UNIVERSITE DE NEUCHATEL
INSTITUT DE ZOOLOGIE
S63
Biologie â
JIxodes ricinus L. sur le Plateau Suisse
Une contribution à l'écologie de ce vecteur
-par
FRANCOIS GIGON
Thèse présentée à la Faculté des Sciences de l'Université de Neuchâtei
pour obtenir le grade de docteur es sciences
-1985-UNIVERSITE DE NEUCHATEL INSTITUT DE ZOOLOGIE
Biologie & Ixodes ricinus L. sur le Plateau Suisse
-Une contribution à l'écologie de ce
vecteur-par
FRANCOIS GIGON
Thèse présentée à la Faculté des Sciences de l'Université de Neuchâte! pour obtenir le grade de docteur es sciences
-1985-IMPRIMATUR POUR LA THÈSE
Biologie d [Ixodes ricinus[L. sur le plateau
s u i s s e . - Wie. contribution à 1 'écologie de
ce vecteur.
de M o n s i e u r François.Gigon
UNIVERSITÉ DE NEUCHATEL
FACULTÉ DES SCIENCESLa Faculté des sciences de l'Université de Neuchâtel, sur le rapport des membres du jury,
Messieurs A. Aesçhlimann, E. Hess
rB.__ Horning
(Berne) et W. flurgdorfer (Hamilton, U.S.A.)
autorise l'impiession de la présente thèse.
Neuchâtel, le ....„?
s.<?PÇeraùre ..1985
Le doyen:
Jf '?> /
T A B L E D E S N A T I E R E S + + + + + + + + + + + + + + + + + + + + + + + + +++*• + + + + + + + I N T R O D U C T I O N
R
m
IV
VVI
VIL
VM
SV57£flA7IClU£
ÙIS7RIBU7ION ÇtOÇRhPHIQat
cycu
HOTCS
IN7CRC7 CPIDtRIULOQIQU£
BU7S DU TRAVAIL
CÜN7CX7C DE LA LI77CRA7URC
CONCCP7ION QeNERALe
£7
PRCSlNlA7I0N
M A T E R I E L E T M E T H O D E S
1 . Capture 2 . Elevage 3. Marquage
LA 7IQUe (I. *j.cJ.nu./>)
1313
13
13
E LES 7CRRAlXS 'U
1IKPCRIPiCNTATION 15
1 . Choix 15
2. Situation 1 5 3. Historique 1 6 4. Climat 16 5. Végétation et phytosociulogie 1 7 6. Faune 187. Situation des diverses expérimentations IB
m L'AC7ivne. ne QUCTC 27
1. Observation 27
2. Drapeau 2 7
3. Battage 27 4. Systèmes de capture sélectifs en hauteur 27
5. Lâchers de tiques 30 B. Sélection du support naturel 3D
7. Supports artificiels 31 B. Enceintes réelles à hauteur de supports imposée 31
9. Enceintes virtuelles à choix de hauteurs 32
10. Interruption provoquée de l'activité de quête 33
11. Influence de l'état hydrique 3a 12. Modèles de laboratoire 34 13. Terrarium 36
IV PHAStS CA'DOÇjCS
5 1
1, Hivernage ^ l a) Extraction 51 b) Flottation 52 c) Tiques libres 5 2 d) Tiques en sacs 52s) Coupe de sol en terrarium 54
3. Clue et ponte
a) Tiques en sacs
b) Technique "d'Ariane"
il.
Prédation
54
55
55
56
MIGRATIONS HORIZOHl7AL£S
67
1. Lâchers de tiques marquées
2. "Tiquodromes" {enceintes virtuelles à choix de hauteurs]
3. Femelles "en Ariane"
67
67
67
VI
neiiOROLOQie.
69
1. Enregistrements officiels
2. Enregistrements personnels permanents
3. Mesures microclimatiques sporadiques
h.
Stockage des données
5. Exploitation informatique
6?
69
69
70
71
vu
RCSIS7ÂNC& AU 7R0ID
75
R E S U L T A T SP)LIiOROLOQIl
791. Climat régional
2. Climat local
3. Microclimat
79
79
60
E
SELECTION DU SUPP0R7
9 11 . Supports naturels 91
2. Supports a r t i f i c i e l s 91
JZT HAUTEUR DL QUE7E 93
1 . Observation directe 93
2. Brouette 93
3. Enceintes à choix de hauteurs (tiquodromes) 94
h. Enceintes à hauteur imposée 95
5. Battage 96
IV POSlIlON DE ÛUE7L 1 0 3
1 . Par rapport au sommet du support 103
2. Orientation dans l e plan v e r t i c a l 103
3. Orientation dans l e plan horizontal 104
4. Par rapport aux f e u i l l e s 105
V CARACTERISTIQUES 7ICiPORLLLLS DE LA ÛUE7L 109
1. Journalières 109
2. Saisonnières 109
3. Annuelles 112
VI EVOLUTION DCS HAUTEURS DL OUETL 119
1 . Saisonnière 119
2. Journalière 119
VE
INfLUlNCL DtS TACTtURS CiITtOROLOQIUUtS
123
1 . Traitement s t a t i s t i q u e 123
2 . Cas p a r t i c u l i e r de l a p l u v i o s i t é 126
Vm INTLUtNCt DtS TAC71URS INTtRNtS 137
1 . Etat hydrique 137
2 . Perturbation de l ' a c t i v i t é 139
IX PÌ1QRATI0NS HORIlONlALtS 14 3
1 . Lâchers de tiques marquées IA3
2. Tiquodromes 14 3
3. Foyers de larves 143
4 . Femelles "en Ariane" 144
X DISTRIBUTION NORIZONTALt 145
1 . Dans l e sol 145
2. Durant l ' a c t i v i t é 145
Xl C0npÛR7£ntN7 Dt DtBUT £7 Dt TIN Dt QUtIl 149
XH NlVtRNATlON 151
1 . En terrarium 151
2 . En place (sans lâchers) 151
XM
RCSlSlANCt AU TkOIh
161
XIV
PHASES ENdOQEiS
1 6 5
1 . En p l a c e 2 . F e m e l l e s " e n A r i a n e " 3. En sacs
165
165
166
XVPKLdAIlON
1 7 1
D I S C U S S I O NLA QUÊTE ve L'HOTE
177
1. Le choix du support
2. La hauteur de quête
3. La position de quête
4. Périodes et rythmes dé quête
177
177
179
179
a
METEOROLOGIE ET ACTIVITE
183
1. Sélection des facteurs
2. Les influences de la météorologie
1B3
1B4
m
IN7LUENCE DES 7AC7EURS INTERNES
189
1 . L ' é t a t hydrique
2. La p o t e n t i a l i t é de quête e t l ' â g e physiologique
189
190
IV
LA PHJNOLOQie
193
1. Les theories de la littérature
2. Les particularités de nos régions
3. L'influence des hôtes
193
194
197
US CAaSCS DL MR7AL171
190
1. Le froid hivernal
2. La prédation
3. Le parasitisme
4. Bilan général
199
201
202
203
VI
LA RLPAR7I7I0N HORIZONTALS,
205
1. Type de distribution
2. La part des phéromones d'agrégation
3. Les migrations
2 0 5
2 0 5
206
VR
VM
ri£7H0D0L0Gl£
CONCLUSION
2 0 7
209
S Y N T H E S E E T R £ 5 U M E221
E n g l i s h summary2 2 5
R E P l E R C ï E P l E N T S229
B I B L I O G R A P H I E2 3 1
T A B L E O E S I L L U S T R f t T I O N S
++++++++++++++++++++++++++++++++++++++++++++4
1 . Figures
Figure Page Figure Page Figure Page
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
m
25
26
27 "'
28
29
30
31
32
33
34
35
8
ID
10
14
19
20
25
37
38
38
39
39
40
41
41
42
42
43
43
44
44
46
47
47
48
48
49
49
50
50
57
58
59
59
60
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
61
68
69
70
60
62
62
63
63
64
64
65
65
72
84
85
87
87
88
88
89
89
90
9B
99
100
101
113
113
114
114
115
116
117
117
118
118
120
120
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
BB
89
90
91
92
93
94
95
96
97
98
99
100
101
128
128
133
135
135
136
140
141
147
148
155
156
156
157
157
158
158
159
159
162
167
168
172
173
212
213
214
215
216
217
ZlB
2. Tableaux
Tableau Page
4
5
6
7
B
9
10
11
12
13
14
15
16
17
IB
19
20
21
22
23
24
25
26
27
28
29
30
14
21
22-23
24
37
40
45
57
56
61
62
72
72
73
83
84
85
86
90
97
97
102
102
106
106
107
107
Tableau Page
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
108
108
115
116
121
129
130
131
132
133
134
136
140
147
155
160
162
163
167
169
172
210
210
211
211
212
213
219
220
220
INTRODUCTION
I Systématique
La position systématique d'i",
/LÌCÀJULÓ
est précisée, parmi les
Arthropo-des, sur le Tableau 1. Cet organisme fait partie de la super-famille des
Ixodo-ixLe.a,
ou tiques, qui comprend quelque BDO espèces (Hoogstraal et Aeschlimann,
1982). Elles se répartissent en deux familles principales : les
Angcuidae.,
ou
"tiques molles", sans scutum et les
IxocLidae.,
ou "tiques dures", dont
l'anato-mie externe présente toujours un scutum, ou bouclier dorsal. C'est à cette
der-nière famille qu'appartient le genre
Ixodes,
le plus riche parmi les tiques,
avec ses 212 espèces.
H Distribution géographique
I, /Ltcirnu,
est largement réparti dans toute l'Europe, dont il déborde
sur 1'U.R.S.S. et l'Afrique du Nord (Algérie et Tunisie). Parmi ce large
é-uentail de distribution, cette tique sélectionne rigoureusement les milieux
qu'elle occupe. Une fois satisfait le critère des hôtes, qui doiuent se trouver
en suffisance dans le biotope considéré, météorologie et micrométéorologie
se-ront à la base de cette sélection :
- La température ne doit pas être supérieure à 35
0C, dans l'air et le
sol, ni s'abaisser au-dessous de -10
0C dans les couches superficielles du sol
(Hc Leod, 1935a, 1936).
- L'humidité relative de l'air doit excéder BO It (fie Leod, 193S), celle
du sol se tenir proche de la saturation, à plus de 95
%
(Lees, 1946).
Bien que les deux types d'exigences ne soient pas indépendants l'un de
l'autre, un léger déficit hydrique pouvant être mieux supporté par une plus
basse température (PIc Leod, 1935a), c'est tout de même l'humidité relative qui
s'impose en tant que facteur limitant. Dès lors, le .rôle de la végétation
de-vient primordial : on peut établir une relation tique/phytosociologie (Gilot
et al., 1975). Cette relation va bien entendu varier d'une contrée à l'autre,
en fonction du climat général : présente dans les collines pâturées de
Grande-Bretagne, la tique ne se rencontre che2 nous que dans des milieux couverts. En
plaine, les sous-bois sont leurs biotopes de prédilection, mais buissons, haies,
bosquets et lisières peuvent aussi les héberger (Aeschlimann, 1972).
m Cycle
/.
A-icû/ULo
est une tique dite "triphasique" : elle possède trois stades
(larve, nymphe et adulte), chacun devant trouver un hâte, s'y fixer à l'aide
du rostre (ensemble formé par les deux chélicères et l'hypostome) et absorber
un repas de sang (Fig. 1 ) . Après s'être laissée
tamber.au
sol, elle subit une
période de digestion qui sera suivie de la mue (larve et nymphe) ou de- la
pon-te (femelle) donnant naissance au stade suivant. Le cycle est donc la
succes-sion de phases libres exoqées (recherchede l'hâte), de phases parasitaires
(trophiques) et d'un deuxième type de phases libres, les phases endoqées
(re-pos, diapause, rrorphogenèse et développement) : Fig 2,
IU Hôtes
Plutôt que de reprendre la liste exhaustive des espèces animales ayant
hébergé l'un ou l'autre des stades d'-f.
/LLCÀJUIA
(Arthur, 1963; Aeschlimann,
1972), nous préférons donner ici une idée du rôle que joue la faune vertébrée
de notre principal terrain d'étude (le Staatsmald, Fig. 5, Fl.E), en tant
qu'hô-tes. La liste de ces hôtes potentiels (Tableau 7) doit en effet être commentée,
car si
I. AÀ.cÀJuu>
est une tique remarquablement dépourvue de spécificité
para-sitaire, les différents stades n'en montrent pas moins des préférences non
né-gligeables, résumées sur le Tableau 2 (Aeschlimann, 1972; Nilne, 1949 b ) . Il
faut encore ajouter que le rôle des insectivores (Hérisson mis à part) et des
oiseaux est en principe modeste dans la nutrition des larves, mais qu'il peut
devenir important en cas de pénurie de petits rongeurs, dont les populations
sont soumises à de fortes fluctuations (Nosek et al., 1973). L'Homme est un
cas très particulier : quiconque en a fait l'expérience sait qu'il ne laissera
pas volontiers une tique se gorger sur lui I Lorsqu'il intervient dans le
cy-cle biologique de la tique, c'est éventuellement pour en accepter un
micro-organisme, mais jamais pour le transmettre. Il représente par conséquent un
cul-de-sac épidémiologique.
Une dernière remarque au sujet d'I
/UCÙTUIA
adulte : si chaque sexe se
rencontre sur l'hôte, seule la femelle est capable de se gorger. Encore qu'il
puisse exceptionnellement se fixer et absorber un frugal repas, comme on
l'ob-serve en élevage, le mâle n'a pas besoin de se nourrir : contrairement à la
plupart des
Ixottidae.
(Oliver, 19B2), les
IKOCUA
sont capables d'élaborer
tou-te la Spermatogenese sans l'intou-tervention d'un repas sanguin, et l'accouplement
en est aussi indépendant. De fait, il a lieu dans la végétation, avec la
fe-melle également à jeun, dans 40 à 60
%
des cas (Graf, 1975). Le restant des
femelles sera fécondé sur l'hôte, d'où la présence des deux sexes sur ce
der-nier : ce comportement du mâle n'est donc pas que la manifestation d'un atavisme.
V Intérêt épidémiologique
De la simple démangeaison à des maladies dont l'issue peut être fatale, les tiques sont à l'origine de toute une série de désagréments à l'égard de l'Homme et des animaux.
Les lésions mécaniques (déprédation des cuirs, voies d'infections se-condaires), l'anémie (grosses pertes de sang dues à une charge parasitaire trop élevée), les réactions allergiques et diverses toxicoses entraînent des baisses de productivité considérables dans les élevages. Plus graves sont en-core les maladies dues à un agent pathogène transmis par la tique telles que les tristement célèbres theilerioses et babesioses (fleschlimann, 197B). Les êtres humains peuvent souffrir de maux comparables qui ne sont, encore au-jourd'hui, pas toujours bien connus.
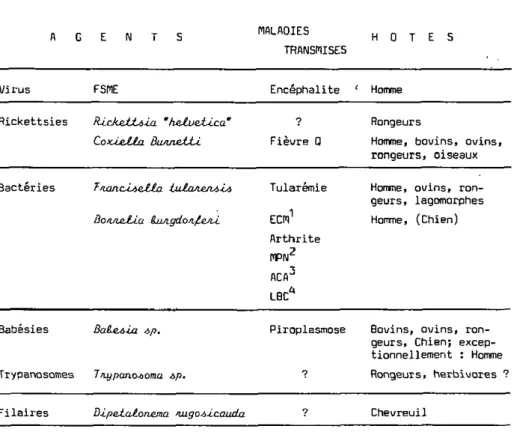
La liste, déjà longue, des agents pathogènes dont les tiques sunt vectri-ces, s'allonge constamment (Aeschlimann et al.. 1979). Ainsi, deux affections dermatologiques jusqu'ici imputées à des toxines sont en réalité le fait d'une Borrélie qui vient d'être décrite (Barbour et al., 19B3; Burgdorfer et al., 1983; Johnson et al-, 1984). Le Tableau 3 ne présente que les quelques organis-mes pathogènes transmis en Europe par /. ticjjuu.
Dans ces conditions, on comprendra l'intérêt toujours renouvelé que sus-cite l'étude des tiques au travers de leur incidence économique et de leur rôle épidémiologique.
Un concept aussi vaste que 1'epidemiologie englobera donc des domaines de recherche aussi variés que la clinique, l'immunologie, la faunistique, la culture microbienne, l'écologie ou le comportement des tiques, pour n'en citer que quelques uns. C'est précisément dans le cadre de ces deux derniers domaines que se situe la présente étude.
VI Buts du travail
La Fig. 2 révèle qu'une grande partie du cycle se déroule en dehors de l'hôte, dans deux types de milieux différents. Si l'on suit un individu du-rant l'un de ses stades (Fig. 3 ) , an s'aperçoit qu'il est soumis au climat des couches superficielles du sol dès sa chute de l'hâte ("drop-off"). Il en sera ainsi jusqu'à sa première apparition dans la végétation, peut-être des mois plus tard, ou l'année suivante. A ce moment débute la recherche de l'hôte, que nous désignerons par le terme "quête" (ou activité de quête), qui soumet
1 : Les termes soulignés dans ce chapitre composent une terminologie qui sera désormais régulièrement employée.
la tique aux conditions régnant à l'air libre. La tique gagne alors un
pro-montoire, le support (ou support de quête). La majeure partie de la quête s'y
passe sans mouvement, la première paire de pattes repliée ("resting posture",
Lees, 19ÛB). La détection d'un hâte de passage engendre une excitation
visi-ble sur la'première paire de pattes : tout d'abord étendues en "V", elles
sont ensuite balancées d'avant en arrière ("waving") si le stimulus se
pour-suit. Contrairement à d'autres espèces (0.
^auignyi,
certains
Am&Jifomma;
Ulaladde
et Rice, 1982) qui "chassent à courre",
I. ilcÀmu,
est "un chasseur à l'affût".
Des phases de repos peuvent interrompre la quête, lorsque la tique descend de
son support.
Ainsi, dans l'ensemble deson cycle, qui peut s'étendre sur deux à six
ans (Donnelly, 1977), la tique ne passera-t-elle que trois semaines au maximum
sur l'hôte et sera soumise tout le reste du temps aux conditions du milieu
extérieur (air ou sol).
Quantitativement, la vie libre est donc prépondérante dans l'étude
épi-démiologique de ce vecteur potentiel qu'est
1. /UCÀJUIÒ.
L'aspect que nous nous
proposons d'étudier peut se résumer en une phrase :
Quelles sont les caractéristiques des différentes phases de quête et de
repos, comment s'articulent-elles entre elles et quels sont les éléments qui
déterminent ces articulations ?
Plus précisément, l'étude porte sur deux grands volets :
1° La quête : déterminisme, temporisation, localisation et mouvements
verticaux en corrélation avec le milieu; accent particulier sur la
météorologie.
2° Le repos : localisation et survie; expérimentation axée
principale-ment sur l'hivernation.
En réalité, le travail s'est émaillé d'un certain nombre de manipulations
et de résultats complémentaires qui nous aideront à brosser un portrait un tant
soit peu plus fouillé des activités d'Z.
IÌCÀJUIA
durant sa vie libre.
V U Contexte de la littérature
Parmi tous les aspects que recouvre 1'epidemiologìe d'un vecteur d'agents
pathogènes, l'écologie et l'éthologie ne bénéficient guère de priorités.
Ce-pendant, lorsque ce vecteur est étudié de longue date, comme c'est le cas
d'.T.
sLicJjuLòt
une littérature relativement abondante peut être consultée sur
ces sujets. Nous ne citons ici que celle qui est à l'origine de notre étude.
- En Grande-Bretagne, toute une série de travaux ont été menés dans un
domaine que nous pourrions désigner par "éco-éthologie" dans le terrain et au
4
laboratoire : Mc Leod (1932, 1934 a et b, 1935 a et b, 1936, 1936, 1939), m i n e (1913, 1944, 1945 a et b, 1946, 1947 a et b, 1949 a et b, 1950 a et b ) , Lees (1946, 1969), Lees et Nilne (1951), Gray et al. (1978) et finalement Gray (1980, 1981 » 1982, 1984) en couvrent la grande majorité.
- En Europe continentale de l'Ouest : Tatze (1933), Bauch (1971, 1972), Walter (1979), Gilot et al. (1975) et Perez et Rodhain (1977) ne présentent que des données rares sur les phases libres, à l'exception de la phénologie. Les travaux suisses de Plermod et al. (1973, 1974, 1975), de Graf (1974, 1975, 1976, 1978 a, b et c) et de Graf et al. (1978, 1979) faisant suite à ceux d'fleschlimann (1972) revêtent un intérêt supplémentaire, dans le sens où ils sont tous issus d'observations menées sur le Plateau Suisse et plus précisé-ment au Staatsuald.
- En Europe de l'Est et en U.R.S.S-, ce sont surtout Cerny (1959, 1961, 1962), Daniel et al. (1972, 1976, 1977), Daniel (1978), Dusbabek et al, (1971), Slonov (1963) qui ont travaillé sur les phases libres, l'hivernage en parti-culier, alors que Selozerou (1982) fournit de précieux renseignements sur les phénomènes de diapause.
Dans le contexte de cette littérature, notre étude s'inscrit donc en parallèle à celles des auteurs de Grande-Bretagne et en complément à celles menées sur le Continent. Elle est la continuation des travaux suisses cités précédemment. Si l'aspect original de travaux complémentaires est évident, il faut préciser ce qu'il en est du caractère "parallèle" : si Vl. JLÌCMWA
des lies Britanniques appartient bien à la même espèce que le nôtre, son ha-bitat est très différent. Toujours inféodé à des milieux couverts en Suisse
(fleschlimann, 1972), il se trouve principalement dans les pâtures collinéennes en Grande-Bretagne. Cette nuance écologique fondamentale entraîne des diffé-rences épidémiologiques que nous avons voulu mettre à jour, en ayant plusieurs fois recours à des techniques originales.
Nous ne présenterons pas plus avant cette littérature, qui sera abon-damment commentée en relation avec nos résultats dans la partie "discussion" de ce volume.
UHI Conception générale et présentation
De propos délibéré, l'essentiel du travail est axé sur le terrain. Les manipulations strictement de laboratoire n'interviennent qu'en tant que corol-laire, destiné à certifier expérimentalement la véracité d'observations réali-sées dans le milieu naturel, ou en tant qu'épreuve pour telle hypothèse venue du terrain, mais difficile à tester dans ce dernier.
La fréquentation régulière du milieu naturel mène de soi à l'accumula-tion d'un certain nombre de petites observal'accumula-tions, difficiles à présenter sous forme de résultats, mais qui contribuent, du moins est-ce notre avis, à la compréhension de l'organisme vivant et de son comportement au sein de son bio-tope. C'est"pourquoi le chapitre "Résultats" peut, à l'occasion, prendre des allures de petite "Discussion". Nous avons tenté de maintenir ces digressions au minimum.
La littérature écologique relative à I . /LLCÀJULA est largement évoquée dans le chapitre "Discussion". Il s'agit d'y atteindre deux objectifs :
1° Comparer nos résultats à des données précédemment recueillies, les expliquer grâce à des connaissances précédemment acquises et les intégrer dans ce contexte.
2° Sans faire figurer une bibliographie exhaustive ni aboutir à un tra-vail de compilation, dresser un panorama des connaissances actuelles de "l'éco-éthologie" d'i". nÀ-clmiò.
Dans ce double but, nous présentons la "Discussion" sous forme synthé-tique, par le biais de quelques thèmes recouvrant naturellement nos propres résultats.
Nota : les renvois en cours de texte mentionnent les numéros figurant dans la Table des Matières, précédés d'une lettre s'il s'agit d'un passage hors cha-pitre en cours. I : Introduction M : Matériel et Méthodes R : Résultats D : Discussion C : Conclusion
6
Embranchement
A H HROPOD A
Classe
Sous-classe
Ordre
Sous-ordre
Super-famille
fly/iiapoda, 1 ii£o&JjLomo/Lpha
Cle/iOAtoma, PycnogonUda
Ann/ieù., Opttion&A, Scoip-ion&ó
PAeudLQòcoipion&ò, SaLLfugae.,
tUopìgae., PalpigtacU,
Amtly-pygae.1
RÀ.cJjuxteJ-NotoàLigmatai 7'eVioàLigmata,
Oiyptoòtigmata, ft&AOALLgmaLa,
AóLigmaia, PioòLigmata
Ixodo-id&a
Famille
Sous-famille
Genre
Espèce
Ajig(Lòìda&, Nidjtattl&ttldae.
Am&£yomnUnae.t äyatomnujiae.
RJiip-lc&phati/iae.,
Haèmaphy-•òatinae.
211 autres espèces
Tableau 1 : Position systématique d'I, sUcUniu, parmi les Arthropodes.
Fjq. 1 : Les trois stades d'I.
iicinui.
De gauche à droite et de haut
en bas : larve à jeun, larve gorgée, nymphe à jeun, nymphe gorgée,
fe-melle et mâle à jeun, fefe-melle gorgée.
H O T E S
Oiseaux
Insectivores
Rongeurs
Lagomorphes
Carnivores sauvages
Ongulés
Chien
Homme
Adultes
++
+++
+Nymphes
++ + + + + + +Larves
+ +++ +M
M
+ +Tableau 2 : Fréquentation des hôtes par les divers stades d'Z.
/ticunu-i.
(•) : exceptionnelle ++ : normale
MALADIES TRANSMISES H O T E S V i r u s FSPlE E n c é p h a l i t e Homme R i c k e t t s i e s ÎUckettàla "helvetica" CoxJjeJtta Bann&iÀÀ. ? Rongeurs F i è v r e Q Homme, b o v i n s , o v i n s , r o n g e u r s , o i s e a u x B a c t é r i e s f'lanclAella iulwierU-Li BOIJIJEJLLU (Uuigdo/i^ejiJ, Tularemie ECM1 A r t h r i t e MPN2 ACA3 LBC* Homme, o v i n s , r o n -g e u r s , la-gomorphes Homme, (Chien)
Babésies BoAeA-La op.
Trypanasomes 7/typanorama &p.
Piroplasmose Bovins, ovins, ron-geurs, Chien; excep-tionnellement : Homme Rongeurs, herbivores ?
Filaires Dipeioéonema ingoàicauda Chevreuil
Tableau 3 : Organismes et maladies transmis par I . JU.CÙJULÒ en Suisse.
1. &u/ihema chA.onU.cwn uiigiarvù
2. M é n i n g o - p o l y n é u r i t e
3 . AcAod&uMLÜjLLi chA.oru.ca aijiophlcarta 4 . LymphadenoA-Lb benigna cutlò
Fig. 2 : Cycle Û%1. ILCÀIUIA,
R.S. : repas sanguin
L : larve
N : nymphe
r
Hâte
J
Repss sanguin
\
„Drop off"
Digestion
^ »
Mue
/tepos
^ ».
Jeûne
I I I Ouête —«
*
I I I £uete
»
Pression du milieu:
I I I air libre
10
(YlATERIEL ET METHODES
I LA 7IQUt fi. ùcìnuò)
1.1 Capture
Les tiques sont récoltées grâce à la méthode dite "du drapeau" (Aeschlimann, 1972) et transportées dans des tubes de verre ou de plas-tique (10 cm ) herméplas-tiquement fermés et dont l'atmosphère est saturée en humidité è l'aide d'un simple brin d'herbe.
1.2 Elevage
La technique est reprise de Graf (1976). Nymphes et adultes sont disposés sur les oreilles de lapins (de race "russe"), tandis que les larves se nour-rissent sur la tête de souris blanches.
1.3 Marquage
Afin d'assurer la reconnaissance durant l'expérimentation, il a été nécessaire de mettre au point une technique de marquage; différents pro-cédés ont été testés : le Tableau 4 en fait le bilan. Ces essais ont fi-nalement conduit à adopter le système suivant :
Couleurs acryliques (réf. Pliocolor) en 7 tons différents aisément re-pérables.
Adultes : 2 taches permettant de réaliser un code à 49 numéros. Immatures : 1 tache permettant de réaliser un code à 7 numéros. La peinture est appliquée à l'aide d'une aiguille coudée et emman-chée (Fig. 4 ) . Etant donné son séchage rapide, elle peut être posée sur les tiques dans la nature, sans prélever celles-ci de leur support et par conséquent, en les dérangeant un minimum.
Nous avons procédé à des essais de marquages en laboratoire sur des lots de 100 individus de chaque stade : la mesure des taux de mortalité (heb-domadaire sur une période de 3 mois) n'a pas montré de différence signi-ficative avec l'observation de lots de tiques non marquées placées dans les mêmes conditions (Pl.I.2) et ceci pour tous les stades.
SYSTEME AVANTAGES INCONVENIENTS
Une tache de couleur (peinture cellulosique)
Vite fait, visibilité en fonction de la cou-leur
Pas de reconnaissance individuelle, solvant toxique, sèche trop vite
Idem, peinture acrylique Idem, non toxique, di-luable dans l'eau
Pas de reconnaissance individuelle
Code à 9 points d'encre de Chine sur tache de peinture
Marquage individuel Réalisation délicate dans le terrain, mauvaise te-nue, décodage pénible
Poudre fluorescente + colle (colle vinylique, cyano-acrylate, peinture acrylique ou cellulosique transparente)
Visible de jour comme de nuit, tenue plus ou moins bonne, selon la fraction collante
Peu de couleurs à dispo-sition, surface trop pe-tite pour être visible la nuit (à l'aide d'une source U.V. "lumière noi-re")
Tableau 4 : Les différents types de marquages testés.
Fiq 4 : Tiques adultes marquées à l'aide d'un code à deux couleurs.
R US 7CRRAIlVS D'ÌXPERMIN7A7I0N
2.1 Choix
Le terrain principal devait remplir les conditions suivantes : - Etre un biotope typique d'I, /U-cJjjiu,.
- Abriter une population de tiques nombreuses.
- Se trouver relativement près de l'Institut de Zoologie de Neuchâtel. - Etre peu perturbé par le tourisme, le trafic routier ou
l'exploita-tion forestière.
Le Staatsuiald remplissait toutes ces conditions» plus une cinquième très intéressante : il avait déjà servi de milieu d'étude aux travaux de Plermod et al. (1973, 1974, 1975) sur la biologie d'Z. nXciniu. Les présentes recher-ches pouvaient donc bénéficier des résultats de ces auteurs et, réciproque-ment, compléter le portrait d'une forêt à tiques du Plateau Suisse.
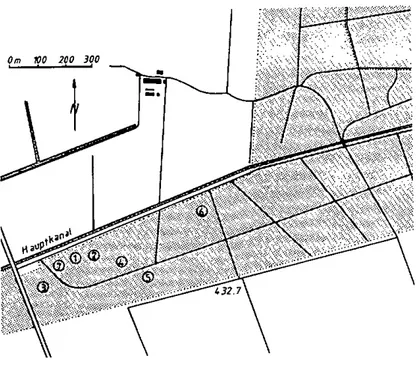
E . 2 Situation
Sise dans le "Grand Marais" (Coord. 203/575-575, près Anet (BE), Fig. 5) sensiblement entre les trois lacs de Neuchâtel, Bienne et Plorat, cette forêt peut être considérée comme un biotope typique d'I, /LICUSIUÒ. (Altitude : 433 m, centre du Plateau Suisse, végétation et climat idéaux qui seront discutés plus loin).
Afin de prendre un étalon représentatif de la situation des tiques en dehors de ce type de forêt, nous avons choisi d'examiner brièvement la situa-tion sur le flanc dE "Chaumont" (Coord. : 563,700/207,10G, en-dessus de Haute-rive (NE) Fig. 5 ) , le premier pli du Jura, dont la pente Sud-Ouest domine le lac de Neuchâtel. Les différences peuvent être regroupées selon trois critè-res :
1° L'altitude (650 m ) de 22D m environ supérieure à celle du Staatsuiald. 2 Le climat : situation de pente, soumis à la réverbération du soleil
sur le miroir du lac.
3 La végétation (chênaie buissonnante), conséquence du climat et du sol mince reposant sur une dalle calcaire.
Ce biotope peut donc être mis en opposition avec celui du Staatsiuald (Fig. 2 1 ) .
H.3 Historique
Toute la région du Grand Marais et de ses environs repose sur le
substrat molassique tertiaire. Son histoire, depuis la dernière
glacia-tion, peut être résumée ainsi (Ludi, 1935) :
Le retrait des glaces a fait place à un lac d'une centaine de
kilo-mètres de long, s'étendant de Soleure, au N-E, au Passage d'Entreroches,
au S-D (Fig. 6) et dont le niveau était probablement supérieur d'une
ving-taine de mètres au niveau actuel. De grandes quantités d'alluvions
fluvia-les se sont accumulées, préparant le sous-sol du futur Marais que
l'abais-sement des eaux aurait déjà pu faire apparaître à l'époque romaine. Dès
lors, l'alternance de périodes d'émersion et d'immersion a permis
l'éta-blissement sporadique de cultures. C'est afin de garantir ces zones contre
les perpétuelles inondations que la première correction des eaux du Jura
a été entreprise (1B68-188D). Par un jeu de canaux de drainage et grâce à
un renforcement des berges, les fluctuations des trois lacs ont été
rame-nées de 4 m à 2,8 m, leur niveau s'abaissant de 1,4 m (lac de Bienne)
et 2,4 m (lacs de Neuchâtel et Ctorat). Ainsi, une grande surface du
Ma-rais put être "assainie". Cependant, les drainages effectués entraînèrent
un affaisement du sol de 0,7 à 1 m et comme, d'autre part, toutes les
crues ne pouvaient pas être absorbées ou contenues, il a fallu compléter
les travaux : une deuxième correction a été nécessaire, celle de 1930.
L'établissement de la forêt actuelle du Staatstuald (alors nommée
"Kanalwald") date de la première correction. Elle croît sur un sol
tour-beux, de 20 à 60 cm d'épaisseur, superposé à une couche de marne : ces
deux éléments s'accordent bien pour une rétention durable de l'eau.
D'au-tre part, l'épais couvert arbustif et arborescent (voir ci-dessous)
s'op-pose efficacement à l'assèchement de l'atmosphère de la forêt durant la
période de végétation. Ces deux groupes de facteurs collaborent à
réali-ser un fort "tamponnage" hygrométrique, tant pédologique qu'atmosphérique,
dans le sous-bois.
H.4 Climat
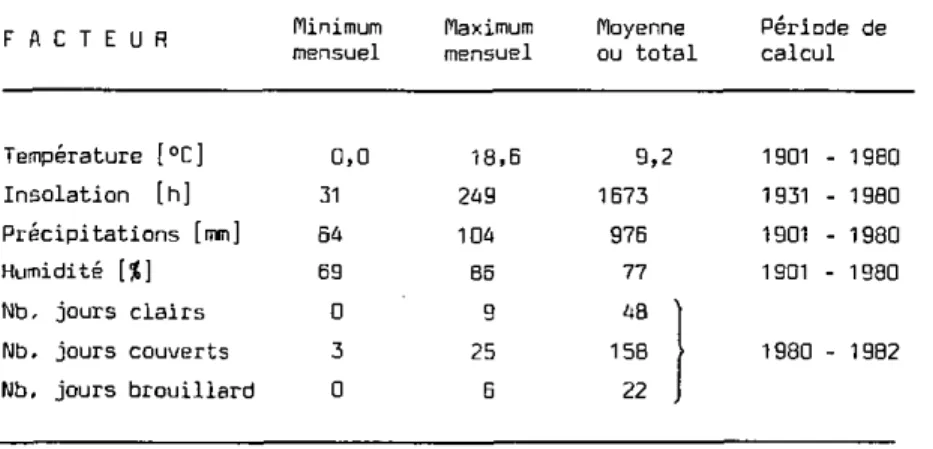
Afin d'avoir une bonne vue générale du climat de cette région, nous
nous sommes référés aux données de l'Observatoire de Neuchâtel, station
météorologique la plus proche de notre terrain (Jornod, 1980 à 1984).
Les mesures figurant au Tableau 5 montrent que nous avons à faire à un
cli-mat tempéré doux, sur lequel l'influence océanique régularisante est
complétée par celle des lacs.
Les particularités du Staatswald seront présentées plus loin (R.I), mais disons déjà que son climat ne s'écarte guère de celui du Plateau Suis-se en général.
H,5 Végétation et phytosociologie
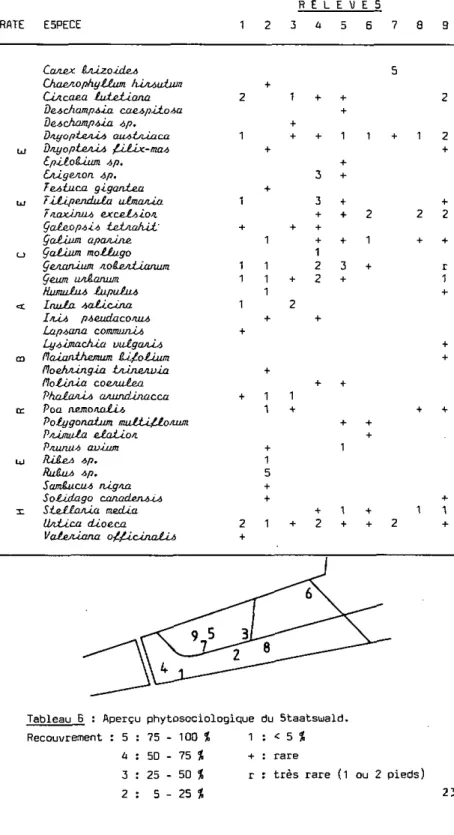
Sien que présentant plusieurs types de végétation, le Staatsuald par-ticipe entièrement du Pnuno-'t'/iwcineiiim (alliance : Aino-Padion). C'est cet-te frênaie à Plerisier qui s'est installée naturellement après la dis-parition du marais. Actuellement, et selon l'influence de l'homme et des forestiers, différentes zones peuvent être individualisées (Oerscheid, 1961) :
a) Feuillus adultes : la végétation se caractérise par une riche et épaisse strate arbustive (relevés 5, 7, 9; les relevés indiqués figurent sur le Tableau B ) .
b) Jeunes feuillus (relevé 8) à sous-bois d'autant moins fourni que la plantation est serrée et récente.
c) Forêt mixte (feuillus et conifères) dont les strates arbustive et arborescente sont bien fournies (relevés 1, 4 ) .
d) Conifères adultes : pessières à sol pratiquement nu ou pinèdes aux épais fourrés de Framboisiers et de Ronciers (relevé 6 ) . e) Plantations de jeunes conifères (Pj.cea OIHEA), serrées sans
sous-bois (pépinières).
f) Clairières plus ou moins humides, de tailles restreintes (Ron-ciers dans les plus sèches, faux Roseaux et Iris, dans les plus humides).
g) Les chemins voient la strate arbustive de la zone gu'ils traver-sent s'enrichir en graminées à leur contact.
Etant donné le caractère essentiellement labile de la phytosociolo-gie du lieu depuis les corrections des eaux et l'influence continuelle de l'homme, nous sommes en présence d'une mosaïque, très fine, de végétation. Cette mosaïque présente néanmoins une grande communauté de caractères :
- Le sol, tourbeux (20-6D c m ) , repose sur une couche de marne im-perméable : les fortes rétentions d'eau qui s'ensuivent évitent le dessèchement lors de périodes arides, même prolongées (été S3, par exemple).
- L'humus, de type mull (pH k-S), montre une décomposition rapide.
- L'épaisseur des strates arborescente et arbustive, pour ne pas
di-re la luxuriance, limite fortement les vents et l'ensoleillement,
réalisant ainsi un fort tamponnage hygrométrique du milieu en
géné-ral, et de ses strates basales en particulier.
- La forte productivité du milieu, en partie due à la présence
d'es-pèces nitrophiles, contribue à maintenir cette situation.
Il faut toutefois relever que les pessières, à plus forte raison les
jeunes, n'entrent pas dans ce cadre : un ombrage trop épais ainsi qu'un sol
trop acide à litière se décomposant mal, empêchent l'établissement de presque
toute végétation de sous-bois.
H.B Faune
Les caractéristiques physiques de la végétation sont en mesure
d'of-frir de nombreux abris à une faune mammalienne et avienne riche, compte
tenu de la relative exiguïté du biotope. Le Tableau 7 présente cette
fau-ne, eu égard a son rôle d'hôtes potentiels d'i".
ticUniiA.
C'est pourquoi
seules les espèces d'oiseaux à forte activité au sol sont citées
(Der-scheid, 1981).
H.7 Situation des diverses expérimentations
Afin de mieux saisir l'utilisation du terrain d'étude, nous faisons
figurer sur la carte de la Fig. 7 les lieux de chaque expérimentation.
Des rappels à cette carte seront faits dans le courant des présentations
des dites expérimentations.
s
ì
tv IO O) O) V) "D in 19Ml
F A C T E U R Minimum mensuel Maximum mensuel Mayenne ou total Période de calcul Température [0C] Insolation [h] Précipitations [mm] Humidité [%] Nb. jours clairs Nb. jours couverts Nb. jours brouillard 0 , 0 31 64 69 0 3 D 18,6 249 104 BD 9 25 6 9 , 2 1673 976 77 48 158 22 1901 - 1980 1931 - 1980 1901 - 1980 1901 - 1980 1980 - 1982
Tableau 5 : Météorologie régionale annuelle, selon la station de 1'Observatoire de Neuchâtel.
STRATE
LJ^-Z Ld
(_)
tn LiJrx
O^
CC ŒE5PECE
Recouvrement t o t a l (¾]
Acesi pòGJjdoplaijaruLò
Atnxiò giuLùiòòa
AÙIUÓ AnconaBctuta penduta
Taguò Aituaiica
T/iaxMuu exc&tòioi
Picea a&i&A
PinuA AOIO&UAPopuùiA aMa
PoputuA nig/ia
ùuE/lCUA A.O&UASamfUicuA nig/ia
llùnuA Aca&Aa
1
8D
+
+
il+
2
3
4
+
1
2
R
3
80
2
2
2
2
3
r
r
E L
4
60
+1
1
+4
+
1
E U
5
75
+
3
2
+
3
+
-L_5
6
80
5
1
1
7
75
3
3
2
2
B
85
5
+
+3
9
85
2
2
2
2
1
Recouvrement t o t a l [%] 30 65 40 80 1D 10 1D 75kce/L pAcurtoplaianuA
AimiA ghïtinooa
AlmiA incarta
Co/iyiuA aocitùwa
OiatacguA monogyna
CvonymuA eiiAopaeuA
T/ianguta alnuò
T/wxJjuiA exc&tò-ion.
flumitùi/, tupuiuA
Liguói-num vulgate.
PU&UA ca&4piÀoAu.APicca adLcA
PmimiA avium
P/iunuA padiu,
PxunuA òpinoòa
QuCA-CUA lOÙlAPiAcA -6/3.
PuIUiA idaeuA
Pu(UiA cacA-ùiA
SamlUicuA nigna
UùmiA Aca&sia
ViAunnum opuluA
+r
+
+
1
r
+
+r
+
2
2
2
r
2
+ +• r3 4
+
1
r
r
+ +1
2
+ 2
1
+• ++
+
+
2
2
+
++
+
+ +2
+1
+2
4
1
Recouvrement t o t a l [%] 50 10 80 35 5 95 5 30Aceji pAeudaplaixinuA +
Acgopodium podagiania +
Angctica AylucAtnAA 2
AApcAuta odo/iota 1
Athy/Uum /llix.-£emina +
Avena Ap, + 1B/iachypodMim AÜuaticum 2 + 2
Btyonia di.oe.ca
1 2
COA&X acutifanmi/, + + 1 1 2 2R E L E V E S
STRATE Lu LiJ LJ H CD Qf LU X ESPECEOwesiophyttum hÀsióutum
Cùicaea £uteiU.ana
deAchampAia cae.Apu.oia
DeóchompA-ia òp.
D/Ufopt&iJsi auAtnÀn.ca
D/Lyopte/uA £LLbc-mai
tZpitoêJMm òp.
digestori òp.
TeAtu.cn. gigantea
Filipendula uJbncuUa
7/iaxJjuu exceJjtion.
ÇaleopiiA tejbiafUt'
Galium apanÀne.
Çalium mollugo
ÇeAanium /lofLejiLianum
Çeum unAanum
HumuluA lupuluA
Inula Aalicina
IILLA pA&udaconuAbipiana communti
LyòimacJu.a vulgasilA
ftaianthemum tLi^olium
Pioehtingin t/üjie/ujjxi
flo-tùtia coejiulen.
PhalatiA ammdinacca
Poa nemonaliò
Polygonahm multi£lo/utm
P/ujnula eÂation.
PAAUUIA aviumIUÄ2J> òp.
RuÂuA Ap.
Sam&ucuA nû.g/ia
Solidago ca/iade/Ló-iA
SteMasUa meaUa
(Ittica aU.oe.ca
ValejUana o£{j,cÀnaJLu>
1 2 1 1 + 1 11
+
+2
+
2 ++
+
1 11
1
++
1
1
+
1
5
+
+
1 31
+
+ + + 2 1 + + h + + 3 3+
+
+ 1 2 2 + + + 2 5+
+
1+
+
+ + + 3 + ++
1
1 + 6 1 2 1 +•+
+
+ +7 a
5 + 1 2 + + 1 2 9 2 2 + + 2 + r 1 + + + + + 1 +Tableau B : Aperçu phytosociologique du Staatswald.
Recouvrement : 5 : 75 - 100
%
h
: 50 - 75
%
3 : 25 - 50
%
2 : 5 - 25
%
1 : < 5
%
+ : rare
r : très rare (l ou 2 pieds)
23
1
5
I
• " J&
•5
a
•3
5^
1O O O•3
3
•ä»
O Ol O£
«s!a
e
5
I
• 9 3 W3
3 C24
Fig. 7 : Plan du Staatswald et implantation des dispositifs expérimentaux dans les différentes stations.
Station 1 : TqD1 I1 2, 3, 11 et 12; influence de l'état hydrique; cabane météorologique (enregistrements locaux permanents).
Station 2 : TqO 13 et U ; femelles "d'Ariane". Station 3 : TqO 15 et 16.
Station 4 : TqD 17 et 18.
Station 5 : TqD 4, 5, G, 19, et 20; influence de l'état hydrique; femelles "d'Ariane"; dérangement des phases de quête; piégeage de Carabes.
Station 6 : TqD 21 et 22.
Station 7 : "sacs" pour les phases endogées.
1 : "tiquodrome"
m
L*ACTIVITE D€ QU£7£
HI. 1 Observation directe
Quoique langue et laborieuse, l'observation directe a été pratiquée .
pour repérer les tiques directement sur leur support de quête, et,
par-tant, s'assurer que les résultats expérimentaux correspondent à la
réali-té. Les principales difficultés sont dues aux conditions lumineuses et à la
nature du substrat sous-forestier. En moyenne, une douzaine de tiques
peu-vent être repérées en une heure.
Après repérage, l'obseruation comportementale doit être menée à
l'aide de jumelles ou d'une longue-vue, à courtes focales, afin de
s'éloi-gner au maximum du sujet. C'est la seule manière de prolonger lé temps de
travail auquel la détection de l'observateur par la tique met
irrémédia-blement fin : ce laps de temps est en effet fonction directe de la
distan-ce observateurf= hôte potentiel)/tique.
IH.2 Drapeau
La méthode dite "du drapeau" a été utilisée chaque fois qu'il était
nécessaire de récolter des tiques pour l'expérimentation ou la mise en
é-levage. Cette méthode, imaginée par Philip (1937), a été modifiée par de
nombreux auteurs. Nous utilisons ici le système décrit par Aeschlimann (1972).
Le drapeau doit être relevé tous les 10 à 15 m pour éviter la perte
d'indi-vidus se laissant tomber peu après leur capture. Ces pertes concernent
sur-tout les adultes et plus particulièrement les femelles, c'est-à-dire les
stades gros et lourds, non retenus par la structure bouclée du tissu. Un
morcellement du piégeage en plus petits tronçons de capture n'apporte pas
d'améliorations. Le rendement est illustré au Tableau 8.
m . 3 Battage
Il s'agit de la méthode classique, dite du "parapluie japonais",
décri-te par PlatthBy et al. 19B4. Dans notre cas, le niveau maximal exploré dans la
couronne d'un arbre est de 4 m.
Ht.û Systèmes de capture sélectifs en hauteur
Afin de mesurer la hauteur, de quête des différents stades
d'I. sU.cJsuu>,
deux voies peuvent être suivies : l'observation en place et la capture. Trois
dispositifs différents ont été imaginés et testés dans cette dernière optique :
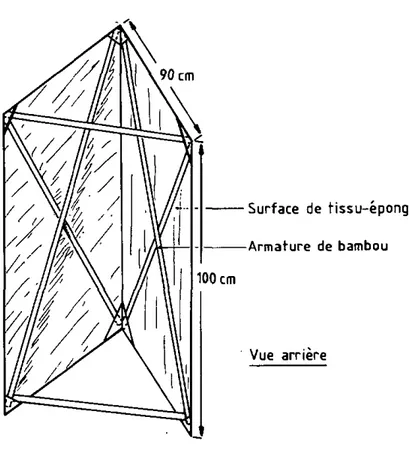
a) Le "triangle". C'est une transcription verticale du drapeau : une surface d'interception en tissu-éponge est tendue sur une armature de bambou en forme d'étrave, qui sera poussée à trauers la strate her-bacée. L'angle d'ouverture de cette étrave peut être modifié en fonction de la densité de la végétation, donc de la difficulté de pénétration. Son angle frontal est renforcé par un empiècement de fort coton, fWec un angle d'ouverture de 6D , la surface frontale d'interception est de 1 m (Fig. B ) .
Trois critères doivent être satisfaits :
1 On doit pouvoir pousser le dispositif à travers la uégéta-tion et au ras du sol, auec une relatiue facilité.
2 La hauteur de quête des tiques doit être comparable à celle à laquelle elles se fixent sur le tissu-éponge.
3 Le rendement doit être satisfaisant, comparé à celui du dra-peau traditionnel.
Des essais pratiques ont été menés dans différents terrains et mon-trent les lacunes suivantes :
1 La résistance de la uégétation peut être très forte (brous-sailles à HuAnA Ap, par exemple). Il faut donc diminuer l'an-gle d'ouverture du trianl'an-gle et ramener ainsi la surface d'in-terception à 0,5 m .
2 Les herbes sont écartées du dispositif par leur base : il y a donc trop peu de chance pour que leurs sommets, au voisina-ge desquels les tiques se trouvent, entrent en contact avec le tissu.
3 Dans des conditions comparables, le rendement du triangle at-teint la moitié de celui du drapeau, ce qui peut être consi-déré comme suffisant pour échantillonner valablement le bio-tope concerné.
b) A partir de ces préliminaires, un deuxième projet a pu être dessi-né, puis réalisé. Il réunit les modifications suivantes :
- Angle frontal plus faible : meilleure pénétration.
- Flancs inclinés en surplomb : les sommets des plantes touchent le tissu avant les tiges; ils ne sont donc plus écartés, as-surant du même coup un meilleur contact végétation/surface de capture. D'autre part, une tique en train de se laisser tomber du tissu ne pourra s'y raccrocher plus bas : la mesure n'est ainsi pas faussée.
- Pour seri/ir de manière répétitive (trois jours par semaine)
durant une saison, ce dispositif devait être maniable et
résis-tant : il est donc construit en bois croisé (le tissu-éponge
tendu à sa surface) et monté sur une roue pneumatique {Fig. 9
et 10).
Ainsi modifié, le "triangle", devenu "brouette", est au point et
accu-se un rendement accu-sensiblement égal à celui du drapeau, compte tenu d'une
lar-geur d'interception de moitié inférieure à celle de ce dernier (Tableau 8 ) .
Son seul inconvenient réside dans le fait que la tranche de végétation
com-prise entre D et 1D cm n'est pas explorée, le bas de la brouette ne pouvant
toucher le sol. Il ne sera donc pas un moyen adéquat pour capturer les
lar-ves, ces dernières se trouvant en majorité près du sol.
c) Afin d'améliorer le rendement et de simuler l'hôte d'une manière
plus complète, nous avons imaginé un autre dispositif.
Une combinaison (type "over-all" de mécanicien) est réalisée en
tissu-éponge. Toutes les coutures latérales sont remplacées par des
fermetures-éclair. Les ouvertures (encolure, poignets, chevilles) sont serrées autour
du corps par des élastiques (Fig. 11). L'expérimentateur qui la revêt
four-nit donc sa chaleur et certains facteurs olfactifs à cette "fourrure
arti-ficielle". Après avoir cheminé dans le biotope en question, il retire la
combinaison et l'étalé complètement, grâce à l'ouverture de toutes les
ferme-tures(!)-éclair. Il peut alors relever la hauteur de fixation de chaque
ti-que dans le tissu (Fig. 12).
Or, les résultats escomptés n'ont de loin pas été atteints (Tableau 9 ) .
Ce qui laisse supposer que :
1 La largeur du front d'interception (tissu/végétation) est trop
faible.
2
Cet inconvénient n'est pas compensé par l'attractivité du
sys-tème.
3 Cette absence, ou insuffisance de compensation, prouve que les
facteurs olfactifs sont marginaux dans la rencontre de l'hôte.
4 Au contraire, les stimuli mécaniques (en particulier de
con-tact) occupent une place primordiale dans cette rencontre.
5 Cette stratégie est tout à fait compatible avec une "chassE
à l'affût" telle que la pratique
I. lÀcUnu-ò,
par opposition
à la "chasse à courre" de certains
Amê-tyomma
et
Hyalomma.
6 L'efficacité étonnante d'un système classique de capture.
simulant si grossièrement un hôte, le drapeau, devient alors
très compréhensible.
Finalementi la brouette décrite sous b) sera retenue comme moyen de
mesure des hauteurs de quête d'-f.
/U.<Uruu>
dans la végétation.
HI.5 Lâchers des tiques
Dans les expériences nécessitant le lâcher de tiques, il a été procédé
comme suit :
1 Capture des tiques, au drapeau, dans le milieu le plus
sembla-ble et le plus proche possisembla-ble du futur lieu d'expérimentation.
2° Stockage en tubes de verre (40 x 19 m m ) ; l'humidité de
l'at-mosphère est assurée en enfermant les tiques avec un brin
d'her-be.
3 Transport au laboratoire pour le marquage des individus, s'il
y a lieu.
û Lâcher dans le terrain : les tiques sont manipulées avec
pré-caution à l'aide de pincettes fines (No 4 ) ; on évitera de les
saisir par les pattes antérieures, celles-ci portant de
nom-breux organes sensoriels susceptibles d'être endommagés
(or-gane de Haller en particulier). Cette manipulation doit être
aussi rapide que possible, afin que le comportement des tiques
soit influencé un minimum par la présence de l'opérateur.
m . B Sélection du support naturel
Dans l'optique de définir le support de quête typique, nous avons
procédé de trois manières différentes :
- Déposer les tiques sur, ou au pied de supports choisis et
observer leur comportement.
- Déposer les tiques dans un périmètre dont la végétation a été
tondue et remplacée par les supports à tester. Observer la
sé-lection et le comportement.
- Observation directe dans la végétation. Relever l'espèce et
les caractéristiques du support.
HI.7 Supports artificiels
L'observation répétée des tiques dans leur phase de quête est trop laborieuse sur les supports naturels qu'offre la végétation. C'est pour-quai nous avons décidé de réaliser des supports artificiels standards dont voici les caractéristiques :
- Baguettes rondes de hêtre, 0 5 mm, longueur émergée 8D cm. - Fil de cuivre rond, 0 2 mm, langueur 10 ou 20 cm, peint à
la couleur acrylique jaune ou blanche.
- Tiges rondes de PUC dur, 0 3 mm, longueur 10, 20, 30 ou 50 cm. Tous ces supports sont enfoncés verticalement dans le sol (de 10 ou 20 cm, selon leur longueur); ils sont regroupés sur de petites surfaces, précédemment tondues, de manière à éviter la concurrence supports naturels/ supports artificiels. (Des expériences (R.IX.1) ont montré que les déplace-ments horizontaux sont suffisamment limités pour qu'une telle surface n'en-traîne pas ou peu de pertes en tiques).
Si les tiges de bois peuvent parfois se déformer lors d'intempéries, les deux autres matières ont une tenue parfaite.
Les tiques se détachent bien sur la couleur claire de ces matériaux, qui reste constante malgré son vieillissement. Aucun entretien n'a été nécessaire durant les mesures qui ont duré deux ans.
HI.8 Enceintes réelles à hauteur de supports imposée
Dans le premier temps, il nous a semblé utile de réunir ces supports artificiels à l'intérieur de petites palissades (Fig. 1 3 ) . On obtient alors un système relativement fermé :
- Les tiques observées sont celles quis'y trouvaient naturelle-ment et qui ont subi leur mue à l'intérieur du périmètre. Il n'y a pas d'immigrations ni d'émigrations actives. Dès leur émergence, elles doivent se satisfaire des supports fournis ou rester sur le sol : on ne peut observer que les tiques sur supports.
- Le passage des hôtes mammaliens est limité par la barrière; on réduit ainsi fortement l'émigration passive des tiques. En réalité, la palissade est parfois non pas franchie, mais escaladée par les tiques, l'arête leur servant de support. Pour éliminer ce biaisa-ge de nos mesures, nous avons tenté d'enduire le bord supérieur de la bar-rière de différentes substances anti-fuites : talc, vaseline, lanoline et
finalement glu ont tour à tour été testés, mais les tiques franchissent plus ou moins bien tous ces obstacles. Il a donc fallu se résoudre à ad-mettre ces fuites. Une invasion de l'extérieur n'est pas à redouter : le milieu non modifié fournit tous les genres de supports naturels nécessaires aux tigues."
Ce type d'installation a été utilisé pour tester la nature du support et en sélectionner un succédané (Pl.HI.7). Il a ensuite servi à déterminer la longueur de ces supports artificiels pour chaque stade d'I. /ticimiA. A cet effet, nous ayons disposé deux batteries de trois enceintes dans deux milieux différents. Les supports proposés sont respectivement de 10 cm, 20 cm
(cuivre laqué blanc ou jaune) et BO cm (bois). Chacun de ces dispositifs, baptisé "tiquodrome" est repéré sur la Fig 7.
Les relevés ont lieu trois fois par semaine et comprennent pour cha-que individu les données mentionnées sur le Tableau 10 (TqD 1-6).
Ht.9 Enceintes virtuelles à choix de hauteurs
Après une année d'utilisation des tiquodromes, certaines modifications se sont imposées :
- Simplification et rapidité d'observation grâce à une géomé-trie en rectangle étroit.
- Possibilité pour les tiques de choisir le support de hauteur convenable.
- Suppression de la palissade : le choix de support à disposi-tion recouvre les besoins des tiques et leurs migradisposi-tions ho-rizontales actives se sont révélées très faibles.
On aboutit alors au dispositif épuré de la Fig. 1ü : les supports, numé-rotés de 1 à 49, sont tous réalisés en PUC dur, blanc et rond, 0 3 mm. Douze de ces dispositifs sont répartis par paires dans six stations caractéristiques du Staatsiuald (Fig. 7, 15-20). Une paire, située sur le flanc de Chaumont (Fig. 2 1 ) , donne un aperçu du comportement dans une station d'altitude (ait. 650 m ) .
Les tiques sont marquées, en place, à la peinture acrylique selon Pl.I.3, lors de leur première apparition.
Les relevés ont lieu trois fois par semaine et comprennent pour chaque individu les données mentionnées sur le Tableau 10 (TqD 11 - ¾ ) .
Ramenées du terrain, elles sont ensuite réduites à une ligne, par indivi-du et par relevé et ainsi introindivi-duites dans l'ordinateur grâce à un symbolisme numérique.
m . 1 0 Interruption provoquée de l'activité de quête
Certaines observations directes ont montré que des tiques en quête, mises en présence d'un hôte à très faible distance, le détectent et entrent dans un état d'excitation tel qu'elles peuvent tomber au sol. Plais il serrv ble qu'elles regagnent volontiers rapidement leur poste, puisqu'on peut ob-server les supports réoccupés peu de temps après (2-3 jours).
Il nous a donc paru intéressant de concevoir une expérience de terrain pour examiner si tel est bien le cas. Il s'agit en fait d'infirmer une des deux hypothèses suivantes :
1 La tique doit être soumise à des conditions externes bien par-ticulières pour que son comportement de recherche de l'hôte {mise en poste de quête) se déclenche.
2 La tique se trouve dans un état physiologique privilégié et durable qui programme de manière permanente son comportement de recherche de l'hôte.
Dans le premier cas, les tiques descendues de leur support y remonte-ront après des délais variables d'une fois à l'autre.
Dans le second, elles remonteront toujours dans le même délai, le plus court possible et inlassablement tant que dure cet état physiologique.
D'un point de vue collectif, la première hypothèse entraîne des pha-ses de quête synchronisées entre individus; la seconde doit montrer des séries d'apparitions non superposables entre individus.
Remarque : il est très vraisemblable que, même dans le cas de la pre-mière hypothèse, la tique doive se trouver dans un état physiologique par-ticulier pour répondre aux stimuli extérieurs. C'est pourquoi il est pri-mordial de travailler avec des tiques en quête au moment de leur capture et de les introduire immédiatement dans l'expérimentation. Cette possibi-lité est offerte par la capture au drapeau.
Protocole : 5D 9 et 50 <t capturés sont immédiatement marqués (marquage individuel : Pl.I.3) puis lâchés au centre d'un dispositif comprenant 2? supports de 50 cm de haut (PVC dur, 0 3 mm ) : Fig. 22 (voir carte Fig. 7 ) .
On procède de même pour les nymphes, mais le lâcher a lieu dans un deu-xième dispositif dont les supports mesurent 20 cm de haut (Fig. 2 2 ) . Ces hauteurs correspondent aux préférences des tiques, mises en valeur dans les expériences décrites sous Pl.IH,9 et H.HI.9. La petite taille des nymphes contraint à réduire le code de marquage à 29 possibilités : 28 N sont donc lâchées.
ft chaque relevé (3 fois par semaine), toutes les tiques en quête sont prélevées sur les supports et relâchées au centre du dispositif, après qu'on ait noté le numéro de l'individu, et sa distance par rapport au centre du dispositif.
III.11 Influence de l'état hydrique
D'après Lees (1948), I . /UcÀnuò présente une réaction de fuite si elle est plongée dans un environnement à l'humidité relative comparable à celle ayant régné durant une période de préconditionnement : en effet, une tique desséchée pendant 16 h. dans une ambiance à 50 % d'humidité relative restera dans une ambiance à 95 % H.r., mais fuira une ambiance à 34 % H.r. La situa-tion est symétrique pour une tique précondisitua-tionnée à 100 % H.r. (évitement de la saturation).
Dans la nature, cette particularité comportementale pourrait être à l'origine des alternances quête-repos observées au cours de la période d'ac-tivité. Pour tester cette hypothèse dans le terrain, nous avons lâché dans des dispositifs à supports de PVC dur (0 3 mm, longueur 10 et 50 cm; Fig. 2 2 ) , des lots de tiques d'élevage conditionnées et marquées comme suit :
- 1 lot soumis à une perte hydrique par un séjour de 15 h. dans une atmosphère à 50 % H.r. Marquage rouge.
- 1 lot préservé de perte hydrique par conservation en atmosphè-re saturée, Marquage jaune.
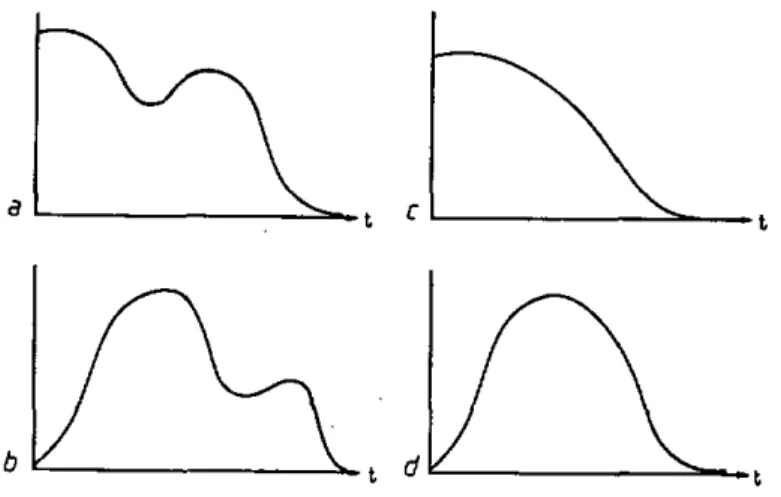
Ces lots comprennent les trois stades. Eu égard à leur fragilité, nous n'avons pas procédé au marquage des larves. Une éventuelle influence du pré-conditionnement sera néanmoins détectable puisqu'une partie de la population différera sa mise en poste de quête; deux pics se marqueront sur la courbe d'activité totale (Fig. 23 a et b ) . Dans le cas contraire (indépendance vis-à-vis du préconditionnement), les deux lots montreront deux périodes d'activi-té confondues, d'où une courbe monomodale (Fig. 23 c et d ) .
La situation des trois dispositifs de ce type employés au Staatswald est mentionnée sur la carte de la Fig. 7.
HI.12 Modèles de laboratoire
En parallèle aux dispositifs de terrain décrits ci-dessus (Cl. UI.8-11}, il faut disposer d'un système de laboratoire pour observer la phase d'ac-tivité en atmosphère contrôlée. Dans un premier essai, les tiques (marquées individuellement) se trouvent dans une atmosphère saturée en humidité, à 20 C (Fig. 2 4 ) . Dans de telles conditions, elles occupent volontiers leur
poste de quête, mais ne l'abandonnent plus guère par la suite. Il est donc souhaitable de pouvoir changer les conditions en cours d'expérience et, en particulier, de renverser les gradients de température et d'humidité rela-tive précédemment établis; ainsi, on peut espérer reproduire les stimuli naturels capables d'induire le mouvement vertical des tiques. Deux types de dispositifs ont finalement été mis au point :
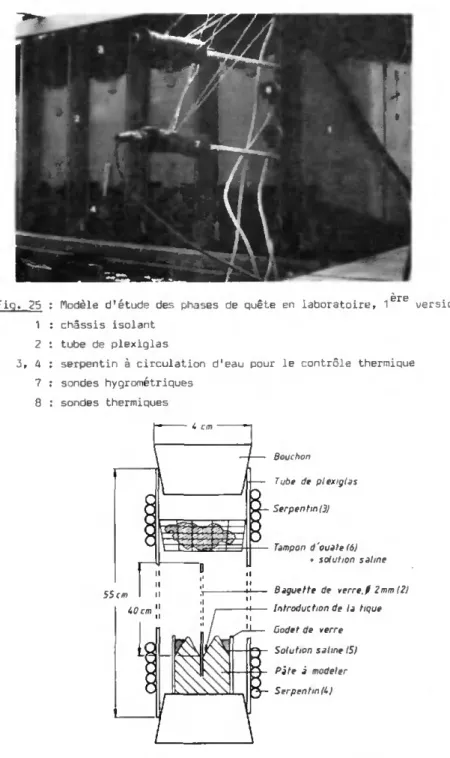
- Le premier (Fig. 25, 26) place les tiques isolément dans des tubes fermés contenant un support de verre ( 2 ) . Le gradient de température est établi par deux serpentins à circulation d'eau chaude et froide (bains thermostatés) en contact avec chaque extrémité du tube (3, 4 ) . Le gradient d'humidité s'obtient à l'aide de solutions salines saturées (Handbook Ch. Ph., 1974) disposées dans un godet basai entourant la ti-ge de verre (5) et dans un coton suspendu au sommet du tube (6). Neuf de ces tubes sont réunis en batterie, dont un, ne contenant pas de ti-ques, est muni de trous permettant l'introduction de sondes hygrométri-ques et thermihygrométri-ques (7, 8) : l'établissement des gradients est ainsi contrôlé et leur allure typique apparaît sur la Fig. 27. Si l'humidité relative montre un gradient quasi linéaire sur toute la longueur du tu-be, la température en revanche se laisse moins bien manipuler : il faut compléter les pôles froids et chauds du tube par un réglage approprié de la température ambiante. Les tiques employées sont des adultes âgés de 2,5 mois, stockés à 200C et 95 % H.r.
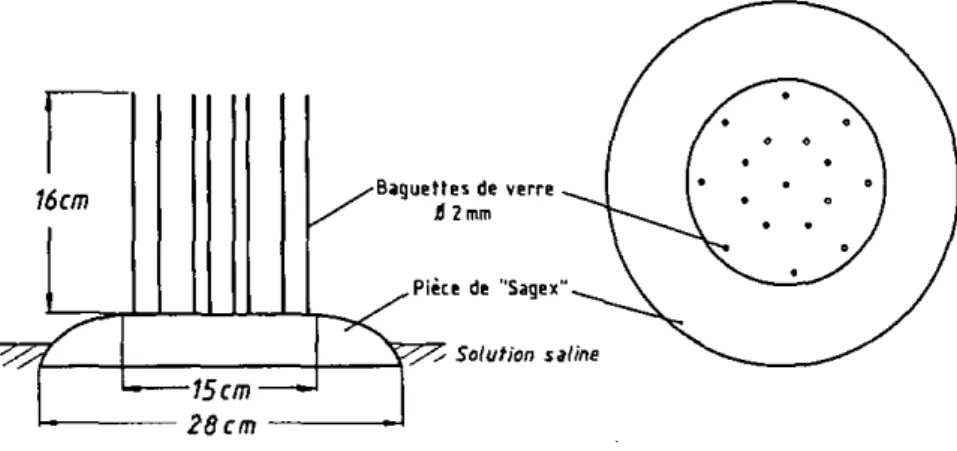
- Le second dispositif, plus simple, ne peut établir de gradient, mais travaille en atmosphère contrôlée, variable à loisir. Son principal in-térêt est l'observation simultanée d'un grand nombre de tiques (Fig. 2 8 ) . Tout le système expérimental est placé dans une chambre d'environnement réglant la température, l'humidité étant ajustée au moyen de la solution contenue dans les petits bacs. Cette solution sert en outre d'anti-fuites, les tiques étant disposées sur des surfaces de "sagex" portant les tiges de verre. Il est important d'arrondir les bords des pièces de "sagex", les tiques ayant tendance à en suivre l'arête (Fig. 2 9 ) . Un grand bac recou-vert d'une plaque de verre contient le tout et suffit à éliminer les in-fluences de l'observateur.
Les tiques (adultes de 4 mois) sont disposées sur la partie périphéri-que des dispériphéri-ques (Fig. 2 9 ) , après avoir subi des conditionnements de trois jours à 95, 66 ou 30 % d'humidité relative et à 20,4 ou 37 C. Chaque lot, de conditionnement homogène, est ensuite exposé aux conditions variables sym-bolisées sur la Fig. 30 : une première période de 49 h., dite de stabilisa-tion (S1), présente un climat "normal" qui devrait permettre l'activité