EFFETS PHYSIOLOGIQUES DE L'ÉLAGAGE CHEZ LES PEUPLIERS HYBRIDES
MÉMOIRE
PRÉSENTÉ
COMME EXIGENCE PARTIELLE
DE LA MAÎTRISE EN BIOLOGIE
PAR
VINCENT MAURIN
Mise en garde
La bibliothèque du Cégep de l’Témiscamingue et de l’Université du Québec en Abitibi-Témiscamingue a obtenu l’autorisation de l’auteur de ce document afin de diffuser, dans un but non lucratif, une copie de son œuvre dans Depositum, site d’archives numériques, gratuit et accessible à tous.
L’auteur conserve néanmoins ses droits de propriété intellectuelle, dont son droit d’auteur, sur cette œuvre. Il est donc interdit de reproduire ou de publier en totalité ou en partie ce document sans l’autorisation de l’auteur.
Warning
The library of the Cégep de l’Témiscamingue and the Université du Québec en Abitibi-Témiscamingue obtained the permission of the author to use a copy of this document for non-profit purposes in order to put it in the open archives Depositum, which is free and accessible to all.
The author retains ownership of the copyright on this document. Neither the whole document, nor substantial extracts from it, may be printed or otherwise reproduced without the author's permission.
Je tiens à remercier ma directrice de recherche Annie DesRochers de m'avoir donné l'opportunité de travailler sur ce projet. Ses conseils, son accessibilité ainsi que les nombreuses relectures m' ont permis d ' être bien encadré tout au long de ces trois années. Je tiens aussi à remercier Line Blackburn, pour ses compétences et son aide sur le terrain. Merci aussi à mes collègues du laboratoire Mathilde Chomel, Amélie Trottier-Picard, Émilie Tarroux, Clémentine Pernot, et Emmanuel Adonsou pour leurs conseils, les services rendus et leur amitié.
Je tiens à remercier Marc Mazerolle de l'UQAT et Jean-Pierre Durbec pour l' aide en statistiques. Merci aussi à tout le campus de l'Université du Québec en Abitibi-Témiscamingue d ' Amos pour son soutien.
Merci à tous les amis qui m' ont aidé de près ou de loin pour avancer dans ce proj et.
Enfin, un merci particulier à ma famille; ma mère Marie-Christine, mes sœurs Mathilde, Anne et Hélène, qui m' ont appuyé et suiv i tout au long de mon cursu s et à mon père Jean-François qu i malheureusement ne sera pas là p our lire les conclusions de mes travaux.
LISTE DES FIGURES ... vi
LISTE DES TABLE AUX ... vii
LISTE DES ABRÉVIATIONS, SIGLES ET ACRONYMES ... ... ... viii
RÉSUMÉ ... ... ... ... ... ... ix
CHAPITRE! INTRODUCTION GÉNÉRALE ... ... ... ... 1
1.1 L ' élagage: définition et principe d' application ... ... ... 2
1.2 Effets physiologiques de l'élagage ... 5
1.2.1 Photosynthèse ... ... ... ... ... 5
1.2.2 Dynamique des glucides ... 6
1.2.3 Concentration foliaire en azote ... 7
1.2.4 Stress hydrique et isotop es du carbone ... ... ... ... 8
1.3 Effets de l 'élagage sur la croissance ... 9
1.3.1 Accroissement en diamètre et défilement.. ... ... ... ... 9
1.3.2 Accroissement en hauteur ... 9
1.4 Branches épicormiques ... ... ... ... ... 10
1. 5 Expérimentation ... 11
1.6 Bibliographie ... ... ... ... ... ... 12
CHAPITRE II HYBRID POPLAR PHYSIOLOGICAL AND GROWTH RESPONSES TO PRUNING SEASON AND INTENSITY ... 17
2.1 Abstract ... ... ... ... ... ... 17
2.2 Introduction ... ... ... ... ... 18
2.3 Materials and methods ... ... ... ... ... 20
2 .3.1 Study sites ... ... ... ... ... 20
2 .3.2 Field sampling ... 21
2 .3.3 Laboratory work ... ... ... ... ... 22
2.4 Statistical analyses ... 23
2.5.1 Growth responses ... 26
2.5.2 Net photosynthesis ... ... ... ... ... 28
2.5.3 Stomatal conductance ... 30
2.5.4 Total non-structural carbohydrates (TNC) ... ... ... ... 30
2.5.5 Leaf nitrogen concentrations ... ... ... ... 31
2.5.6 Carbon isotope discrimination ... 31
2.6 Discussion ... ... ... ... ... 32
2.7 I'vianagement implications ... 36
2.8 References ... 36
CHAPITRE III PRODUCTION AND ROLE OF EPICORMIC SHOOT FORMATION IN HYBRID PO PL ARS: EFFECTS OF PRUNING SEASON AND INTENSITY ... .43
3.1 Abstract ... ... ... ... ... ... 43
3.2 Introduction ... 44
3.3 I'viaterials and methods ... ... ... ... ... 45
3 .3.1 Study site ... ... ... ... ... 45
3 .3.2 Field sampling ... 46
3 .3.3 Laboratory work ... ... ... ... ... .47
3 .3.4 Statistical analyses ... 48
3.4 Results ... ... ... ... 50
3 .4.1 Number of epicormic shoots ... ... ... ... 50
3 .4.2 Growth responses ... 52
3 .4.3 Net photosynthesis ... ... ... ... 53
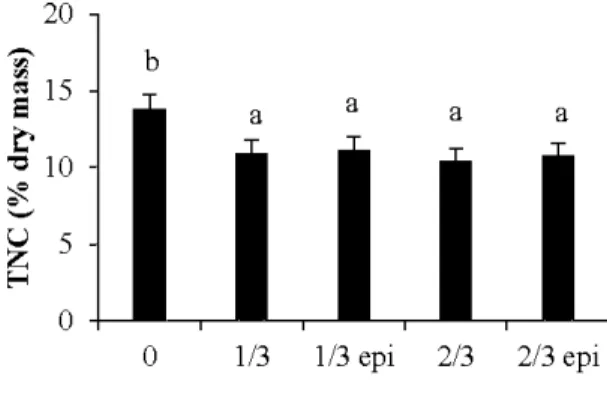
3 .4.4 Total n on-structural carbohydrates (TNC) ... ... 55
3 .4.5 Epicormic shoots morphological responses ... ... ... 55
3.5 Discussion ... ... ... ... ... 58 3.6 I'vianagement implications ... ... ... ... ... 6 1 3.7 Acknowledgements ... ... ... ... ... 6 1 3. 8 References ... 6 1 CH APITRE IV CONCLUSION GENERALE ... 67 4.1 Bibliographie ... ... ... ... ... ... 72
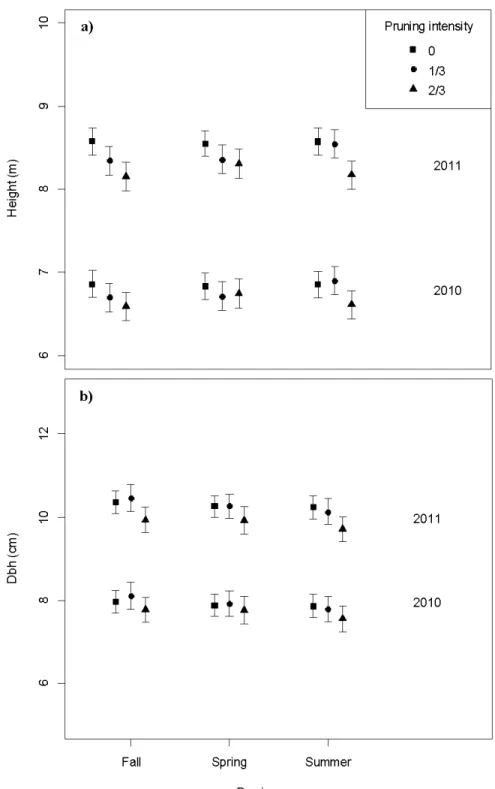
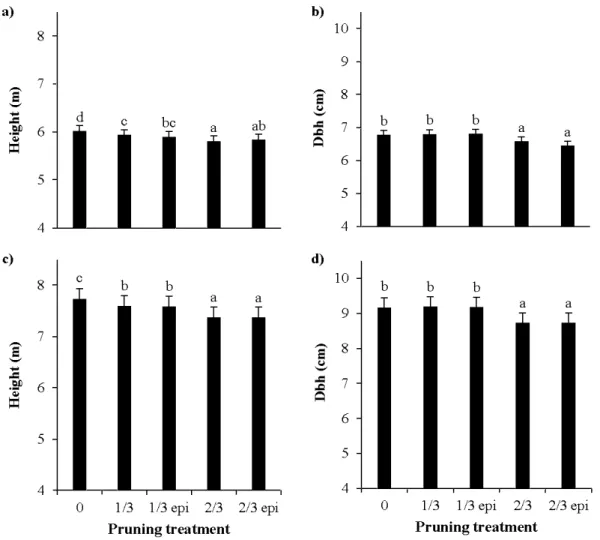
Figure Page 2.1 Model-averaged predictions for height (a) and dbh (b) in 2010 and 2011, for all
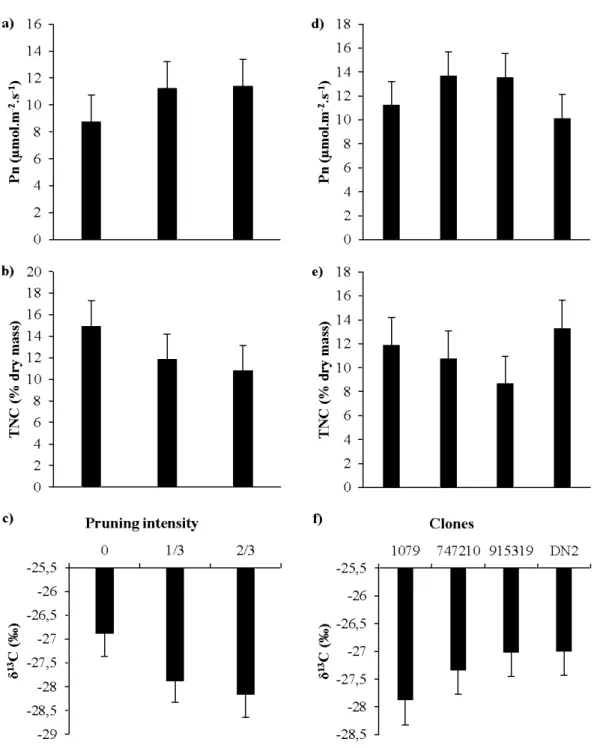
pruning seasons and intensities . . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . . 27 2.2 Model-averaged predictions for net photosynthesis (Pn; a,d), Total
non-structural carbohydrates (TNC; b,e) and leaf carbon isotope ratios (813C; c,d)
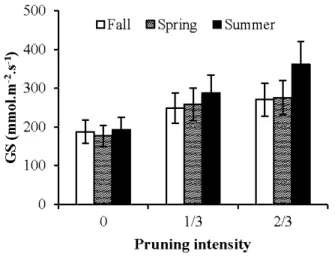
for all pruning intensities and clones... 29 2.3 Model-averaged predictions for stomatal conductance across pruning intensities
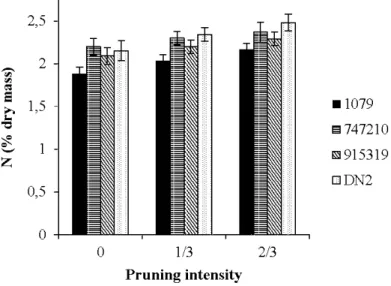
and seasons .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . . 30 2.4 Model-averaged predictions for foliar nitrogen concentration across pruning
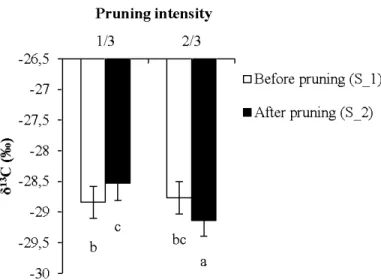
intensities for each clone... 31 2.5 Model-averaged predictions for 813C values across pruning intensities for
leaves of summer-pruned trees .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . .. . . 32 3.1 Predictions for the number of epicormic shoots produced (a) after one growing
season (epi2010), (b) during the second growing season (epi2011 new) and (c)
after two growing seasons ( epi20 11_ old) for all pruning intensities and seasons 51 3.2 Predictions for h eight and dbh, one (a; b) and two (c; d) years after pruning on
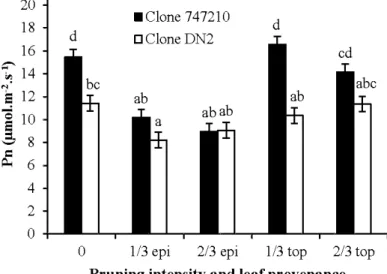
trees with or without epicormic shoots... 52 3.3 Predictions for mean net photosynthesis (Pn) of leaves from (0) unpruned trees,
( 113 epi) epicormic shoots of 113 pruned trees, (2J3 epi) epicormic shoots of 2/3 pruned trees, ( 113 top) canopy of 113 pruned trees and (2J3 top) canopy of
2/3 pruned trees, for hybrid poplar clones 747210 and DN2 ... 53 3.4 Predictions for mean total non-structural root carbohydrates for unpruned trees
(0), 113 pruned trees without epicormic shoots (113), 113 pruned trees with epicormic shoots (113 epi), 2J3 pruned trees without epicormic shoots (2J3) and
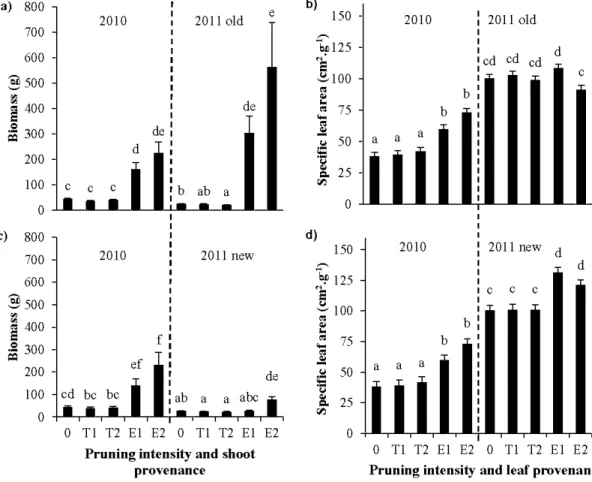
2/3 pruned trees with epicormic shoots (2J3 epi) ... 55 3.5 Predictions for mean shoot biomass (a, c) and specifie leaf area (SLA; b, d) of
canopy shoots from unpruned trees (0), 113 pruned trees (Tl), 2J3 pruned trees (TI) and of epicormic shoots from 113 pruned trees (El), 2J3 pruned trees, after one growing season (E2) after one growing season (20 1 0), during the second
T~km ~~ 2.1 Selected models according to results of Akaike information criterion adjusted
for small samplesizes . . . 25 2.2 Mean tree height, diameter at breast height and estimated volume per hectare
for each clone . . .. .. . . .. .. . . .. .. . . .. .. . . .. .. . . .. .. . . .. .. . . .. .. . . .. .. . . .. .. . . .. .. .. . .. .. . . .. .. . . .. .. . . .. .. . . .. .. . . .. .. 26 3.1 Comparison of the candidate models considered to explain the number of
epicormic shoots produced after one and two growing seasons... 49 3.2 Best fit models chosen with stepwise selection testing the effect of pruning
intensity and epicormic branching on net photosynthesis and root total
non-structural carbohydrate concentrations... 54 3.3 Best fitted models chosen with stepwise selection testing the effect of pruning
AICc Akaike Information Criterion corrected
Dbh Diameter at breast height
N Nitrogen
TNC Total non-structural carbohydrates
L'élagage est une opération sylvicole qui consiste à enlever les branches situées sur le tronc jusqu'à une certaine hauteur de la couronne verte. Le but est de produire des billes de bois sans nœuds. C'est une pratique ancienne qui a fait l'objet de nombreuses études, mais rares sont celles qui se sont intéressées au choix de la saison d'élagage et son incidence sur la physiologie et la croissance du peuplier. Comprendre ces effets permettrait de développer des scénarios sylvicoles plus adaptés selon les conditions actuelles ou futures. L'objectif de cette étude était de déterminer les effets de la saison et de l'intensité d'élagage sur l'activité physiologique de 4 clones de peuplier hybride. L'apparition de branches épicormiques ou gourmands (rameaux qui poussent spontanément sur une partie du tronc exposé à la lumière) étant une réponse courante des peupliers à l'élagage, l'étude était dotée d'un second objectif: déterminer si ces branches constituent un puits ou une source d'énergie pour l'arbre. L'étude s'est déroulée dans une peupleraie localisée à New Liskeard, ON, dont les arbres ont été plantés au printemps 2007 à un espacement de 3,5 rn (816 tiges.ha.1). Les élagages ont été
effectués à l'automne 2009, au printemps 2010 et, à l'été 2010, à deux intensités différentes; 113 et 2J3 de la cime vivante, et un témoin non élagué. Les arbres élagués avaient une photosynthèse nette plus élevée que les témoins, mais l'effet était similaire pour les arbres élagués à 113 (+21.7%) et à 2J3 (+23.6%). L'élagage entrainait aussi une diminution des réserves de glucides dans les racines, qui était plus importante pour les arbres élagués à 2/3 (-29%) qu'à 1/3 (-22%). Ce résultat suggère que la réponse photosynthétique mise en place suite à l'élagage était insuffisante pour reconstituer l'ensemble des réserves. L'élagage a augmenté la concentration en azote foliaire et la discrimination du 13Carbone (813C), ce qui suggérait une meilleure résistance au stress hydrique. Deux ans après les traitements, élaguer 2J3 de la cime entrainait un ralentissement de la croissance peu importe la saison d'élagage, alors qu'élaguer à 113 en été n'affectait pas la croissance des arbres. De plus, élaguer en été entrainait une augmentation de la concentration d 'azote dans les feuilles, de la conductance stomatique et de la discrimination du 13Carbone, ce qui indiquait de meilleures dispositions pour récupérer comparativement aux arbres taillés en automne et au printemps. La saison d'élagage jouait auss i un rôle important dans la production de branches épicormiques, puisque les arbres élagués en été ont produit significativement moins de gourmands que les arbres élagués en automne et au printemps, et ce indépendamment de l'intensité d'élagage. Les feuilles des branches épicormiques contribuaient à la photosynthèse globale de l'arbre, sans pour autant restaurer les réserves de glucides. La présence de ces branches n ' a pas influencé la croissance en diamètre. Ces branches semblaient donc être auto-suffisantes en énergie. Nous avons conclu qu'élaguer à 113 de la cime en été était la meilleure option pour produire du bois de qualité sans affecter la croissance et la physiologie des peupliers. De même, si des branches épicormiques apparaissaient elles pouvaient être émondées sans perturber l'activité physiologique et la croissance subséquente des peupliers hybrides.
INTRODUCTION GÉNÉRALE
Au Canada l'aménagement forestier extensif est dominant, et les taux de croissance des forêts primaires sont faibles. Les longues révolutions qui en découlent sont un frein à la demande future de bois et de matière ligneuse à l'échelle mondiale. Les plantations d'essences à croissance rapide sont considérées comme la principale alternative pour produire plus de bois sur des surfaces plus petites tout en préservant les forêts primaires (Anderson et Luckert, 2007 ; Heilman, 1999). Au Canada, bien que la ligniculture soit récente, les plantations de peuplier hybride (Populus spp.) peuvent atteindre des accroissements de 6 à 29 m3/ha/an (Park et Wilson, 2007). Contrairement à il y a quelques décennies, le peuplier est maintenant une essence prisée par l'industrie. Son bois est utilisé dans la fabrication des panneaux de particules, des poutres lamellées, des contre-plaqués, de sciages destinés à des utilisations multiples et pour la production de pâte et de papier. De plus, la pratique de l'élagage permet d'augmenter la proportion de bois avec une plus forte valeur ajoutée et de valoriser des débouchés tels que le contre-plaqué et le déroulage.
La production de bois sans défauts est en effet accélérée dans les peuplements en plantation par des élagages. Bien que les réponses en termes de croissance à différentes intensités d'élagage aient été étudiées, les effets physiologiques de l'élagage sont encore largement inconnus. La connaissance de ces effets nous permettrait de développer divers scénarios sylvicoles pouvant s'adapter à des conditions de croissance variées, plutôt que d'y aller à tâtons, par essais et erreurs. Par exemple, l'enlèvement d'une partie du feuillage causé par l'élagage agit sûrement sur le régime hydrique des arbres, qui pourrait alors être utilisé en conditions de sécheresses temporaires. Les changements physiologiques qui s'opèrent chez les arbres élagués pourraient également varier en fonction de la saison d 'élagage, de par les
différents stades physiologiques causés par les changements de sa1sons. De la sorte, le traitement pourrait être modifié en fonction des conditions climatiques. Enfin, une réponse fréquente des peupliers à l'élagage est la production de branches épicormiques ou gourmands. Bien que l'on sache que leur production est reliée à la mise en lumière soudaine du tronc, on ne sait pas si la production de ces gourmands constitue un coût énergétique pour
1 'arbre (au détriment de la croissance en diamètre), ou alors si cela constitue un bénéfice, par la production importante et rapide de feuillage qui contribuera à la photosynthèse de l'arbre.
1.1 L'élagage: définition et principe d'application
D'après les différents guides sur la populiculture disponibles au Canada (Boysen et Strobl, 1991 ; Portier et al., 2011 ; van Oosten, 2006), l'élagage est un terme générique qui englobe 3 opérations :
- la taille de formation; elle vise à donner rectitude et solidité à un arbre en éliminant les têtes doubles ou multiples ainsi que les branches qui par leur fort développement déséquilibrent le port de l'arbre .
-L' élagage; il a pour but de produire du bois sans nœuds en coupant des branches mortes ou vivantes de la partie inférieure du tronc.
-L'émondage; il complète l'élagage en supprimant les gourmands qui se développent sur le tronc à partir de bourgeons épicormiques.
Ces diverses opérations sylvicoles permettent d'améliorer la forme et la qualité des bois en contenant les défauts engendrés par la coupe des branches à un noyau central, augmentant ainsi la proportion de bois sans défaut (Kerr et Morgan, 2006 ; Montagu et al., 2003 ; Polge, 1969; Tak:iya et al., 2009). En effet, chaque branche produit un nœud sur le tronc et abaisse ainsi la valeur commerciale du bois (Neilsen et Pinkard, 2003). La résistance mécanique est altérée par la présence de nœuds, et le fil du bois est dévié au voisinage des nœuds ce qui peut occasionner des défauts dans les produits finis. De la sorte, en Europe le prix des feuillus précieux peut varier de 200 à 1000 € par m3 selon la qualité (Oosterbaan et al., 2009). Dans les premières années de la plantation on privilégie la taille de formation car la rectitude de la bille de pied est primordiale pour un bois de qualité (Hubert et Courraud, 1994) . Pour l'élagage il est préférable d'intervenir quand les arbres sont âgés entre 3 et 5 ans, ou quand le diamètre à 1,30 rn de l' arbre se situe entre 8 et 10 cm (Gagnon, 2006). Néanmoins, ces
chiffres peuvent varier en fonction de la vitesse de croissance des arbres. Si la plantation est vigoureuse il se peut qu'il faille intervenir avant. Généralement l'élagage doit être précoce et progressif (Hubert et Courraud, 1994). Précoce, il coûte moins cher et il est plus efficace car les branches sont de plus faible diamètre. De plus les défauts sont contenus dans un noyau de plus faible diamètre. Progressif, il traumatise moins les arbres. En effet l'élagage représente une forme d'agression pour l'arbre. De ce fait de nombreuses recommandations sont basées sur le diamètre des branches. Notamment, il est préférable d'élaguer dans le jeune âge des branches de faible diamètre pour diminuer la section où les pathogènes peuvent entrer et accélérer la cicatrisation (Montagu et al., 2003 ; Polge, 1969). Pour faire face aux blessures et agressions l'arbre réagit de deux manières (Shigo, 1984) :
- en isolant la zone infestée, pour ce faire il oppose à la progression des agents pathogènes des barrières chimiques dans lesquelles on trouve des substances antifongiques et antibiotiques.
- en développant sur le pourtour de la blessure un bourrelet cicatriciel et des barrières internes qui à terme doivent refermer les plaies.
Dans les deux cas, les tissus infectés ou blessés ne se régénèrent pas, ils sont enkystés dans les tissus sains. Pour le clone P. delta ides x (P. laurifolia x P. nigra) var. Walker, Block et al.
(2009) mentionnent que l' on peut intervenir une à deux années après l'établissement de la plantation. Rytter et Jansson (2009), recommandent d 'élaguer quand la tige atteint un diamètre (dhp) de 15 cm, et de couper les branches dont le diamètre est compris entre 3 et 5 cm. L' important serait l'âge des tissus blessés par la coupe. En effet, les petites blessures n'affectent que les jeunes tissus, ceux contenant des cellules de parenchyme actives, donc le plus fort taux de réserves en énergie. Plus la plaie est petite, plus la cicatrisation est rapide. La coupe de grosses branches entraîne quant à elle une cicatrisation plus longue et augmente le risque d'une coloration du bois plus étendue (Soutrenon, 1995). Enfin, d 'un point de vue économique, une branche de petit diamètre âgée de 1 ou 2 ans se coupe sans difficulté alors qu'après 3 ou 4 saisons de végétation, couper la même branche demande 3 ou 4 fois plus de temps et rend l'élagage plus couteux (Hubert et Courraud, 1994).
L ' intensité d 'élagage correspond à la hauteur jusqu 'à laquelle on enlève des branches. La hauteur de bois sans nœuds correspond généralement à la longueur utilisée par l'industrie
soit 6 à 8m. Bien entendu l'opération ne se fait pas d'un seul coup. On conseille d'atteindre cette hauteur à l'aide de 4 à 5 élagages répartis sur les 10 premières années de la plantation (Boysen et Strobl, 1991 ; Portier et al., 2011 ; van Oosten, 2006) soit en 2 ou 3 élagages
(Hubert et Courraud, 1994).
Les plaies d'élagage représentent une porte d'entrée pour les agents pathogènes, et plus une blessure reste ouverte longtemps, plus elle a de chance de s'infecter. Polge ( 1969), considère que la période la plus propice pour l'élagage semble être celle qui précède le départ de la végétation i.e., au printemps, car l'entrée en action de l'assise cambiale entraîne une
cicatrisation plus rapide des plaies, qui sont à ce moment-là chez les conifères recouvertes d'une pellicule de résine. Cependant à la même période, les écoulements par les plaies de taille attirent les insectes et favorisent les pourritures (Hubert et Courraud, 1994). Chez Pinus sylvestris, élaguer à l'automne augmentait le risque d'infection des plaies par Phacidium coniferarum (Uotila et Mustonen, 1994). Montagu et al. (2003) précisent que même si en
Tasmanie Eucalyptus nittens peut être élagué à n ' importe qu'elle saison, la cicatrisation est
bien plus rapide quand l'arbre est en pleine croissance et que le diamètre des branches est plus faible. En effet, les mécanismes de compartimentation dépendent de 1' activité physiologique des cellules des parenchymes ainsi que de la disponibilité des réserves emmagasinées. Pour cette raison, la saison a une influence sur la réaction des arbres suite à des blessures (Dujesiefken et Stobbe, 2002). Il semble donc que la taille des branches ne devrait pas être faite durant la période hivernale et que les blessures faites pendant la période de végétation sont plus efficacement compartimentées (Dujesiefken et al., 2005).
Lonsdale (1993), dans son étude sur la saison d'élagage a établi un calendrier des "bonnes" et des "mauvaises" saisons de l'élagage pour 9 espèces représentatives de la Grande Bretagne. Il préconise un élagage au milieu de 1 'été durant la saison de croissance car 1' arbre est actif et ses réserves ont été restaurées par la photosynthèse. Les composants de la défense des arbres dépendent de la croissance des cellules, qui sont plus actives qu'en période de dormance. En été, la disponibilité en sucres pour la conversion en défenses chimiques, comme la production de phénols, n'est pas réduite par le débourrement ou la floraison (Lons dale, 1993). De plus les blessures seraient moins susceptibles de s'infecter. En zone boréale au Canada, où la saison de végétation est plus courte qu'en Europe, les guides de
populiculture préconisent d'élaguer le peuplier au début de l'été (Boysen et Strobl, 1991 ; Portier et al., 2011 ; van Oosten, 2006). Cependant ces recommandations découlent
d'avantage des pratiques courantes et ne sont pas basées sur des études scientifiques qm auraient testé l'effet de la saison d'élagage sur la croissance ou la vigueur des arbres.
1.2 Effets physiologiques de l'élagage
1.2.1 Photosynthèse
Le taux de photosynthèse de la feuille varie considérablement pendant la journée. Il est communément bas tôt le matin, et au fur et à mesure que 1 'intensité lumineuse augmente et que l'air se réchauffe, les stomates s'ouvrent et la photosynthèse nette commence à augmenter rapidement pour atteindre un maximum vers midi. Aux alentours de midi il y a une baisse du taux de photosynthèse qui serait due à la fermeture des stomates causée par une réponse à la tension importante subie par la colonne d'eau à cette heure de la journée. À
nouveau, le maximum est atteint dans l' après-midi. Ensuite, une baisse de la photosynthèse est observée en début de soirée quand l'intensité lumineuse et la température diminuent (Kozlowski et Pallardy, 1997). Les réponses physiologiques à l'élagage s'opèrent sur deux plans; le premier, lié directement au processus photosynthétique, se traduit par une augmentation de la photosynthèse de la feuille. Le second se traduit par une accélération du développement de la feuille et une augmentation de la longévité des feuilles résiduelles (Beadle et al., 2004). Hart et al. (2000) ont démontré qu'après une forte défoliation chez le
peuplier faux-tremble (Populus tremuloides Michx.), on observait une augmentation de la
conductance stomatique et une photosynthèse compensatoire des feuilles résiduelles au bout de huit jours. La photosynthèse compensatoire correspond donc à l'augmentation du taux de photosynthèse d'un arbre partiellement défolié, comparé à un arbre non défolié sur un feuillage du même âge (Nowak et Caldwell, 1984). Cependant, une défoliation plus sévère (98%) n'augmentait plus la photosynthèse compensatoire ce qui suggère qu'il y ait une limite à cette augmentation de la photosynthèse qui pourrait être due à une contrainte physique : à 50% de défoliation, les stomates étaient peut-être déjà ouverts au maximum. La plasticité des feuilles à augmenter leur photosynthèse serait donc limitée Hart et al. (2000).
(élagage) chez des Eucalyptus (E. globulus) âgés de 1 an variait dans le temps :une semaine après la défoliation les valeurs du Amax étaient respectivement 1,5 et 1,7 fois plus élevées pour des taux d'élagage de 50 et 70%, pour retrouver une valeur proche des arbres non élagués au bout de cinq semaines (Tumbull et al., 2007). Bassman et Dickmann (1982), ont observé que le taux de photosynthèse des feuilles résiduelles de peupliers partiellement défoliés était plus élevé que dans des feuilles similaires de peupliers intacts. Cette différence était mesurable pendant cinq semaines. Cependant, défoliation et élagage sont deux stress différents, bien que ces deux stress entrainent un débalancement entre la surface foliaire et la masse racinaire (ratio tige/racines). Les observations faites chez des peupliers défoliés pourraient donc différer chez des peupliers élagués.
1.2.2 Dynamique des glucides
La photosynthèse est un processus essentiel à la fabrication de l'énergie chez les végétaux, notamment les hydrates de carbone non structuraux totaux (TNC) ou glucides. Les TNC se retrouvent chez l'arbre sous différentes formes telles les sucres solubles (sucrose, fructose, glucose) et 1 'amidon. Les glucides s'accumulent dans les racines et les tissus v ivants des tiges sous forme de réserves, qui fourniront 1 'énergie nécessaire à la croissance de la plante et au maintien des processus cellulaires. La concentration en glucides peut varier en fonction de l'espèce, du site, et de l'âge des peuplements (Kozlowski et Pallardy, 1997). Cette concentration connait également des variations naturelles tout au long de la saison de croissance, régulée par les changements de saison et la respiration racinaire (DesRochers et al., 2002). En forêt boréale chez le peuplier faux-tremble (Populus tremuloides), les concentrations racinaires en glucides à l ' automne sont faibles. Elles augmentent au cours du printemps et de l' été jusqu 'à l'arrêt de l'élongation de la partie aérienne et des feuilles (DesRochers et al., 2002). La concentration en TNC diminue ensuite à cause de la croissance des racines en automne. L'amidon est alors converti en sucre, pour soutenir la croissance racinaire (Landhausser et Lieffers, 2003). Chez le peuplier faux-tremble, la variation de la concentration en glucides selon les périodes de la saison de végétation diffère du schéma classique des espèces à feuilles caduques (DesRochers et al., 2002), chez lesquelles la concentration est maximale en automne et minimale après l'éclosion des bourgeons (Larcher, 2003). Enfin chez de jeunes peupliers hybrides (Populus maximawiczii Henry x P. nigra L.
'l'v1N9') âgés d'un an, Tschaplinski et Blake (1995), ont constaté que les racines des cultivars intacts contenaient 1,8 fois plus de glucides que les racines d'arbres décapités.
Chaque année la racine fabrique un cerne de croissance à partir de son assise subéro-phéllodermique. Cette couche qui renferme le xylème et le phloème produit annuellement une nouvelle pellicule d'écorce vers l'extérieur et une nouvelle couche de bois vers l'intérieur. Les glucides sont entreposés majoritairement dans les rayons de parenchyme du phloème. Plus la racine grandit, plus la proportion de bois par rapport à celle de 1 'écorce est importante. De ce fait, les concentrations en glucides sont plus faibles dans les grosses racines que dans les racines fines, car ces dernières sont composées d'une plus faible proportion de bois.
1.2.3 Concentration foliaire en azote
L'azote est important dans la croissance des tiges et des feuilles. Il contribue entre autre au développement de la chlorophylle nécessaire à la photosynthèse. La quantité d 'azote varie selon les tissus, l'âge ou le stade de développement et la saison. Les plus fortes concentrations en N se trouvent dans les tissus composés principalement de cellules physiologiquement actives comme les tissus méristématiques et les feuilles, notamment dans l'enzyme Rubisco, impliquée dans la fixation du carbone durant la photosynthèse. Ces dernières pourraient contenir près de 40% de l' azote total de l'arbre. Pour la plupart des espèces feuillues, la concentration foliaire en azote a tendance à diminuer pendant la période d'élongation des feuilles. Il y a ensuite une augmentation à la fin de l'été, qui cesse quand la croissance stoppe et, qui est suivie d'une forte diminution jusqu'à ce que les feuilles tombent. Cette diminution s'explique par la translocation de 1 'azote contenu dans les feuilles vers les branches et le tronc (Kozlowski et Pallardy, 1997).
Après une défoliation partielle surE. globulus, (Turnbull et al., 2007), ont montré que la
concentration foliaire en azote avait augmenté significativement une semaine après l'intervention, et que l 'effet était persistant encore après quatre semaines. Malgré une forte relation entre
Amax
et 1 'azote foliaire, ce dernier ne s'était retrouvé ni sous forme de Rubis co ni de chlorophylle. L'augmentation de la capacité biochimique pour la photosynthèse en réponse à une défoliation partielle n'était donc pas fonction de la quantité de Rubisco ou dechlorophylle, mais d'une augmentation de l'activité spécifique de la Rubisco (Turnbull et al.,
2007).
1.2.4 Stress hydrique et isotopes du carbone
Les peupliers étant de grands consommateurs d'eau, leur implantation loin des cours d'eau peut poser problème, surtout en période de sécheresse estivale. L'élagage réduit inévitablement la surface foliaire d'un arbre, ce qui en cas de déficit hydrique, pourrait être un avantage en réduisant l'évapotranspiration et en minimisant la fermeture des stomates comparativement à des arbres non élagués (Polge, 1969). L'air contient naturellement deux isotopes stables 12C et 13C, respectivement présent à hauteur de 98,9% et 1, 1% du COz
atmosphérique (Kozlowski et Pallardy, 1997). L'acquisition de COz par les plantes commence quand le COz est diffusé à travers les stomates dans les espaces d'air internes de la feuille. La concentration interne en COz de la feuille est réduite par rapport à celle de l'atmosphère, ceci est dû à la ftxation du COz par les enzymes photosynthétiques. Parmi ces deux isotopes, la Rubisco, principale enzyme carboxylase pour les plantes de type C3, ftxe
préférentiellement le 12COz, car plus léger et donc plus facilement assimilable. C'est le phénomène de la discrimination isotopique du carbone (813C) (Kozlowski et Pallardy, 1997). Chez les plantes ~' il a été estimé que la discrimination isotopique pour le 13C est de l'ordre de -4%o lors de l'étape de la diffusion du COz à travers les stomates et de -29%o lors de l'étape decarboxylation (Farquhar et al., 1989). En été, lorsque les températures s'élèvent et que la disponibilité en eau dans le sol diminue, les stomates se ferment pour limiter la transpiration. Par conséquent, la concentration en COz à l'intérieur de la feuille est réduite et comme la Rubisco préfère le 12COz, le résidu d ' air à l'intérieur du mésophylle s'enrichit en
13
C0z. Dans ces conditions de stress hydrique, une plus grande quantité de 13C0z est donc ftxé par la Rubisco (Kozlowski et Pallardy, 1997). Les valeurs de 813C sont ainsi fréquemment utilisées pour estimer l'impact d'un manque d'eau pour les plantes. Ainsi, la mesure de la discrimination isotopique du carbone devrait nous permettre de savoir si les arbres élagués ont été moins sensibles aux périodes de sécheresse estivales compte tenu de leur surface foliaire réduite.
1.3 Effets de l'élagage sur la croissance
1.3.1 Accroissement en diamètre et défilement
L'élagage de branches mortes n'aura aucun effet sur la croissance. Par contre, l'élagage de branches vivantes aura un effet sur la croissance et dépendra de la quantité de surface foliaire enlevée. Chez les Eucalyptus et Acacias, une diminution de la croissance est observée à des taux d'élagage de 40 à 50% de la surface foliaire enlevée, alors que pour les conifères le même effet est observé mais pour des intensités d'élagage plus faibles, de 25 à 40% (Pinkard et Beadle, 2000). Plus l'intensité de l'élagage augmente, plus le ralentissement de la croissance en diamètre est fort. Keller ( 1979) dans son expérience sur 1' élagage de 1 'hybride de peuplier 1214 (Populus x canadensis Monch), a démontré un ralentissement de l'accroissement en diamètre dans la partie inférieure des troncs, mais au voisinage de la cime le ralentissement était moins marqué. L'élagage améliore donc la cylindricité des arbres (Funk, 1979 ; Keller, 1979 ; Neilsen et Pinkard, 2003) du fait d'une réduction du défilement (forme de l'arbre sur sa longueur). L'effet est d'autant plus marqué avec des intensités d'élagage plus sévères. Cependant seuls des élagages sévères (> 60% de la surface foliaire) avaient eu un effet dépressif marqué sur le tronc (Keller, 1979). En opposition, DeBell et al.
(2002) n'avaient pas constaté d'effets significatifs de l'élagage (50% de la hauteur) sur l'accroissement moyen en diamètre chez le clone 47-174 (Populus trichocarpa Torr. & Gray x Populus deltoides Bartr. ex Marsh.).
1.3.2 Accroissement en hauteur
Pour beaucoup d'espèces d'arbres, l'apex a la priorité pour la distribution du carbone comparé au reste de la tige, et ceci explique pourquoi la croissance en hauteur est moins affectée par l'élagage que la croissance en diamètre (Kerr et Morgan, 2006). Or, lors de ses travaux sur Juglans nigra, (Funk, 1979), a constaté que l'élagage avait favorisé la croissance en hauteur. Pour le clone I 214, la croissance en hauteur n 'était ralentie dans aucun des cas par l'élagage (Keller, 1979). Dans une expérience menée sur le chêne (Quercus robur L.), le hêtre (Fagus sylvatica L.) et le merisier (Prunus avium L.), Kerr et Morgan (2006) n 'observèrent à long terme aucun effet significatif de la taille de formation dans le jeune âge, sur la croissance en hauteur. Pour les eucalyptus, un taux d 'élagage supérieur à 50% a été
nécessaire pour en affecter la croissance en hauteur (Montagu et al., 2003 ; Pinkard et al.,
2004). Chez le jeune sapin baumier, il n'y avait pas d'effet de l'élagage sur la hauteur jusqu'à une amputation de 40% de la cime vivante. Au-delà de ce seuil une réduction de la croissance était observée (Margolis et al., 1988). Sur Pinus radiata, Neilsen et Pinkard (2003) ont constaté un effet immédiat et persistant pour des taux d'élagage de 60 et 75%.
1.4 Branches épicormiques
Un gourmand ou pousse épicormique, est une pousse qui apparaît sur la tige à partir d'un bourgeon latent (Fontaine et al., 2002). Chez le chêne sessile (Quercus petraea), il a été montré que ces bourgeons étaient tous d'origine proventive (Fontaine et al., 1999), i.e., qu'ils étaient mis en place lors du développement d'un bourgeon préexistant. Ils sont donc localisés à l'aisselle d'un organe foliaire et demeurent même après la chute des feuilles. Leur localisation est directement liée à la phyllotaxie i.e., à leur disposition sur le rameau. Ils sont qualifiés de primaires s'ils sont formés la même année que la pousse sur laquelle ils sont insérés, et de secondaires s'ils se sont développés ultérieurement sur une ramification de cette pousse (Fontaine et al., 2002). Chez d'autres espèces commeAcer pseudoplatanus L. et Salix alba L. des bourgeons épicormiques peuvent apparaître de façon spontanée suite à des blessures, ils sont alors appelés adventifs (Fink, 1983). Ils se développent à partir des tissus périphériques du tronc et leur localisation est indépendante de la phyllotaxie. Quelle que soit son origine, un bourgeon ne forme pas obligatoirement une pousse à l'année, il peut évoluer tardivement et rester au moins un an au stade de bourgeon (Fontaine et al., 2002). L'apparition importante de branches épicormiques dans un peuplement serait la conséquence d'un stress. Par exemple, la mise en lumière brutale des troncs suite à un épisode de chablis, une éclaircie ou un élagage de trop forte intensité. Chez le chêne pédonculé (Quercus robur
L.), il a été montré que l'élagage avait un effet à court terme sur l'apparition des branches épicormiques mais qui disparaissait sur une période de sept ans (Kerr et Hanner, 2001). Les auteurs n'ont trouvé aucune incidence de la saison d'élagage sur la production de gourmands. Cependant des observations apportées par Hubert et Courraud ( 1994) font part d'une repousse des gourmands nulle voire très faible lorsque les peupliers sont élagués en été durant la saison de végétation. Ceci indique que le choix approprié de la saison d'élagage pourrait aider à contrôler la production de branches épicormiques. Takiya et al. (2009) ont
examiné l'influence de l'élagage sur l'intensité lumineuse dans des peuplements de mélèzes hybrides (Larix gmelinii var. japonica x L. kaempferi). Ils ont illustré que l'élagage
augmentait la production de nouvelles branches épicormiques. L'émergence de ces branches épicormiques était attribuée à la rupture de la dominance apicale (Takiya et al., 2009).
1.5 Expérimentation
Le chapitre 2 présente une expérience dont l'objectif principal était d'expliquer les réponses en croissance de 4 clones de peupliers hybrides par les modifications physiologiques causées par la saison et l'intensité d'élagage. La première hypothèse était que les feuilles résiduelles des arbres élagués feraient de la photosynthèse compensatoire, afm de compenser la perte de feuillage. Cette photosynthèse compensatoire devrait être plus élevée pour les arbres élagués en été, car le stress environnemental provoqué par l'élagage à lieu lorsque que les feuilles sont présentes et actives. La seconde hypothèse était que l'élagage ferait baisser les concentrations en glucides dans les racines, car les arbres puiseront dans leurs réserves pour la formation de nouvelles feuilles suite à l 'élagage. D'autre part la concentration des glucides devrait être légèrement plus élevée pour les arbres élagués en été car la conservation de leur feuillage la première partie de l'été devrait leur permettre de constituer des réserves d' énergie plus importantes. De plus, le débalancement du ratio biomasse aérienne 1 biomasse racinaire causé par l'élagage devrait entrainer une augmentation de la concentration d'azote foliaire chez les arbres élagués. Enfm, les arbres élagués ayant une surface foliaire réduite, devraient avoir des feuilles moins stressées en eau et cela devrait se traduire par des valeurs de 813C plus faibles. Nous pensons également que la magnitude des réponses physiologiques devrait augmenter avec l ' intensité d'élagage. L'étude s'est déroulée dans une peupleraie localisée à New Liskeard, ON, dont les arbres ont été plantés au printemps 2007 à un espacement de 3,5 rn (816 tiges.ha-1
). Les élagages ont été
effectués à l'automne 2009, au printemps 2010 et, à l'été 2010, à deux intensités différentes; 1/3 et 2/3 de la cime v ivante, et un témoin non élagué. La photosynthèse, la concentration foliaire en azote, la concentration en sucres dans les racines ainsi que la discrimination isotopique du carbone ont été mesurées . Les clones ont été choisis sur la base de leur origine parentale différente. Il s'agit des clones 1079 (P. x jackii (P. balsamifera x P . deltoides)),
DN2 (P. deltoides x P. nigra) dont le suivi individuel de la croissance (hauteur et diamètre) a été effectué tout au long de l'expérience. Le chapitre 3 décrit la seconde expérience de l'étude dont l'objectif était de déterminer si les branches épicormiques représentaient un puits ou une source d'énergie pour l'arbre, mais aussi d'observer l'incidence de la saison et de l'intensité d'élagage sur leur production. Partant du principe que les branches épicormiques apparaissaient en réponse au stress infligé par l'élagage, leurs feuilles devraient avoir une photosynthèse équivalente à celle des feuilles résiduelles situées dans les deux tiers supérieurs de la tige, malgré leur position plus basse sur le tronc, ce qui pourrait expliquer la forte croissance de ces branches. Les branches épicormiques fournissant de la photosynthèse supplémentaire, la seconde hypothèse était que les arbres ayant des gourmands devraient avoir des réserves en TNC plus importantes. Enfin nous supposions que le nombre de branches épicormiques devrait être plus élevé chez les arbres élagués aux deux tiers, afin de compenser une perte du feuillage plus importante. Pour cette expérience une partie des arbres a été émondée (taille des branches épicormiques). La mesure de l'ensemble des paramètres physiologiques s'est fait de manière identique à la première expérience, de même pour le suivi de croissance. Enfin, les paramètres physiologiques et les caractéristiques morphologiques (surface foliaire spécifique et biomasse notamment) des branches épicormiques ont été mesurés. Le chapitre 4 présente quant à lui les conclusions générales de l'étude.
1.6 Bibliographie
Anderson, J.A., et M.K. Luckert. 2007. «Can hybrid poplar save industrial forestry in Canada?: A financial analysis in Alberta and policy considerations». The Forestry Ch ronicle, vol. 83, no 1, p. 92-104.
Bassman, J.H., et D.I. Dickmann. 1982. <<Effects of defoliation in the developing leaf zone on young Populus x euramericana plants. 1. Photosynthetic physiology, growth, and dry weight partitioning». Forest Science, vol. 28, no 3, p. 599-612.
Beadle, C., L. Pinkard, J. Medhurst, M. Cherry, et D. Worledge. 2004. Physiological responses to pruning in mixed species plantings: Blackwood management: L earning from New Zealand (Rotorua, New Zealand). 84-88 p.
Block, R.M., J.D. Knight, N.W. Booth, et K.C. Van Rees. 2009. «Nursery stock type, nitrogen fertilization and shoot pruning effects on the growth of juvenile hybrid poplar in Saskatchewan». Canadian Journal of Plant Science, vol. 89, no 2, p. 289-301.
Boysen, B., et S. Strobl. 1991. A Grower's Guide to Hybrid Poplar: Ministry of Natural Ressources, Ontario, 148 p.
DeBell, D.S., R. Singleton, C.A. Harrington, et B.L. Gartner. 2002. «Wood density and fiber length in young Populus stems: Relation to clone, age, growth rate, and pruning».
Wood and Fiber Science, vol. 34, no 4, p. 529-539.
DesRochers, A., S.M. Landhausser, et V.J. Lieffers. 2002. <<Coarse and fme root respiration in aspen (Populus tremuloides)». Tree Physiology, vol. 22, no 10, p. 725-732.
Dujesiefken, D., W. Liese, W. Shortle, et R. Minocha. 2005. <<Response ofbeech and oaks to wounds made at different times of the year». European Journal of Forest Research,
vol. 124, no 2, p. 113-117.
Duj esiefken, D., etH. Stobbe. 2002. «The Hamburg Tree Pruning System - A framework for pruning of individual trees». Urban Forestry and Urban Greening, vol. 1, no 2, p. 75-82.
Farquhar, G.D., J.R. Ehleringer, et K.T. Hubick. 1989. <<Carbon isotope discrimination and photosynthesis». A nnual Review of Plant Physiology, vol. 40, p. 503-53 7.
Pink, S. 1983. «The occurrence of adventitious and preventitious buds within the bark of sorne temperate and tropical trees». American Journal of Botany, vol. 70, no 4, p. 532-542.
Fontaine, F., P. Jarret, et J. L. Druelle. 2002. <<Étude et suivi des bourgeons épicormiques à l'origine des gourmands chez le Chêne sessile». Revue Forestière Française, vol. 4, p. 337-356.
Fontaine, F., E. Kiefer, C. Clément, M. Burrus, et J. L. Druelle. 1999. «Ünto geny of the proventitious epicormic buds in Quercus petraea. II. From 6 to 40 years of the tree's life». Trees, vol. 14, no 2, p. 83-90.
Portier, J. , B. Bigué, S. Morissette, J. Couture, et J. Ferland. 2011. Le Guide la populiculture au Québec: guide pratique sur la culture du peuplier hybride au Québec Québec: Réseau Ligniculture Québec, 124 p.
Funk, D. T. 1979. «Stem form response to repeated pruning of young black walnut trees».
CanadianJournal of Forest Research, vol. 9, no 1, p. 114-116.
Gagnon, H. 2006. L'élagage des peupliers hybrides: Élaguer pour enrichir nos forêts! Colloque sur l'élagage forestier (Maniwaki, QC).
Hart, M., E.H. Hogg, et V.J. Lieffers. 2000. <<Enhanced water relations of residual foliage following defoliation in Populus tremuloides». Canadian Journal of Botany, vol. 78, no 5, p. 583-590.
Heilman, P.E. 1999. <<Planted forests: poplars». New Forests, vol. 17, no 1, p. 89-93.
Hubert, M., et R. Courraud. 1994. Élagage et taille de formation des arbres forestiers, 2nd. Paris: IDF, 303 p.
Keller, R. 1979. <<Premiers résultats d'une expérience d'élagage du peuplier I 214». Annals of Forest Science, vol. 36, no 1, p. 59-92.
Kerr, G., et R. Hanner. 2001. <<Production of epicormic shoots on oak (Quercus robur):
Effects offrequency and time ofpruning». Forestry, vol. 74, no 5, p. 467-477.
Kerr, G., et G. Morgan. 2006. <<Does formative pruning improve the form of broadleaved trees?». CanadianJournal ofForest Research, v ol. 36, no 1, p. 132-141.
Kozlowski, T.T., et S.G. Pallardy. 1997. Physiology of woody plants, Second Edition. San Diego: Academie Press, 411 p.
Landhausser, S.M., et V.J. Lieffers. 2003. «Seasonal changes in carbohydrate reserves in mature northern Populus tremuloides clones». Trees - Structure and Function, vol. 17, no 6, p. 471-476.
Lonsdale, D. 1993. Choosing the time of the year to prune trees. Arboriculture Research Note. 117/93/PATH. Arboricultural Advisory and Information Service, Farnham, England: 6 p.
Margolis, H.A, R.R. Gagnon, D. Pothier, et M. Pineau. 1988. «The adjustment of growth, sapwood area, heartwood area, and sapwood saturated permeability of balsam fir after different intensities of pruning». Canadian Journal of Forest Research, vol. 18, no 6, p. 723-727.
Montagu, K.D., D.E. Kearney, et R.G.B. Smith. 2003. «The biology and silviculture of pruning planted eucalypts for clear wood production--a review». Forest Ecology and
Management, vol. 179, no 1-3, p. 1-13.
Neilsen, W.A, et E.A Pinkard. 2003. <<Effects of green pruning on growth of Pinus
radiata». Canadian Journal of Forest Research, vol. 33, no 11, p. 2067-2073.
Nowak, R.S., et M.M. Caldwell. 1984. <<A test of compensatory photosynthesis in the field: Implications for herbivory tolerance». Oecologia, vol. 6 1, no 3, p. 311-318.
Oosterbaan, A, E. Hochbichler, V.N. Nicolescu, et H. Spiecker. 2009. «Silvicultural principles, goals and measures in growing valuable broadleaved tree species».
Bodenkultur, vol. 60, no 3, p. 45-51.
Park, A, et E. R. Wilson. 2007. «E3eauti:ful Plantations: Can intensive silviculture help Canada to fulfill ecological and timber production objectives?». The Forestry
Chronicle, vol. 83, no 6, p. 825-839.
Pinkard, E.A, et C.L. Beadle. 2000. «A physiological approach to pruning». International
Forestry Review, vol. 2, no 4, p. 295-305.
Pinkard, E.A, C. Mohammed, C.L. Beadle, M.F. Hall, D. Worledge, et A Mollon. 2004. «Growth responses, physiology and decay associated with pruning plantation-grown
Eucalyptus globulus Labill. and E. nitens (Deane and Maiden) Maiden». Forest Eco/ogy and Management, vol. 200, no 1-3, p. 263-277.
Polge, H. 1969. <<Densité de plantation et élagage de branches vivantes, ou pourquoi, quand, et comment élaguer?». Revue Forestière Française, vol. 21, no Progrès techniques en sylviculture, p. 451-466.
Rytter, L., et G. Jansson. 2009. «Influence of pruning on wood characters in hybrid aspen».
Silva Fennica, vol. 43, no 4, p. 689-698.
Shigo, AL. 1984. «Compartmentalization : A conceptual framework for understanding how trees grows and defend themselves». Annual Review of Phytopathology, vol. 22, p. 189-214.
Soutrenon, A 1995. <<Période d'élagage et traitement des plaies sur feuillus». Ingénieries
-EAT, vol. 3, p. 3-12.
Takiya, M., H. Koyama, K. Umeki, M. Yasaka, Y. Ohno, I. Watanabe, et K. Terazawa. 2009. «The effects of early and intense pruning on light penetration, tree growth, and epicormic shoot dynamics in a young hybrid larch stand». Journal of Forest Research, vol. 15, no 3, p. 149-160.
Tschaplinski, T.J., et T.J. Blake. 1995. «Growth and carbohydrate status of coppice shoots of hybrid poplar following shoot pruning». Tree Physiology, vol. 15, no 5, p. 333-338.
Tumbull, T.L., M.A Adams, et C.R. Warren. 2007. <<lncreased photosynthesis following partial defoliation of field-grown Eucalyptus globulus seedlings is not caused by increased leafnitrogen». Tree Physiology, vol. 27, no 10, p. 1481-1492.
Uotila, A, et S. Mustonen. 1994. «The effect of different levels of green pruning on the diameter growth of Pinus sylvestris L.». Scandinavian Journal of Forest Research,
vol. 9, no 3, p. 226-232.
van Oosten, C. 2006. Hybrid poplar crop manual for the Prairie Provinces. Forest D evelopment Fund final report. Prince Albert, Saskatchewan Forest Centre: 232 p.
HYBRID POPLAR PHYSIOLOGICAL AND GROWTH RESPONSES TO PRUNING SEASON AND INTENSITY
2.1 Abstract
Pruning is part of the silvicultural practices recommended for poplar plantations in order to produce clearwood. Common wisdom proposes pruning during the summer season, however little is known on trees' physiological responses to pruning season and intensity. This study aimed at determining the effects of pruning season and intensity on growth and physiology of 4 hybrid poplar clones. Using three year old trees, we compared 9 combinations of pruning season (fall, spring and summer) and intensity (unpruned, 1/3 and 2J3 of crown length pruned). We showed that pruning increased net photosynthesis of residual foliage, nitrogen foliar concentrations and reduced root total non-structural carbohydrates reserves. Leaf carbon isotopie ratios (813C) values of pruned trees decreased compared to unpruned trees, reflecting their greater stomatal conductance and suggesting that pruned trees could have better drought stress resistance. Two growing seasons after pruning, 1/3 pruned trees had reduced height growth except for summer-pruned trees. Irrespective of the pruning season, pruning 2J3 of the live crown reduced both breast height diameter and height two years after treatments. Summer pruning increased foliar nitrogen concentrations, stomatal conductance and decreased leaf carbon isotopie ratios (813C) more than for trees pruned in faU or spring, suggesting better capacities to recover from pruning. We concluded that removal of 113 of the lower crown in summer was the best option to produce clear wood without compromising growth rates and physiological processes in hybrid poplar.
2.2 Introduction
The area of primary forests has decreased by more than 40 million ha since 2000 (F AO, 20 10). Establishment of fast growing plantations is considered as the main tool to produce more wood on reduced land areas by preserving the remainder of native forests while ensuring long-term timber supplies (Anderson and Luckert, 2007 ; Heilman, 1999). To maintain high growth rates, fast growing plantations require silvicultural interventions right from stand establishment to the final eut. The timing and intensity of silvicultural operations can significantly affect profitability, but more importantly the physiology and growth of trees. Pruning treatments to remove branches from the lower crown are usually clone to improve the shape of trees during the establishment period and /orto create knot free wood and increase the value ofboles (Hubert and Courraud, 1994).
Most pruning prescriptions are based on empirical data combining operational needs with tree growth responses (Keller, 1979 ; Neilsen and Pinkard, 2003 ; Shock et al., 2003 ;
Uotila and Mustonen, 1994). As a consequence, pruning prescriptions tend to be conservative and designed to ensure that growth is not affected by a particular level of pruning under most conditions (Pinkard and Beadle, 2000). Little is known, still, on the physiological responses to pruning which could help us recommend better silvicultural practices. Removal of foliage and branches should trigger gas exchange responses, modifications of carbohydrate and nitrogen (N) stocks, and a reduction in transpiration possibly enhancing trees' resistance to drought stress. Such responses are also likely to be affected by the season when trees are pruned, in turn affecting tree growth and vigour.
Pruning intensity (or height to which branches are removed) is usually determined by the desired length of saw logs and veneer log grades (Montagu et al., 2003). Recommendations
for hybrid poplars in Canada suggest reaching a 6-7 rn clear bole, in 3 to 5 lifts depending on tree growth, only removing one-third of the live crown at each lift (Boysen and Strobl, 1991 ; Portier et al., 2011 ; van Oosten, 2006).
Trees develop physiological responses to compensate pruning or defoliation such as compensatory photosynthesis, which is defined as an increase in the photosynthetic rates of
foliage of partially defoliated plants relative to foliage of the same age on undefoliated plants (Nowak and Caldwell, 1984). This mechanism also triggers an increase in the rate of the leaf development and in the longevity of existing leaves (Pinkard and Beadle, 1998a). An
increased utilisation of carbohydrates reserves is also a compensatory mechanism to support the production of new foliage (Tschaplinski and Blake, 1994), such as observed in the needles of pruned Pinus sylvestris (Langstrom et al., 1990). It has also been observed that pruning increased nitrogen concentrations in the remaining leaves of pruned trees (Pinkard et al., 1998 ; Tumbull et al., 2007). Such improved nutrient status in residual tissue may result
directly from modified root-shoot ratios (McNaughton, 1983). Moreover, pruning, by reducing leaf area and therefore transpiration, appeared to increase leaf water potential (Elfadl and Luukkanen, 2003). In situations of a water deficit, the reduction of foliage in pruned trees could avoid or delay stomata closure compared to unpruned trees by reducing the overall tree transpiration. These responses depend on species (Reich et al., 1993) and
generally increase with pruning intensity (Alcom et al., 2008; Medhurst et al., 2006).
Physiological processes are subjected to seasonal variations (Kozlowski and Pallardy, 1997), and physiological responses to pruning could also vary according to the season when pruning is clone. Many recommendations for pruning season are based on wound h ealing and insects or fungi damages at wound sites, and the extension of wood discoloration (Duj esiefken et al. , 2005 ; Dujesiefken and Stobbe, 2002 ; Lonsdale, 1993 ; Shigo, 1985 ;
Shigo et al., 1978 ; Soutrenon, 1995). However, prescriptions for pruning season in the
literature are contradictory. It is commonly recommended not to prune during the dormant season, because wound closure is slower, tree defenses are less operational and wounds are less effectively compartmentalized (Duj esiefken et al., 2005 ; Lonsdale, 1993). Hence, it has
been suggested that the best time to prune is at the end of winter when the tree defence system is about to become active, but before carbohydrate reserves are depleted by leaf flushing (Shigo, 1989), resulting in quick wound healing without spread of potential diseases (Soutrenon, 1995). Then again, prescriptions for pruning season depend on species and climate. In Europe, literature suggests pruning pop lars at the end of the summer (Bonduelle, 1971 ; Hubert and Courraud, 1994), while in Canada where the growing season is shorter, it is mostly recommended to prune poplars in late spring or early summer (Boysen and Strobl,
1991 ; Portier et al., 2011 ; van Oosten, 2006). These recommendations, however, do not
seem to be based on any physiological rationale or experimental data, but rather on conventional wisdom and traditional practice.
The main objective of this study was to assess physiological and growth responses of pruning season and intensity in different hybrid poplar clones. Gas exchange, root total non-structural carbohydrates and foliar nitrogen concentrations, and water stress were evaluated following three seasonal pruning (fall, spring or summer) and three pruning intensities (unpruned, 1/3 or 213 of crown height pruned). Since summer-pruned trees keep the totality of their photosynthetic area during the first part of the growing season, we expected higher root carbohydrates reserves hence stronger physiological responses to compensate for the loss of foliage. We anticipated that the magnitude of the response would increase with pruning intensity.
2.3 Materials and methods 2.3.1 Study sites
The study was conducted in a plantation established in 2007, at the New Liskeard Agricultural Research Station of Guelph University in North-Eastern Ontario ( 47°52'N, 79°66'W). The area is part ofHaileybury Clay forest section of the Great Lakes-St. Lawrence Forest Region (Rowe, 1972). From 1971 to 2000 annual precipitation for the region has averaged 819 mm (rainfall 625 mm, snowfall 195 cm) with an average daily temperature of 2. 8
oc
and an average of 2605 degree-days above 0ac
(Ville-I\.1arie station ( 47°21 'N, 79°26'W)), (Environment Canada, 2011). The regional surficial geology is characterized by lacustrine clays and sands from post-glacial Lake Barlow (Rowe, 1972), and soil type was a Humic Gleysol (Canada Soil Survey Committee, 1987).The site was ploughed in October 2006 and cross-cultivated with disks followed by herbicide application in spring 2007 before planting at a 3.5 rn x 3.5 rn spacing (816 stems.ha·
1
). Four hybrid poplar clones were chosen based on their different parentage: 1079 (Populus
x jackii (P. balsamifera x P. deltoides)), 747210 (P. balsamifera x P. trichocarpa), 915319
(P. maximowiczii x P. balsamifera) and DN2 (P. deltoides x P. nigra). The planting stock
local fertilization with 18-23-18 (N-P-K, 110 g.tree.1). Yearly weed control was done by
cross cultivation with disks followed by herbicide application (Roundup™).
Trees were planted in a randomized block design with three blocks (replicates ), each containing four plots of 100 trees (10 rows x 10 trees), one for each clone. Each plot was randomly divided into 3 pruning seasons (October 2009 = fall, March 2010 = spring or June 2010 = summer) and 3 intensities (unpruned, 1/3 or 213 of crown length removed). One row of trees was used for each treatment combination (9). Pive trees of each row were randomly selected for the study (pseudo-replicates; n = 540). If epicormic branches sprouted from the
stem of pruned trees during the two growing seasons after pruning, they were removed with respect to the pruning treatment they were allocated.
2.3.2 Field sampling
Diameter at breast height (dbh) and height were measured in fall 2009 and at the end of the 2010 and 2011 growing seasons. Stem volume was calculated from the following equation developed by Boysen and Strobl (1991) then converted into volume per hectare:
V = (exp (-1.064079 + 1.562891 * ln(D) + 0.101423 * ln(H)) * 1.013914) / 1000
where Vis the stem volume in m3, D the dbh in cm and H the tree height in m.
Instantaneous net photosynthesis and stomatal conductance was measured at weekly intervals with a CIRAS-2 portable infra-red gas analyser (PP Systems, Amesbury, MA). Repeated measurements on the same tree were done weekly between the second and fifth week following the summer pruning. Measures were done on leaves located in the upper third of the canopy, using the youngest fully expanded leaf. Trees were selected randomly and one measure was made for each treatment i.e., n = 108 for each week, except at week 2 where
only two blocks (replicates) were measured due to a battery problem. One root sample per treatment combination (n = 108) was collected for determination of total non-structural carbohydrates (TNC) concentrations at the end of august 2010 by cutting a disk from one of the main roots. Root disks of similar size were collected 30 cm from the trunk from roots directly connected to the base of the tree.
Four to six leaves were collected from the upper third of the canopy of each tree in July (five weeks after the summer pruning) for nitrogen (n = 108) and carbon isotopie discrimination (813C) analyses. For the 813C analysis, two types of leaves were distinguished for the trees pruned in summer; leaves formed before or after the pruning (n = 132).
2.3.3 Laboratory work
Roots samples for TNC analysis were oven-dried at 65 °C until constant weight, and ground to pass a 40 mesh screen of a Wiley mill (model 3383-L10; Thomas Scientific, Swedesboro, NJ). Soluble sugars were extracted by boiling three times in 80% ethanol at 95
oc.
Phenol-sulfuric acid was used to analyze the ethanol extract for total sugar concentrations. Starch was digested using an enzyme mixture of a-amylase and amyloglucosidase followed by the colorimetrie measurement of the glucose hydrolysate using a peroxidase-glucose oxidase-o-dianisidine reagent (Chow and Landhausser, 2004).Leaf samples were oven-dried at 65
oc
until constant weight then ground fmely with a Mixer Mill :MM301 hall grinder (Retsch Inc. , Newtown, PA). Carbon isotopie composition (813C) and total nitrogen concentration were determined using a Costech Elemental Analyzer 4010 CHNS (Costech Analytical Technologies Inc., Valencia, CA) at the Soil Biochemistry Laboratory, University of Alberta, Edmonton. Samples were flash combusted at 1800 °C. The resulting gas were separated on a 2 rn x 6 mm OD stainless steel Porapak QS 80/100 mesh packed chromatographie column and detected quantitatively by a Thermal Conductiv ity Detector.The isotopie composition of the sam pl es was calculated from:
Ù13C = ( ( Rample / R.tandard ) - 1 ) X 1000
where Rample and R.tanctarct are the ratios of 13C/12C in the sample and standard, respectively. Spring wheat grain was used as the working standard, with a carbon isotope composition of-23.4 7%o relative to Pee Dee Belemnite.
2.4 Statistical analyses
All statistical analyses were conducted with R software version 2.15.1 (R Development Core Team, 2012). Linear mixed-effects models were used to model treatment effects on each response variable. To reckon with the plantation design, all models included random effects that consisted of block, and clone nested in block. The lme function from the nlme package (Pinheiro et al., 2012) was used to estimate the parameters of all models, whereas
package AICcmodavg was used for model selection and multimodel inference (Mazerolle, 20 12). For each response variable, a model set was built containing candidate models from which model selection was clone using Akaike information criterion corrected (AICc) for small sample sizes (Burnham and Anderson, 2004). Differences in AICc values (~AICc) were calculated for the respective models relative to the "best" model, i.e., the model with the lowest AICc (Table 2.1). Models with ~AICc < 2 and high Akaike weights (w, interpreted as probabilities) were deemed to have the grea test statistical support (Burnham and Anderson, 2004). If one model had an Akaike weight of more than 80%, it was considered as the most parsimonious model of all tested models. Predictions and SE associated were then calculated using the function predictSE.lme (Mazerolle, 2012). Otherwise, we calculated model-averaged estimates and unconditional 95% confidence intervals with multimodel inference (Mazerolle, 2006). Ninety-five percent (95%) confidence interval which did not include 0, indicated a strong effect.
The most biologically relevant models were built for our analyses with the following factors: pruning intensity, pruning season, clone, week (weekly measurement for gas exchange variables), year (year of measurement for growth variables) and leaf age and their interactions. Since a limited number of models should be compared (Anderson and Burnham, 2002) ten models were compared for physiological variables and 26 for growth analyses. For more clarity, Table 2.1 only presents models which had the greatest statistical support i.e.,
with ~AICc < 4 (Burnham and Anderson, 2004).
Model sets D, and H correspond to growth variables dbh and height, while model sets PN, N Jeaf, and DELTA _1 correspond to the physiological variables net photosynthesis, leaf
nitrogen and leaf carbon isotopie ratio, respectively. Parameter tree was incorporated as a random effect in the model sets with repeated measures (model sets PN, D and H; Table 2.1).
To consider the influence of root diameter on TNC concentrations (DesRochers et al.,
2002), we recorded root diameter for each sample and included it in the models as a covariate (model set TNC). In para11el, growth data were analysed using initial size of trees as covariates (fall 2009). Stomatal conductance (model set GS) was log-transformed to normalize residuals and to improve homoscedasticity, results and predicted values are presented in back-transformed units.
For 813C analyses, a separate data set was created to account for the two types of leaves; those formed before (S_l) or after (S_2) pruning (old vs new leaves), these data being only available for summer pruning at intensities 113 and 2J3. Model set DELTA_2 was built to analyse the effects of pruning treatment, age of leaves and clones, on the 813C relative to the summer pruning. We used multiple comparisons of means (Tuckey's tests) to analyse the interaction between leaf age and pruning intensity. A significance level of P = 0.05 was used.