les traitements sauf la concentration en collagène total qui a été diminuée par le supplément de vitamines (P=0.03). Ces résultats indiquent que les suppléments d’acide folique et de vitamine B12 ont peu d’impact chez des vaches de boucherie qui ne sont pas en carence et que les teneurs en hydrates de carbone non-structuraux n’étaient vraisemblablement pas suffisantes pour affecter les performances animales.
v
Abstract
Thirty-two spring calving cows (760 ± 91 kg BW) and calves (44 ± 4.53 kg BW) were blocked according to parity and calving date (8 blocks of 4 cows) according to a randomised complete block design. Within each block, 2 cows were feda low total non-structural carbohydrate (LTNC) diet, while 2 were fed a high TNC (HTNC) diet. Within each diet, cows were administered either no vitamins (v-) or weekly intramuscular injections of 160 mg of folic acid plus 10 mg of vitamin B12 (v+).From parturition to 100 d post-calving cows received grass silage ad libitum with (HTNC) or without 1 kg of dried molasses (LTNC). From day 100 to 200, cow-calf pairs were allotted new pasture daily at 0630h (LTNC) or 1830h (HTNC). During days 200 to 300, cow-calf pairs were fed ad libitum silage cut at 0600h (LTNC) or 1800h (HTNC). Calves were slaughtered at 305±12 d. Treatments had no effect on milk yield (P=0.17) but increased milk vitamin B12 excretion (P<0.01) at 195 days post-calving. Treatments had no effect on calf performance except for meat collagen (P>0.05). These results indicate that folic acid and vitamin B12 supplements have little impact on healthy beef cattle and that plant TNC levels did not differ sufficiently to elicit an animal response.
vii
Avant-Propos
J’aimerais d’abord remercier mon directeur Dany Cinq-Mars et mes co-directeurs, Christiane Girard, Robert Berthiaume et Luigi Faucitano pour l’encadrement qu’ils m’ont offert tout au long du projet, des analyses et de la rédaction. Votre support et votre patience ont été indispensables tout au long du parcours.
J’ai une pensée spéciale pour l’équipe de la ferme expérimentale de Kapuskasing. Maurice, Kevin, Daniel, Larry, Pierre, Guilaine, Marcelle, Aurélien et Élodie. Vos efforts ont été grandement appréciés. J’aimerais aussi remercier Sophie, Sylvie, Chrystiane, Véronique et Ghislain du Centre de recherche et de développement sur le bovin laitier et le porc pour leur assistance avec les analyses de laboratoire.
Merci à Développement Économique Canada pour leur contribution financière au projet et pour ma bourse d’étude.
Je ne saurais passer sous silence ma famille et mon mari qui m’ont supporté et encouragé dans ce périple et sans qui rien n’aurait été possible.
viii
ix
Table des matières
Résumé ... iii
Abstract... v
Avant-Propos ... vii
Table des matières ... ix
Liste des tableaux ... xi
Liste des figures ... xiii
Introduction ... 1
Références de l’introduction... 3
Revue des travaux antérieurs ... 5
1.1- Fourrages ... 5
1.1.1– Effet du moment de la journée sur la concentration de glucides non structuraux des plantes ... 5
1.1.2- Effet du moment d’allocation d’herbe sur le comportement au pâturage 6 1.1.3– Effet d’une quantité supérieure de glucides non structuraux sur l’utilisation de la protéine ... 7
1.1.4– Effet d’une quantité supérieure de glucides non structuraux sur la composition et la digestibilité des plantes fourragères ... 9
1.1.5– Effet d’une concentration plus élevée de glucides solubles dans les fourrages sur la production laitière ... 12
1.1.6- Effet d’une concentration plus élevée de glucides solubles dans les fourrages sur la performance de croissance... 13
1.1.7– Effet de la concentration en protéine d’un pâturage sur les performances animales ... 14
1.2- Supplémentation de la diète en mélasse ... 16
1.2.1 – Effet de la supplémentation de la diète en sucrose sur la production laitière ... 16
1.3 –Folate ... 20
1.3.1- Structure et fonctions de la molécule de folate ... 20
1.4 - Vitamine B12... 22
1.4.1 - Structure et fonctions de la molécule de cobalamine ... 23
1.4.2 - Besoins en acide folique des vaches laitières ... 24
1.4.3 - Impact de la vitamine B12 sur le métabolisme et la production laitière 28 1.5 – Impact des vitamines sur les performances des veaux et bouvillons ... 31
1.5.1 – Impact de l’acide folique sur les performances des veaux en croissance ... 31
1.5.2 - Impact de la vitamine B12 sur la performance de bouvillons ... 32
1.6 - Amélioration de la valeur nutritive de la viande de bœuf ... 35
1.6.1 - Symptômes de carences en cobalamine ... 35
1.6.2 - Consommation de produits animaux et de cobalamine dans la population ... 36
1.6.3 - Boeuf nourri à l’herbe et conditions économiques ... 37
x
Chapitre 2 ... 47
GROWTH PERFORMANCE,CARCASS CHARACTERISTICS AND METABOLISM OF COW -CALF PAIRS RECEIVING HIGH OR LOW TNCDIETS WITH OR WITHOUT FOLIC ACID AND VITAMIN B12SUPPLEMENTS ... 47
Introduction ... 48
Material and methods ... 49
Animals and diets ... 49
Animal Weight ... 51
Milk production ... 51
Forage sampling and analyses ... 52
Meat quality assessment ... 53
Vitamin analysis... 55
Statistical analysis ... 56
Results and discussion ... 57
Forages ... 57
Vitamins in cow plasma ... 59
Milk production ... 59
Calf weight ... 60
Carcass quality characteristics ... 61
Meat quality characteristics ... 65
Meat tenderness ... 66
Conclusions and implications ... 71
Acknowledgements ... 71
Références du chapitre 2 ... 72
xi
Liste des tableaux
Tableau 1.1 adapté de Broderick et Radloff (2004). Effets de variation de glucides totaux sur la production laitière.
Tableau 1.2- adapté de Tiffany et al., 2003. Impact de la concentration de cobalt sur la performance de bouvillons en croissance et finition
Tableau 1.3- adapté de Tiffany et al., 2003. Impact de la concentration de cobalt sur la concentration plasmatique de vitamine B12, d’acide méthylmalonique, de glucose et de propionate.
Tableau 1.4 Coûts de production et de gain selon le mode de production. Adapté de Berthiaume et al., 2006.
Table 2.1 Chemical composition (% DM) of silage, pasture and molasses fed to cows and calves.
Table 2.2 Effects of total non-structural carbohydrate level in diet on forage ingestion. Table 2.3 Total sunshine hours in June, July and August of 2008 compared to a 83 year
mean.
Table 2.4 Effects of total non-structural carbohydrate level in diet and vitamin supplementation of dams on calf average daily gain (ADG).
Table 2.5 Effects of total non-structural carbohydrate level in diet and vitamin supplementation of dams on carcass characteristics of steers.
Table 2.6 Effects of total non-structural carbohydrate level in diet and vitamin supplementation of dams on meat quality traits and composition of steers.
xiii
Liste des figures
Figure 1.1 adaptée de Griggs et al. (2005). Coupe de dactyle à la hauteur des zones grises à intervalles de six heures au début et à la fin des intervalles de six heures dans des séquences de défoliation de 24 heures initiées à 19h00 et à 7h00.
Figure 1.2 Structure chimique des différentes formes de folates réduits ainsi que celle de l’acide folique, Scott, 1999.
Figure 1.3 Fonctions du folate et de la cobalamine, source : Scott, 1999 Figure 2.1 Temperature of carcasses 30 hours after slaughter.
1
Introduction
Au Canada, la plupart des veaux de boucherie naissent au printemps, passent l’été avec leur mère au pâturage et sont envoyés dans des parcs d’engraissement à l’automne, ou encore sont transportés directement à l’abattoir à la fin d’un engraissement à la ferme (Paradis et al. 2008). Lors d’engraissement à la ferme, l’herbe et l’ensilage jouent un rôle majeur dans la nutrition de la vache et du veau de boucherie.
Avant le sevrage, la production laitière est le facteur qui a le plus d’impact sur le gain du veau de boucherie (Totusek et al., 1973). Les veaux de boucherie allaités qui reçoivent un supplément de lait ont un gain moyen quotidien (GMQ) supérieur aux veaux non supplémentés (Berthiaume, observations non-publiées). Ces résultats sont en accord avec les dires de Jasper et Weary (2002) selon lesquels des veaux nourris de lactoremplaceur à volonté ont un GMQ de 0.78 kg avant le sevrage versus 0.48 kg pour les veaux recevant 10% de leur poids vif en lactoremplaceur.
La production laitière des vaches Holstein est augmentée de 1 kg j-1 lorsqu’elles sont nourries avec de l’ensilage fauché en après-midi par rapport à celles nourries avec de l’ensilage fauché le matin (Brito et al. 2008). De même, l’allocation de bandes de pâturage l’après-midi par opposition à l’allocation le matin augmente la production laitière puisque la quantité de matière sèche et de glucides non structuraux est supérieure dans l’herbe en après-midi (Orr et al. 2001). Des concentrations plus élevées de glucides non structuraux ont été retrouvées chez le ray-grass annuel (Gregorini et al. 2006), la fléole (Morin et al. 2012a), la fétuque élevée (Fisher et al. 1999) et la luzerne (Morin et al. 2012b) fauchée en après-midi par rapport au matin.
Dumoulin et al. (1991) ont démontré que des veaux supplémentés avec de l’acide folique ont eu un meilleur GMQ que les veaux témoins. Chez la vache laitière, des suppléments d’acide folique et de vitamine B12 ont augmenté la production de lait corrigé pour
2
l’énergie de 25. 8 kg j-1 chez les vaches recevant des suppléments d’acide folique seul à 29.0 kg j-1 ainsi que la concentration d’acide folique et de vitamine B12 dans le lait (Girard and Matte 2005). L’effet de ces vitamines sur la production laitière et la composition du lait n’est pas documenté sur les vaches de races bouchères. Cependant, en se basant sur les observations faites sur les vaches laitières, on peut supposer qu’un supplément combinant l’acide folique et la vitamine B12 pourrait augmenter les concentrations de ces vitamines dans le lait et leur transfert aux veaux allaités.
Ce projet avait pour but d’étudier les effets de différents niveaux de sucres non structuraux dans les fourrages ou la diète et de suppléments d’acide folique et de vitamine B12 ainsi que leur interaction sur le poids de bouvillons semi-finis, le rendement de carcasse et la qualité de leur viande.
3
Références de l’introduction
Brito, A.F., Tremblay, G.F., Bertrand, A., Castonguay, Y., Belanger, G., Michaud, R., Lapierre, H., Benchaar, C., Petit, H.V., Ouellet, D.R. et Berthiaume, R. 2008. Alfalfa cut at sundown and harvested as baleage improves milk yield of late-lactation dairy cows J. Dairy Sci. 91:3968-3982.
Dumoulin, P.G., Girard, C.L., Matte, J.J. et St-Laurent, G.J. 1991. Effects of a parenteral supplement of folic acid and its interaction with level of feed intake on hepatic tissues and growth performance of young dairy heifers. J. Anim. Sci. 69:1657-1666. Fisher, D.S., Mayland, H.F. et Burns, J.C. 1999. Variation in ruminants’ preference for tall fescue hays cut either at sundown or at sunup. J. Anim. Sci. 77:762-768.
Girard, C.L. et Matte, J.J. 2005. Effects of Intramuscular Injections of Vitamin B12 on Lacation Performance of Dairy Cows Fed Dietary Supplements of Folic Acid and Rumen-Protected Methionine. J. Dairy Sci. 88:671-676.
Gregorini, P., Eirin, M., Refi, R., Ursino, M., Ansin, O.E. et Gunter, S.A. 2006. Timing of herbage allocation in strip grazing: Effects on grazing pattern and performance of beef heifers. J. Anim. Sci. 84:1943-1950.
Jasper, J. et Weary, D.M. 2002. Effects of Ad Libitum Milk Intake on Dairy Calves J. Dairy Sci. 85:3054–3058.
Morin, C., Bélanger, G., Tremblay, G.F., Bertrand, A., Castonguay, Y., Drapeau, R., Michaud, R., Berthiaume, R.R. et Allard, G. 2012. Diurnal variations of
nonstructural carbohydrates and nutritive value in timothy Can. J. Plant Sci. 92 : 883-887.
Morin, C., Tremblay, G.F., Bélanger, G., Bertrand, A., Castonguay, Y., Drapeau, R., Michaud, R., Berthiaume, R.R. et Allard, G. 2012. Nonstructural Carbohydrate Concentration during Field Wilting of PM- and AM-Cut Alfalfa. Agron. J. 104 : 649-660. Orr, R.J., Rutter, S.M., P. D. Penning et Rook, A. J. 2001. Matching grass supply to grazing patterns for dairy cows. Grass Forage Sci. 56:352-361.
Paradis, C., Berthiaume, R., Lafrenière, C., Gervais, R. et Chouinard, P.Y. 2008. Conjugated linoleic acid content in adipose tissue of calves suckling beef cows on pasture and supplemented with raw or extruded soybeans. J. Anim. Sci. 86:1624-1636.
4
Totusek, R., D. W. Arnett, G. L. Holland, et Whiteman J. V. 1973. Relation of estimation method, sampling interval and milk composition to milk yield of beef cows and calf gain. J. Anim. Sci. 37:153–158.
5
Revue des travaux antérieurs
1.1- Fourrages
1.1.1– Effet du moment de la journée sur la concentration de glucides non structuraux des plantes
Les plantes produisent des glucides tels du glucose, du fructose et du sucrose par photosynthèse (Mayland et al., 2005). Durant les heures d’ensoleillement les plantes n’exportent pas les substrats photosynthétiques au même rythme que s’effectue leur synthèse (Fisher et al. 1999). Il en résulte une augmentation de la concentration de glucides non structuraux dans la partie aérienne tant par l’augmentation de la quantité de glucides non structuraux dans la plante que par la diminution de la concentration d’eau dans celle-ci (Huntington et Burns, 2007). La concentration de glucides non structuraux de la plante diminue ensuite durant la nuit lorsque la plante utilise une partie des glucides accumulés durant la journée pour sa respiration (Mayland et al. , 1998; Mayland et al. 2005). Les plantes fourragères contiennent donc plus de glucides solubles en après-midi (Lechtenberg et al., 1971).
La variation quotidienne de glucides non structuraux est une source de variation de ces glucides dans les fourrages récoltés à différents moments de la journée. Fisher et al (1999) ont analysé 2 foins de fétuque élevée du cultivar « HighMag». Le premier foin, récolté au coucher du soleil le 20 août contenait 8,15 % de glucides non structuraux sur une base de matière sèche par rapport à 6,21 % pour le deuxième foin récolté le lendemain au lever du soleil. Cette variation est connue chez plusieurs espèces, c'est-à-dire chez le ray-grass annuel (Gregorini et al., 2006), la luzerne (Morin et al., 2012b) et la fléole des prés (Morin et al., 2012a)
6
La variation quotidienne de la concentration en glucides solubles des plantes fourragères est aussi une source de variations de la quantité de glucides consommés par les animaux au pâturage. La variation de la consommation des glucides est cependant différente en condition de pâturage parce que les plantes sont consommées en couches successives plutôt que d’être coupées en une seule fois (Griggs et al., 2005).
1.1.2- Effet du moment d’allocation d’herbe sur le comportement au pâturage
Le temps passé à brouter par des ruminants au pâturage est exprimé en plusieurs épisodes de paissance, qui sont des repas discrets (Gregorini et al., 2006). Un épisode de paissance est un moment où « l’animal est directement engagé dans l’acte de manger c’est à dire chercher, acquérir dans la gueule, mastiquer et avaler l’herbe (Gregorini et al., 2006). Les ruminants effectuent de 3 à 4 épisodes de paissance majeurs par jour, dont les deux principaux sont au lever et au coucher du soleil, le second étant plus long et plus intense (Gregorini et al., 2006).
La distribution de ces épisodes de paissance dans une journée est flexible selon le moment où les animaux se font allouer une nouvelle parcelle d’herbe, soit le matin ou le soir (Gregorini et al., 2006). Orr et al. (2001) ont observé que des vaches auxquelles on allouait une nouvelle parcelle d’herbe à 7h45 ou à 16h45 passaient le même temps total dans la journée à manger tandis que Gregorini et al. (2006) ont observé que des génisses à qui on allouait une nouvelle parcelle à 7h00 passaient 54 minutes de plus chaque jour à se nourrir que celles à qui on allouait une nouvelle parcelle à 15h00. Néanmoins, les deux équipes ont observé des comportements similaires chez les animaux au niveau de la distribution temporelle et de l’intensité des épisodes de paissance.
L’épisode de paissance du soir est plus long et plus intense chez les vaches qui reçoivent une nouvelle parcelle en après-midi (Orr et al., 2001). En effet, les vaches ayant participé à cette expérience ont effectué un épisode de paissance de plus de quatre heures en moyenne, entre l’allocation de la parcelle et le coucher du soleil. En contrepartie, les vaches recevant une nouvelle parcelle le matin ont effectué un épisode de paissance
7 intermittent d’environs trois heures en après-midi, puisqu’elles avaient déjà eu un épisode de paissance de deux heures le matin. Les vaches du traitement de l’après-midi avaient eu seulement de petits épisodes de paissance dispersés le matin (Gregorini et al. 2006). Durant l’épisode de paissance du soir, le rythme de prise de bouchées était plus rapide chez les vaches recevant une nouvelle parcelle en après-midi (Gregorini et al., 2006), surtout lors de la première heure suivant l’allocation (Orr et al., 2001). Bien que la masse des bouchées demeure la même pour les deux traitements, il en résulte une préhension d’herbe plus intense durant cet épisode de paissance (Orr et al., 2001).
Puisque les animaux effectuent leur plus important épisode de paissance lorsqu’ils reçoivent une nouvelle parcelle d’herbe, les génisses recevant leur nouvelle parcelle le matin passent 36 % de leur temps de paissance quotidien à paître le matin, par rapport à 28 % pour celles qui reçoivent leur allocation d’herbe le soir (Gregorini et al., 2006). Puisqu’en avant midi, l’herbe contient une moins grande concentration de matière sèche et un ratio
Glucides non structuraux Protéines
moins élevé que le soir, l’énergie consommée par les génisses chaque jour peut avoir été différente même si la quantité totale de matière sèche ingérée était la même (Gregorini et al., 2006). On peut donc déduire de ces deux études que d’allouer une nouvelle parcelle d’herbe le soir ou l’après-midi favorise un épisode de paissance plus long et plus intense, au moment où l’herbe est la plus nutritive (Gregorini et al, 2006).
1.1.3– Effet d’une quantité supérieure de glucides non structuraux sur l’utilisation de la protéine
Les pertes d’azote constituent un problème autant au niveau du gaspillage de nutriments, causant des pertes monétaires, qu’au niveau de la pollution causée par l’excrétion de l’urée contenue dans l’urine. Les pertes ruminales d’azote sont les plus importantes lors de la paissance d’un foin jeune et feuillu, parce qu’il procure à l’animal une grande
8
quantité de protéines solubles (Van Vuuren, 1993). La concentration en glucides solubles des plantes est plus faible le matin (Gregorini et al., 2006), au moment où la concentration ruminale d’azote est à son plus élevé (Chilibroste et al., 2005). Ce surplus d’azote ammoniacal, en présence d’un déficit de glucides solubles cause un déséquilibre dans la disponibilité de l’énergie et de la protéine qui nuit à la prolifération des microorganismes ruminaux (Chilibroste et al., 2005).
Des études ont donc démontré que la concentration d’azote ammoniacal du rumen de bouvillons était moins élevée lorsqu’ils ingéraient un fourrage contenant plus de glucides non structuraux (Lee et al., 2002). L’impact de cette réduction demeure cependant mal compris. Moorby et al. (2005) et Lee et al. (2002) suggèrent tous deux qu’il n’y a pas d’amélioration dans l’efficacité ruminale même s’il y a diminution de la concentration d’azote ammoniacal. Broderick et Radloff (2004) et Miller et al. (2001) ont eux suggéré une meilleure utilisation de l’azote par des vaches recevant une diète plus élevée en glucides solubles. Des bouvillons à qui ont a offert du ray-grass vivace ayant un contenu élevé en glucides solubles ont produit 15,9 g d’azote microbienne par kg de matière organique digérée, par rapport à 17,8 g pour les bouvillons du traitement témoin ce qui demeure statistiquement similaire. De même, l’efficacité du transfert d’azote non-ammoniacal et d’azote microbien au duodénum a été similaire dans les deux traitements. L’absorption d’azote et d’acides aminés au niveau du duodénum a néanmoins été plus grande chez les animaux recevant un fourrage d’une concentration plus élevée en glucides solubles. Celà serait dû à une plus grande consommation d’azote par les animaux, puisque les bouvillons mangent plus d’un fourrage contenant plus de glucides solubles (Lee et al., 2002).
Miller et al. (2001) ont observé une consommation d’azote similaire chez des vaches recevant elles aussi du ray-grass ayant une concentration plus élevée de glucides non structuraux ou un ray-grass témoin. Contrairement à l’étude ci-haut, les vaches avaient une consommation de fourrage similaire pour les deux traitements. Ces auteurs ont cependant observé que l’azote ingérée étaient métabolisée différemment. Dans les traitements élevés en glucides solubles il y a eu une augmentation proportionnelle de 30
9 % de l’azote alimentaire qui se dirigeait vers le lait, compensée par une diminution proportionnelle de 26 % de l’azote excrété dans l’urine. Il est donc nécessaire de continuer la recherche pour connaître les facteurs qui font varier l’efficacité de la fermentation ruminale et découvrir les facteurs qui promeuvent cette efficacité.
1.1.4– Effet d’une quantité supérieure de glucides non structuraux sur la composition et la digestibilité des plantes fourragères
La quantité de glucides non structuraux influence la composition des plantes qui varie quotidiennement. Ainsi, des fourrages sélectionnés génétiquement pour contenir une grande quantité de glucides solubles, récoltés l’après-midi, contiennent moins de fibres solubles au détergent neutre (NDF) et de fibres solubles au détergent acide (ADF) (Lee et al., 2002; Moorby et al., 2005). Ils contiennent donc moins de cellulose et de lignine (Fisher et al., 1999). La concentration en matière sèche des plantes coupées l’après-midi est plus élevée et la concentration en acides gras à courte chaîne plus faible (Huntington et Burns, 2007).
Au niveau de la concentration en azote et en protéine brute, les résultats se contredisent. Huntington et Burns (2007) et Moorby et al. (2005) ont trouvé des quantités moindres de protéine brute et d’azote, respectivement. Cependant, Lee et al. (2002) ont trouvé une concentration d’azote similaire pour deux fourrages différents en concentration de glucides solubles, coupés l’avant-midi ou l’après-midi. Ces résultats sont d’autant plus surprenants que les cultivars de ray-grass vivaces utilisés sont les mêmes que pour Moorby et al. (2005) et les heures de coupes sont également les mêmes. Il serait donc intéressant de connaître la variation en protéine brute d’un ensilage fermenté coupé l’avant-midi ou l’après-midi.
La digestibilité vraie de la matière sèche est plus élevée chez de la fétuque élevée coupée l’après-midi, avec une digestibilité de 85,1 %, par rapport à 83,4 % pour le même cultivar coupé le matin (P<0,01) (Fisher et al., 1999). Les études se contredisent sur la digestibilité apparente de la matière sèche et de la matière organique. Lee et al. (2002)
10
n’ont pas trouvé de différence entre deux fourrages de concentration en glucides non structuraux différente, tandis que Moorby et al. (2005) ont trouvé des différences significatives pour ces deux analyses (P < 0,05) en faveur des fourrages à haute teneur en glucides solubles. Ces résultats sont d’autant plus surprenants que les deux ont, comme indiqué précédemment, utilisé les mêmes cultivars de ray-grass vivace et les ont coupés au même moment de la journée. Il est possible que le moment de l’année où a été effectué les coupes, c'est-à-dire au début avril pour Lee et al. (2002) et à la mi-mai pour Moorby et al. (2005) ait pu influencer les résultats puisqu’il est bien connu que le climat influence les teneurs en glucide des fourrages.
Plus spécifiquement, la digestibilité apparente de l’azote et des fibres solubles au détergent neutre est similaire entre des fourrages à teneur élevée ou moyenne en glucides non structuraux (Lee et al., 2002).
La digestibilité des fourrages, tout comme leur composition, sont différentes lorsque l’animal est au pâturage, puisque la plante n’est pas consommée d’un seul coup, mais plutôt par couches successives (Griggs et al., 2005). Griggs et al. (2005) ont effectué deux séquences de quatre coupes à l’intérieur de 24 heures sur du dactyle, tel que démontré dans la figure 1.1.
11 Figure 1.1 Coupe de dactyle à la hauteur des zones grises à intervalles de six heures au début et à la fin des intervalles de six heures dans des séquences de défoliation de 24 heures initiées à 19h00 et à 7h00. Adaptée de Griggs et al. (2005).
Ils ont découvert que la digestibilité ne variait que légèrement et diminuait avec les coupes successives dans les deux séquences. Cependant, la défoliation initiale, c'est-à-dire l’herbe coupée pour la première fois à 19h00, a une digestibilité vraie in vitro significativement plus élevée que l’herbe coupée pour la première fois à 7h00. Bien que la variation de digestibilité et de concentration en glucides solubles soit faible sur 24 heures, un repas du soir plus important au moment où la digestibilité et la concentration en glucides solubles du dactyle sont significativement plus élevées pourrait expliquer le gain de performance obtenu par d’autres chercheurs avec des animaux se faisant allouer des parcelles d’herbe le soir (Griggs et al., 2005).
12
1.1.5– Effet d’une concentration plus élevée de glucides solubles dans les fourrages sur la production laitière
Une étude antérieure de Berthiaume (observations non publiées) a démontré que de supplémenter la diète de veaux avec un lactoremplaceur offert dans une chaudière a significativement augmenté le gain moyen quotidien de ces veaux. De leur naissance jusqu’à l’âge de 100 jours, les veaux recevant 2 litres de lactoremplaceur par jour ont effectué un gain moyen quotidien de 1,06 kg par jour par rapport à 1,02 kg par jour pour des veaux ne recevant pas de lactoremplaceur (P < 0,001). De même, de l’âge de 200 à 290 jours, les bouvillons recevant 5 litres de lactoremplaceur par jour ont effectué un gain moyen quotidien de 0,825 kg par jour par rapport à 0,599 kg par jour pour des bouvillons ne recevant pas de lactoremplaceur (P < 0,001). Cependant, puisque la consommation volontaire de lait varie beaucoup entre les veaux de la naissance au sevrage, cette supplémentation apparaît difficile (Berthiaume, observations non publiées). Cette étude a donc démontré que la croissance des veaux est limitée par la production de lait des vaches. Il est possible de croire que d’améliorer la production laitière des vaches allaitantes à l’aide de fourrages à plus haute teneur en glucides solubles aurait un effet favorable sur les performances de gain des veaux allaités.
Kim (1995) a démontré que des vaches laitières nourries avec du foin de luzerne récolté l’après-midi ont produit environs 10 % plus de lait que des vaches nourries avec le même foin de luzerne, mais récolté l’avant-midi, entre autres parce que les vaches du traitement de l’après-midi ont mangé 10 % plus de foin que les autres. Mertens et Mayland (non publié), cité par Mayland et al. (2005) ont obtenu des résultats variables sur ce même sujet. Quatre groupes de vaches laitières, deux en Idaho et deux au Wisconsin, ont été nourries avec soit du foin de luzerne récolté au lever ou au coucher du soleil. Les vaches recevant du foin récolté au coucher du soleil ont produit 6 % plus de lait que celles recevant du foin récolté au lever du soleil en Idaho. Il n’y a pas eu d’effet significatif du temps de récolte sur la production laitière au niveau des vaches du Wisconsin (Mertens et Mayland, non publié, cité par Mayland et al., 2005). De même, Orr et al. (2001) ont fait pâturer deux groupes de vaches laitières dans des champs de ray-grass vivace. Un groupe
13 recevait une nouvelle parcelle à 7h45, tandis que l’autre recevait leur nouvelle parcelle à 16h45. La production laitière a été similaire entre les deux groupes durant les 6 premières semaines de l’expérience pour ensuite se différentier avec 5 % plus de lait produit par les vaches recevant leur parcelle à 16h45 (P < 0,07) (Orr et al.,2001) La production laitière tendait à être plus élevée chez les vaches recevant leur parcelle l’après-midi, mais le groupe était trop petit pour voir une variation significative (Orr et al., 2001). Devant la diversité des résultats et l’absence d’études sur des vaches de races bouchères il serait intéressant de voir l’impact de l’allocation de parcelle le matin ou le soir sur la production laitière de celles-ci, afin d’améliorer les gains de leurs veaux.
Tout comme les données de production laitière, les données de variations de composantes du lait varient. La consommation de foin riche en glucides non structuraux augmente la production de protéine vraie et de caséine dans le lait (Miller et al., 2001; Moorby et al., 2005). Miller et al. (2001) ont trouvé des quantités supérieures de gras, de protéine brute, de petit-lait et de lactose tandis que Moorby et al. (2005) ont trouvé des quantités supérieures de petit lait, de protéine vraie et de caséine. Cependant, dans les deux études, les quantités de composantes étaient plus élevées dans le lait de vaches nourries avec du foin contenant plus de glucides non structuraux, que la variation soit significative ou non, sauf dans le cas de l’azote non protéique, dont la quantité était moindre dans le lait des vaches de ce traitement (Miller et al., 2001; Moorby et al., 2005).
1.1.6- Effet d’une concentration plus élevée de glucides solubles dans les fourrages sur la performance de croissance
Une étude antérieure de Berthiaume (communication personnelle) a démontré que des veaux de races bouchères qui avaient accès entre 100 et 200 jours, avec leur mère, à une nouvelle parcelle de pâturage à 19h00 le soir avaient un gain moyen quotidien de 1,14 kg en comparaison avec un gain moyen quotidien de 1,05 kg pour des veaux semblables qui recevaient une nouvelle parcelle à 7h00 (P < 0,001). Ces mêmes animaux ont par la suite été nourris avec de l’ensilage récolté le soir ou le matin. Les bouvillons nourris d’ensilage récolté le soir et le matin ont eu des gains moyens quotidiens de 0,69 et de 0,73,
14
respectivement (P < 0,001). La plus faible croissance des animaux recevant l’ensilage récolté le soir a pu être dû à la mauvaise conservation des ensilages qui a réduit le gain des bouvillons (Portelance, communication personelle).
Les résultats de Gregorini et al. (2006) confirment ceux de Berthiaume. Ils ont alloué de nouvelles parcelles de ray-grass annuel en pâturage à des génisses bouchères à 7h00 et à 19h00. Au printemps, les génisses recevant leur allocation de parcelle à 19h00 ont gagné en moyenne 549g (P < 0,01) et 0,01 points de condition de chair (P < 0,05) de plus que les génisses recevant leur allocation de parcelle à 7h00. L’avantage sur le gain moyen quotidien procuré par l’allocation d’une nouvelle parcelle de pâturage le soir a été observé dans les deux expériences, mais cela demeure un sujet peu étudié. De plus, il serait intéressant d’observer les performances de bouvillons nourris avec de l’ensilage bien conservé récolté le matin ou le soir, afin de découvrir si la conservation de l’ensilage avait influencé les résultats de l’expérience de Berthiaume (observations non publiées).
1.1.7– Effet de la concentration en protéine d’un pâturage sur les performances animales
Carruthers et Neil (1997) ont examiné l’impact de pâturages contenant 17,6 % et 13,2 % de protéine brute sur la production laitière de seize vaches Holstein. La consommation volontaire des vaches n’a pas été influencée par le traitement. Cependant les vaches consommant un fourrage plus concentré en protéines ont produit plus de lait (19,9 vs 18,7 kg/vache), de gras (0,93 vs 0,86 kg/vache), de protéine (0,64 vs 0,61 kg/vache) et de lactose (0,99 vs 0,92 kg/vache) (P<0.001). Les concentrations des composantes du lait n’ont pas varié sauf pour la concentration en protéine qui a diminué (3,23 vs 3,31 %; (P < 0,05) et celle d’urée dans le lait qui a augmenté (5,8 vs 2,8 mmol/l; P < 0,001). Il est possible que l’impact de la variation des pâturages en protéine ait été aussi important parce que les vaches consommant un fourrage moins concentré en protéines étaient probablement déficientes en protéine, ce qui influençait négativement leurs performances (Carruthers et Neil, 1997).
15 Ces résultats confirment ceux de Petit et al. (1994) qui ont trouvé que des bouvillons à qui on a offert une diète supplémentée en tourteau de canola comprenant 23,09 % et 26,11 % de protéine ont eu des gains moyens quotidiens de 1,09 et 0,22 kg par jour respectivement. Ils ont également démontré une conversion alimentaire accrue. Bien que ces données soient éloquentes, les bouvillons y étaient en déficience protéique ou recevaient un supplément protéique. Il serait intéressant d’examiner la performance de vaches et de bouvillons auxquels on offre seulement des fourrages dont la teneur protéique varie.
16
1.2- Supplémentation de la diète en mélasse
1.2.1 – Effet de la supplémentation de la diète en sucrose sur la production laitière L’équipe de Huhtanen (1987) a découvert que d’ajouter un kg de mélasse par jour dans la ration des vaches a augmenté la production laitière de 23,3 à 24,0 kg/d et la production de protéine de 0,72 à 0,76 kg/d. Cependant l’effet était quadratique car ajouter deux kg de mélasse par jour dans la ration des vaches diminuait significativement la production de lait, de protéine et de gras. Toutefois, ces vaches recevaient un supplément de concentrés à un niveau de 6 kg par jour.
L’effet de la supplémentation de la diète de vaches laitières comparativement aux performances de vaches nourries uniquement de fourrages à été mesurée par Carruthers et Neil (1997). Seize vaches Holstein ont eu accès à un pâturage composé de 77-83 % de ray-grass vivace, 8-16 % de brome et de millet, 4-5 % de trèfle blanc, 1-2 % de plantes adventices et 1-3 % de matière morte. Huit de ces vaches ont reçu un supplément de glucides non structuraux, surtout sous forme d’amidon, composé de farine de maïs et de monohydrate de dextrose, à 10 % de la consommation d’énergie métabolisable de fourrage des vaches. Ces vaches ont donc consommé une diète comprenant 29,4 % de glucides non structuraux par rapport à 22,0 % pour les vaches nourries uniquement au pâturage. La concentration de protéines brutes de la diète était de 16,1 % et de 17,6 %, respectivement. Les vaches supplémentées ont consommé plus que les vaches non supplémentées (15,6 kg par vache vs 14,5 kg par vache), bien que les vaches supplémentées aient consommé moins de fourrages (14,3 kg par vache vs 14,5 kg par vache). La supplémentation n’a pas eu d’effet sur la digestibilité de la matière sèche ou de la matière organique, mais a diminué la digestibilité de l’azote de 4 %. Le supplément d’amidon a diminué la quantité d’azote excrétée dans l’urine mais a augmenté de 3 g seulement par jour la rétention d’azote, puisque plus d’azote était excrété dans les fèces. La concentration en azote ammoniacal ruminal a été plus faible chez les vaches recevant un supplément, de même que le pH ruminal. Cela peut être expliqué par une meilleure
17 efficacité de production de protéine microbienne et une augmentation de la production de protéine microbienne. Le supplément n’a pas eu d’effet sur la production laitière ou sur la production totale de gras ou de lactose dans le lait. La supplémentation en amidon a cependant augmenté la production totale de protéines laitières et diminué la concentration en urée du lait (Carruthers et Neil, 1997). L’effet presque nul de la supplémentation en glucides solubles sur la production laitière pourrait être le résultat de la faible quantité de protéine présente dans le pâturage plus que du manque d’effet du traitement (Carruthers et Neil, 1997).
Les diètes composées en majorité d’ensilage contiennent une grande quantité de protéines dégradables dans le rumen (Muck, 1987; MacDonald et al., 1991) qui peuvent être mal utilisées si une quantité suffisante de glucides facilement fermentescibles n’est pas ingérée au même moment (Kim et al., 1999). Chamberlain et al. (1993) ont démontré que les sucres sont fermentés plus rapidement dans le rumen que l’amidon. Potentiellement pour cette raison, les sucres solubles contribuent à produire 18 % plus de protéine microbienne dans le rumen que l’amidon (NRC, 1996). Hall (2002) avance que la mélasse est une source pratique de sucres alimentaires pour les vaches laitières.
Puisque la production laitière est améliorée avec un supplément d’amidon chez des vaches nourries de fourrages et qu’elle l’est également chez des vaches à qui l’on offre un supplément de sucres solubles sous forme de mélasse, il est intéressant d’observer si la mélasse, donc le sucrose, est plus efficace que l’amidon pour améliorer les performances laitières. Broderick et Radloff (2004) ont donc effectué une étude sur l’impact de la supplémentation en mélasse par rapport à la supplémentation de maïs-grain humide sur la production laitière de vaches nourries à base d’ensilage de luzerne et de maïs. Quatre diètes ont été offertes avec des niveaux de sucres totaux variables comme montré dans le tableau 1.1.
18
Tableau 1.1 : Effets de variation des sucres totaux de la ration sur la production laitière.
Sucres totaux % Contrastes orthogonaux
Production 2,4 4,2 5,6 7,2 SE Linéaire Quadratique Cubique
Consommation (kg/jour de MS) 25,3b 25,7ab 26,3a 26,0ab 0,3 0,05 0,23 0,37 Gain de poids (kg/d) 0,41 0,16 0,36 0,19 0,09 0,27 0,65 0,11 Lait (kg/d) 38,0ab 37,5ab 38,9a 36,7b 0,6 0,34 0,16 0,04 N du lait/ consommation de N (%) 25,5a 24,4ab 25,4a 23,1b 0,6 0,02 0,29 0,03 Gras (kg/d) 1,52ab 1,60ab 1,61a 1,50b 0,04 0,82 0,02 0,72 Protéine (kg/d) 1,19a 1,15ab 1,20a 1,11b 0,03 0,11 0,34 0,05
Azote uréique du lait (mg/dL) 15,1ab 14,2b 15,4a 14,7ab 0,4 0,97 0,82 0,02
Adapté de Broderick et Radloff (2004).
19 Il y a donc eu une augmentation de la consommation volontaire, potentiellement due à l’augmentation de l’appétence des rations contenant plus de mélasse sèche. La consommation plus élevée n’a cependant pas eu d’effet sur le gain de poids, la production de lait, de protéine ou d’azote uréique du lait. On a observé une diminution de l’efficacité d’excrétion d’azote dans le lait. On a également observé une augmentation quadratique de la production de gras, ce qui est probablement dû à l’augmentation de la production de butyrate dans le rumen (Khalili et Huhtanen, 1991). Les résultats sont confondus par une diminution de la quantité de sucres non structuraux lorsque la quantité de mélasse augmente dans la ration et que la quantité de maïs-grain humide diminue parce que la mélasse contient plus de fibres et donc moins de sucres non structuraux que le maïs-grain humide. La mélasse a été séchée sur du tourteau de soya qui contient beaucoup de fibres solubles au détergent neutre et a ainsi apporté une variable de plus à prendre en compte pour les résultats (Broderick et Radloff, 2004). En effet, Broderick et Radloff (2004) ont observé une augmentation quadratique de la production laitière (P < 0.05) et de la production de protéine laitière (P < 0.02) lors de la substitution de maïs-grain humide par de la mélasse humide. Il y a donc augmentation de la productivité avec une supplémentation de sucrose par rapport à une supplémentation d’amidon. De plus, les auteurs indiquent que des résultats optimaux pourraient être obtenus avec une diète comprenant 5,0 % de sucre, ce qui correspond aux dires de Hall (2002).
Il est observé que le sucrose améliore les performances laitières par rapport à l’amidon et qu’un supplément d’amidon améliore les performances laitières de vaches nourries de fourrages. Il serait aussi intéressant d’observer l’effet d’un supplément de sucrose sous forme de mélasse chez des vaches nourries de fourrage, et plus encore chez des vaches de race bouchère chez qui il n’existe pas de données à ce sujet.
20
1.3 –Folate
1.3.1- Structure et fonctions de la molécule de folate
Les folates sont constitués d’un anneau pteridine, d’un p-aminobenzoate et d’une chaîne de polyglutamate comme l’illustre la figure 1.2 (Scott, 1999). Leur unique fonction étant d’accepter des unités d’un carbone pour ensuite les transmettre à d’autres molécules (Scott et Weir, 1994), ils peuvent être retrouvés dans les cellules soit sous la forme de tétrahydrofolates monoglutamylés, soit sous une forme comprenant un groupement monocarboné (Scott, 1999). Les formes comprenant un groupement monocarboné sont le 10-formyltetrahydrofolate, le 5,10-méthylenetetrahydrofolate et le 5-méthyltetrahydrofolate (Scott, 1999). Les folates réduits sont extrêmement instables et faciles à couper entre le C-9 et le N-10 (Scott, 1999). Cette instabilité des folates se trouve également dans la nourriture, ce qui rend leur absorption alimentaire difficile à prévoir. Il est donc bénéfique de supplémenter l’alimentation des monogastriques avec de l’acide folique. L’acide folique est une molécule de synthèse beaucoup plus stable que les folates naturels (Scott, 1999). Elle n’est également pas réduite, contrairement à ses homologues naturels (Scott, 1999). La figure 1.2 démontre la structure chimique des différentes formes de folates réduits ainsi que celle de l’acide folique.
Les folates, ou l’acide folique réduit par les cellules en tétrahydrofolate, ont pour rôle de d’accepter et de transmettre des unités d’un carbone (Choi et Mason, 2000). Ils contribuent ainsi à la formation de purines et de pyrimidines d’ADN (Bailey et Gregory, 1999). Afin de participer à la formation de pyrimidines, les molécules de tetrahydrofolates reçoivent une unité d’un carbone de molécules de sérine et les tetrahydrofolates deviennent ainsi des molécules de 5,10-méthylenetetrahydrofolate (Scott, 1999). Ces dernières donnent une unité d’un carbone à l’enzyme thymidylate synthase qui elle, transforme des bases d’uracile en bases de thymine (Scott, 1999). De
21 même, les molécules de tetrafolates reçoivent une unité d’un carbone du formate pour former du 10-formyltetrahydrofolate. Ce dernier fournit des unités d’un carbone aux enzymes qui synthétisent des purines, afin que celles-ci apposent les carbones aux C-2 et C-8 de l’anneau des purines (Scott, 1999).
Figure 1.2- Structure chimique des différentes formes de folates réduits ainsi que celle de l’acide folique, Scott, 1999.
Le folate est également essentiel au cycle de méthylation (Girard et Matte, 2005b). Les molécules de 5,10-méthylenetetrahydrofolate sont transformées en méthyltetrahydrofolate par l’enzyme 5,10-méthylenetetrahydrofolate reductase. Le
5-22
méthyltetrahydrofolate donne ensuite un groupement méthyle à l’enzyme méthionine synthase qui l’appose sur de l’homocystéine qui devient ainsi de la méthionine (Scott, 1999). L’apport d’une molécule d’adénosine triphosphate active la méthionine qui devient ainsi de la S-adénosylmethionine. Cette dernière agit comme donneur d’un groupe méthyle pour une foule d’enzymes transférant des groupes méthyles (Scott, 1999). Les substrats de méthylation sont principalement des lipides, des hormones, de l’ADN et des protéines (Scott, 1999). Une schématisation de l’action des folates au niveau cellulaire est illustrée dans la figure 1.3.
Figure 1.3- Fonctions du folate et de la cobalamine, source : Scott, 1999
23 1.4.1 - Structure et fonctions de la molécule de cobalamine
La cobalamine, mieux connue sous le nom de vitamine B12, est composée d’un anneau de porphyrine fait de quatre pyrroles avec une molécule de cobalt en leur centre. Elle est la plus grande des vitamines du complexe B avec un poids moléculaire de plus de 1000.
La cobalamine sert de cofacteur à plusieurs enzymes dont les principales sont méthionine synthase et méthylmalonyl-CoA mutase. Dans le cycle de méthylation, la cobalamine sous forme de méthylcobalamine transporte un groupement méthyle entre les méthyltetrahydrofolates et l’homocystéine (Girard et Matte, 2005) (Scott, 1999). La cobalamine est également nécessaire à la néoglucogenèse à partir de propionate, d’acides gras à chaînes de carbone comprenant un nombre impair de carbones ainsi que certains acides aminés (Girard et Matte, 2005) (Scott, 1999). La cobalamine sous forme de 5’-deoxy-5’-adenosylcobalamine est impliquée dans la transformation du méthylmalonyl-CoA en succinyl-CoA à l’aide de l’enzyme méthylmalonyl-CoA mutase (Girard et Matte, 2005). La vitamine B12 est donc particulièrement importante chez les ruminants qui doivent synthétiser la majorité de leur glucose à partir de propionate à travers ce procédé (Scott, 1999).
Puisqu’à la fois le folate et la cobalamine sont nécessaires au bon fonctionnement de la méthionine synthase, il y a un lien entre le métabolisme des folates et celui des cobalamines (Girard et Matte, 2005). Si l’un des deux n’est pas présent en quantité suffisante, des signes de carences apparaissent même si l’autre est suffisant (Scott, 1999).
Dans le contexte de la production bovine, des suppléments d’acide folique et de cobalamine pourraient affecter la performance des veaux de deux façons, soit en augmentant la production de lait ou en augmentant la production de composantes du lait. Il a été prouvé dans une étude antérieure qu’un apport supplémentaire de lait augmente le gain des veaux de race bouchère (Berthiaume, non publié) et chez les
24
veaux de race laitière (Jasper et Weary, 2002). L’autre effet pourrait être plus directement une augmentation du gain chez les veaux par l’apport de ces vitamines. Cependant, puisque les veaux pré-ruminants ne mangent pas de minéral, les vitamines devraient être consommées dans le lait. Comme une étude antérieure a démontré que les veaux étaient récalcitrants à boire du lactoremplaceur lorsqu’ils étaient encore allaités (Berthiaume et al., non publié), le moyen le plus simple serait d’augmenter la quantité d’acide folique et de cobalamine dans le lait maternel. Pour toutes ces raisons il est important de regarder l’impact de l’acide folique et de la cobalamine sur la production des vaches.
1.4.2 - Besoins en acide folique des vaches laitières
L’équipe de Girard a effectué une étude sur la variation de folates dans le sérum sanguin de vaches laitières, avec comme hypothèse que des variations pourraient indiquer qu’en certaines périodes les besoins en folate ne sont pas complètement comblés (Girard et al., 1989). Ils ont découvert que la quantité totale de folates était maximale deux mois après la parturition (836,63 µg), avec une augmentation entre la quantité à la parturition (503,59 µg) et ce point, suivi décroissance entre 3 mois de gestation (685,24 µg) et la parturition suivante (P< 0,05) (Girard et al., 1989). Ces résultats concordent avec des données sur les truies chez qui la gestation et la lactation n’ont pas lieu de façon concomitante. La concentration de folates dans le sérum sanguin diminue donc durant la gestation pour ensuite augmenter durant la lactation, jusqu’au sevrage (Matte et al., 1984; Matte et Girard, 1989). Une étude plus récente de Girard et al. (1994) a produit des résultats similaires au niveau de la variation des folates dans la lactation et la gestation.
Parallèlement, l’équipe de Girard a également effectué une étude sur l’impact d’injections d’acide folique sur la concentration de folates dans le sérum de vaches laitières. Les injections ont effectivement augmenté la concentration de folates dans le
25 sérum sanguin chez les vaches taries du jour 0 au jour 1, avec la concentration la plus élevée obtenue après une injection de 160 mg d’acide folique (Girard et al., 1989). Les injections n’ont pas eu d’impact sur la concentration en folates du sérum ou du lait des vaches en lactation (Girard et al., 1989). L’effet nul de l’acide folique sur les vaches en lactation peut être expliqué par le fait qu’une concentration élevée d’acide folique dans le sérum sanguin provoque une excrétion rénale rapide (Girard et al., 1989). De plus, les vaches taries qui sont gestantes possèdent des protéines qui se lient aux folates et ainsi les protègent de l’excrétion rénale (Girard et al., 1989).
La variation des folates dans le sérum démontre que la production d’acide folique par les bactéries du rumen n’est pas suffisante pour prévenir les fluctuations. De même, l’effet d’une injection d’acide folique chez les vaches taries indique qu’un apport exogène pourrait augmenter la quantité de folates disponible pour le fœtus (Girard et al., 1989).
Suite à ces découvertes, l’équipe de Girard ont poursuivi leurs recherches afin de découvrir si une injection d’acide folique aurait un effet sur les folates présents dans le sérum des vaches et sur les performances de la vache (Girard et al., 1994). Vingt-quatre vaches laitières ont soit reçu une injection de saline ou de 160 mg d’acide folique à partir de 45 jours pré-partum jusqu’à 6 semaines post-partum (Girard et al., 1994). Les injections de 160 mg d’acide folique ont été suffisantes pour élever le taux de folates présentes dans le sérum des vaches (Girard et al., 1989). Il est cependant intéressant de noter que le taux était inférieur chez les primipares par rapport aux multipares (P ≤ 0,03) (Girard et al., 1994).
Contrairement aux résultats de Girard et al. (1989), les folates présents dans le lait des vaches tendent à être plus élevés chez les vaches supplémentées avec 44,0 ± 2,3 ng/ml chez les vaches contrôle et 53,9 ± 4,8 ng/ml pour les vaches supplémentées (P = 0,12) (Girard et al. 1994). Cependant, pour les folates du lait, la supplémentation n’aurait aucun effet de la parturition jusqu’à 6 semaines après celle-ci. Cela pourrait être dû à un
26
mécanisme de régulation du folate laitier qui maintient la concentration en folate du lait, ce au détriment des réserves maternelles (Tamura et al., 1980). Si tel est le cas, la baisse du taux de folates dans le sérum sanguin indiquerait que les besoins des vaches multipares sont plus élevés en fin de lactation (Girard et al., 1994).
La production laitière des vaches supplémentées a été augmentée par l’acide folique. Bien que la différence n’ait pas été significative (P = 0,09), l’augmentation est importante, de l’ordre de 14 %, avec 11,0 ± 0,6 kg/j pour les vaches contrôle et 12,5 ± 0,6 kg/j pour les vaches supplémentées (Girard et al., 1994). L’augmentation a été similaire pour les primipares et les multipares. La supplémentation a également eu l’effet d’augmenter la concentration en protéine du lait des multipares de 3,23 ± 0,06% à 3,51 ± 0,09% (P = 0,07) sur toute la lactation (Girard et al., 1994). Ces nouvelles données ont donc indiqué que, bien que l’acide folique produite par les bactéries du rumen soit suffisante pour prévenir les déficiences et ainsi soutenir la gestation et la lactation chez la vache laitière multipare, elles sont probablement insuffisantes pour permettre à cette vache multipare d’atteindre un maximum de production de lait et de protéine laitière durant la gestation et la lactation (Girard et al., 1994). Il est suggéré que l’effet plus marqué du supplément d’acide folique chez les multipares soit dû aux nombreuses lactations et gestations qu’ont subies ces vaches, qui ont eu pour effet de diminuer leurs réserves d’acide folique. De plus, ces vaches produisent des veaux plus gros et plus de lait (Girard et al., 1994). La demande en folate du fœtus est possiblement la raison pour laquelle la teneur en folates du lait ne serait pas influencée par le supplément à partir de la parturition jusqu’à 6 semaines après celle-ci, moment où les vaches ne sont pas gestantes (Girard et al., 1994).
L’équipe de Girard a par la suite effectué une étude sur l’impact d’un supplément alimentaire d’acide folique sur la production laitière et la composition du lait chez les vaches laitières (Girard et Matte, 1998). Ils ont découvert que la concentration en folates du sérum sanguin augmentait plus suite à une dose alimentaire de 2 ou de 4 mg/kg de poids vif qu’à une dose de 160 mg sous forme d’injection (Girard et Matte, 1998). De
27 plus, la concentration de folates a augmenté grâce au supplément (P = 0,08). Cependant l’apport de 4 mg d’acide folique par kg de poids vif par jour n’a pas augmenté le taux de folates du lait plus que l’apport de 2 mg. Il est suggéré que la sécrétion de folates dans le lait soit régulée par des protéines qui se lient aux folates. Ainsi, lorsque ces protéines sont saturées, la quantité de folates excrétée dans le lait n’augmente plus (Girard et Matte, 1998).
En accord avec des données antérieures, la production laitière des vaches multipares a été augmentée par l’apport d’un supplément d’acide folique de 4 mg par kg de poids vif (Girard et Matte, 1998). Elles ont produit 2,2 kg/j ou 6 % de plus durant le premier 100 jours de lactation (P = 0,02) et 3 kg/j de plus ou 10 % entre 100 et 200 jours (P = 0,03) (Girard et Matte, 1998). La production laitière n’a pas été modifiée dans les derniers 100 jours de la lactation, contrairement aux résultats de Girard et al. (1994) où ils avaient été augmentés de 45 jours après l’insémination jusqu’au tarissement.
Ces données sont confirmées par une autre étude de Girard et al. (2005), où un supplément alimentaire d’acide folique de 2,6g/j a fait varier la production laitière de 38,0 ± 0,9 kg/jour chez les vaches témoin à 41,4 ± 1,0 kg/jour chez les vaches recevant un supplément.
Contrairement aux données de Girard et al. (1994), la composition du lait n’a pas été modifiée par le supplément (Girard et Matte, 1998). Ainsi, la concentration du lait en protéine n’a pas été augmentée par l’apport d’acide folique (Girard et Matte, 1998). Il est suggéré que, puisque l’acide folique est essentielle à la synthèse de méthionine, l’apport limitant de lysine à partir de laquelle il est possible de produire de la méthionine, dans l’alimentation, aurait limité l’impact de l’acide folique sur la synthèse protéique pour le lait (Girard et Matte, 1998). Puisque la composition du lait n’a pas été modifiée et que la production laitière a augmenté chez les vaches supplémentées, la quantité de solides, de gras, de lactose, de cendres et de caséine produit dans le lait a augmenté, sans qu’il n’y ait une augmentation de la prise alimentaire, ce qui laisse
28
présager une meilleure efficacité d’utilisation, entre autres de l’azote puisque le taux d’azote non protéique du lait est moins élevé chez les vaches supplémentées (Girard et Matte, 1998).
Des résultats plus récents de Girard et al. (2005) démontrent eux aussi qu’il n’y a pas eu de modification de la composition du lait suite à un supplément d’acide folique. Cependant, seulement la quantité de protéines brutes produite et de folates sécrétée dans le lait ont été augmentées significativement malgré une tendance à la hausse pour les solides du lait et le gras (Girard et al., 2005).
Il est intéressant de remarquer que l’impact significatif de l’acide folique sur la production a eu lieu chez les vaches multipares, tant dans les résultats de Girard et Matte (1998), que ceux de Girard et al. (1994). Il est suggéré qu’une interaction avec la vitamine B12, dont la concentration est plus faible dans le sérum de vaches primipares, soit la cause des faibles impacts de l’acide folique sur ces animaux. En effet, la vitamine B12 est essentielle à la participation de l’acide folique dans le cycle de méthylation en tant que cofacteur de l’enzyme méthionine synthase (Girard et Matte, 1998). Les vaches primipares seraient plus affectées par un manque de vitamine B12 puisque leur concentration plasmatique de vitamine B12 sont plus basses. (Girard et Matte, 1998).
1.4.3 - Impact de la vitamine B12 sur le métabolisme et la production laitière Les découvertes antérieures ont poussé l’équipe de Girard à investiguer l’impact d’injections de 10 mg vitamine B12 sur la performances de vaches primipares en début de lactation recevant également des suppléments alimentaires d’acide folique et de méthionine non dégradée dans le rumen (Girard et Matte, 2005a). La vitamine a augmenté le taux d’hémoglobine dans le sang (P = 0,04) et le taux d’érythrocytes (P = 0,06) mais a diminué la concentration d’acide méthylmalonique dans le sérum sanguin
29 (P = 0,02). Le supplément a également augmenté la concentration de vitamine B12 dans le lait des vaches supplémentées (Girard et Matte, 2005; Graulet et al., 2007).
En terme de performances, la production de lait corrigée pour l’énergie des vaches a été de 25,8 kg/d et de 29,0 kg/d pour les vaches contrôle et supplémentées, respectivement (P = 0,03) (Girard et Matte, 2005). Graulet et al. (2007) ont cependant eu des résultats contraires, selon lesquels un supplément de vitamine B12 n’augmentait pas la production laitière, bien que l’acide folique, avec ou sans vitamine B12, augmentait cette production chez des vaches multipares. Il est donc suggéré que la vitamine B12 ne soit pas le facteur faisant varier la production de lait entre les vaches recevant un supplément d’acide folique (Graulet et al., 2007).
La production totale de solides dans le lait, de gras et de lactose a également été augmentée par la vitamine B12, bien que la composition du lait n’ait pas été modifiée, ce qui correspond à des données obtenues avec un supplément d’acide folique chez des vaches multipares ayant un taux de vitamine B12 dans le sérum plus élevé que les primipares non supplémentées (Girard et Matte, 1998; Girard et Matte, 2005).
Il est donc possible de conclure qu’en début de lactation, la quantité de vitamine B12 disponible pour les vaches primipares n’était pas suffisante, ce qui limitait la production laitière de celles-ci (Girard et Matte, 2005). Un apport insuffisant de vitamine B12 peut avoir influencé négativement à la fois le cycle de méthylation, la production d’ADN et l’utilisation du propionate comme source d’énergie (puisque ce sont toutes des réactions nécessitant la vitamine B12 (Girard et Matte, 2005b). Cependant, chez les vaches multipares, il semble que la vitamine B12 ne soit pas le facteur limitant l’action de l’acide folique (Graulet et al., 2007).
Bien que la vitamine B12 ne soit pas le facteur limitant, elle améliore le métabolisme des vaches lorsque administrée conjointement à de l’acide folique (Graulet et al., 2007). En effet, l’augmentation de la production laitière et de la production de protéine laitière
30
occasionnés par un supplément d’acide folique, sans pour autant qu’il y ait une augmentation de la consommation volontaire quotidienne, cause possiblement une mobilisation des réserves corporelles des vaches parce que la glande mammaire a des besoins en nutriments plus importants (Graulet et al., 2007). Un supplément d’acide folique a causé chez les vaches laitières une augmentation des gras totaux, des triacylglycérols et du cholestérol hépatique, tandis que des suppléments de vitamine B9 et B12 conjointement n’ont pas provoqué ces augmentations. La vitamine B12 a possiblement pour effet de réduire les accumulations de méthylmalonyl-CoA, ce qui permet la β-oxydation des acides gras dans le foie et ainsi les molécules d’acétyl-CoA peuvent entrer dans le cycle de Krebs pour fournir de l’énergie à l’animal (Graulet et al., 2007).
Le poids vif, la condition de chair et la consommation volontaire quotidienne n’ont pas été modifiés par le supplément de vitamine B12 seul (Graulet et al., 2007).
Il est donc démontré que, sous certaines circonstances, l’acide folique augmente la production laitière et la production de composantes du lait. La cobalamine, pour sa part, augmente parfois les performances, mais améliore surtout le métabolisme des vaches. Il est donc possible que les veaux de mères supplémentées croissent plus vite puisqu’ils boivent plus de lait. De plus, le lait des vaches supplémentées en acide folique et en vitamine B12 en contient plus.
31
1.5 – Impact des vitamines sur les performances des veaux et
bouvillons
1.5.1 – Impact de l’acide folique sur les performances des veaux en croissance En 1987, Zinn et al. ont supplémenté la diète de veaux avec des vitamines du complexe B, avec entre autres de la vitamine B9 et B12. Les vitamines n’avaient pas influencé le gain ou la conversion alimentaire des veaux (P > 0,20), mais les chercheurs ont trouvé que respectivement seulement 10 % et 3 % de l’acide folique et de la cobalamine se rendaient à l’intestin (Zinn et al., 1987). Ces données correspondent à celles de Girard et al. (1992) qui démontrent qu’il est possible d’offrir une dose d’acide folique plus faible aux pré-ruminants qu’aux ruminants et obtenir la même concentration de folates dans le sérum sanguin (Girard et al., 1992).
Il semble donc favorable d’offrir un supplément alimentaire d’acide folique dans le lait aux veaux pré-ruminants (Girard et al., 1992), si l’acide folique absorbée améliore les performances animales. Une étude de Dumoulin et al. (1991) a démontré que des génisses ayant un rumen fonctionnel, recevant 40 mg d’acide folique par injection intramusculaire ont effectué un gain moyen quotidien de 7,6 % plus élevé que celles recevant une solution saline du sevrage à 5 semaines après le sevrage (P < 0,05). L’acide folique a aussi eu pour effet d’augmenter les concentrations de folates dans le sérum sanguin, la quantité d’hémoglobine et le volume d’érythrocytes (Dumoulin et al., 1991). Cet effet de l’acide folique suggère que l’apport en folates de la diète ne suffit pas à maximiser le statut hématologique de veaux près du sevrage, entre autres parce que la flore ruminale n’est pas assez développée (Dumoulin et al., 1991).
Au contraire, il semble que les porcelets nourris avec du lait comprenant 11,8 ± 0,7 ng/ml de folates ont eu un taux de croissance similaire à des porcelets nourris avec du lait comprenant 7,9 ± 0,4 ng/ml (P > 0,47) (Matte et Girard, 1989). Cependant ces
32
porcelets avaient accès à de la nourriture contenant de l’acide folique, en surplus du lait maternel, ce qui diminue les besoins à combler avec le lait maternel.
1.5.2 - Impact de la vitamine B12 sur la performance de bouvillons
En 1966 Hayes et al. ont découvert que le liquide ruminal de bouvillons nourris avec du foin contenait moins d’acide folique que celui de bouvillons nourris exclusivement avec du maïs. De même, le liquide ruminal des bouvillons nourris avec du maïs contenait plus de vitamine B12 que celui des bouvillons nourris avec du foin haché (Hayes et al., 1966). Ces données sont importantes si l’on considère le mode de finition exclusivement au fourrage comme le bœuf VitaliPré (Pelletier, 2008). Si l’on considère supplémenter la diète de veaux en vitamine B12, il faut aussi prendre en compte que la vitamine B12 n’est pas absorbée en quantités appréciables dans le rumen (Zinn et al., 1987) et qu’il est favorable d’en supplémenter le lait qui n’est pas dégradé dans le rumen plutôt qu’une autre forme de nourriture qui, elle, subit la dégradation microbienne du rumen, ou encore d’augmenter la capacité des bactéries à produire cette vitamine.
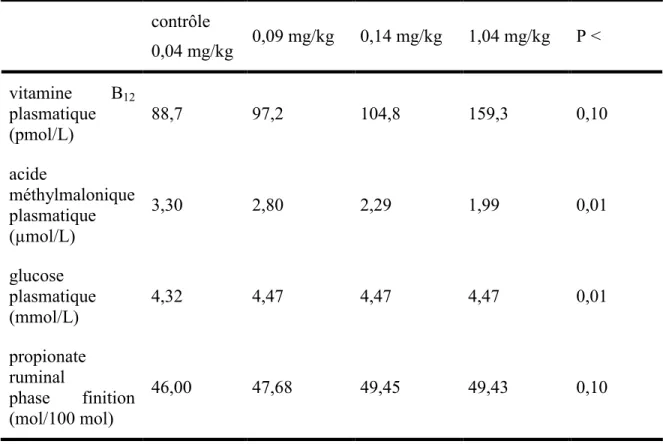
Tiffany et al. ont effectué en 2003 une étude qui compare les performances de bouvillons dont le statut en vitamine B12 est amélioré par une augmentation de la quantité de cobalt dans la ration. Soixante bouvillons de race Angus ont reçus une ration comprenant 0,04, soit le témoin, 0,09, 0,14 ou 1,04 mg/kg de CoCO3 dans la ration. La quantité de cobalt dans la ration n’a pas modifié le gain moyen quotidien dans la phase de croissance. Or, durant la finition, les bouvillons ayant au moins 0,09 mg/kg de cobalt dans la ration ont eu un gain moyen quotidien et un poids final plus élevé (Tableau 1.2) (Tiffany et al., 2003). Une cause possible de ce gain est l’augmentation de la consommation quotidienne d’aliments lorsque la ration contient plus de 0,09 mg/kg de cobalt (Tableau 1.2). Bien que les bouvillons mangent le plus à une concentration de