Microbial Protease or Yeast Extract—Alternative Additions for Improvement of Fermentation Performance and Quality of Beer Brewed with a High Rice Content
9
0
0
Texte intégral
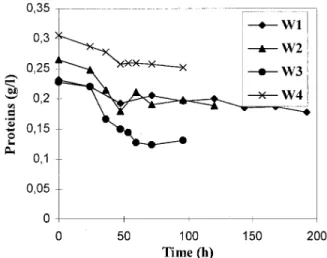
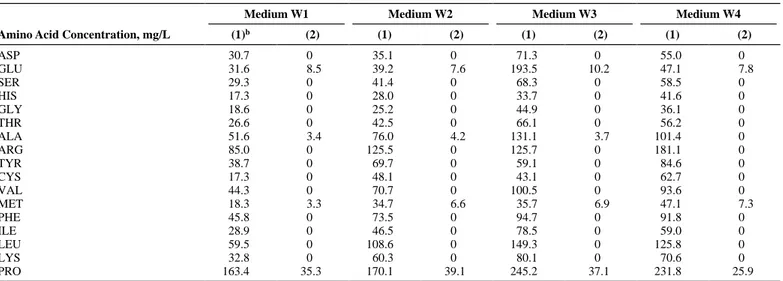
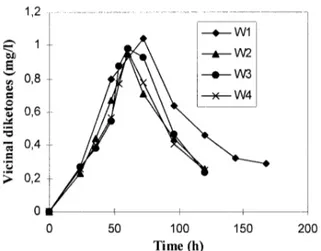
Figure
Documents relatifs