Extraction of lipids and pigments of Chlorella vulgaris by supercritical carbon dioxide: influence of bead milling on extraction performance
9
0
0
Texte intégral
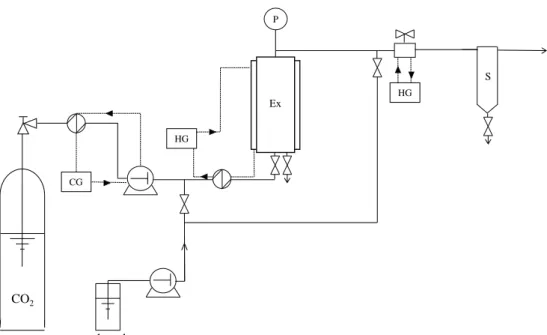
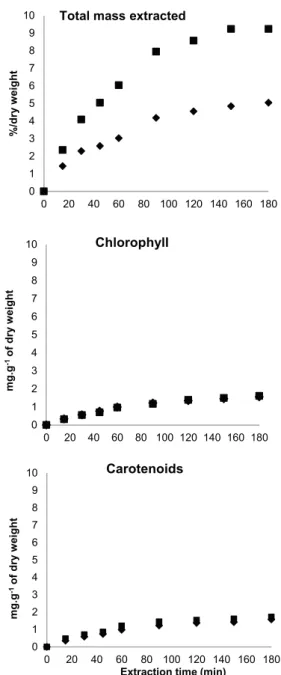
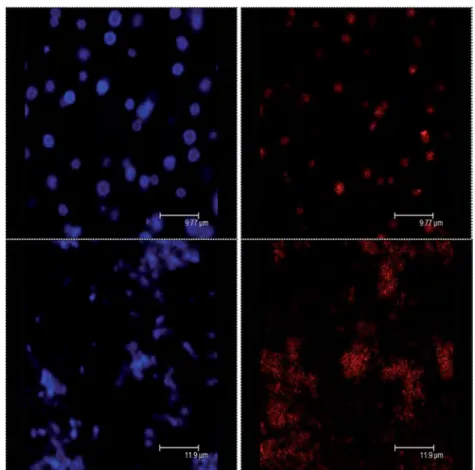
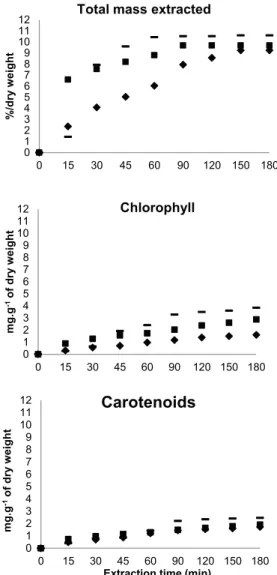
Figure
Documents relatifs
