Impacts de l’urbanisation sur la diversité floristique des
marécages
Mémoire
Léo Janne Paquin
Maîtrise en biologie végétale - avec mémoire
Maître ès sciences (M. Sc.)
Impacts de l’urbanisation sur la diversité
floristique des marécages
Mémoire
Léo Janne Paquin
Sous la direction de :
Monique Poulin, directrice de recherche
Stéphanie Pellerin, codirectrice de recherche
Résumé
L’urbanisation est l’une des causes principales du déclin de la biodiversité à l’échelle mondiale, mais ses conséquences sur les milieux humides urbains n’ont que rarement été étudiées. Nous examinons ici comment la richesse spécifique et la diversité bêta des marécages varient en fonction de l’urbanisation dans l’agglomération de Québec, Canada. Nous avons échantillonné 34 marécages distribués le long d’un gradient d’urbanisation, évalué en fonction de la couverture de canopée dans une zone tampon d’un rayon de 100 m à la périphérie des marécages. Les espèces vasculaires ont été recensées et des variables environnementales furent mesurées dans 92 quadrats. Nous avons testé la variation de la richesse spécifique selon le gradient d’urbanisation en tenant compte du statut des espèces (indigène indicatrice de milieux humides, indigène terrestre, et exotique). La diversité bêta a été calculée comme la similarité de la composition floristique entre les sites et les différences obtenues furent évaluées en utilisant un test de dispersion de l’homogénéité de la variance. Nous avons également effectué un partitionnement la diversité bêta en ses deux composantes (remplacement et différence de richesse) afin d’expliquer la variation de la composition en espèces selon le niveau d’urbanisation. Nos résultats ont montré que la richesse en espèces exotiques augmentait avec l’urbanisation alors que la richesse en espèces indigènes demeurait constante. Nous avons également constaté une plus grande diversité bêta dans les marécages urbanisés et que cette différenciation biotique était principalement causée par le remplacement d’espèces. Ces patrons, combinés à une augmentation des espèces exotiques terrestres avec l’urbanisation, suggèrent un relâchement de la contrainte anoxique au moins dans quelques sites, ce qui a pu contribuer à la diversification biotique à l’échelle régionale. Cependant, ce résultat pourrait n’être que temporaire et refléter les premières étapes de la colonisation des espèces exotiques dans les marécages.
Abstract
Urbanization is one of the main drivers of biodiversity decline worldwide, but this process has seldom been evaluated for urban wetlands. Here, we examine how plant species richness and beta diversity of swamps varied with urbanization in the Quebec City, Canada. We inventoried 34 swamps distributed along an urbanization gradient (forest cover between 25 and 87% in a 100 m radius buffer zone). Vascular species were sampled in 92 plots and abiotic variables as well as proxies (bryophyte abundance, microtopography, and pedologic data) were measured. We tested whether species richness varied along the urbanization gradient and according to the species type (native wetland, native upland or exotic species). Beta diversity was calculated as between-site similarities in composition, and differences were evaluated using tests for homogeneity in multivariate dispersion. We also partitioned beta diversity into its component (turnover and richness difference) for each species type to better understand their response to an increase of urbanization. We found that exotic species richness increased with urbanization while native wetland and upland species richness were similar to each other and remained constant along the same gradient. Yet, exotics were six to 27 times less diversified than native species. We also found a greater beta diversity in urbanized swamps, mostly caused by species turnover. We also observed a decrease of bryophyte abundance and a decrease of microtopography, thus fewer microhabitats, with an intensification of urbanization. These changes, combined with an increase of exotic upland species suggest the release of the anoxia constraint at least in a few sites, which might have favored spontaneous colonization by exotics, and enhance diversification at the regional scale. This result might reflect only the first stages of exotic colonization and thus might be temporary.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des figures ... vi
Liste des tableaux ... vii
Liste des annexes ... viii
Remerciements... x
Avant-propos ... xii
Introduction ... 1
Les mesures de diversité ... 1
Homogénéisation et variation de la diversité bêta ... 2
Les marécages ... 3 Définition ... 3 Hydrologie ... 4 Microtopographie ... 6 Urbanisation ... 6 Fragmentation ... 6 Hydrologie ... 7
Dette d’extinction et patron de diversité bêta... 10
Objectifs et Hypothèses ... 12
Chapter 1 Urbanization fosters differentiation of swamp vegetation ... 13
Résumé ... 14
Abstract ... 15
Introduction ... 16
Methods ... 18
Study area ... 18
Site Selection and Sampling ... 19
Results ... 23
Urbanization and species richness ... 23
Beta Diversity ... 26
Environmental Changes and Species Composition ... 29
Discussion ... 31 Species richness ... 31 Beta diversity ... 32 Environmental factors ... 34 Conclusion ... 35 Conclusion générale ... 36 Apports à la recherche ... 36 Avenues de recherche ... 37 Bibliographie ... 39
Liste des figures
Figure 1: Location map of the 34 sampled swamps (dots) characterized by three
levels of urbanization (low, intermediate and high) in Quebec City, Quebec (Canada), 2016. ... 19
Figure 2: Differences in mean species richness (%) according to urbanization
levels and species status, bars provide standard deviation. Capital letters indicate differences between levels of urbanization within a single species status and lower case letters indicate differences between species status within a single level of urbanization. Contrasts were used to assess significance (p <0.05). ... 26
Figure 3: Influence of urbanization levels on the multivariate dispersion of species
composition in swamps. Urbanization levels were determined in immediate surroundings of swamps in a 100 m radius buffer. Taxonomic beta diversity is measured as the distance of each site to their group centroid using Hellinger distance and represented here on the first two axes of PCoA, circles corresponding to standard deviation. Boxplots show swamp-to-centroid distance (median and quartiles) for each urbanization level. On the PCoA, changes in dispersion around centroids reflect variation in beta diversity; a change of centroid position indicates species turnover. Circles are ellipses of standard deviation Dots colours represent levels of urbanization (white: low, gray: intermediate and black: high). ... 27
Figure 4: Correlation triplot relating five selected environmental variables (arrows),
34 swamps (dots) and 29 species (see code for species in appendix III). Last letter of species name’s indicate their status (W: wetland, U: upland, E: exotic) and only species most fitted to the model are shown. Angle between sites, environmental variables and themselves reflects their correlation. Orthogonal projection of site on an environmental variable approximates its value. Dot colours represent levels of urbanization (white: low, gray: intermediate and black: high). ... 30
Liste des tableaux
Table 1: Effect of urbanization level (low, intermediate, high) and species status
(wetland, upland, exotic) as well as their interactions on mean species richness tested with a linear mixed-effect model and contrasts. Bold indicates significant differences. ... 25
Table 2: Partition of beta diversity (BD) into replacement (turnover between
species) and richness difference (changes in the number of species) at different levels of urbanization (low, intermediate, high) on a) all, b) wetland, c) upland and d) exotic species. Replacement and richness difference are reported in percentages of the total beta diversity value. ... 28
Liste des annexes
Appendix I: Classes of land use in Quebec City and mean cover for each level of
urbanization. ... 56
Appendix II: Principal coordinate analysis on site by land use composition.
Principal component analysis on land-use composition matrix by sites. Arrows represent land use composition and circles arounds site indicate site group. Groups were defined using a non-hierarchical clustering K-means. Dots colour represent level of urbanization (white: low, gray: intermediate and black: high). Land use were photointerpreted in a 100-m buffer zone outside wetlands border. Symbol of arrows: Comm: commercial areas, FWtld: forest and wetland, Hwy: Highway; Hydro: water courses and lakes; Ind : industrial sites; Road: secondary network; Parks: parks and turfs; Resid: residential areas; Wild: wildland. ... 57
Appendix III: Plant species found in the 34 sampled swamps in Quebec City,
Canada. Status refers to wetland (Wet.), upland (Upl.) or exotic (Exo.) species. Species are also sorted in trees, shrubs and herbs and forbs. ... 58
Remerciements
Je remercie d’abord Monique Poulin de son accueil dans son laboratoire et de m’avoir offert ce projet. Nos rencontres furent des plus agréables, merci d’avoir été attentive et disponible, toujours prête à répondre à mes questions et m’encourager dans les moments plus difficiles. Je souhaite également remercier Stéphanie Pellerin, dont les commentaires et la rétroaction rapide furent grandement appréciés. Je veux également remercier Didier Alard pour sa vision en écologie du paysage et ses nombreuses suggestions qui m’ont permis de considérer mon projet sous un angle différent.
Je souhaite souligner la contribution financière de l’Organisme des Bassins Versants de la Capitale et de la ville de Québec qui ont rendu ce projet possible, ainsi que le Centre de la Science de la Biodiversité du Québec pour les ressources statistiques et financières pour la réalisation d’activités essentielles à ma formation.
Je remercie également les étudiants-chercheurs du laboratoire, Milène, Jean-François, Alexandre et Naren pour vos conseils, vos commentaires et votre bonne humeur au quotidien, ainsi que Jérôme qui m’a initiée au monde des SIG. Une mention particulière à Maxime, sans qui l’aventure n’aurait pas été la même ; je te remercie pour nos discussions écologiques enrichissantes, tes conseils et ton aide qui furent inestimables.
Je suis infiniment reconnaissante envers mes assistantes sur le terrain, Agathe, Emma, Geneviève, Marianic et Sandra sans qui la récolte de données sur le terrain aurait été impossible. Votre bonne humeur et votre entrain persistants malgré les maringouins et l’herbe à puce ont su rendre ces étés d’autant plus agréables. Je souhaite également souligner l’importante contribution de Gilles Ayotte et de Geneviève Leblanc pour l’identification de nombreux spécimens, ainsi que Geoffrey Hall et Luc Brouillet pour leur aide plus que précieuse et leur accueil à l’herbier
Marie-Victorin. Un merci du fond du cœur à Luc Brouillet pour m’avoir transmis, sans le savoir, sa passion pour la botanique.
Je tiens à remercier particulièrement Maya Favreau, amie depuis toujours, qui a su répondre à mes questions, d’ordre technique, professionnel et personnel. Je n’aurais pu terminer ce projet sans Romain et Wusllus, avec qui j’ai partagé ma vie durant les quatre dernières années et qui ont su me soutenir au quotidien et dans les moments les plus difficiles, le premier avec ces mots raisonnés et le second avec ses ronronnements. Un énorme merci à mes amis et amies qui ont su me faire rire et me changer les idées.
Finalement, je souhaite remercier ma famille qui m’a toujours grandement soutenue et encouragée à persévérer.
Avant-propos
Ce projet a été réalisé dans cadre d’une collaboration avec la ville de Québec et l’Organisme des bassins versants de la Capitale nationale, dirigé par Monique Poulin de l’Université Laval. Le projet porte sur les impacts de l’urbanisation sur la diversité floristique des marécages de la ville de Québec.
Le chapitre 1 de ce mémoire intitulé « Urbanization fosters differentiation in swamp
vegetation » constitue le corps du présent mémoire et a été rédigé sous la forme
d’un article scientifique. Cet article sera soumis à la revue internationale « Journal
of Urban Ecology ». En tant que première auteure, je fus responsable des
hypothèses de recherche, de l’élaboration du protocole, de la récolte de données à l’été 2016, de la compilation des données, des analyses statistiques et de la rédaction du manuscrit. Les coauteurs ont participé à la planification du projet et à l’élaboration du protocole d’échantillonnage et ont procuré des conseils sur les analyses statistiques et l’interprétation des résultats et ont collaboré à la révision du présent manuscrit.
Les coauteurs sont :
• Monique Poulin, directrice de recherche, professeure titulaire au Département de phytologie de l’Université Laval ;
• Stéphanie Pellerin, codirectrice de recherche, professeure associée au Département de sciences biologiques de l’Université de Montréal ;
• Didier Alard, collaborateur, professeur titulaire à l’Université de Bordeaux affilié à l’INRA en France.
Introduction
La population mondiale augmente et se concentre davantage dans les villes. En 2014, 54 % des humains vivaient en zone urbaine et on estime que cette proportion atteindra 66 % en 2050 (MEA, 2005). Afin de répondre aux besoins des populations et à la croissance économique, les zones urbaines et périurbaines s’étendent pour permettre la construction d’infrastructures et le développement industriel et agricole. L’étalement urbain se produit au détriment des milieux naturels présents qui se retrouvent fragmentés dans la matrice urbaine. Les dynamiques naturelles régulant l’assemblage des communautés végétales sont altérées, entraînant des changements dans leur structure et leur composition ainsi que l’extinction locale d’espèces (p. ex. Galatowitsch et Van Der Valks, 1996 ; Burton et al., 2009 ; Moffatt
et al., 2004). Les milieux humides ont été particulièrement touchés par la destruction
des habitats. En effet, il est évalué que plus de 87 % des milieux humides ont été détruits à l’échelle mondiale (MEA, 2005 ; Davidson, 2014) ; les milieux restants sont soumis aux perturbations anthropiques et à l’invasion des espèces exotiques (Qian et Gio, 2010). Cependant, une meilleure compréhension de la réponse des habitats à l’urbanisation permettrait de mieux protéger la faune et la flore qui y habitent.
Les mesures de diversité
La diversité alpha représente la diversité en espèces d’un site. Elle peut être exprimée en un nombre d’espèces ou avec un indice (p. ex. Shannon-Wiener, Simpson). La diversité gamma correspond au pool total d’espèces d’une région donnée, souvent l’aire d’étude. Elle peut être calculée avec les mêmes métriques que la diversité alpha (Legendre et al., 2005). Entre ces deux niveaux se trouve la diversité bêta qui correspond à la variation de la composition en espèces entre les sites de l’aire d’étude (Whittaker, 1960, 1972 ; Legendre et al., 2005). Ce niveau de diversité est décomposable en deux éléments, soit la différence de richesse (« richness difference ») et le remplacement (« turnover » ou « replacement »). Le remplacement correspond au nombre d’espèces qui se succèdent le long d’un gradient écologique, temporel ou spatial, résultant des processus d’introduction et
d’extinction. La différence de richesse réfère à la variation du nombre d’espèces entre les sites et peut être attribuée à la variation du nombre de niches écologiques d’un site à l’autre (Legendre, 2014). Lorsque les espèces présentes dans un site plus pauvre sont un sous-ensemble exact des espèces présentes d’un site plus riche, il est alors question d’emboîtement (« nestedness ») (Legendre, 2014).
Homogénéisation et variation de la diversité bêta
L’homogénéisation biotique correspond à une augmentation de la similarité des communautés entre les sites, ou plus précisément une diminution de la diversité bêta. L’homogénéisation biotique affecte tous les groupes taxonomiques en plus d’être une cause importante du déclin de la biodiversité à l’échelle mondiale (McKinney, 2006, 2008 ; Baiser et al., 2012). Les environnements urbains et périurbains imposent des pressions environnementales différentes de la matrice naturelle initialement présente et favorisent notamment les espèces synanthropiques (McKinney, 2006 ; 2008). La faune et la flore des villes sont alors davantage similaires entre elles à travers le monde (McKinney, 2002). De plus, l’affaiblissement des barrières biogéographiques par les activités humaines facilite le déplacement d’espèces et favorise leur propagation au-delà de leur zone de répartition naturelle (La Sorte et al., 2007).
L’homogénéisation biotique a auparavant été largement attribuée aux espèces exotiques (McKinney et Lockwood, 1999). Toutefois, certaines études récentes ont souligné la futilité de la distinction exotique/indigène lorsqu’on considère l’homogénéisation de la flore (Mayfield, 2010 ; Brice et al., 2017) puisque les espèces responsables de l’homogénéisation répondent à une série de critères (reproduction rapide, tolérance à la perturbation et à des milieux eutrophes) sans égard à leur origine. Ainsi, plusieurs études ont montré que la diminution de la diversité bêta pouvait également être causée par un gain en espèces indigènes (McCune et Vellend, 2013 ; Beauvais et al., 2016 ; Naaf et Wulf, 2010). Par exemple, McCune et Vellend (2013) ont observé la disparition d’espèces spécialistes, ayant
une niche écologique restreinte et étant peu tolérantes à la perturbation au profit d’espèces « généralistes » occupant des niches plus larges.
Certaines études affirment que le rôle des espèces exotiques dans le changement de la diversité bêta serait lié à leur temps de résidence et à leur répartition. À l’échelle de l’Europe de l’Ouest, les archéophytes désignent les espèces dont l’introduction est antérieure à l’an 1500. Les archéophytes favorisent une homogénéisation biotique, de façon similaire aux espèces généralistes indigènes, puisque les archéophytes ont atteint leur aire maximale de répartition. À l’inverse, les néophytes (dont l’introduction est postérieure à l’an 1500) contribuent généralement à différencier les flores puisqu’elles n’ont pas atteint leur aire de répartition maximale (Kühn et Klotz, 2006 ; Lososová et al., 2016). En effet, si deux espèces différentes s’introduisent dans autant de sites, elles causent une augmentation de la diversité bêta ; à l’inverse, si une même espèce est introduite dans deux sites, elle augmente la similarité et contribue à l’homogénéisation (Olden et Rooney, 2006). Au Québec, puisque la quasi-totalité des espèces a été introduite après 1500 (Lavoie et al., 2003 ; Lavoie et al., 2007), elles peuvent donc être considérées comme des néophytes.
Les marécages
Définition
Au cours des 30 dernières années, plusieurs définitions et systèmes de classification des milieux humides ont été proposés (pour un historique, voir Ménard et al. (2007)). La définition retenue par le Canada est celle du Groupe de travail national des terres humides (GTNTH, 1997) qui caractérise la terre humide comme une « terre saturée
d’eau pendant une période assez longue pour que naissent les processus de terre humide ou aquatique, qui se caractérisent par un faible drainage des sols, des hydrophytes et différentes sortes d’activités biologiques adaptées aux milieux humides ». Cette définition reprend les trois caractéristiques du milieu humide :
classification se divise en cinq grandes classes qui séparent les différents milieux humides selon leur physionomie et leurs caractéristiques hydrologiques (Zoltai et Vitt, 1995). La classe d’intérêt dans ce mémoire est celle du marécage qui se définit comme « une terre humide dominée par les arbres ou grands arbustes (aussi
appelés taillis) et influencée par l’eau souterraine minérotrophe, sur des sols minéraux ou organiques » (Adams et al., 1997). Selon cette classification, les
marécages incluent également les tourbières boisées. La végétation est dominée par des arbres ou de grands arbustes qui forment un couvert minimal de 30 %. Le pH du marécage varie largement ; on retrouvera des milieux oligotrophes (pH inférieurs à 4,5) et des milieux eutrophes (pH de plus de 7,0).
La position des milieux humides par rapport aux éléments hydrographiques du paysage permet une distinction supplémentaire : les milieux riverains et les milieux isolés. Le terme isolé réfère à l’absence de lien hydrologique en surface avec un élément hydrographique. Autrement dit, un milieu humide dit isolé est enclavé dans un milieu mésique (Tiner, 2003 ; Tiner et Lebowitz, 2005). Cette notion présente des aspects intéressants au point de vue écologique puisque les dynamiques et processus des régimes sont propres aux milieux isolés et riverains.
Hydrologie
L’hydrologie est un élément clé dans la formation des marécages. L’hydropériode, la variation des niveaux d’eau au cours d’une saison est très variable entre les marécages. Par exemple, dans les « Cypress swamps », des bogs forestiers bombés dominés par les cyprès présents dans le sud-est des États-Unis, il est possible d’observer de l’eau libre durant presque toute l’année, puisque la nappe phréatique est plus élevée que le niveau du sol (Mitch et Gosselin, 2015 ; Ehrenfeld et Schneider, 1991), alors que les « bottoms land hardwood », des marécages situés dans la plaine inondable de la même région, peuvent être inondés seulement lors des crues printanières et avoir une nappe phréatique très basse pour le reste de l’année (Hodges, 1997 ; Mitch et Gosselink, 2015). Cette baisse des niveaux d’eau
est due à une diminution des précipitations ainsi qu’à l’évapotranspiration de la végétation (Mitch et Gosselink, 2015).
La nappe phréatique des marécages est très variable et sa hauteur change au cours de la saison ainsi qu’entre les années (Zoltai et Vitt, 1995 ; Golet et al., 1993). Selon les travaux de Lowry (1984) effectués dans des érablières rouges au Massachusetts, la nappe phréatique est affleurante lors des saisons de croissance particulièrement humides et peut descendre à plus d’un mètre de profondeur dans les années les plus sèches, alors qu’elle se trouve à environ 20 cm sous la surface lors des saisons normales. Cette profondeur atteint une moyenne d’un mètre après la crue printanière dans les marécages canadiens (Mitch et Gosselink, 2015).
Les milieux riverains
Les marécages riverains sont à la fois dépendants de l’hydrologie de la rivière, mais aussi de la géomorphologie de l’écosystème riverain, c’est-à-dire des conditions géologiques, topographiques et édaphiques du bassin versant ainsi que de son climat et de sa végétation (Naiman et al., 2005 ; Poff et al., 2007). Le débit, la fréquence, la durée, la prévisibilité et l’instantanéité des crues agissent comme filtres abiotiques et déterminent la composition des grandes communautés le long du continuum riverain (Poff et al., 2007). La crue joue un rôle sur la succession des communautés en permettant à la fois le transport de propagules de l’amont vers l’aval et le maintien d’un cycle de perturbation régulateur dans les communautés déjà établies (Nilsson et Svedmark, 2002 ; Nilsson et al., 2010). La nappe phréatique des marécages riverains est suffisamment influente durant la saison de croissance pour permettre le développement d’une végétation hydrophile et d’un sol hydromorphe (Nilsson et Svedmark, 2002 ; Groffman et al., 2003 ; Mitch et Gosselink, 2015). Toutefois, à l’étiage, le faible niveau d’eau cause un abaissement de la nappe phréatique entraînant un stress hydrique. Les espèces présentes doivent donc résister à la crue printanière et aux crues sporadiques au cours de la saison de croissance, ainsi qu’à un stress hydrique à l’étiage (Mitch et Gosselink, 2015).
Les milieux isolés
Les marécages « isolés » sont enclavés localement par un milieu mésique ou un habitat caractérisé par un sol non hydromorphe (Tiner, 2003). Cette définition est toutefois loin d’être parfaite puisque l’isolement géographique des milieux humides dans le paysage n’implique pas que ces derniers soient fonctionnellement isolés (Leibowitz, 2003 ; Tiner, 2003 ; Mushet et al., 2015 ; Cohen et al., 2016). De plus, la distance d’isolement minimale entre les milieux humides ou hydriques varie entre les études (Mushet et al., 2015). Bien que le terme ne soit pas idéal, il est utilisé faute de meilleure nomenclature. Dans le cadre du présent travail, les milieux entourés d’une matrice de milieux mésiques ou urbains et qui ne sont pas caractérisés par des eaux de surfaces permanentes sont considérés comme isolés (Fossey et Rousseau, 2016).
Microtopographie
La microtopographie contribue à la richesse spécifique importante des marécages (Brunland et Richardson, 2005 ; Alsfeld et al., 2009). Résultant des perturbations naturelles, par exemple les chablis, la succession de buttes et de creux génère un gradient d’humidité, de température et de texture du sol, offrant plusieurs niches écologiques. La microtopographie favorise également une hétérogénéité dans la fonte de la neige au printemps et l’accumulation d’eau lors des pluies (Valtera et Schaetzl, 2017). Cette microtopographie permet la coexistence de nombreuses espèces ; les buttes favorisent les espèces mésiques et ligneuses alors que les espèces adaptées à l’anoxie croissent dans les dépressions (Paratley et Fahey, 1986 ; Ehrenfeld, 1995 ; Courtwright et Findlay, 2011).
Urbanisation
Fragmentation
L’étalement des milieux urbains et périurbains cause une conversion de l’usage des terres, ce qui entraîne l’isolement des milieux naturels dans la matrice urbaine. Cette fragmentation des habitats altère les conditions abiotiques des milieux naturels
(Ehrenfeld et al., 2003 ; Faulkner, 2004) suite à l’eutrophisation des cours d’eau, le changement des régimes hydrologiques et l’augmentation de la luminosité et de la température (Pickett et al., 2001 ; Hogan et Waldbridge, 2009 ; Kowarik, 2011 ; Kaushal et al., 2014). La fragmentation modifie également les régimes de perturbations naturelles comme les cycles de feu (Stevens et al., 2016) et les inondations (Poff et al., 2007 ; Brice et al., 2017). De plus, l’isolement des populations affecte les patrons de colonisation et d’extinction au sein des fragments de végétation restants (Aguilar et al., 2006) et la variabilité génétique au sein des populations (Honnay et Jacquemyn, 2007). Suite à l’enclavement dans la matrice, on note des événements d’extinction de population d’espèces spécialistes et une colonisation par des espèces généralistes (Laurance et al., 2012 ; Vellend et al., 2017).
Les changements dans la matrice de paysage suite à l’étalement urbain permettent à de nouvelles espèces de s’établir dans les fragments isolés. L’effet de lisière, causé par les changements environnementaux de la matrice urbaine, notamment l’abondance de lumière au sol, permet à plusieurs espèces synanthropiques de s’installer à la marge des fragments sur une distance pouvant dépasser 30 mètres (Gurevitch et al., 2008, Hamberg et al., 2009 ; Vallet et al., 2010). La fréquentation des sites cause une destruction la végétation sous l’action du piétinement et entraîne la création de sentiers. Ces sentiers génèrent un effet de lisière interne qui facilite la dissémination des espèces synanthropiques vers l’intérieur du site et en altèrent la composition (Cutway et Ehrenfeld, 2009, 2010 ; Bergeron et Pellerin, 2014). Dans les marécages cependant, l’anoxie du milieu pourrait limiter la propagation de ces espèces au-delà de la zone du sentier (Ehrenfeld, 2008).
Hydrologie
L’imperméabilisation du sol modifie les régimes hydrologiques des milieux humides. À l’échelle du bassin versant, l’imperméabilisation des surfaces réduit l’infiltration d’eau dans le sol et diminue l’évaporation, ce qui entraîne généralement une diminution (Grischek et al., 1996 ; Garcia-Fresca, 2007) de la nappe phréatique
régionale et du débit de base des rivières. Dans les régions plus chaudes (p. ex. Australie (Barron et al., 2013)), l’imperméabilisation des sols favoriserait un rehaussement de la nappe phréatique dû à une diminution de l’évapotranspiration. En l’absence d’études spécifiques, il est cependant difficile de prédire l’ampleur des changements au niveau de la nappe phréatique d’une région donnée, suite à l’urbanisation (Bhaskar et al., 2016). Plusieurs conditions naturelles telles que le climat et la quantité de précipitations, la topographie, le type de sol (lié à l’infiltration et au ruissellement de surface) ainsi que le type et la densité de végétation (lié à l’évapotranspiration) déterminent le régime hydrologique local (Bhaskar et al., 2016). Les facteurs anthropiques comme la densité de la couverture des terres et la proportion de végétation, les méthodes de gestion des eaux pluviales et la vétusté des réseaux d’aqueducs interagissent avec les conditions naturelles pour influencer le comportement de la nappe phréatique (Price, 2011 ; Hamel et al., 2013 ; Bhaskar
et al., 2016).
Un changement des niveaux d’eau dans les marécages modifie la composition des communautés végétales. Une augmentation trop importante des niveaux d’eau cause la mort des arbres composant la canopée (Kingsford, 2000). À l’inverse, une diminution de la contrainte anoxique favorise la progression de la végétation terrestre dans le milieu (Ehrenfeld et Schneider, 1993 ; Ward et Stanford 1995 ; Toner et Keddy, 1997). Plusieurs études effectuées sur la végétation riveraine rapportent un déclin des espèces indigènes et une augmentation des espèces exotiques et rudérales suite à la modification du chenal de la rivière et du contrôle des niveaux d’eau (Merritt et Cooper, 2000 ; Naiman et al., 2005 ; Brice et al., 2017). Par exemple, à l’échelle des États-Unis le contrôle des débits des rivières harnachées masque les variabilités naturelles en homogénéisant les conditions environnementales, facilitant l’implantation d’espèces exotiques et cosmopolites (Nilsson et Svedmark, 2002 ; Poff et al., 2007). Il est difficile de déterminer si les milieux humides enclavés dans la matrice urbaine recevront plus ou moins d’eau puisque chaque site est soumis à un ensemble de facteurs environnementaux très locaux (Ehrenfeld et al., 2003). Toutefois, l’urbanisation est généralement associée
à des sites dont les niveaux d’eau sont plus variables au cours d’une même saison (Booth, 1991 ; Stander et Ehrenfeld, 2009 ; Bhaskar et al., 2016).
Dans les milieux riverains, l’imperméabilisation des terres favorise l’incision du chenal. Lors d’un épisode de pluie, les surfaces imperméables accélèrent et concentrent les écoulements de surface dans les cours d’eau urbains, causant des crues soudaines et rapides qui contribuent à une érosion des berges et du lit de la rivière (Groffman et al., 2003). L’incision de chenal combinée à un débit de base plus faible du cours d’eau entraînent un abaissement de la nappe phréatique dans le milieu adjacent (Walsh et al., 2005 ; Hardison et al., 2009). Les activités anthropiques modifient non seulement les caractères physico-chimiques et la quantité de nutriments disponibles (Paul et Meyer, 2001 ; Hogan et Waldbridge, 2007), mais également la connectivité amont-aval en altérant les patrons de dispersion des espèces. Ceci modifie la composition des communautés végétales (Naiman et al., 2008 ; Aronson et al., 2017). Par exemple, la présence de barrages sur les rivières diminue la hauteur des niveaux d’eau atteints lors des périodes de grandes crues. Or, ces crues correspondent au moment de dispersion des propagules hydrochores (Poff et al., 2007 ; Merritt et Wohl, 2002). On observe une diminution du nombre de propagules puisqu’elles sont retenues par le barrage, ainsi qu’une diminution de la dispersion des propagules dans les milieux adjacents due à des niveaux d’eau plus bas (Nilsson et al., 2010).
Dans les milieux humides isolés, quelques auteurs ont rapporté que, suite à l’urbanisation, l’apport en eau se faisait principalement par le ruissellement des eaux de surface (Owen, 1995 ; Barksdale et al., 2014 ; Bhaskar et al., 2016). Il est cependant difficile de conclure la direction des écoulements de surface sans une cartographie détaillée de la microtopographie des terres adjacentes à chaque milieu humide.
Dette d’extinction et patron de diversité bêta
Dans les îlots naturels, la réponse de la végétation à la destruction de la matrice naturelle adjacente n’est pas immédiate (Tilman et al., 1994 ; Jackson et Sax, 2010). En effet, les communautés n’atteignent l’équilibre (l’état de « relaxation », selon Diamond, (1972)) qu’après plusieurs années. Même lorsque les perturbations sont
in situ, c’est-à-dire dans le site à l’étude, le délai peut être marqué. En comparant
deux forêts soumises à des perturbations anthropiques in-situ similaires, Vellend et
al. (2006) ont estimé à près d’un siècle le délai nécessaire pour la disparition
d’espèces qui étaient présentes avant le début de la dégradation. Le délai n’est pas le même pour toutes les espèces : celles qui ont un temps de colonisation et de disparition rapide réagissent plus rapidement, contrairement à celles ayant un taux plus lent (Vellend, 2004). Ces dernières produisent également souvent une faible quantité de graines par individu, et sont caractérisées par un âge de reproduction tardif et une courte distance de dispersion (Verheyen et al., 2004 ; Vellend et al., 2006). Les espèces longévives sont également plus résilientes puisqu’elles se maintiennent dans le milieu malgré un échec reproducteur (Jackson et Sax, 2010). Dans les communautés qui ne sont pas à l’équilibre, il est possible d’observer la superposition de plusieurs cortèges floristiques. Certaines des espèces présentes initialement qui sont appelées à disparaître localement cohabitent avec de nouvelles espèces favorisées par le changement des conditions du milieu suite à l’urbanisation.
Cette superposition de cortèges floristiques peut se traduire par une augmentation de la diversité bêta due à l’introduction d’espèces différentes selon les sites, causée par l’hétérogénéité de la matrice urbaine et des contraintes environnementales divergentes (Ramalho et Hobbs, 2012 ; Godefroid et Koedam, 2007) et par la pression de diaspores (Pickett et al., 2001 ; Lockwood et al., 2005). Ceci a pour conséquence de présenter, à première vue, un phénomène de diversification, mais, puisque ces espèces sont synanthropiques ou généralistes, on observe une diminution de la diversité de la flore à l’échelle régionale puisque les fragments gagnent en homogénéité avec les espèces des milieux adjacents (Brice et al., 2017).
Objectifs et Hypothèses
L’objectif de ce projet de maîtrise est de décrire les conséquences de l’urbanisation sur la composition floristique des marécages dans la région de Québec. Plus précisément, les questions suivantes ont été adressées :
1) L’urbanisation est-elle associée à une augmentation ou une diminution de la diversité bêta à l’échelle régionale ?
2) Quelle est la réponse spécifique des espèces indicatrices de milieux humides, terrestres et exotiques ?
L’hypothèse générale est que l’urbanisation entraîne un relâchement des principales contraintes environnementales des marécages, notamment la contrainte anoxique, modifiant ainsi les communautés végétales des sites enclavés dans la matrice urbaine. Plus spécifiquement :
1) La complexité des éléments du paysage urbain favorise l’introduction d’espèces différentes entre les sites causant une augmentation de la diversité bêta.
2) L’augmentation de l’intensité de l’urbanisation favorisera une plus grande richesse spécifique des espèces exotiques dans les sites plus perturbés. Ces espèces s’implanteront notamment au détriment des espèces de milieux humides dues à une perturbation des cycles hydrologiques naturels.
Chapter 1 Urbanization fosters differentiation of swamp
Résumé
L’urbanisation est l’une des causes principales du déclin de la biodiversité à l’échelle mondiale, mais ses conséquences sur les milieux humides urbains n’ont que rarement été étudiées. Nous examinons ici comment la richesse spécifique et la diversité bêta des marécages varient en fonction de l’urbanisation dans l’agglomération de Québec, Canada. Nous avons échantillonné 34 marécages distribués le long d’un gradient d’urbanisation, évalué en fonction de la couverture de canopée dans une zone tampon d’un rayon de 100 m à la périphérie des marécages. Les espèces vasculaires ont été recensées et des variables environnementales furent mesurées dans 92 quadrats. Nous avons testé la variation de la richesse spécifique selon le gradient d’urbanisation en tenant compte du statut des espèces (indigène indicatrice de milieux humides, indigène terrestre, et exotique). La diversité bêta a été calculée comme la similarité de la composition floristique entre les sites et les différences obtenues furent évaluées en utilisant un test de dispersion de l’homogénéité de la variance. Nous avons également effectué un partitionnement la diversité bêta en ses deux composantes (remplacement et différence de richesse) afin d’expliquer la variation de la composition en espèces selon le niveau d’urbanisation. Nos résultats ont montré que la richesse en espèces exotiques augmentait avec l’urbanisation alors que la richesse en espèces indigènes demeurait constante. Nous avons également constaté une plus grande diversité bêta dans les marécages urbanisés et que cette différenciation biotique était principalement causée par le remplacement d’espèces. Ces patrons, combinés à une augmentation des espèces exotiques terrestres avec l’urbanisation, suggèrent un relâchement de la contrainte anoxique au moins dans quelques sites, ce qui a pu contribuer à la diversification biotique à l’échelle régionale. Cependant, ce résultat pourrait n’être que temporaire et refléter les premières étapes de la colonisation des espèces exotiques dans les marécages.
Abstract
Urbanization is one of the main drivers of biodiversity decline worldwide, but this process has seldom been evaluated for urban wetlands. Here, we examine how plant species richness and beta diversity of swamps varied with urbanization in the Quebec City, Canada. We inventoried 34 swamps distributed along an urbanization gradient (forest cover between 25 and 87% in a 100 m radius buffer zone). Vascular species were sampled in 92 plots and abiotic variables as well as proxies (bryophyte abundance, microtopography, and pedologic data) were measured. We tested whether species richness varied along the urbanization gradient and according to the species type (native wetland, native upland or exotic species). Beta diversity was calculated as between-site similarities in composition, and differences were evaluated using tests for homogeneity in multivariate dispersion. We also partitioned beta diversity into its component (turnover and richness difference) for each species type to better understand their response to an increase of urbanization. We found that exotic species richness increased with urbanization while native wetland and upland species richness were similar to each other and remained constant along the same gradient. Yet, exotics were six to 27 times less diversified than native species. We also found a greater beta diversity in urbanized swamps, mostly caused by species turnover. We also observed a decrease of bryophyte abundance and a decrease of microtopography, thus fewer microhabitats, with an intensification of urbanization. These changes, combined with an increase of exotic upland species suggest the release of the anoxia constraint at least in a few sites, which might have favored spontaneous colonization by exotics, and enhance diversification at the regional scale. This result might reflect only the first stages of exotic colonization and thus might be temporary.
Introduction
Urbanization is one of the main drivers of biodiversity change worldwide (e.g., Chapin et al., 2000; Thuiller, 2007). At broad spatial scale, urbanization has been mostly associated with biotic homogenization (BH) which corresponds to an increase in similarity of species composition (i.e., decrease of beta diversity) over time or across sites (McKinney and Lockwood, 1999; Olden and Rooney, 2006; McKinney, 2006; La Sorte et al., 2014). This loss of beta diversity is often caused by widespread generalist or exotic species replacing local specialist ones (McKinney and Lockwood, 1999; La Sorte et al., 2007). However, despite the attention that the concept of BH has received in the last decade, uncertainties persist toward the underlying causal factors as well as the plurality of outcomes related to this process (Olden and Poff, 2003; Olden and Rooney, 2006). For example, even though a decline in beta diversity has been largely associated with an increase of exotic species following land-use changes, BH has also been attributed to the spread of native species (Tabarelli et al., 2012; McCune and Vellend, 2013; Beauvais et al., 2016). In addition, differentiation rather than homogenization can occur with urbanization (McKinney, 2008), depending on the size and composition of the initial species pool (Olden and Poff, 2003), the balance between native vs. non-native species and their residential time (Kühn and Klotz, 2006; Lososová et al., 2012; 2016) and the intensity and type of urbanization (Flynn et al., 2009; Allan et al., 2015, Newbold et al., 2015). It follows that urbanization is a complex process (Grimm et al., 2008) that can accentuate or buffer environmental constraints prevailing in urban ecosystems (Pennington et al., 2010; Brice et al., 2017) and gaps remain in our knowledge of the mechanisms behind homogenization or differentiation of urban ecosystem remnants, such as grasslands, forest or wetlands.
Wetlands are receiving increasing attention due to their great supply of ecosystem services (MEA, 2005; Maltby and Acreman, 2011), notably in urban landscapes where they are recognized as critical green infrastructure for flood control, water purification, aesthetics, cooling effect and recreation (Taha, 1997; Bolund and Hunhammar, 1999; Lee and Scholz, 2006; Sun et al., 2012; McLaughlin and Cohen,
2013). Understanding wetland response to urbanization is mandatory to develop management plans that will make it possible to maintain the integrity of their species composition and the sustainability of their ecosystem services. Wetlands might be prone to homogenization following urbanization as wetland plants usually spread more widely than those associated with mesic forests (Santamaría, 2002; Ricklefs et al., 2008). Wetlands are also highly susceptible to invasion by exotic species (Galatowitsch et al., 1999; Zedler and Kercher, 2004; Larson et al., 2016), although studies mostly focused on riparian and herbaceous dominated wetlands. On the other hand, differentiation (increase of beta diversity) following environmental changes and subsequent species establishments have been shown in wetland communities (Ehrenfeld and Schneider, 1991; Favreau et al., 2019).
Compared to mesic forests or other wetlands, forested wetlands such as swamps show the particularity of being simultaneously constrained by both low light availability on the ground and periodic anaerobic soil conditions during the growing season (Conner et al., 1981; Mitsch and Gosselink, 2000; Colmer and Voesenek, 2009), which lead to a strong selection pressure for species establishment (Battaglia et al., 2000; Lin et al., 2004). Land use changes arising from urban sprawl may alter stress regime in swamps, notably due to rain collectors trapping surface run-off, thereafter impeding groundwater recharge in swamps and releasing the anoxia stress constraint (Azous and Horner, 1997; Kentula et al., 2004). This may cause shifts of ecological conditions, allowing exogenous species, either native generalists or exotics, to disperse from surrounding heterogeneous anthropic lands and establish themselves in such urban swamps (Ehrenfeld and Schneider, 1991; Findlay and Bourdages, 2000; Ehrenfeld, 2008; Cutway and Ehrenfeld, 2009). For example, urbanization has been associated with a decrease of flooding period and duration in riparian forests, a stress release process that led to a greater variation in species composition across sites (Brice et al., 2017). Following this process, we may assist to biotic differentiation rather than homogenization in such systems. It remains that studies on the impact of urbanization on swamps are scarce and most were conducted on sites greatly variable in size (between less than 1 ha to 440 ha), which
makes it hard to draw clear conclusions on the impact of urbanization on forested wetland flora.
In this study, we examine how plant species richness and beta diversity of swamps varies with urbanization in the Quebec City metropolitan area, Canada. More precisely, we addressed the following questions: (i) Is urbanization associated with biotic homogenization or differentiation in swamp flora? (ii) How urbanization affects species composition of swamps? (iii) What is the specific response of wetland and upland species as well as exotics to different urbanization levels? Because we expected stress release after urbanization, we hypothesized a biotic differentiation related to urbanization intensity. We predicted an increased beta diversity and a divergence in species composition between swamps surrounded by urbanized landscapes compared to less disturbed ones. We also anticipated urbanization to foster upland and exotic species richness.
Methods
Study area
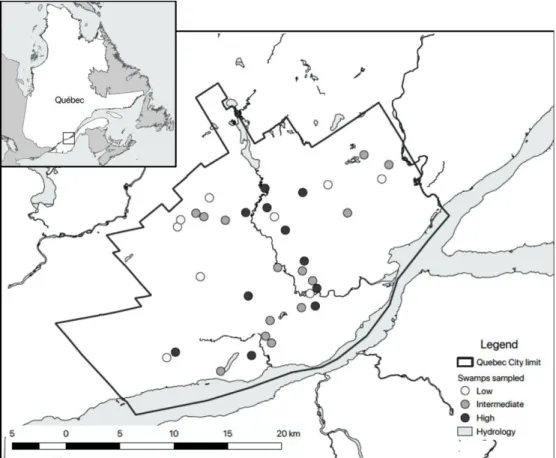
The study was conducted during the summer of 2016 in the Quebec City metropolitan area (Figure 1, hereafter referred to as Quebec City), the seventh most populous metropolitan region of Canada (548 km2, 569 717 inhabitants; Statistics Canada, 2016). Although built-up areas increased by 79% in the last 35 years in this region (Nazarnia et al., 2016), remnants of natural habitats still represent half of the territory while agricultural lands represent 11% and built-up surfaces 39% of the landscape. Built-up surfaces can be further divided into residential (24%), industrial/commercial (5.5%), road network and mining areas (5%) with the remaining space occupied by vacant lots (4.5%) (Cimon-Morin and Poulin, 2018). Wetlands represent 8% (4 921 ha) of the natural territory and half of them are swamps (Beaulieu et al., 2014).
Figure 1: Location map of the 34 sampled swamps (dots) characterized by three
levels of urbanization (low, intermediate and high) in Quebec City, Quebec (Canada), 2016.
Site Selection and Sampling
The site selection process was based on an initial map where wetlands larger than 0.3 ha had been delineated and classified by photointerpretation into seven classes (bog, fen, forested peatland, marsh, swamp, wet meadow and shallow water). From these 791 mapped wetlands, we selected swamps according to the following criteria: 1) a distance of at least 300 meters between sampling sites, 2) a balance between riparian and isolated swamps (i.e. not directly connected to a permanent water course (Blanchette et al., 2019), and 3) sites ranging from 1 to 6 ha to avoid size effect bias. We also rejected all swamps surrounded by agricultural fields. The remaining 102 wetlands were field-validated to remove bush-dominated and destroyed swamps, as well as forested peatland and misinterpreted wetlands.
Finally, 34 wetlands met our criteria and their land use composition was characterized by a 100 m buffer zone with 9 categories (Appendix I) by photointerpretation of color orthophotos (Google, 2018) in QGIS 3.0.0 (QGIS Development Team, 2018). We then used a non-hierarchical k-means clustering (Legendre and Legendre, 2012) to define groups of landscape urbanization level surrounding swamps. A Principal Coordinate Analysis (PCoA) (Legendre et Legendre, 2012) based on land use composition in the immediate surroundings of each sampling site was performed in order to determine the land use gradients underlying this urbanization gradient (Appendix II).
A total of 92 plots were sampled across the 34 swamps, with 2 to 5 plots per swamp depending on its size. Within each swamp, plots of 400 m2 (20 × 20 m) were randomly positioned while respecting a 30-meter distance between each plot to avoid spatial autocorrelation and a 25-meter distance from the edge to avoid edge effect (Alignier et al., 2014). Vegetation sampling was carried out between the end of June (avoiding spring ephemeral) and beginning of September 2016 to avoid spring ephemeral. Species cover in each plot was assessed using seven classes: <1%, 1–5%, 6–10%, 11–25%, 26–50%, 51–75%, 76–100% and using nomenclature from VASCAN (Brouillet et al., 2019). We found no correlation between swamp size and species richness (r = 0.06, P = 0.738), indicating that the sampling method did not induce species-area bias.
Environmental variables and proxy were sampled or evaluated in each plot to further understand how plant communities respond to urbanization. Following Saucier (1994), we evaluated soil texture with a quantitative scale (0: sand to 12: clay) as well as the size, depth and abundance of soil mottles and humus thickness. Soil mottles and humus and peat thickness account respectively for water table depth and near-surface water saturation, as humus is degraded more rapidly in aerobic conditions (Zoltai and Vitt, 1995; Mitsch and Gosselink, 2015). Variation in microphotography was assessed with an index correlated with the amplitude between pits and mounds (0: flat, 1: <0.5 m, 2: 0.5-1 m, 3: more than 1 m of
amplitude). The cover of bryophytes (largely dominated by the genus Sphagnum), vernal pools and bare ground surfaces were estimated with the same classes as for vegetation. This variable accounts for water presence at ground level as above ground, as the most common bryophyte found in Canadian forest requires wet or humid condition (Goguen and Arp, 2017).
Data Analysis
Species classification
To better understand the impacts of urbanization levels on plant communities, we classified all species in three mutually exclusive groups (native wetland, native upland and exotic species). First, we discriminated species according to their origin (native or exotic). Then all native species were sorted according to their habitat preference (wetland or upland species; Appendix III). We used wetland indicator status (Reed, 1988) to sort native species into Wetland and Upland species. We considered wetland species those that are classified as “obligate” or “facultative wetland” while upland species included those classified as “facultative,” “facultative upland” and “upland” species. Wetland indicator status follows Bazoge et al. (2014) and the PLANT database (USDA, 2019). Only two species, Lythrum salicaria and
Lysimachia nummularia were at the same time exotics and wetland species; they
were neither frequent (15% and 9% of the sites respectively) nor abundant (<1% of cover in each site for both species) and were thus exclusively classified as exotic. Those three mutually exclusive groups (wetland, upland and exotic species) are hereafter referred to as species status.
Species Richness
We tested whether species number per site varied along the urbanization gradient (low, intermediate, and high) and according to species status (wetland, upland or exotic species), as well as the interaction between both factors, using linear mixed-effect models. Because a significant interaction was found between urbanization intensity and species status, we tested the individual effect of each factor with linear mixed-effect model for each level of the other factor and computed least square
Beta Diversity
Species beta diversity was compared between swamps of different urbanization levels with tests of homogeneity for multivariate dispersions (PERMDISP; Anderson et al., 2006). PERMDISP calculates the distance of each site to the centroid in ordination space (PCoA) and then tests whether these distances are different between groups (i.e., low, intermediate and high level of urbanization) through permutations. More precisely, a site-by-species matrix was first computed using importance values of species (IV: Barbour et al., 1997). The IVs were computed as the means of the species relative frequency (number of plots per swamp in which a species occurred relative to the total number of occurrences of all species) and relative dominance (cover of a species per swamp relative to the total cover of all species, using the median of the cover classes). We used IVs because our sampling design implied a different number of sampling plots per site. Then, a site-by-site distance matrix was computed using Hellinger distance (Legendre and Gallagher, 2001). This matrix was used to compute the centroid of each group. Then, the distance of each site to its associated group centroid was calculated, and the dispersion of these distances (within-group variance) was used as an estimate of beta diversity (the greater the within-group variance, the higher the beta diversity). Finally, the site-to-centroid distances were subjected to an ANOVA with 9,999 permutations to determine whether the dispersion (variance) differed between groups. A distinct analysis was performed for the entire flora (total pool of species) as well as for each species status (wetland, upland or exotic species).
To detect species shifts (turnover) among urbanization levels, we tested for location differences between centroids, using PERMANOVA with pseudo-F ratios (9,999 permutations; Anderson, 2001). Because this test is sensitive to differences in multivariate dispersion (Anderson and Walsh, 2013), data visualization was used to support interpretation of the statistical test. The differences in multivariate dispersion and centroid location among urbanization levels were illustrated in PCoA ordinations based on their respective distance matrix.
To better understand the mechanisms driving beta diversity, we further looked at the changes occurring inside communities: i.e., species turnover and richness difference. We partitioned beta diversity with the Sørensen dissimilarity following Legendre (2014) using presence-absence data, as the partitioning cannot be done on relative measures (Borcard et al., 2018). Within each level of urbanization, we conducted four partitionings, one of the total pool of species and one for each group of species status (wetland, upland and exotic species).
Environmental Factors
To assess how community composition is related to environmental factors, we performed a Redundancy Analysis (RDA) using Hellinger-transformed community data (Legendre and Gallagher, 2001). Explanatory variables were subjected to a stepwise selection to choose a parsimonious model.
All statistical analyses were performed using R (v. 3.5.1; R Foundation for Statistical Computing, Vienna, AT). Non-hierarchical clustering, multivariate dispersion and variable transformation were done with the vegan package (Oksanen et al., 2016), while beta diversity partitioning was computed with {adespatial} (Dray et al., 2017). Linear models (lme) and multiple comparison (pairs) were realized using respectively nlme (Pinheiro et al., 2017) and lsmeans packages (Russel, 2017) (Rcore Team, 2019).
Results
Urbanization and species richness
A total of 280 taxa (112 wetland, 119 upland and 49 exotic taxa) were identified in sampled swampsspecies status. The most frequent species (in> 90% of the swamps) were Acer rubrum and Dryopteris carthusiana, both native wetland species (see Appendix III for species nomenclature). The most frequent upland species were
Athyrium filix-femina (89% of sites), Abies balsamea (83%) and Amelanchier arborea
(83%) while Epipactis helleborine (40%) and Ranunculus repens (29%) were the most frequent exotic species.
The impact of urbanization on species richness differed according to the species status (Table 1). Only exotics responded to urbanization gradient, with about 2 and 4 times more species at intermediate and high urbanization levels than at low urbanization level, respectively (Figure 2). The richness of wetland and upland species did not vary along the urbanization gradient and were similar within each urbanization level (Figure 2). However, exotic species were 27, 11 and 6 times less diversified than native species (both wetland and upland species summed together) at low, intermediate and high levels of urbanization, respectively (Figure 2).
Table 1: Effect of urbanization level (low, intermediate, high) and species status
(wetland, upland, exotic) as well as their interactions on mean species richness tested with a linear mixed-effect model and contrasts. Bold indicates significant differences. Species status ~ Urbanization level DF F p Urbanization 31 0.32 0.7285 Status 62 216,158 <.0001 Urbanization: Status 62 5.512 0.0007
Urbanization | Status Estimate SE t-ratio p
Wetland species Low — Int. 0.074 0.381 0.193 0.980 Low—High 0.298 0.387 0.768 0.725 Int. —High 0.224 0.352 0.637 0.801 Upland species Low — Int. 0.281 0.381 0.739 0.743 Low—High 0.619 0.387 1.599 0.261 Int. —High 0.338 0.352 0.961 0.607 Exotic species Low — Int. -0.844 0.381 -2.215 0.084 Low—High -1.522 0.387 -3.928 0.001 Int. —High -0.678 0.352 -1.928 0.148
Status | Urbanization Estimate SE t-ratio p
Low Exo—Wet. -4.218 0.373 -11.32 <.0001 Exo. —Up. -4.827 0.373 -12.954 <.0001 Wet. —Upl. -0.609 0.373 -1.635 0.239 Intermediate Exo – Wet. -3.301 0.310 -10.646 <.0001 Exo. —Upl. -3.702 0.310 -11.941 <.0001 Wet. —Upl. -0.401 0.310 -1.294 0.404 High Exo—Wet. -2.399 0.323 -7.433 <.0001 Exo. —Upl. -2.686 0.323 -8.324 <.0001 Wet. —Upl. -0.287 0.323 -0.891 0.648
Figure 2: Differences in mean species richness (%) according to urbanization
levels and species status, bars provide standard deviation. Capital letters indicate differences between levels of urbanization within a single species status and lower case letters indicate differences between species status within a single level of urbanization. Contrasts were used to assess significance (p <0.05).
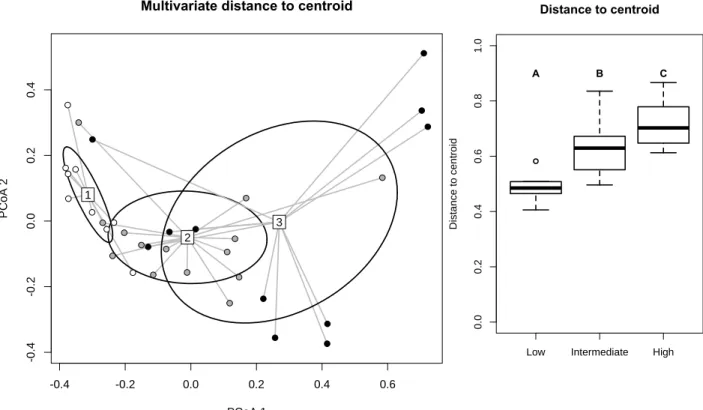
Beta Diversity
When computed on the total pool of species, beta diversity in swamps differed between urbanization levels (F = 20.7; P = 0.0001). More precisely, there was a significant increase in site dispersion around centroids, from low to high urbanization levels (Figure 3; as indicated by ellipse size). Differences in beta diversity were associated with changes in species composition among urbanization levels as the test for centroid location was significant (F = 4.3; P = 0.00006). The even spacing between pairs of centroids further indicates a constant change of species composition among all urbanization levels (Figure 3).
Low Intermediate High
Mean Species Richness
S p e c ie s r ic h n e s s 0 1 0 2 0 3 0 4 0 5 0 a a b a a b a a b A A A A A A A AB B Species Status Wetland Upland Exotic
Figure 3: Influence of urbanization levels on the multivariate dispersion of species
composition in swamps. Urbanization levels were determined in immediate surroundings of swamps in a 100 m radius buffer. Taxonomic beta diversity is measured as the distance of each site to their group centroid using Hellinger distance and represented here on the first two axes of PCoA, circles corresponding to standard deviation. Boxplots show swamp-to-centroid distance (median and quartiles) for each urbanization level. On the PCoA, changes in dispersion around centroids reflect variation in beta diversity; a change of centroid position indicates species turnover. Circles are ellipses of standard deviation Dots colours represent levels of urbanization (white: low, gray: intermediate and black: high).
This pattern of swamp differentiation also appeared when analyzing wetland and upland species distinctively; beta diversity increased from low to high level of urbanization (Table 2bc). Yet, when considering exotic species only, beta diversity remained constant along the urbanization gradient (Table 2d). The processes involved in beta diversity patterns also differed between groups of species status.
For all species, and especially wetland or upland species, species replacement was the main process explaining beta diversity patterns (Table 2 abc). When considering
1
2
3
Multivariate distance to centroid
method = "euclidean" PCoA 1 P C o A 2 -0.4 -0.2 0.0 0.2 0.4 0.6 -0 .4 -0 .2 0 .0 0 .2 0 .4
Low Intermediate High
0 .0 0 .2 0 .4 0 .6 0 .8 1 .0 Distance to centroid D is ta n c e t o c e n tr o id A B C
all species or wetland species only, replacement was more influential than richness differences at all urbanization levels but especially at high level. For upland species, the respective contribution of both processes was constant among urbanization levels, with a peak at intermediate level (Table 2abc). For exotic species, beta diversity of swamps was driven by richness differences, especially at low urbanization where it was four times more influential than species replacement (Table 2d).
Table 2: Partition of beta diversity (BD) into replacement (turnover between species)
and richness difference (changes in the number of species) at different levels of urbanization (low, intermediate, high) on a) all, b) wetland, c) upland and d) exotic species. Replacement and richness difference are reported in percentages of the total beta diversity value.
Urbanization level BD total Replacement (%) Richness Difference (%) a) All species Low 0.16 60.7 39.3 Intermediate 0.24 70.4 29.6 High 0.28 79.7 20.3 b) Wetland Low 0.18 55.5 44.5 Intermediate 0.25 61.4 38.6 High 0.27 72.3 27.7 c) Upland Low 0.15 63.3 36.7 Intermediate 0.24 70.6 29.4 High 0.30 63.7 36.3 d) Exotic Low 0.30 17.9 82.1 Intermediate 0.38 43.9 56.1 High 0.35 47.2 52.8
Environmental Changes and Species Composition
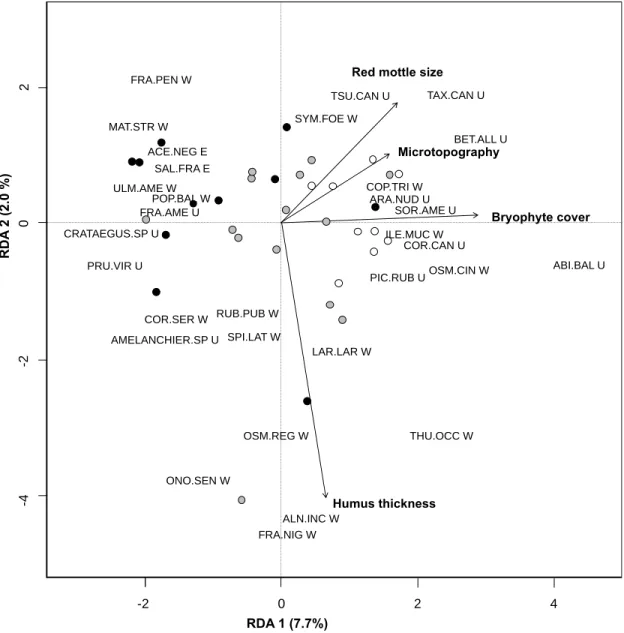
The urbanization gradient was mainly represented on the first axis of the RDA which explained 7.7% of total variation (58.4% of explained variation) in species composition (Figure 4). Each different level of urbanization was characterized by co-occurring groups of species. Less urbanized sites were associated with abundant bryophyte cover, hydromorphic soil feature (red mottle) as well as a well-developed microtopography (Figure 4). Those sites were characterized by the presence of wetland (Coptis trifolia, Ilex mucronata, Larix laricina, Osmundastrum
cinnamomeum, and Thuya occidentalis) and upland (Abies balsamea, Aralia nudicaulis, Betula alleghaniensis, Picea rubens, Taxus canadensis and Tsuga canadensis) species. Moderately urbanized sites were associated the presence of
both wetland (Alnus incana subsp. rugosa, Onoclea sensibilis and Spirea alba subsp. latifolia) and upland species (Athyrium filix-femina and Betula paperyfera). Finally, highly urbanized sites were associated with species of different habitat preference, with exotic species almost restricted to this end of the urbanization gradient (Acer negundo, Crataegus spp., Fraxinus spp., Populus balsamifera,
Figure 4: Correlation triplot relating five selected environmental variables (arrows),
34 swamps (dots) and 29 species (see code for species in appendix III). Last letter of species name’s indicate their status (W: wetland, U: upland, E: exotic) and only species most fitted to the model are shown. Angle between sites, environmental variables and themselves reflects their correlation. Orthogonal projection of site on an environmental variable approximates its value. Dot colours represent levels of urbanization (white: low, gray: intermediate and black: high).
FRA.PEN W MAT.STR W ACE.NEG E SAL.FRA E ULM.AME W POP.BAL W FRA.AME U SYM.FOE W TSU.CAN U TAX.CAN U BET.ALL U ARA.NUD U ABI.BAL U PIC.RUB UOSM.CIN W ILE.MUC W SOR.AME U COP.TRI W LAR.LAR W THU.OCC W ALN.INC W FRA.NIG W ONO.SEN W COR.CAN U PRU.VIR U CRATAEGUS.SP U COR.SER W AMELANCHIER.SP U SPI.LAT W OSM.REG W RUB.PUB W
Red mottle size
Microtopography Humus thickness Bryophyte cover 2 0 -2 -4 -2 0 2 4 RDA 1 (7.7%) R D A 2 ( 2. 0 % )