O
pen
A
rchive
T
OULOUSE
A
rchive
O
uverte (
OATAO
)
OATAO is an open access repository that collects the work of Toulouse researchers and
makes it freely available over the web where possible.
This is an author-deposited version published in :
http://oatao.univ-toulouse.fr/
Eprints ID : 11172
To link to this article : doi:10.1016/j.ecoleng.2012.12.105
URL : http://dx.doi.org/10.1016/j.ecoleng.2012.12.105
To cite this version : Cavaillé, Paul and Dommanget, Fanny and
Daumergue, Nathan and Loucougaray, Gregory and Spiegelberger,
Thomas and Tabacchi, Eric and Evette, André Biodiversity assessment
following a naturality gradient of riverbank protection structures in
French prealps rivers. (2013) Ecological Engineering, vol. 53 . pp.
23-30. ISSN 0925-8574
Any correspondance concerning this service should be sent to the repository
administrator: [email protected]
Biodiversity
assessment
following
a
naturality
gradient
of
riverbank
protection
structures
in
French
prealps
rivers
Paul
Cavaillé
a,∗,
Fanny
Dommanget
a,
Nathan
Daumergue
a,
Gregory
Loucougaray
a,
Thomas
Spiegelberger
a,
Eric
Tabacchi
b,
André
Evette
aaIrstea,UREMGR,2ruedelapapeterieBP76,38402Saint-Martin-d’Hères,France
bCNRS,UniversityofToulouse,InstitutNationalPolytechnique,EcoLab,Bât.4R1,118routedeNarbonne,31062Toulousecedex9,France
Keywords: Beetlediversity Bioengineering Plantdiversity Riverbank Ripariancorridors
a
b
s
t
r
a
c
t
Erosioncontrolofriverbankisfrequentlynecessarytoprotecthumaninvestmentssituatedalongrivers.
Thetechniquechosenforsucherosioncontrolconstructionmayhavemajorimpactsonbiodiversityand
onthefunctioningofrivercorridors.Evenifthereisagreement,thatbiodiversityshouldbeonecriterion
forchoosingembankmentstechniqueslittleisknownaboutwhethersuchtechniquescanaccommodate
biodiversity.
Weaimedtodeterminecoleopteranandplanttaxonomicdiversitiesalonganaturalitygradientof
riverbankprotectionsystems,rankingfromentirelycivilengineeringstructures,throughcombined
con-structions(mixingcivilengineeringandbioengineering),topurelybioengineeringstructures.
Fifteensites(fivesitesofeachtechnique)weresampledintheRhône-Alpesregion(S.E.France).On
eachsite,vegetationwassampledalongthreetransectsfromthebottomtothetopoftheriverbankand
flyingbeetlesbytrapping.
Intotal,werecorded148plantspeciesand78beetlegenera.Wefoundsignificantlyloweranimaland
plantdiversitieswithincivilengineeringconstructionsthanintheothertwotechniques.Diversitiesof
bothtechniquestendedtobehigher,althoughnotsignificantly,incombinedtechniquesthaninpurely
bioengineeringones.Furthermore,civilengineeringstructuresweremoresubjecttoinvasionbyexotic
plantspeciesthanthetwoothertechniques.Theseresultsquantifyandhighlighttheinterestof
bioengi-neeringtechniquescomparedtocivilengineeringinenhancingbiodiversityandlimitinginvasivespecies
techniques.
1. Introduction
In addition to its high level of biodiversity, riparian corri-dorssupplymultipleecosystemservicessuchasnutrientcontrol, floodcontrol,shadingeffectandsocialaspects.Ripariancorridors havethereforebeenwidelyrecognizedaskeycomponentin land-scapes(Décamps,2011).Thereisastrongcausallinkagebetween biodiversityandecosystemfunctioning(Maestreetal.,2012). Bio-diversity loss may induce a modification of natural ecosystem functioningandecosystemservices(McNeely,2010).
However,WesternEuropeanripariancorridorsandtheir veg-etation have been widely affected by centuries of human use. Rivercorrectionsandassociatedfloodcontrolhavebeen primar-ilyperceivedhasagreat humanachievementin environmental
∗Correspondingauthor.
E-mailaddress:[email protected](P.Cavaillé).
control.However,shortlyafterinitialsuccesssuchartificial land-scapes spotlight a major conflict between riparian ecosystem conservationandhumanpatrimonyprotection(Pahl-Wostl,2006), assuchriverstraighteninghasleadtoadrasticlossofbiodiversity in riparian systems. In urbanized areas as well as in agricul-tural landscapes,spaceavailableforriparianvegetationisoften reducedtotheriverbankonly.Thus,organismscanhardly circu-latealongthereremnantsofthepastripariancorridorsespecially whencivilengineeringerosioncontrolstructuresareused(Nilsson etal.,2005).Suchinterruptionsinthecorridormosaicmaylower biologicalcontinuitybyreducingcirculation,refugeandfeeding opportunities,andbyincreasingthermalcontrastsduringsummer. Alteredlandscapesanddisruptedcorridorresultinaglobal ripar-ianecosystemloss(Poffetal.,1997;TocknerandStanford,2002). Consequently,biodiversityconservationshouldbeastrategyfor practitionerstoenhanceecosystemfunctioning(Isbelletal.,2011). In addition, direct exposure to flood and high flood fre-quencyleadtohighpropagulefluxesonbaresoilsandpromote
colonizationbyexoticspeciesandsubsequentlyinvasion(Décamps et al., 1995). Therefore, exotic species richness is significantly greaterinriparianzonescomparedtouplandsites(Stohlgrenetal., 1998).
Habitat restoration hasbeen clearly identified as a key ele-menttoreversenegativeanthropic impactsonbiodiversityand ecosystemsfunctioning(Helfieldetal.,2012).Publicpolicysuch astheEuropeanWaterFrameworkDirective(WFD;2000/60/EC) encouragesaglobal“goodecologicalstatus”ofaquatic environ-mentincludingmorphologicalconditionssuchasstructureofthe riparianzone.Astudyorderedin2005bytheFrenchMinistryof EcologyandSustainableDevelopmenthighlightsthat51%ofthe surfacewatermasswouldnotreachtheobjectiveof“good ecologi-calstatus”by2015(IFEN,2006).Improvingbiodiversityonartificial riverbanksmayactinfavourofthisobjectiveandinparticularif alternativestocivilengineeringareusedsuchasbioengineering techniques(LiandEddleman,2002).
Bioengineeringtechniquesforerosioncontrolconsistsinthe uses of living plantsin order tomimic natural vegetationable to resist intense scouring (Gray and Sotir, 1996; Norris et al., 2008;EvetteandFrossard,2009).Bioengineeringappliedto river-banksaimsatcombiningbothartificialandnaturalcomponents toprovideprotectionattheentirebanklevel(Evetteetal.,2012). Inadditiontotheirerosioncontrolcapacity,bioengineering struc-turesaresupposedpromotingtherecoveryofindigenousspecies andtoensurebetterplantcoveragethanartificiallyreconstructed embankmentsusingcivilengineeringtechniques(Pezeshkietal., 2007).One oftheargumentsgiven forchoosingbioengineering structuresinsteadofcivilengineeringonesisthatthefirst pro-mote the re-establishment of a functional and self-sustainable ecosystem;however,toourknowledge,noquantitativescientific evidenceshave been produced to supportthis idea. Untilnow socio-economicalissuesarethemainreasonstowhetherprotect ornot.Riverandlandmanagers’finaldecisiononwhichriverbank protectiontechniquewillbeusedismainlybasedonseveral cru-cialpointssuchashydro-engineeringaspects,bedmaterialand watercoursebankstability(SchiechtlandStem,1996).However,no quantitativeecologicalaspectisforthemomenttakenintoaccount inthedecisionline.
Todate,many scientificstudiesonbioengineeringstructures alongriversfocusonmethodologicalaspectssuchasstructures, construction methods, mechanical resistanceand thechoice of species(Bretonetal.,underreview),comparedtothefewworks whicharededicatedtobiodiversitysubsequentlyrelatedto ero-sioncontrolstructuresonwatercoursebanks.Since2002andthe earlyworkontheenvironmentalandecologicaladvantagesof bio-engineeringstructures(LiandEddleman,2002),fewstudieshave usedquantitative methodstoinvestigaterelation between bio-diversityanderosioncontrolstructures.Someworksaredealing withplantspeciesand habitatdiversity assessmentbut remain studycasesonveryfewsites(Lietal.,2006;SudduthandMeyer, 2006).Ontheotherhand,studieshavebeendoneonriverbank restorationbyremovingbankfixationorprotectionand compar-ativebiological surveyonrestoredandnon-restored siteswere made(Helfieldetal.,2007;Januschkeetal.,2011).Thoughthereis abigknowledgegapconcerningquantitativebiological compari-sonofriverbanktechniquesforerosioncontrolandtheirimpact onecosystemproperties(Pahl-Wostl,2006).Toourknowledge, nostudyhasyetdemonstratedtherelationshipbetweenmaterials andtechniquesusedforerosioncontrolandassociatedchangesin biodiversityontheconstructions.
Theaimsofthisstudywere(i)toevaluateandcompareanimal andvegetalbiodiversityonthreeriverbanktechniquesforerosion controland(ii)toassessthecapacityofdifferentriverbank pro-tectiontechniquestolimitinvasionofexoticspecies.Theapplied
aimofthisstudywastointegrateecologicalaspectsintothe deci-sionlinebyprovidingecologicaldata.Weprovideguidelinesto practitionersonwhatecologicalconditionstheycanexpectonthe riverbanktechniquechoseninordertodevelopstrategieslinkedto riverbankmanagementandallowrestorationofecosystemgoods andservices.Wehypothesizethatcivilengineeringtechniquesdo notpromotebiodiversityasgoodasbioengineeringtechniques. 2. Materialsandmethods
2.1. Sites
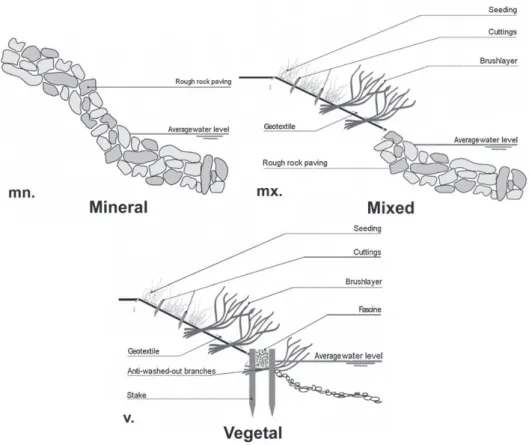
Weassessedbiologicaldiversityonfivereplicatesofthree river-banktechniques(Fig.1):
-Civilengineeringembankments:riprapprotection(rocksusedto stabilizeshorelines,againstscouranderosion)notedas “Min-eral”.
-Mixed embankments combining civil engineering techniques (riprapatthelowerpartofthebank)andbioengineering tech-niques(cuttingsandwoodyplantationattheupperpart)noted as“Mixed”.
-Completely bioengineeringembankments: (willow fascines at thelowerpartofthebankwithcuttingsandwoodyplantation attheupperpart)notedas“Vegetal”.
Thesethreetechniqueswereconsideredasthreelevelofa nat-uralitygradient.Naturalitycanbedefinedbytheamountofliving vegetationusedintheconstructionasclassificationkeycriterion (Brunel,2009;Werdin-Pfistereretal.,2009).Mineralstructures wereconsideredasman-madeenvironmentswithlittleplant cov-erage,differentfromtheenvironmentfoundonnaturalmountain riverbank.Mixed methods showintermediate plant cover with revegetationon halfof thebank.Bioengineering structuresare completelyvegetalandthereforeareexpectedtoshowthe high-estvegetationcoverandthusthehighestlevelofnaturality.These threeengineeringtechniquestherefore constitutethreedistinct levelsonanaturalitygradient.
WeselectedfifteenriverbanksalongsixdistinctFrenchprealps riversbelongingtotheRhonecatchmentarea.Alargeandintense prospectionatterritorialunitsforrivermanagementorganizations wasperformedtolistenoughcomparablebanks.Table1listssites characteristics.Asaltitudeprofoundlyaffectsecological commu-nitycomposition,studysiteswereselectedatanarrowrangeof 270m(between200and470masl).Onlysitesbetween20and 30mlongandwithanestablishedvegetationwerechosenfor anal-ysis.AllthestudysiteswerelocatedinthefoothillsoftheFrench Alpscharacterizedbyhighlyfragmentedhabitats.
2.2. Vegetation
VegetationwasassessedusingthecontactpointmethodinJune andJuly2009.Thismethodencompassesthevertical organiza-tionofthevegetation(tree,shrubandherbaceouslayers).Plant speciesdiversityandfrequencieswereestimatedusinga2-m-long stickwitha diameterof1cm. Measurementsweretakenevery metrealongthree20mtransectsplacedparalleltotheshoreone locatedatthewaterline(Transect1),oneinthemiddle(Transect 2)andoneatthetop(Transect3)oftheriverbankembankment (sixtymeasurementsper site).Vegetationwasidentified tothe specieslevelusingvariousidentificationkeysandflora(Rameau, 1994;LauberandWagner,1998;Aeschimannetal.,2004).Exotic speciesweredefinedonthebasisofestablishedlistsrelevantto thearea(Mulleretal.,2006),alistofexoticinvadersinSwitzerland
Fig.1.Schematicrepresentationofriverbanktechniquesstudied.(a)Civilengineeringembankments:riprapprotection,notedas“Mineral”.(b)Mixedembankments combiningcivilengineering(riprapatthelowerpartofthebank)andbioengineeringtechniques,notedas“Mixed”.(c)Completelybioengineeringembankments:willow fascinesatthelowerpartofthebankwithcuttings,notedas“Vegetal”.
(LandoltandBäumler,2010)andalistestablishedforthe admin-istrativedistrictofIsère(Gentiana,2006).
2.3. Fauna
Coleopteranswereidentifiedtogenus(Insecta,Coleoptera).This order, and more specifically theCarabids, is used as proxy for habitatdiversity,hydromorphologicalprocessesandhabitathealth status(Boscainietal.,2000;Kleinwaïchteretal.,2003).Moreover, morphologicalattributesofColeopteranscanprovideinformation ontheirfunctionalandtrophicstatus(VanLooyetal.,2005).Flying coleopteransweretrappedusingtwoFlora®“YellowWell”traps,
locatedatthetwoextremitiesin themiddle ofthebank (tran-sect2)andplacedatthecanopylevel.TrapsweresetinJuneand
July2009andremainedinthefieldforsevendays.Insectswere identified using a binocular microscope and appropriate refer-enceworks(Picard,1929;Paulian,1941;Hoffmann,1945;Guignot, 1947;Balachowsky,1949).
2.4. Statisticalanalysis
Differencesinplantspeciesandcoleopterangenerarichness, andinfrequencyandnumberofexoticspeciesamongthethree typesofdevelopmentwasassessedusingKruskal–Wallistests.In caseofasignificantmaineffect,pairedMann–Whitneytestswere carriedouttodetectdifferencesbetweeneach technique. Over-allstatisticalriskwasassessedusingtheHolm’scorrection(Holm, 1979).Aninter-classprincipalcomponentanalysis(PCA)(Doledec
Table1
Characteristicsandlocalizationsofembankments.
Sitecode Constructiontype Locality River Latitude Longitude Constructionworkyear Altitude(masl) 6V Vegetal LesEchelles Guiersvif 45◦26’19.15”N 5◦45’36.75”E 2005 393
35V Vegetal VillardBonnot Vorz 45◦15’01.29”N 5◦54’01.62”E 2006 242
7V Vegetal StGeoireenValdaine Ainan 45◦28’02.24”N 5◦39’40.90”E 2004 381
38V Vegetal LeGrandSerre Galaure 45◦15’45.62”N 5◦06’29.65”E 2007 370
39V Vegetal Cluse Arve 46◦04’13.98”N 6◦33’18.72”E 2005 470
6Mix Mixed LesEchelles Guiersvif 45◦26’19.15”N 5◦45’36.75”E 2005 393
30Mix Mixed Grenoble Isère 45◦11’55.65”N 5◦44’02.36”E 2002 210
32Mix Mixed Gière Isère 45◦11’20.94”N 5◦47’20.62”E 2005 210
39Mix Mixed Cluse Arve 46◦04’13.98”N 6◦33’18.72”E 2005 470
40Mix Mixed Bonneville Arve 46◦04’33.18”N 6◦24’26.63”E 2004 444
7Min Mineral StGeoireenValdaine Ainan 45◦28’02.24”N 5◦39’40.90”E 2004 381
35Min Mineral VillardBonnot Vorz 45◦15’01.29”N 5◦54’01.62”E 2006 242
37Min Mineral ChateauneufdeGalaure Galaure 45◦14’09.59”N 4◦58’30.83”E 2004 257
38Min Mineral LeGrandSerre Galaure 45◦15’45.62”N 5◦06’29.65”E 2007 370
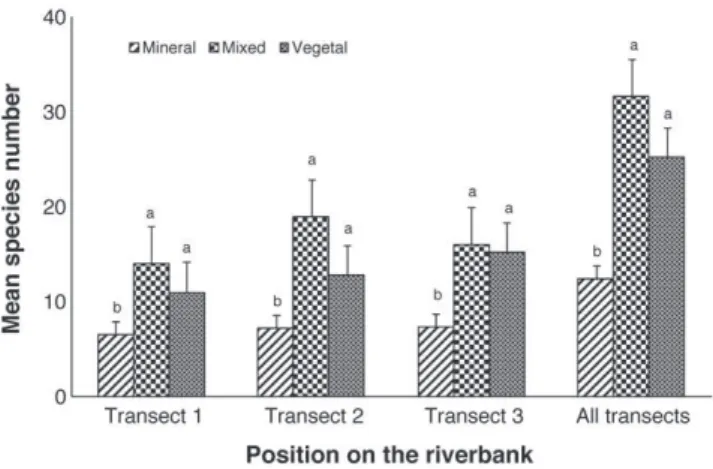
Fig.2.Meanplantspeciesnumbersforeachriverbankprotectiontechnique accord-ingtotheirpositionsonthebank.Differentlettersindicateastatisticaldifference betweensamples.Errorbarsindicatestandarderrors.The“Alltransect”bar repre-sentthemeanofthetotalnumberofplantspeciesfoundforeachtechnique.
and Chessel,1987)wascarriedouttoanalyzetheoverallplant pattern. AMonte-Carlo test wasprocessedtodetectsignificant differencesinfloristic compositiononstructuresusingdifferent engineeringtechniques(MetropolisandUlam,1949).Alltestswere performedusingRstatisticallanguage(R,2005)withade4package (Thioulouseetal.,1997)formultivariateanalysis.
3. Results 3.1. Vegetation
Weidentified126plantspeciesfromthe15sampledsites. Plant species richness differedalong thenaturality gradient (Kruskal–Wallistest:p-value=0.006,Table2).“Vegetal”or“Mixed” techniques having higher species plant diversity compared to “Mineral”technique(Mann–WhitneyUtestwithHolmcorrection: p-value=0.024and0.036,Table2).Therewasnosignificant dif-ferencebetweenmeanspeciesrichnesson“Vegetal”and“Mixed” techniquesalthoughdiversitytendedtobehigheronthelatter, especiallyinthemiddleandatthebottomofthebank(Fig.2).
Floristiccompositiondifferedsignificantlyalongthenaturality gradient(Monte-Carlotest:p-value=0.003).Thedistributionand spacingofthethreesphericalclusters(mn:Mineral;mx:Mixed; v:Vegetal)onthePCAfactorialmapprovidesinformationonthe variabilityoffloristiccompositions(Fig.3).However,somespecies contributemorethanotherstothesegregationofthethreesets,e.g. multipleSalicaceaespecies(SalixviminalisL.,SalixpurpureaL.,Salix triandraL.,SalixfragilisL.,SalixmyrsinifoliaSalisb.andSalix pen-tandraL.)characterized“Vegetal”technique.“Mineral”technique wascharacterizedbyspecieslikeBuddlejadavidiiFranch.,Humulus lupulusL.,RobiniapseudoacaciaL.,UrticadioicaL.and Parthenocis-susquinquefoliaL.“Mixed”techniquewasmainlycharacterizedby thepresenceofCornussanguineaL.,CalamagrostisepigeiosL.,Salix incanaL.,ViburnumopulusL.andLoniceraxylosteumL.(Fig.4).The inertiaoftheinter-classanalysisrepresents18.19% ofthetotal varianceamongthesites.Inaddition,66.16%oftheinformation explainedbythisinter-classPCAwasexplainedbyaxis1.Salicaceae speciesaswellasshrubslikeC.sanguineaandV. opulusmainly explainedaxis1componentonthepositivesideoftheaxis,andto alesserextentbyspecieslikeB.davidiiandR.pseudoacaciaonthe negativeside.Axis2expressed33.84%oftheinformation.Inthe upperpartofthisaxis,thediscriminatoryspeciesweremainly:S. viminalis,S.purpureaandS.triandra.Inthelowerpart,C.sanguinea,
Fig.3. Inter-classanalysisofvegetationonthreeriverbanksprotectiontechnique. mn:Mineral;mx:Mixed;v:Vegetal.Thedistributionandspacingbetweenthethree sphericalclustersprovidesinformationonthevariabilityoffloristiccompositions betweenthedifferenttechniques.Thesizeoftheellipseprovidesinformationabout intra-techniqueplantspeciesvariability.
C.epigeios,S.incanaandV.opuluswerethediscriminatoryspecies (Fig.4).
The area of the ellipse provided information about intra-techniquesplantspeciesvariability(Fig.3).“Mixed”and“Vegetal” techniquesshowcomparablevariabilityofplantspecies composi-tion,higherthanthatof“Mineral”technique.
Fig.4. Spatialdistributionofallspeciesencounteredonthethreeriverbanks pro-tectiontechnique.Speciescodesrefertothefirstthreelettersofthegeniusand thespeciesnames.Percentagesofvarianceobservedaregivenoneachaxis.A correspondencetableisgivenintothesupplementaryfiles.
Table2
ProbabilityvaluesoftheKruskal-WallistestandMann-Whitneytestforvegetationsurvey,flyingcoleopteraandinvasivespecies.
Statisticaltests Pairwisecomparison Vegetationsurvey Aeriancoleoptera Invasivespecies
Probabilityvalues Probabilityvalues Number Frequency
Kruskal–Wallis 0.006** 0.006** 0.308 0.027*
Vegetal/Mixed 0.247 0.083 0.817 1
Mann–Whitney Mixed/Mineral 0.024* 0.03* 0.363 0.063
Vegetal/Mineral 0.036* 0.038* 0.812 0.045*
*Indicatestatisticalsignificantdifferences(p-value<0.05). **Indicatehighlysignificantdifferences(p-value<0.01).
3.2. Fauna
Weidentified42distinctgeneraofflyingColeopterafromthe 14sampledsites(everytrapshavebeenlostforonesitefromthe “Vegetal”technique).Fig.5indicatesthemeannumbersofflying ColeopterageneracollectedinFlora®yellowtrapsforeachthree
techniques.“Vegetal”supportedameanof12.2differentgenera ofcoleopterans,“Mixed”onessupported7.2and“Mineral” tech-niquessupported3.0genera.Ageniusnameslistwasincludedin
supplementaryfiles.
Riverbank techniques significantly influenced the number of coleopterangenera (Kruskal–Wallisp-value=0.006,Table 2). “Mixed”and“Vegetal”techniquesweresignificantlyricherthan “Mineral”technique(Mann–WhitneyUtestwithHolmcorrection: p-value=0.03and0.038;Table2).
WeobservedmanygeneraassociatedwithSalicaceaeincluding varioussaproxylicCetoniaspecieswithlarvaegrowingin decom-posingwood(Cetoniaaurata,Potosiacupreabourginii).Therewere alsoCerambycidaespecieslikeLamia textor,ofwhich thelarvae developinwillowstumpsandroots,Necydalismajorinthehigh cavitiesoftrees,andAromiamoschata,ofwhichthelarvaedevelop in rottingstumps and branches.Many herbivorouscoleopteran specieswereidentified,especiallyVariimordassp.(oftenseenon Apiaceae),Hopliafarinosa(whichfeedsonpollen),Oedemerassp. (the adultof which isherbivorous and thelarva xylophagous). SomewereassociatedwithSalicaceaelikeC.aurata,Chaetocnema aridulaandAlticaaenescens.Predatoryspecieswerealsoidentified, includingthecarnivorousAdrastuslimbatusandOrchesiamicans. 3.3. Exoticplantspecies
Althoughexoticplantspeciesweresometimesfoundonsites thathavebeenconstructedusingbioengineering,theywerenot amongthetendominantspeciesobservedinthestudy.In con-trast, two exotic species, B.davidii and R. pseudoacacia, belong
Fig.5.Coleopterameangenusnumbersinyellowtrapsonthethreeriverbanks pro-tectiontechnique.Differentlettersindicateastatisticaldifferencebetweensamples. Errorbarsindicatestandarderrors.
Fig.6.Exoticmeanspeciesnumbersonthethreeriverbanksprotectiontechnique. Differentlettersindicateastatisticaldifferencebetweensamples.Errorbarsindicate standarderrors.
tothetenmostabundantspecieson“Mineral”structure (100% of observed exotic species). Fallopia sp. was the main genius representedon“Mixed”structure(83%ofobservedexoticspecies) andon“Vegetal”structure(100%ofobservedexoticspecies). Over-all exoticspeciesrichness variedfrom 1.75to 2.2betweenthe techniques (Fig.6)anddidnot differsignificantly betweenthe three techniques(Kruskal–Wallis test p-value=0.308; Table 2). Invasivespeciesfrequencywassignificantlydifferentaccordingto thetechnique(Kruskal–Wallistestp-value=0.027;Fig.7),reaching amaximumonmineraldevelopments(42.5)comparedtovegetal (10)and mixed(13).Frequencyofinvasivespecies was signifi-cantly differentbetween“Mixed” and“Mineral” techniquesbut not between“Vegetal”and “Mixed”techniquesbecauseofhigh variability on“Vegetal”developments(fromf=0.34 tof=22.71; Mann–WhitneyUtestwithHolmcorrection:p-value=0.063and 0.045;Table2).Exoticinvasivespeciesweredifferentaccordingto thetechnique.
Fig.7.Exoticmeanspeciesfrequenciesonthethreeriverbanksprotection tech-nique.Differentlettersindicateastatisticaldifferencebetweensamples.Errorbars indicatestandarderrors.
4. Discussion
4.1. Vegetationdiversity
Thissurvey providesa firstassessment ofplant speciesand Coleoptera genera richness associated with three categories of riverbankprotectiontechniques.Basedonfieldobservations,we highlightsignificantdifferencesinplantdiversitybetween con-ventionalcivilengineeringandbioengineering.Inaccordancewith ourhypothesis,significantlyfewerplantspecieswerefound con-structionusingon“Mineral”structuresthanoneitherofthetwo othersprobablyduetothesubstratein“Mineral”structuresthat physicallylimitscolonizationbyplants. Incontrast,weobserve nosignificantdifferencesinplantspeciesrichnessbetween “Vege-tal”and“Mixed”techniques.Thehighernumberofplantspecies werefoundon“Mixed”structurescomparedto“Vegetal”onesmay beexplainedbyalargeproportionofwillowsfromthefascines installedatthewaterlinewhichshowanimportantgrowthrate; willowsarepartoffast-growing speciesusuallyusedriverbank bioengineering (Adamet al.,2008).Thiswillowcanopy rapidly developsadensecoveragethatcanpartiallyinhibitcolonizationof theunderstorybyherbaceousplants.Forthe“Mixed”technique, thepresenceofvegetationfreeriprapatthebottomofthebank restricts colonizationand coverageby Salicaceae species tothe upperpart.Riprapattheinterfacebetweentheaquatic environ-mentandthefirstwillowcuttingsprovideopenspaceavailablefor colonizationbyhalophyticspecies(Chenetal.,2010).Thislower partconstitutesadifferenthabitat,distinctfromthatofferedby the“Vegetal”techniquewherewoodyspecies,mostlywillows, pre-dominate.Speciestypicallyfoundonthis“Mixed”structuresare(i) typicalwetlandspecieslikeTetragonolobusmaritimus,Juncus acu-tiflorus,CarexpseudocyperusandPhragmitesaustralis,(ii)meadow specieslikeHolcusmollis,(iii)edgespeciesandspeciesofwoodland fringeslikeLigustrumvulgare,L.xylosteumandCrataegus monog-ynaand(iv)treesfromalluvialzoneslikePopulusnigraandPopulus alba.Mostofthosespecieswerenotfoundon“vegetal”structures becauseofthepredominanceofwillowsduetoalackoflightand space.Finally,periodicsedimentationfillsinthegapsbetweenthe blocksandcreatesaloam-richsubstratefavourablefor coloniza-tionbyhelophyticspecies.Ourresultsarethereforeconsistentwith otherstudieswherethepresenceofsomelitterandloamincreases both thedensity and diversity of pioneerplants (Langladeand Décamps,1996).
4.2. Faunadiversity
FlyingColeopteraweremorediverseon“Vegetal”(12.2genera onaverage)and“Mixed”(7.2generaonaverage)structureswhich havemorecomplexvegetationandgreaterfloristicdiversitythan on“Mineral”(3.0generaonaverage)structures.Ourresultsare consistentwithotherstudieswhichshowthattheabundanceof Coleopteranspeciescanberelatedtothepresenceofdifferent veg-etationstrata,fromgrassesthroughbushestotrees(Burel,1989). Ingeneral,insectpopulations––especiallyCarabidae––dependon vegetation structure and heterogeneous habitats (Verdonschot etal.,2007).Alongrivercorridors,hotspotsofbiodiversityare prin-cipallysituatedinregionswherehabitatdiversityishigh(Tews etal.,2004).Suchhabitatheterogeneityprimarilyreflectsgradients ofdisturbance,sedimentgrainsize,moistureandfertilitythatare tightlylinkedtoriverdynamicsthatrecurrentlyrenewsthe habi-tatmosaicanddrivessuccession.Inparticularsmallandmedium floodscreatemorespatialvariabilityandthereforemore hetero-geneity(Pollocketal.,1998;Helfieldetal.,2007).
Although “Mixed”structures harbour greaterbiodiversity in terms of plant species richness (30 species on average), more
generaof flying Coleopterawere foundon “Vegetal”structures (12.2generaonaverage).Thissuggeststhattheplantspecies iden-tityhasamajorimpactoninsectdiversity.Salixspeciesprobably attractsmany nectar-feedinginsectsbyproviding animportant nectarsourceavailableearlyintheyear(inJanuaryandFebruary) when other nectar sources are scarce(Newsholme, 1992).The highnumberofcoleopteraassociatedwithSalicaceaefoundinthis studyconvergeswithNewsholme’s observations.Vegetal cover and biomass werehigher on “Vegetal”structures compared to thetwootherswhatcouldexplainthehighercoleopteran diver-sityon“Vegetal”structures.Moreoverthat shorezonescontain largenumberofpredatorsandscavengersincludingcarabid bee-tlesthatfeedonwrackorcarrionthathasbeendeposedonthe shore(Kleinwächteretal.,2003).Wecanassumethat“Vegetal”and “Mixed”structuressupportmorevegetationcomparedto “Min-eral” structuresand canthus producemore wrack and induce higheranimaldrowningmortality.Ingeneral,shorezonesarean importantfeedingzoneformanyspeciesincludingcarabidspecies (StrayerandFindlay,2010).
4.3. Invasivespecies
Thisstudydidnotrevealanysignificantdifferenceinthe num-berof exotic plants between “Mineral”, “Mixed”and “Vegetal” techniques (respectively 1.75; 2.2 and 1.8). The occurrence of exotic plantspecies hasbeen often related tothe intensity of humanactivity(Pyˇseketal.,2010).Alltechniquesstudiedinvolve heavyhumaninterventionlikeearthmovingordiggingactivities. Itcan beassumedthat thesoilswereallsubject toan equiva-lentdegreeofperturbationand comparablepropagulepressure explainingwhythedifferenttechniquesharbourasimilarnumber ofexoticplantspecies.Incontrast,thisstudyshowsthatthe fre-quencyofexoticspecieswashigheron“Mineral”structuresthan ontheothertechniques.Exotic plantsspecies identifiedin this studyarecharacterizedbyhighgrowthrateswhichgavethema competitiveadvantageinpioneerhabitats(CallandNilsen,2003; Tallent-HalsellandWatt,2009).Moreovertherelativeabundance ofinvasiveplantscouldbeexplainedbybioticinteractions,notably relatedtocompetition(Levineetal.,2004).Thepresenceof com-petitorson“Mixed” and “Vegetal”structures couldrestrict the vigourandinhibitthepropagationofexoticspecieswhatisin par-ticularseenonbioengineeringstructures;whereitisverylikely thatthehighdensityofwillowrestrictedtheperformanceofexotic species. Interestingly,B. davidii wasthe only species on “Min-eral”techniqueandFallopiasp.wastheonlyinvasivespecieson “Vegetal”technique.Asmineralstructurescouldexperiencehigher temperaturesduringsunnyweatherthanvegetalizedbanks,high temperatureonmineraltechniquescouldbeherehypothesized asanenvironmentalfilterforspeciessensitiveforheatstress.B. davidiiisaMediterraneanandtropicalspecies,canbesupposed tostandhightemperatures(Tallent-HalsellandWatt,2009;Watt etal.,2010).Colonisablesubstrataavailablecanalsobean expla-nation;Fallopiasp.rhizomesneedmorespacetodevelopandend upinaviableorganism.
4.4. Perspectivesandapplications
Theapplicativepurposeofthisstudyistoprovideuseful infor-mationtodecision-makersforplanningsustainablemanagement strategiesofmountainriverbanks.Alongsmallrivers,riverbank protectionsshouldbeplantinsteadofself-recoveryofnatural veg-etationin order topromote a rapid vegetal cover and to limit exoticspeciesinvasion.Thebioengineeringtechniquesusing will-ows controls erosion, limit colonization by exotic heliophilous speciesandprovidesfoodresourcesandhabitattoentomofauna.
Salicaceae are particularly adapted to riverbank restoration techniques. The morphological (deep roots, dense coverage), mechanical(flexiblewood)andphysiological(rapidgrowth, pro-duction of large amounts of nectar and pollen) properties of thisgenus limiterosionandensurea goodreturn tonaturality (SchiechtlandStem,1996;FrossardandEvette,2009).However, suchrestorationtechniquesshouldalwaysincludealargediversity ofspeciesandgrowthforms.Thus,creatingdifferentplantstrata (grasses,bushes,andtrees)ensureshabitatdiversity.River restora-tionprogrammecouldalsoaimatincreasinghabitatdiversity.For instance,itispossibletopromotetherecruitmentofhelophytic species by creatinginterface areasbetween thewater and the bank,freeofbushesandtreesandwheresedimentcanbedeposed tofacilitatetheaccumulationofacolonisablesubstrate.“Mixed” techniquewithriprapatthebaseofthebankandbioengineering techniquesontheupperpartrepresentatrade-offbetweenvegetal coveringandhabitatdiversity.
Finally yet importantly, bioengineering structures represent agood alternativeintheFrenchEnvironmentalLaw, as admin-istrative procedures are simpler (Decree n◦93-743 of the law
92-3article10).Forcivil-engineeringstructures,an environmen-talimpactassessmentisrequiredwhatbringstheadministrative delayto6–9monthstoobtainaconstructionpermit.
5. Conclusion
Theintensificationofhumandisturbancealongmountainriver lead to a dramatic decrease in riparianbiodiversity. Maintain-ingbiologicalcontinuitybetweensmallremnantrefugezonesis apriorityfortheconservationofbiodiversitythreatenedspecies (Miller,2006).Thepresentstudyshowsthecapacitiesof restora-tionecology and,more specifically,bioengineering, topromote the recovery of coleopteran and plants of riparian forests. In addition,“Mixed”and“Vegetal”embankmenttechniquesenhance connectivitybetweendifferenthabitatsandthuscontributetothe restorationofecologicalcorridors.Increasedconnectivitybetween fragmentedlandscapepatchesislikelytoincreasegeneticdiversity withinmodifiedlandscapes(Neavesetal.,2009).
Inafurtherstep,thisstudyshouldbeextendedtotheaquatic compartment.Benthic invertebratesseemstobeagood indica-torofnaturalitywiththeirdiversitycorrelatedtotheamountof organicsubstrate(woodand roots)onthebanks(Sudduth and Meyer,2006).Thequestioningcouldalsoberaisedtoafunctional analysisoftherestoredecosystems(LavorelandGarnier,2002; Hooperetal.,2005).
Acknowledgements
Thisworkwouldnothavebeenpossiblewithoutthehelpof theEco-ErosionUnitof theMountainEcosystemResearch Unit ofIrstea,GrenobleandtheUniversityofToulouse,CNRSEcoLab, Toulouse.TheFrench-SwissInterregIVProjectGeni’Alpandthe Agencedel’EauRhone-Alpesarethankedforprovidingfunding. AppendixA. Supplementarydata
Supplementary data associated with this article can be found,intheonlineversion,athttp://dx.doi.org/10.1016/j.ecoleng. 2012.12.105.
References
Adam,P.,Debiais,N.,Gerber,F.,Lachat,B.,2008.Legénievégétal,Unmanuel tech-niqueauservicedel’aménagementetdelarestaurationdesmilieuxaquatiques.
Ministèredel’écologie,dudéveloppementetdel’aménagementdurables,Paris, 290pp.
Aeschimann,D.,Lauber,K.,Moser,D.-M.,Theurillat,J.-P.,2004.FloraAlpina.Belin, Bern,2672pp.
Balachowsky,A.,1949.FaunedeFrance(tome50):ColéoptèresScolytides. Lecheva-lier,Paris,320pp.
Boscaini,A.,Franceschini,A.,Maiolini,B.,2000.Riverecotones:carabidbeetlesasa toolforqualityassessment.Hydrobiologia422,173–181.
Breton,V.,Forestier,O.,Guindon,O.,Evette,A.Ecologicalrestorationunderpressure frominvasiveanimalspecies:useofSalicaceaecuttingsinariverbank.River ResearchandApplications,underreview.
Brunel,J.P.,2009.Sourcesofwaterusedbynaturalmesquitevegetationina semi-aridregionofnorthernMexico.Hydrol.Sci.J.54(2),375–381.
Burel,F.,1989.Landscapestructureeffectsoncarabidbeetlesspatialpatternsin westernFrance.Landsc.Ecol.2(4),215–226.
Call, L.J.,Nilsen, E.T.,2003.Analysisofspatial patternsandspatialassociation betweentheinvasivetree-of-heaven(Ailanthusaltissima)andthenativeblack locust(Robiniapseudoacacia).Am.Midl.Nat.150(1),1–14.
Chen,S.Y.,Zhang,J.L.,Jia,P.,Xu,J.,Wang,G.,Xiao,S.,2010.Effectsofsizevariation andspatialstructureonplasticresponseofplantheighttolightcompetition. Chin.Sci.Bull.55(12),1135–1141.
Décamps,H.,2011.Rivernetworksasbiodiversityhotlines.C.R.Biol.334(5–6), 420–434.
Décamps,H.,Planty-Tabacchi,A.M.,Tabacchi,E.,1995.Changesinthehydrological regimeandinvasionsbyplantspeciesalongripariansystemsoftheAdourRiver, France.Regul.Rivers:Res.Manage.11(1),23–33.
Doledec,S.,Chessel,D.,1987.Seasonalsuccessionsandspatialvariablesin freshwa-terenvironments.I.Descriptionofacompletetwo-waylayoutbyprojection of variables (Rythmessaisonniers et composantes stationnellesen milieu aquatique.I.Descriptiond’unpland’observationcompletparprojectionde variables).ActaOecol.8(3),403–426.
Evette,A.,Balique,C.,Lavaine,C.,Rey,F.,Prunier,P.,2012.Usingecologicaland biogeographicalfeaturestoproduceatypologyoftheplantspeciesusedin bioengineeringforriverbankprotectionineurope.RiverRes.Appl.28(10), 1830–1842.
Evette,A.,Frossard,P.A.,2009.Lesvégétauxontdugénie.EspacesNaturels26, 35–37.
Frossard,P.A.,Evette,A.,2009.Legénievégétalpourlaluttecontrel’érosionen rivière:unetraditionmillénaireenconstanteévolution.I.E.A.T.,99–109. Gentiana,2006.Lesespècesenvahissantesdel’Isère.Conseilgénéraldel’Isère,
Grenoble,32pp.
Gray,D.,Sotir,R.,1996.BiotechnicalandSoilBioengineeringSlopeStabilization– APracticalGuideforErosionControl.JohnWileyandSons,Inc.,NewYork,400 pp.
Guignot,F.,1947.FaunedeFrance(tome48):ColéoptèresHydrocanthares. Lecheva-lier,Paris,294pp.
Helfield,J.M.,Capon,S.J.,Nilsson,C.,Jansson,R.,Palm,D.,2007.Restorationofrivers usedfortimberfloating:effectsonriparianplantdiversity.Ecol.Appl.17(3), 840–851.
Helfield,J.M.,Engström,J.,Michel,J.T.,Nilsson,C.,Jansson,R.,2012.Effectsofriver restorationonriparianbiodiversityinsecondarychannelsofthePiteRiver, Sweden.Environ.Manage.49(1),130–141.
Hoffmann, A., 1945. Faune de France (tome 44): Coléoptères Bruchides et Anthribides.Lechevalier,Paris,184pp.
Holm,S.,1979.Asimplesequentiallyrejectivemultipletestprocedure.Scand.J.Stat. 6(2),65–70.
Hooper, D.U.,Chapin, F.S.,Ewel,J.J.,Hector, A.,Inchausti, P.,Lavorel,S., Law-ton,J.H.,Lodge,D.M.,Loreau,M.,Naeem,S.,Schmid,B.,Setala,H.,Symstad, A.J.,Vandermeer, J.,Wardle,D.A.,2005. Effects ofbiodiversityon ecosys-temfunctioning: aconsensusofcurrentknowledge.Ecol. Monogr.75(1), 3–35.
IFEN,2006.L’environnementenFrance–Edition.IFEN,500pp.
Isbell,F.,Calcagno,V.,Hector, A.,Connolly,J.,Harpole,W.,Reich, P., Scherer-Lorenzen,M.,Schmid,B.,Tilman,D.,vanRuijven,J.,Weigelt,A.,Wilsey,B., Zavaleta,E.,Loreau,M.,2011.Highplantdiversityisneededtomaintain ecosys-temservices.Nature477(7363),199–202.
Januschke,K.,Brunzel,S.,Haase,P.,Hering,D.,2011.Effectsofstreamrestorations onriparianmesohabitats,vegetationandcarabidbeetles.Biodivers.Conserv.20 (13),3147–3164.
Kleinwächter,M.,Eggers,T.O.,Henning,M.,Anlauf,A.,Hentschel,B.,Larink,O.,2003. Distributionpatternsofterrestrialandaquaticinvertebratesinfluencedby dif-ferentgroyneformsalongtheRiverElbe(Germany).LargeRivers15(1–4), 319–338.
Kleinwaïchter,M.,Larink,O.,Eggers,T.,Anlauf,A.,2003.Impactofmodifiedgroynes onCarabidbeetles.In:Proceedingsofthe“LowlandRiverRehabilitation2003” Conference,Wageningen,TheNetherlands.
Landolt, E., Bäumler, B., 2010. Flora indicativa:ökologische Zeigerwerte und biologischeKennzeichen zurFlorader Schweizundder Alpen=ecological indicator values andbiological attributesofthe Floraof Switzerlandand theAlps.ConservatoireetJardinbotaniquesdelavilledeGenève,Genève, 376pp.
Langlade,L.-R.,Décamps,O.,1996.Effetdel’accumulationdelimonsurla colonisa-tionvégétaled’unbancdegaletsenrivière.C.R.Acad.Sci.,Paris318,1076–1082. Lauber,K.,Wagner,G.,1998.FloraHelvetica,secondedition.P.HauptEditions,
Lavorel,S.,Garnier,E.,2002.Predictingchangesincommunitycompositionand ecosystemfunctioningfromplanttraits:revisitingtheHolyGrail.Funct.Ecol. 16(5),545–556.
Levine,J.M.,Adler,P.B.,Yelenik,S.G.,2004.Ameta-analysisofbioticresistanceto exoticplantinvasions.Ecol.Lett.7(10),975–989.
Li,M.H.,Eddleman,K.E.,2002.Biotechnicalengineeringasanalternativeto tradi-tionalengineeringmethods– abiotechnicalstreambankstabilizationdesign approach.Landsc.UrbanPlan.60(4),225–242.
Li,X.P.,Zhang,L.Q.,Zhang,Z.,2006.Soilbioengineeringandtheecological restora-tionofriverbanksattheAirportTown,Shanghai,China.Ecol.Eng.26(3), 304–314.
Maestre,F.T.,Quero,J.L.,Gotelli,N.J.,Escudero,A.,Ochoa,V.,Delgado-Baquerizo, M.,García-Gómez,M.,Bowker,M.A.,Soliveres,S.,Escolar,C.,García-Palacios, P.,Berdugo,M.,Valencia,E.,Gozalo,B.,Gallardo,A.,Aguilera,L.,Arredondo,T., Blones,J.,Boeken,B.,Bran,D.,Conceic¸ão,A.A.,Cabrera,O.,Chaieb,M.,Derak, M.,Eldridge,D.J.,Espinosa,C.I.,Florentino,A.,Gaitán,J.,Gatica,M.G.,Ghiloufi, W.,Gómez-González,S.,Gutiérrez,J.R.,Hernández,R.M.,Huang,X., Huber-Sannwald,E.,Jankju,M.,Miriti,M.,Monerris,J.,Mau,R.L.,Morici,E.,Naseri, K.,Ospina,A.,Polo,V.,Prina,A.,Pucheta,E.,Ramírez-Collantes,D.A.,Romão, R.,Tighe,M.,Torres-Díaz,C.,Val,J.,Veiga,J.P.,Wang,D.,Zaady,E.,2012.Plant speciesrichnessandecosystemmultifunctionalityinglobaldrylands.Science 335(6065),214–218.
McNeely,J.A.,2010.Biodiversity,EcosystemFunctioning,&HumanWellbeing:An EcologicalandEconomicPerspectiveeditedbyShahidNaeem,DanielE.Bunker, AndyHector,MichelLoreauandCharlesPerrings(2009),xiv+368pp.,Oxford UniversityPress,Oxford,UK.ISBN9780199547951(hbk),GBP75.00;ISBN 9780199547968(pbk),GBP37.50.Oryx44(03),461.
Metropolis,N.,Ulam,S.,1949.TheMonteCarlomethod.J.Am.Stat.Assoc.44(247), 335–341.
Miller,J.R.,2006.Restoration,reconciliation,andreconnectingwithnaturenearby. Biol.Conserv.127(3),356–361.
Muller,S.,Aboucaya,A.,Affre,L.,Cassan,S.,2006.PlantesinvasivesenFrance,168 pp.
Neaves,L.E.,Zenger,K.R.,Prince,R.I.T.,Eldridge,M.D.B.,Cooper,D.W.,2009. Land-scapediscontinuitiesinfluencegeneflowandgeneticstructureinalarge,vagile Australianmammal,Macropusfuliginosus.Mol.Ecol.18(16),3363–3378. Newsholme,C.,1992.Willows:TheGenusSalix.Batsford,London,226pp. Nilsson,C.,Reidy,C.A.,Dynesius,M.,Revenga,C.,2005.Fragmentationandflow
regulationoftheworld’slargeriversystems.Science308(5720),405–408. Norris,J.E.L.,Stokes,A.D.,Nicoll,A.,Achim,B.C.A.,2008.SpeciesSelectionforSoil
ReinforcementandProtection.SlopeStabilityandErosionControl.Springer. Dordrecht,EcotechnologicalSolutions,pp.167–210.
Pahl-Wostl, C.,2006. Theimportanceofsociallearningin restoringthe mul-tifunctionalityofriversandfloodplains.Ecol.Soc.11(1),Availableonline:
http://www.ecologyandsociety.org/vol11/iss1/art10/
Paulian,R.,1941.FaunedeFrance(tome38):ColéoptèresScarabéides.Lechevalier, Paris,240pp.
Pezeshki,S.R.,Li,S.,ShieldsJr.,F.D.,Martin,L.T.,2007.Factorsgoverningsurvival ofblackwillow(Salixnigra)cuttingsinastreambankrestorationproject.Ecol. Eng.29(1),56–65.
Picard,F.,1929.FaunedeFrance(tome20):ColéoptèresCerambycîdae.Lechevalier, Paris,168pp.
Poff,N.L.,Allan,J.D.,Bain,M.B.,Karr,J.R.,Prestegaard,K.L.,Richter,B.D.,Sparks,R.E., Stromberg,J.C.,1997.Thenaturalflowregime:anewparadigmforriverine conservationandrestoration.Bioscience47,769–784.
Pollock,M.M.,Naiman,R.J.,Hanley,T.A.,1998.Plantspeciesrichnessinriparian wetlands– atestofbiodiversitytheory.Ecology79(1),94–105.
Pyˇsek,P.,Jaroˇsík,V.,Hulme,P.E.,Kühn,I.,Wild,J.,Arianoutsou,M.,Bacher,S., Chiron,F.,Didˇziulis,V.,Essl,F.,Genovesi,P., Gherardi,F.,Hejda,M.,Kark, S.,Lambdon,P.W.,Desprez-Loustau,M.L., Nentwig,W.,Pergl,J.,Poboljˇsaj, K.,Rabitsch,W.,Roques,A.,Roy,D.B.,Shirley,S.,Solarz,W.,Vilà,M., Win-ter,M.,2010.Disentanglingtheroleofenvironmentalandhumanpressures onbiologicalinvasionsacrossEurope.Proc.Natl.Acad.Sci.U.S.A.107(27), 12157–12162.
R,D.C.T.,2005.R:ALanguageandEnvironmentforStatisticalComputing.R Foun-dationforStatisticalComputing,Austria.
Rameau,J.C.,1994.FloreForestièreFranc¸aise;Tome1:PlainesEtCollines.Institut PourLeDéveloppementForestier.Ministèredel’AgricultureetdelaForêt,Paris, 1785pp.
Schiechtl,H.M.,Stem,R.,1996.WaterBioengineeringTechniquesforWatercourse BankandShorelineProtection.BlackwellScience,Oxford,UK,186pp. Stohlgren,T.,Bull,K.,Otsuki,Y.,Villa,C.,Lee,M.,1998.Riparianzonesashavensfor
exoticplantspeciesinthecentralgrasslands.PlantEcol.138(1),113–125. Strayer,D.L.,Findlay,S.E.G.,2010.Ecologyoffreshwatershorezones.Aquat.Sci.72
(2),127–163.
Sudduth,E.,Meyer,J.,2006.Effectsofbioengineeredstreambankstabilizationon bankhabitatandmacroinvertebratesinurbanstreams.Environ.Manage.38 (2),218–226.
Tallent-Halsell,N.G.,Watt,M.S.,2009.TheinvasiveBuddlejadavidii(butterflybush). Bot.Rev.75(3),292–325.
Tews,J.,Brose,U.,Grimm,V.,Tielbörger,K.,Wichmann,M.C.,Schwager,M.,Jeltsch, F.,2004.Animalspeciesdiversitydrivenbyhabitatheterogeneity/diversity:the importanceofkeystonestructures.J.Biogeogr.31(1),79–92.
Thioulouse,J.,Chessel,D.,Doledec,S.,Olivier,J.M.,1997.ADE-4:amultivariate analysisandgraphicaldisplaysoftware.Stat.Comput.7(1),75–83.
Tockner,K.,Stanford,J.A.,2002.Riverinefloodplains:presentstateandfuture trends.Environ.Conserv.29(3),308–330.
VanLooy,K.,Vanacker,S.,Jochems,H.,DeBlust,G.,Dufrene,M.,2005.Ground beetlehabitat templets and riverbank integrity.River Res. Appl.21(10), 1133–1146.
Verdonschot, R., Noordijk, J., Sykora, K., Schaffers, A., 2007. Ground beetles (Coleoptera:Carabidae)withinavegetationgradientintheMillingerwaard. Entomol.Ber.(Amst.)67(3),82–91.
Watt, M.S., Kriticos, D.J., Potter, K.J.B., Manning, L.K., Tallent-Halsell, N., Bourdot, G.W., 2010. Using species niche models to inform strategic management of weeds in a changing climate. Biol. Invasions 12 (11), 3711–3725.
Werdin-Pfisterer,N.R.,Kielland,K.,Boone,R.D.,2009.Soilaminoacid composi-tionacrossaborealforestsuccessionalsequence.SoilBiol.Biochem.41(6), 1210–1220.