Impacts du cuivre et de la restriction alimentaire chez le tête-de-boule.
Texte intégral
Figure




![Figure 3.1 Relation entre la concentration de cuivre mesur6e dans les tissus corporels ([Cu]](https://thumb-eu.123doks.com/thumbv2/123doknet/5404896.126013/68.926.150.772.89.704/figure-relation-concentration-cuivre-mesur-tissus-corporels-cu.webp)




Documents relatifs
Chapitre V : Analyse Du Flambement Mécanique Des Plaques Rectangulaire P-FGM Avec Epaisseur Parabolique Par La Méthode Des Différences Finies.. 72
ايفيظو تجّزدتمنا داىمهن عاىنأ ةّدع ضزدت فىس تعاطتسا ثاذ لاوّدنا لامعتسا عم , تنزاقم تسازد مهنيب نىكتسو تّيّسلاا لاوّدناو تّيبيجنا لاوّدنا. ضزدن
Pour le deuxième objectif, et en profitant des timings listés dans le Tableau 4.2, nous remarquons que la création de composant (configuration générique) pour la deuxième session
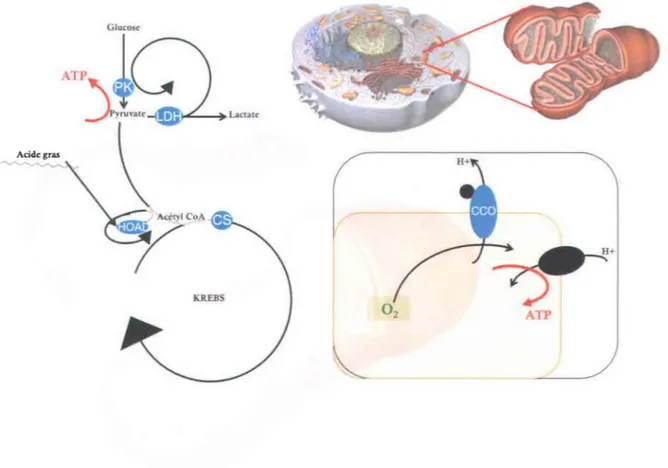
L’exfoliation des cellules épithéliales mammaires chez la vache laitière comme biomarqueur de l’adaptation à la restriction alimentaire. Défis scientifiques Phase, Nov 2019,
The challenge is the classical state space explosion problem: the num- ber of reachable program states grows exponentially with the number of concurrent threads, which renders
Ces donnbes montrent que les liaisons intermolbculaires du type pyridinium jouent un r61e important h la fois dans la sta- bilitb thermique du collagbne et dans
Le but de ce travail est d'examiner les effets d’une restriction alimentaire et de la substitution du concentré par l’ensilage de cactus sur quelques paramètres
Nos agnelles semblent plus résistantes que d’autres races, notamment les brebis de race Awassi chez lesquelles il a été observé des perturbations physiologiques au bout d’un peu
