Application des champignons mycorhiziens à arbuscules dans la culture de la tomate industrielle
132
0
0
Texte intégral
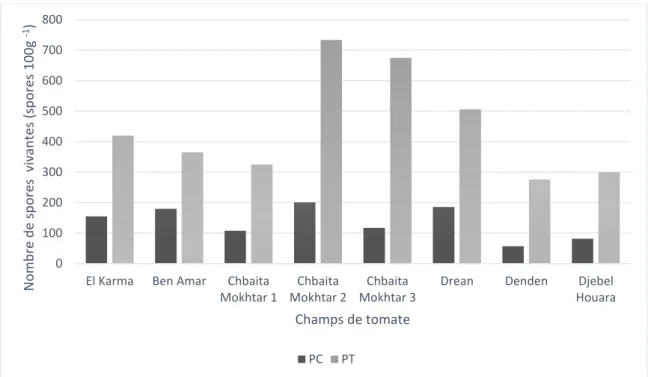
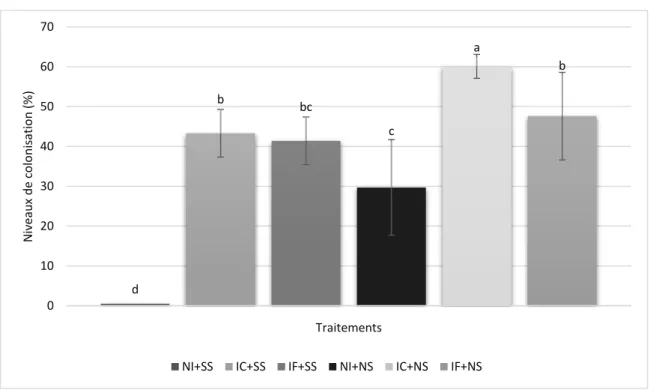
Figure
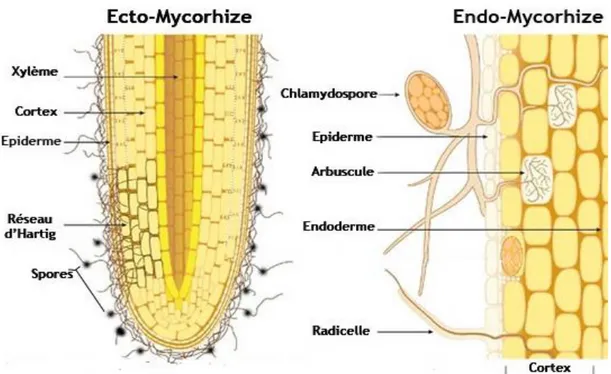
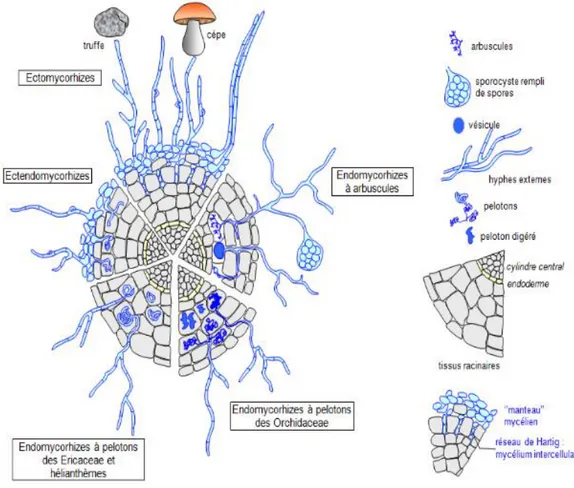
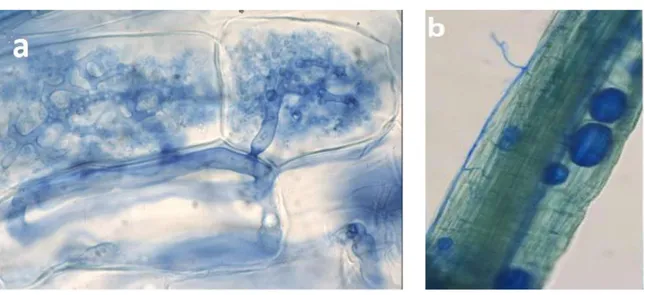
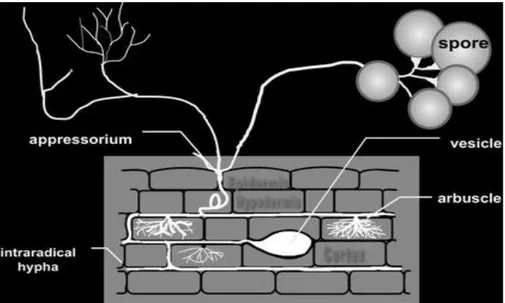
+7
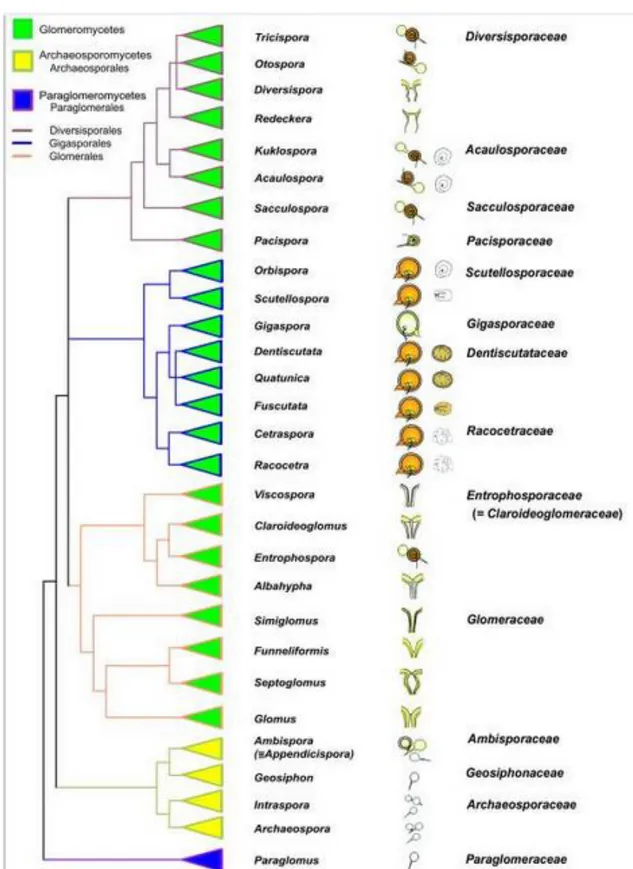
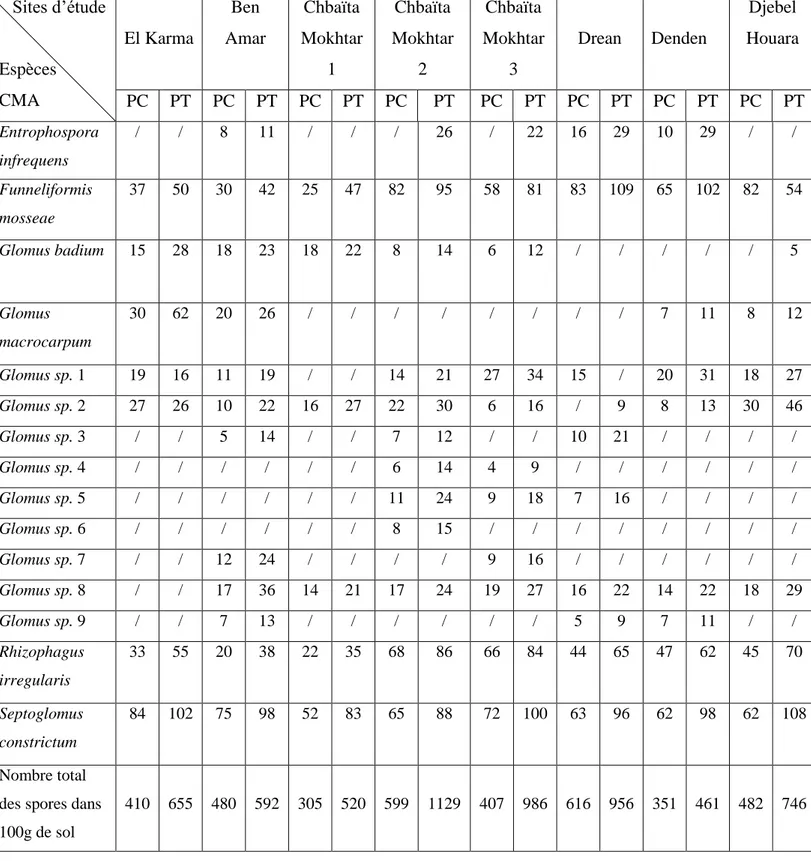
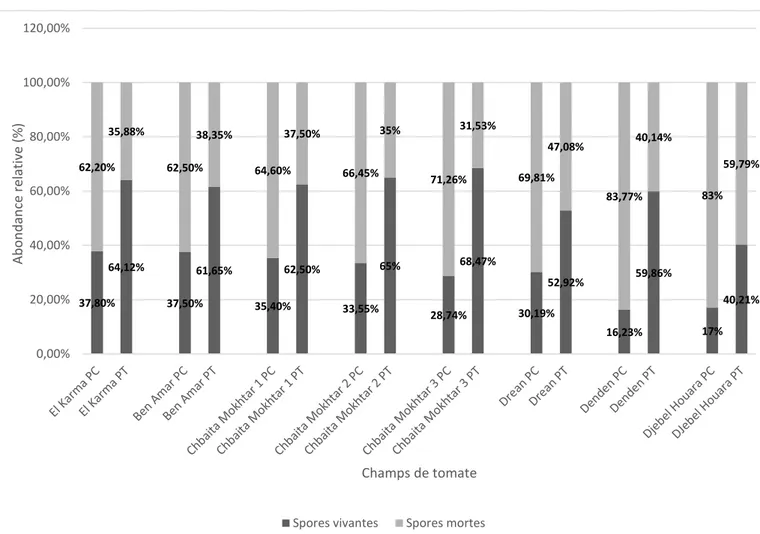
Documents relatifs