Colonisation, par les coléoptères
phloéophages et xylophages, de sapins
baumiers défoliés par l’arpenteuse de la
pruche
Mémoire
Jean-Michel Béland
Maîtrise en sciences forestières
Maître ès sciences (M. Sc.)
Québec, Canada
Colonisation, par les coléoptères
phloéophages et xylophages, de sapins
baumiers défoliés par l’arpenteuse de la
pruche
Mémoire
Jean-Michel Béland
Sous la direction de :
Éric Bauce, directeur de recherche
Christian Hébert, codirecteur de recherche
iii
Résumé
L‟objectif de cette étude était de caractériser l‟impact d‟une épidémie d‟arpenteuse de la pruche dans la sapinière à bouleau blanc, sur la communauté de coléoptères saproxyliques primaires. Pour ce faire, nous avons dans un premier temps comparé différentes approches afin de déterminer laquelle d‟entre-elles pouvait facilement et efficacement estimer l‟état de défoliation des arbres affectés par l‟arpenteuse de la pruche. Par la suite, la réponse hâtive des coléoptères phloéo- et xylophages a été évaluée à l‟aide de pièges à impacts multidirectionnels, de pièges à impact fixés aux troncs et de cages d‟émergence. Les résultats montrent qu‟une méthode d‟estimation globale de la cime supérieure est optimale pour déterminer l‟état de défoliation du sapin baumier suite au passage de l‟arpenteuse de la pruche. Ils montrent aussi qu‟un changement dans l‟assemblage des coléoptères est survenu seulement en 2014 et dans les peuplements les plus défoliés. Un assemblage caractérisé par des espèces polyphages souvent associées aux forêts non-perturbées s‟est remanié en une prédominance d‟un xylomycétophage consommateur de conifère mourant: le scolyte birayé. Seul ce scolyte a été positivement associé avec la hausse de défoliation et de mortalité du sapin baumier mais aussi de la présence de coupes de récupération à proximité des parcelles étudiées. Les résultats ont également montré une réponse dichotomique du scolyte montrant une colonisation massive seulement au moment où les sapins atteignaient 95% de défoliation. Enfin, les pièges à impact se sont avérés utiles pour détecter les variations de fréquence de visite de peuplements et d‟arbres le long du gradient de défoliation. Le scolyte birayé est donc le premier et presque le seul saproxylique à engager le processus de décomposition des arbres post-arpenteuse de la pruche deux années après la détection de l‟épidémie. Par ailleurs, cette colonisation risque d‟entraîner des désagréments car son symbionte fongique noircit les galeries creusées partout dans l‟aubier des arbres infestés.
iv
Abstract
The aim of this study was to characterise the impact of a hemlock looper outbreak, in the balsam fir-white birch forest, on the community of primary saproxylic beetles. To this end, we first compared different approaches in order to determine which of them could profile easily and efficiently the defoliation status of the trees affected by the hemlock looper. Then, the early response of the phloeo- and xylophagous beetles was assessed with flight-interception multidirectional and trunk-window traps along with emergence cages. Results show that an upper crown overall estimation method is optimal to determine the defoliation status of the balsam fir following hemlock looper. They as well show that a change in the beetle assemblage occurred only in 2014 and in the most defoliated stands. An assemblage characterised by polyphagous species often associated to undisturbed forests shifted to the predominance of a xylomycetophagous and dying conifer feeder: the striped ambrosia beetle. This scolytid alone was positively associated with the increase of defoliation and death of balsam firs but also with the presence of salvage logging nearby the study plots. Results have then showed a dichotomic response of the beetle, thus exhibiting massive colonization only when fir trees reached 95% of defoliation. At last, the flight-interception traps were found to be useful to detect variations in beetle visit frequencies of stands and trees along the defoliation gradient. The striped ambrosia beetle is therefore the first and almost the only saproxylic to initiate the decomposition process of post-hemlock looper firs trees two years after the detection of the outbreak. However, this colonization may raise concerns because its fungal symbiont stains the galleries throughout the sapwood of infested trees.
v
Table des matières
Résumé ... iii
Abstract ... iv
Table des matières ... v
Liste des tableaux ... vii
Liste des figures ...ix
Remerciements ... xii
A vant -Propos ... xiii
Introduction générale ... 1
Successions végétales et animales après perturbations naturelles ... 2
Une première épidémie d‟arpenteuse de la pruche dans la Région de la Capitale-Nationale ... 4
La coévolution des coléoptères phloéophages et xylophages avec leurs hôtes ... 6
La colonisation par les coléoptères phloéophages et xylophages en contexte d‟épidémie d‟arpenteuse de la pruche ...11
CHAPITRE 1 - Comparison of several methods for estimating defoliation from the hemlock looper (Lambdina fi scellaria (Guenée)) (Lepidoptera: Geometridae) on balsam fir ...15
Résumé ...16
Abstract ...17
1 Introduction ...18
2 Methods ...20
2.1 Study area ...20
2.2 Stand selection and defoliation estimation ...20
2.3 Statistical analysis ...21 3 Results ...23 4 Discussion...25 Acknowledgements ...28 Referenc es cited...29 Tables ...31 Figures...35
CHAPITRE 2 – Early succession of bark and wood boring beetles during an outbreak of the hemlock looper (Lambdina fiscellaria (Guenée)) (Lepidoptera: Geometridae) in boreal balsam fir fore st ...40 Résumé ...41 Abstract ...42 1 Introduction ...43 2 Methods ...47 2.1 Study area ...47 2.2 Stand description ...47 2.3 Beetle sampling ...48 2.4 Statistical analysis ...49 3 Results ...52
vi
3.1 Early succession of saproxylic beetles in defoliated balsam fir stands ...52
3.2 The variation in saproxylic beetle assemblages explained by environmental variables ...53
4 Discussion...55
Acknowledgements ...59
Referenc es cited...60
Tables ...64
Figures...73
CHAPITRE 3 - Numerical response of the striped ambrosia beetle (Coleoptera: Curculionidae) to hemlock looper (Lepidoptera: Geometridae) defoliation in boreal balsam fir fore sts ...78 Résumé ...79 Abstract ...80 1 Introduction ...81 2 Methods ...83 2.1 Study area ...83 2.2 Stand description ...83 2.3 Beetle sampling ...84 2.4 Statistical analysis ...85 3 Results ...86
3.1 Colonization rates vs tree defoliation ...86
3.2 Visit rates vs tree defoliation ...87
3.3 Flight activity vs stand defoliation ...88
4 Discussion...89
4.1 Host selection and colonization processes ...89
4.2 Forecasting massive colonization to improve salvage logging ...91
Acknowledgements ...93
Referenc es cited...94
Figures...97
Conclusion générale... 102
Comment expliquer la prédominance du scolyte birayé ... 103
Implications pour l‟industrie forestière et perspectives de recherche ... 104
Référenc es citées ... 106
vii
Liste des tableaux
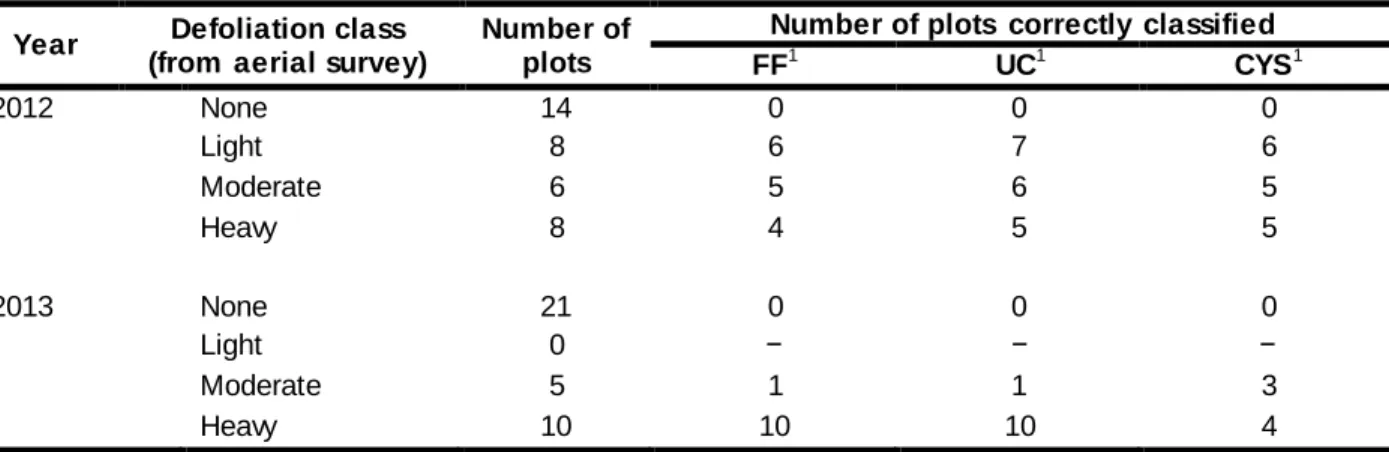
Table 1.1 List of defoliation estimation methods compared in this study. ...31 Table 1.2 Accuracy of defoliation classes determined through aerial survey of hemlock looper
defoliation, according to three ground-level methods estimating defoliation (FF, UC and CYS) on balsam fir trees in 2012 and 201 3 in the Laurentian Wildlife Reserve, Quebec, Canada. ...32
Table 1.3 Regressions between defoliation on Current-Year Shoots and that on each of n−1 to n−4 shoots, as well as with the average defoliation on n-1 to n-4 shoots, also called Previous-Years defoliation (PY). P-values are shown for linear regressions and R² are shown for both linear and non-linear regressions. ...33
Table 1.4 Regressions between defoliation estimated using the Field Fettes (FF) method and that estimated on current-year shoots (CYS) or by the upper crown (UC) method. Significance of Intercept (β0) and slope, (β1) coefficients with standard error (SE), t-statistics, and P-values are shown with the adjusted R² of each linear model. ...34
Table 2.1 Abundance of Cerambycidae and Curculionidae, captured in 2013 and 2014, with multidirectional flight-interception (MFIT), trunk-window (TWT) and emergence cage (EC) traps. Species marked in bold account for more than 1% of total capture. ...72
ix
Liste des figures
Figure 1.1 Validation of aerial survey defoliation classes using defoliation estimated with t hree ground-level methods (Field Fettes, Upper Crown estimates and Current -Year Shoot estimates) in 2012 and 2013. Significantly different classes have different letters, after Tukey pair-wise comparison, based on significant ANOVAs. Dot symbols represent sample trees, Middle bar of the box represents mean defoliation per treatment (with letters indicating differences between means above each boxplot) and box outlines represent the standard error. Grey areas represent % defoliation interval of MFFP classes. For the None class, distinct plots from our controls were represented by red dots. Completely defoliated plots in 2012 were also represented by red dots in 2013 boxplots ...36
Figure 1.2 Relationships between annual defoliation estimated on Current -Year Shoots and cumulative defoliation estimated on previous year shoots (PY; n−1 to n−4 mean) in 2012 and 2013 as assessed by the Field Fettes method. The grey shading in 3b show trees on which current-year shoots were moderate− heavily defoliated, according to MFFP classification. ...37
Figure 1.3 Relationships between two defoliation proxies on balsam fir trees and a complete estimation of defoliation using the Field Fettes method in 2012 and 2013 in the Laurentian Wildlife Reserve in Quebec, Canada. a) Defoliation estimated in the Upper Crown in 2012. b) Defoliation estimated on Current-Year Shoots in 2012. c) Defoliation estimated in the Upper Crown in 2013. d) Defoliation estimated on Current-Year Shoots in 2013. ...38
Figure 2.1 Principal components analyses (PCA) correlation biplots (scaling = 2) of the saproxylic beetle assemblages in balsam fir stands shortly (1−2 years) after a hemlock looper) outbreak in the Laurentian Wildlife Reserve. Biplots show ordinations of beetle assemblages in (a) multidirectional flight-interception traps, (b) trunk-window traps and (c) emergence cages. Dots show the position of each trap (or plot for multidirectional flight -interception traps), and dot filling represent the defoliation level: white for light, hatched for moderate, grey for high and black for dead. The white filled triangles represent the control plots or trees. Beetle species with a score < 0.1 are marked with a + symbol and species with a score > 0.1 are identified on their score coordinates by a species code. The species codes are the following: Draut (Dryocoetes autographus), Drcar (Dryocoetes caryi), Deruf (Dendroctonus rufipennis), Evmon (Evodinus monticola), Hypin (Hylobius pinicola), Pispa (Pityok teines sparsus), Pidub (Pissodes
dubius (striatulus)), Pisim (Pissodes similis), Poruf (Polygraphus rufipennis), Rhmac
(Rhyncolus macrops), Trbet (Trypodendron betulae), Trlin (Trypodendron lineatum). The 2013 and 2014 mean plot or tree scores are shown. ...74
Figure 2.2 Canonical redundancy analyses (RDA) correlation triplots (scaling = 2) of the saproxylic beetle assemblages in balsam fir stands shortly two years after a hemlock looper outbreak in the Laurentian Wildlife Reserve and the variables explaining the species succession. Triplots show ordinations of beetle assemblages in (a) multidirectional flight -interception traps, (b) trunk-window traps and (c) emergence cages. Dots show the position of each trap (or plot for multidirectional flight-interception traps), and dot filling represent the defoliation level: white for light, hatched for moderate, grey for heavy and black for dead. The white filled triangles represent the control plots or trees. Beetle species with a score < 0.1 are marked with a + symbol and species with a score > 0.1 are identified on their score coordinates by a species
x
code. The species codes are the following: Crruf (Cryphalus r. ruficollis), Cusp (Curculionidae
sp.), Draut (Dryocoetes autographus), Drcar (Dryocoetes caryi), Deruf (Dendroctonus rufipennis), Evmon (Evodinus monticola), Pispa (Pityok teines sparsus), Pidub (Pissodes dubius (striatulus)), Pisim (Pissodes similis), Poruf (Polygraphus rufipennis), Rhmac
(Rhyncolus macrops), Scann (Scierus annectans), Trbet (Trypodendron betulae), Trlin (Trypodendron lineatum). The dotted vectors represent the continuous explanatory variables: DDWV (down dead wood volume), Dead A. balsamea (dead balsam fir basal area), Defoliation (upper crown defoliation percentage), Hardwood (hardwood basal area), Logging (logged area in a 500 m buffer zone of the plot) and Soft wood (softwood basal area). ...76
Figure 3.1 Abundance scatter plot of striped ambrosia beetles (Trypodendron lineatum) as sampled by emergence cages in relation with upper crown overall defoliation. Cages placed on fir trees totally defoliated in 2012 are in red. Dashed vertical line represents the defoliation threshold at which massive emergences occurred. a) 2013 collect. b) 2014 collect. ...97
Figure 3.2 Results of binomial (GLMM) regressions between upper crown overall defoliation and the 2014 presence/absence of striped ambrosia beetles (Trypodendron lineatum) as sampled by emergence cages. Black points represent occurred events and their size represents the event frequencies at a defoliation status. Points at 0.00 represent absences while points at 1.00 represent presences. Cages placed on fir trees totally defoliated in 2012 are in red. Dashed vertical line represents the defoliation threshold at which massive emergences occurred. Predicted colonisation probabilities with fixed and nested effects are represented by grey points. ...98
Figure 3.3 Results of negative binomial (GLMM) regressions between upper crown overal l defoliation and the abundance of striped ambrosia beetles (Trypodendron lineatum) as sampled by trunk-window traps. Traps placed on fir trees totally defoliated in 2012 are in red. Predicted abundances with fixed and nested effects are represented by grey points. a) 2013 collect. b) 2014 collect. ...99
Figure 3.4 Results of negative binomial (GLMM) regressions between the mean upper crown overall defoliation and the abundance of striped ambrosia beetles (Trypodendron lineatum) as sampled by multidirectional flight-interception traps. Traps placed in stands totally defoliated in 2012 are in red. a) 2013 collect. b) 2014 collect without the circled outlier trap. ... 100
xii
Remerciements
Je voudrais remercier premièrement mon directeur de recherche, M. Éric Bauce, pour la grande liberté m‟ayant été accordée et pour m‟avoir donné la chance de faire une maîtrise à l‟Université Laval. Je veux ensuite remercier M. Conrad Cloutier (co-directeur) qui m‟a introduit à la recherche en entomologie et qui m‟a, par tout hasard, ouvert la porte vers un monde qui m‟était alors inconnu; l‟entomologie forestière. J‟apprécie qu‟il m‟ait supporté depuis le Baccalauréat jusqu‟à aujourd‟hui. Je voudrais ensuite remercier M. Richard Berthiaume qui s‟est impliqué et qui a fait un travail énorme dans le développement des projets de recherches et dans la rédaction de mon mémoire. N‟étant qu‟un membre de mon comité d‟encadrement à la maîtrise, il a fait un travail digne d‟un co-directeur et il m‟est important de le souligner.
Je voudrais maintenant remercier infiniment M. Christian Hébert, mon codirecteur, pour m‟avoir accueilli au sein de son équipe et de son laboratoire au Centre de Foresterie des Laurentides. Grâce à ce chercheur passionné, j‟ai découvert l‟entomologie forestière, j‟ai pu vivre des expériences à travers le Québec et développer ainsi un grand enthousiasme pour mon projet et pour la foresterie en général. J‟ai bénéficié de ses vastes connaissances et de nombreuses heures de son temps afin de perfectionner mon projet de maîtrise et surtout pour l‟écriture de mon mémoire. Étant « son » seul étudiant à la maîtrise, j‟ai pu être constamment sous sa loupe pour le meilleur et pour le pire. Je n‟aurais pas pu accomplir de tels projets sans son esprit critique, la confiance qu‟il avait en moi et bien sûr, son généreux support financier.
Je tiens à remercier les différents membres du laboratoire qui ont été présents lorsque j‟avais besoin d‟eux. Merci à M. Georges Pelletier pour m‟avoir aidé à identifier des insectes saproxyliques et pour avoir confirmé toutes mes identifications. Merci beaucoup à M. Yves Dubuc pour avoir aidé à développer le dispositif d‟échantillonnage de mon projet, pour m‟avoir assister dans plusieurs expériences et travaux de terrain et pour m‟avoir supporter comme seul voisin de bureau pendant plus de trois ans. Un autre merci à Anne Cotton-Gagnon, Alexis Schab, Charlène Saugey, Étienne Giguère, Guillaume Plante et Laure Guillerand, étudiants de l‟Université Laval ou employés du consortium iFor, pour toute l‟aide dont j‟ai bénéficié afin d‟accomplir les travaux de terrain et de laboratoire. Un grand merci à Jonathan Boucher pour m‟avoir ouvert au monde des statistiques et pour m‟avoir grandement aidé lorsque j‟étais dans l‟impasse. Un gros merci à Olivier Jeffrey pour avoir été mon mentor pendant toutes ces années, pour m‟avoir aidé de toutes les manières possibles dans l‟accomplissement de la maîtrise et enfin pour avoir été un collègue satisfaisant.
Enfin, je voudrais remercier mes parents et ma sœur pour m‟avoir toujours supporté, encouragé et pour être toujours présents quand je suis dans le besoin. Merci à tous d‟avoir fait grandir une passion et d‟avoir rendu possible cet accomplissement.
xiii
Avant-Propos
Ce mémoire de maîtrise est constitué de trois chapitres, « Comparison of several methods for estimating defoliation from the hemlock looper (Lambdina fiscellaria (Guenée)) (Lepidoptera: Geometridae) on balsam fir », « Early succession of bark and wood boring beetles during an outbreak of the hemlock in boreal balsam fir forest » et « Numerical response of the striped ambrosia beetle (Coleoptera: Curculionidae) to hemlock looper (Lepidoptera: Geometridae) defoliation in boreal balsam fir forest ». Ces trois chapitres rédigés en anglais et sous la forme d‟articles scientifiques sont destinés à être publiés dans des journaux scientifiques au cours de l‟année 2017. Ces chapitres sont précédés d‟une introduction et suivis d‟une conclusion rédigées en français. Le candidat à la maîtrise a effectué l‟installation du dispositif d‟échantillonnage, l‟estimation de la défoliation des arbres du dispositif, l‟échantillonnage des insectes saproxyliques, la description des attributs de l‟habitat dans les sites d‟étude, le tri, l‟identification et le montage de la collection d‟insectes étudiés et a fait les différentes analyses statistiques. Les trois chapitres ont été écrits entièrement par le candidat à la maîtrise, encadré et soutenu par les commentaires et conseils des coauteurs Christian Hébert, Conrad Cloutier, Richard Berthiaume et Éric Bauce. L‟étudiant a donc le statut d‟auteur principal pour les trois articles paraissant dans ce mémoire et les coauteurs, énumérés à la phrase précédente, ont été également impliqués dans la conception des trois projets et articles. Cette étude a été financée par le consortium de recherche sur les insectes forestiers (iFor) sous des contributions du Conseil de recherches en sciences naturelles et en génie du Canada (CRSNG), le Ministère des Forêts, de la Faune et des Parcs du Québec (MFFP), le Conseil de l‟industrie forestière du Québec (CIFQ), le Service Canadien des Forêts (SCF-RNCan) et de la Société de Protection des Forêts contre les Insectes et Maladies (SOPFIM).
1
Introduction générale
La zone bioclimatique boréale s‟étend dans tout l‟hémisphère nord de la planète. Au Canada, la forêt boréale (270 millions d‟hectares) constitue 28% de la zone boréale mondiale et elle joue un rôle écologique important pour le stockage du carbone, la purification de l‟air et la régularisation du climat (RNCan 2017). De plus, l‟exploitation des forêts boréales génère environ 60% de l‟activité économique de l‟industrie forestière canadienne (Burton et al. 2003). Le bois d‟œuvre, les produits forestiers non ligneux et le potentiel hydroélectrique des rivières constituent des sources de matières premières renouvelables (RNCan 2015). L‟aménagement forestier est une perturbation humaine omniprésente dont les effets ont été documentés dans la forêt boréale Canadienne. Des superficies appréciables de la forêt boréale sont coupées chaque année mais elles ne représentent que 0,2% du territoire forestier du Canada (RNCan 2017). Les pratiques d‟aménagement changent continuellement en réponse aux changements de politiques, aux conditions du marché, aux avancées technologiques en équipement pour la récolte, aux changements de régimes forestiers et à l‟acquisition de nouvelles connaissances (Venier 2014). L‟aménagement écosystémique et l‟aménagement durable des forêts sont deux concepts récents qui se sont développés en réponse aux débats concernant la gestion environnementale des forêts (Grenon et al. 2010). L‟aménagement écosystémique est une approche qui oriente les pratiques forestières pour que les critères environnementaux, économiques et sociaux de l‟aménagement durable soient respectés (Grenon et al. 2010). Sa mise en œuvre vise à assurer le maintien de la biodiversité et la viabilité des écosystèmes en réduisant les écarts entre la forêt aménagée et la forêt jugée naturelle, dans le respect des valeurs sociales liées au milieu forestier et donc en répondant aux besoins socio-économiques de la population (Grenon et al. 2010).
Maintenir les processus écologiques en s‟inspirant des perturbations naturelles fait partie des nouvelles approches d‟aménagement dans l‟écosystème boréal ayant comme objectif de conserver la biodiversité (Fischer et al. 2006, Mori 2011). Cette dernière est indéniablement affectée par les perturbations, naturelles ou anthropiques, et ce à de multiples échelles temporelles et spatiales (Pickett et White 1985, Levin 2000, Loreau 2010, Turner 2010). Les feux de forêt sont considérés comme la perturbation la plus importante pour renouveler la forêt boréale dans les zones biogéoclimatiques centrales et de l‟ouest (Rowe et Scotter 1973, Bergeron et al. 2006). Dans les régions plus humides de
2
l‟Est du Canada, où le cycle de feux est plus long, les insectes ravageurs sont les agents prédominants du régime de perturbations naturelles (Venier 2014). Au Québec, la dynamique de la pessière est principalement régie par le feu (Payette 1992, Bergeron et
al. 2001, Bond-Lamberty et al. 2007), alors que celle de la sapinière l‟est surtout par les
épidémies de tordeuse des bourgeons de l‟épinette (TBE; Choristoneura fumiferana Clemens) (Lepidoptera: Tortricidae) (Blais 1983). Connaître l‟impact de ces perturbations naturelles sur la biodiversité des écosystèmes forestiers est primordial dans le contexte où on veut s‟en inspirer à des fins d‟aménagement. Indépendamment de la perturbation, elle provoque généralement une quantité considérable d‟arbres stressés et morts qui deviennent une ressource privilégiée pour la faune saproxylique (Speight 1989). Les différentes perturbations forestières affectent alors négativement ou positivement les multiples espèces végétales et animales présentes dans l‟écosystème.
Successions végétales et animales après perturbations naturelles
D‟un point de vue socio-économique, les feux en forêt boréale sont perçus comme des désastres naturels (Drapeau et al. 2010). Pourtant, au plan écologique, ils engendrent des habitats prospères pour certaines communautés végétales et animales (Drapeau et al. 2010, Nappi et al. 2011). Le nouvel écosystème produit suite au passage du feu possède plusieurs attributs uniques qui sont favorables à des espèces opportunistes (Wikars 1997, Boulanger et Sirois 2007) mais également à plusieurs espèces «associées aux feux» (Boucher et al. 2012). Les essences ligneuses retrouvées dans les régions à haute fréquence de feux ont évolué et se sont adaptées pour devenir plus résistantes et/ou résilientes à cette perturbation (Pausas et Keeley 2009). Des traits acquis tels qu‟une écorce épaisse, une discontinuité verticale protégeant les cônes ou la présence de cônes sérotineux peuvent avantager certaines espèces dans le processus de succession végétale après le passage du feu (Deslaurier et al. 1996, Mauri Ortuno et al. 2009). Par ailleurs, aux suites de sévères épidémies de TBE, l‟ouverture des peuplements défoliés peut entraîner un renouvellement complet du peuplement grâce à une régénération préétablie (Bouchard et al. 2005). Par exemple, l‟abondance du sapin baumier (Abiesbalsamea (L.) Mill.) dans le sous-bois sous forme de graines, de semis et de régénération
avancée lui permet un retour rapide suite à ce type de perturbation. D‟autres espèces de lumière, dites pionnières, comme le bouleau blanc (Betula papyrifera Marshall), ont besoin d‟une ouverture de la voûte forestière pour se régénérer efficacement. Des épidémies
3
sévères et répétées de TBE leur procurent des conditions optimales en peuplements mixtes et dominés par les conifères (Bouchard et al. 2007). Les espèces végétales montrent des patrons de succession adaptés aux différentes perturbations forestières et on peut anticiper qu‟il en soit de même pour les insectes, dont les espèces saproxyliques sont les premières à profiter de l‟une des principales caractéristiques de ces perturbations naturelles, soit la production de bois mort en abondance.
Les organismes saproxyliques sont définis comme ceux qui utilisent, pendant au moins une partie de leur cycle vital, du bois moribond ou mort, ou d‟autres organismes saproxyliques présents dans le bois comme les champignons et les bactéries (Speight 1989). Les coléoptères jouent un rôle considérable dans la colonisation du bois, contribuant à la décomposition et au recyclage des nutriments (Speight 1989, Kim 1993, Boulanger et Sirois 2007), en plus de faire partie de l‟alimentation de base de plusieurs vertébrés (Morissette et al. 2002, Kennedy et Fontaine 2009, Nappi et al. 2010). Ainsi, dans le processus d‟utilisation du bois mort issu d‟une perturbation forestière, la première phase est celle de la colonisation primaire des tissus sous-corticaux et ligneux. Cette colonisation est généralement effectuée par les coléoptères. De plus, la communauté qui amorce la décomposition est directement associée au type de bois mort car le niveau de spécialisation de ces colonisateurs primaires est souvent très prononcé (Vindstad et al. 2014). Par conséquent, un coléoptère saproxylique peut normalement exploiter du bois mort à un stade spécifique de décomposition (Vindstad et al. 2014), et la spécialisation peut aller jusqu‟ au niveau de l‟espèce ou même au diamètre de l‟hôte (Lachat et al. 2013). De plus, les caractéristiques du bois mort varient selon le type et l‟intensité de la perturbation (Pedlar et al. 2002). Alors que les feux de surface n‟affectent que le sous-bois forestier, les feux de couronnes tuent la majorité des arbres et peuvent brûler l‟humus jusqu‟au sol minéral (Gauthier et al. 2001). En plus de brûler les arbres, les feux exposent davantage les tiges au soleil (Jonsell et al. 1998), résultant en des tissus sous-corticaux plus secs (Boulanger et Sirois 2006). D‟un autre côté, en contexte d‟épidémie sévère de TBE, la mortalité des arbres commence après quatre à cinq années de défoliation grave, mais cette mortalité s‟échelonne sur plusieurs années (Taylor et MacLean 2009). Au Canada, plusieurs études ont porté sur l‟impact des différentes perturbations naturelles sur les communautés végétales, ce qui n‟est pas le cas pour les coléoptères saproxyliques. Seul l‟impact des feux de forêt sur les coléoptères saproxyliques, en pessière noire à sphaignes (Saint-Germain et al. 2004a et b, Boulanger et Sirois 2007, Boulanger et al.
4
2010, Boucher et al. 2012), a retenu l‟attention, mais très peu d‟études ont porté sur l‟impact des épidémies d‟insectes comme la TBE sur ces mêmes coléoptères (Belyea 1952a et b). En 2012, une épidémie d‟arpenteuse de la pruche (Lambdina fiscellaria (Guenée)) (Lepidoptera: Geometridae), le 2e défoliateur en importance dans l‟Est de l‟Amérique du Nord (Hébert et Brodeur 2013) a été observée pour la première fois dans la Région de la Capitale-Nationale du Québec. Cette nouvelle perturbation naturelle majeure offre une occasion unique d‟étudier son impact sur la communauté des coléoptères saproxyliques.
Une première épidémie d’arpenteuse de la pruche dans la Région
de la Capitale-Nationale
Native de l‟Amérique du Nord, l‟arpenteuse de la pruche cause des pertes de croissance mais surtout une importante mortalité des arbres attaqués sévèrement dans les peuplements de sapins baumiers du domaine bioclimatique de la sapinière à bouleau blanc (Otvos et al. 1971). À Terre-Neuve, de nombreuses infestations d‟arpenteuse de la pruche ont été observées au cours du 20e siècle (Carroll 1956, Iqbal et al. 2011). Au Québec, plusieurs épidémies ont sévi à l‟Île d‟Anticosti (Jobin et Desaulniers 1981) pendant la même période, mais plus récemment, l‟arpenteuse a défolié 924 971 ha de forêt sur la Côte-Nord en 2000 (Bordeleau 2000). Des dommages ont encore été recensés en 2015 sur Anticosti (MFFP 2015). En 2012, une première année de dommages causés par l‟arpenteuse de la pruche a été signalée dans la région de la Capitale-Nationale, notamment à la forêt Montmorency, dans le Parc national de la Jacques -Cartier ainsi que dans la réserve faunique des Laurentides (3379 ha; MRNF 2012). L‟étendue des premiers dommages causés par cette perturbation a alors été évaluée par survol aérien de la région dans le secteur de la Capitale-Nationale à l‟aide d‟un logiciel de «Relevé Aérien à Référence Spatiale» ou RARS (MRNF 2012).
L‟arpenteuse de la pruche est une espèce univoltine qui passe l‟hiver sous forme d‟œuf (Carroll 1956). L‟éclosion se produit entre la fin de mai et le début de juin, après le débourrement des bourgeons du sapin baumier (Hébert et Jobin 2001, Hébert et al. 2001; 2004). Les larves des deux premiers stades consomment les aiguilles des pousses annuelles des conifères puis, au milieu de leur développement, les larves se déplacent sur le vieux feuillage (Hébert et Jobin 2001, Iqbal et Maclean 2010). Elles se développent
5
en passant par quatre ou cinq stades larvaires durant l‟été (Berthiaume et al. 2007). Indépendamment de l‟âge larvaire, l‟arpenteuse consomme partiellement les aiguilles des conifères (Carroll 1956). Ce comportement alimentaire particulier est très dommageable car toutes les aiguilles ayant été partiellement consommées sèchent et finissent par tomber à l‟automne (Hébert et Jobin 2001). Cela peut entrainer une mortalité rapide des arbres défoliés, et ce, dès la première année où les dommages sont observés (Hudak et
al. 1978, Jobin et Desaulniers 1981). Son hôte de prédilection est le sapin baumier mais
elle peut aussi entraîner la mort d‟autres Pinaceae comme la pruche du Canada (Tsuga
canadensis (L.) Carrière) (Trial 1993) et les épinettes blanche et noire (MacLean and
Ebert 1999). Vers la fin de juillet, les larves âgées montrent une phototaxie négative (Carroll 1956) et cherchent un endroit (lichen, mousse ou crevasse dans l‟écorce) pour s‟accrocher et former leur chrysalide. Après environ 20 jours de développement, les adultes émergent des chrysalides, s‟accouplent et pondent leurs œufs (Carroll 1956, Dobesberger 1989). Les infestations débutent habituellement sur de petites surfaces, isolées et bien définies, puis elles forment des îlots rapprochés ou une grande infestation irrégulière. Les épidémies d‟arpenteuse demeurent essentiellement localisées (Carroll 1956) comparativement à celles de la TBE. Conséquemment, une fois que tous les arbres d‟un peuplement sont morts, les larves n‟ont presque plus de ressources alimentaires car les capacités de déplacement de l‟espèce se limitent au « ballooning » des premiers stades larvaires et au vol des papillons femelles, qui s‟exprime seulement après avoir déposé au moins 50% de ses oeufs (Delisle et al. 1998). Les épidémies apparaissent et disparaissent donc soudainement, en persistant rarement plus que trois ans dans un même peuplement (Maclean et Ebert 1999, Hébert et al. 2001, Bordeleau 2002).
Les mortalités causées par l‟arpenteuse de la pruche se situent temporellement entre celles causées par les feux de forêt, qui tuent instantanément la majorité des arbres affectés, et la TBE, qui nécessite plusieurs années de défoliation pour entraîner la mortalité des arbres. La défoliation grave de sapins baumiers par l‟arpenteuse de la pruche cause un stress violent qui peut parfois entraîner la mort des arbres en aussi peu que trois mois. En conséquence, cette ouverture de la voûte forestière augmente l‟entrée de lumière, entraîne une hausse de la température et facilite l‟entrée du vent rendant les tiges mortes susceptibles aux chablis (Ostaff et MacLean 1989). De plus, la décomposition et le recyclage des nutriments contenus dans la biomasse végétale morte devraient alors être déclenchés par les coléoptères phloéophages et xylophages. Ces espèces
6
consomment respectivement les tissus sous-corticaux et les tissus de l‟aubier et du duramen d‟arbres stressés ou récemment morts (Langor et al. 2008). Plusieurs espèces de coléoptères appartenant à la famille des Buprestidae, Cerambycidae et Curculionidae sont parmi les premiers colonisateurs des arbres (Langor et al. 2008) et sont donc les portes d‟entrée pour d‟autres espèces saproxyliques de succession et de décomposition plus avancées. Le rôle écologique de ces colonisateurs primaires est donc important pour l‟ensemble des organismes saproxyliques mais également pour assurer la décomposition des tiges et la remise en circulation d‟éléments nutritifs dans l‟écosystème qui pourraient rester séquestrés dans les chicots pour une longue période.
La coévolution des coléoptères phloéophages et xylophages avec
leurs hôtes
Les coléoptères phloéophages et xylophages utilisent souvent des stratégies alimentaires de colonisation non-agressives car ils s‟attaquent majoritairement aux arbres récemment morts, mais ils peuvent aussi utiliser des stratégies agressives s‟ils s‟attaquent à des arbres vivants. Les espèces très agressives tuent les arbres parfois sains en les colonisant massivement; ils appartiennent à la classe des parasites obligatoires (Raffa et al. 1993). Les espèces qualifiées de parasites facultatifs sont plus ou moins agressives et colonisent normalement des arbres tombés, stressés, blessés ou sous attaque d‟autres insectes ou pathogènes. Ils s‟attaquent seulement aux arbres sains lorsque leurs populations ont suffisamment augmenté grâce à une grande disponibilité d‟arbres affaiblis (Raffa et al. 1993). Enfin, les espèces non-agressives qui colonisent des arbres récemment morts sont dites herbivores saprophytes (Raffa et al. 1993). Les différentes stratégies fournissent des avantages ou désavantages pour la reproduction selon la qualité nutritionnelle des tissus consommés ainsi qu‟en fonction des mécanismes de défense naturelle des arbres. Il a été suggéré que les colonisateurs primaires sont surtout spécialistes dans la sélection de l‟espèce d‟arbre colonisée, tandis que les colonisateurs secondaires (fongivores, détritivores et prédateurs) sont plus généralistes (Langor 2008).
Les conifères et les scolytes (Curculionidae: Scolytinae) coexistent depuis le Mésozoïque précoce. Une relation étroite s‟est développée entre ces insectes et les membres de la famille des Pinaceae qui produisent de la résine de manière constitutive (Seybold et al. 2000). La coévolution entre ces espèces implique une course aux armements entre les
7
colonisateurs et les colonisés (Franceschi et al. 2005). Certaines espèces de conifères, comme le sapin baumier, possèdent un système de défenses naturelles contre les pathogènes et les colonisateurs de tige qui se divise en une partie constitutive et une partie induite (Franceschi et al. 2005). Le système constitutif inclut des cellules et canaux qui accumulent de la résine dans les tissus vasculaires, des cellules dans le phloème qui contiennent des composés chimiques toxiques (e.g. phénols et terpénoïdes), et des propriétés mécaniques de l‟écorce comme des couches de cellules subérisées et lignifiées, des scléréides et des cristaux d‟oxalate de calcium (Franceschi et al. 2005). Le système induit implique une synthèse de novo ou une activation d‟une grande gamme de composés chimiques de défense comme des protéines liées à la pathogenèse et des enzymes en plus d‟autres phénols et terpénoïdes. Ce système induit agit parfois contre une infection en cours (réponse hypersensible et résistance locale) ou contre des infections/agressions futures de xylophages (résistance acquise) (Franceschi et al. 2005). Plusieurs études ont montré, par exemple, que la production de phénols et la régulation d‟enzymes impliquées dans la défense sont rapidement induites dans des arbres précédemment colonisés et que les composés phénoliques sont plus toxiques et plus spécifiques à l‟organisme invasif que ceux qui sont constitutifs (Franceschi et al. 2005). Grâce à ce système complexe de défenses naturelles, les conifères sont protégés contre une grande quantité d‟organismes tentant de les attaquer, mais certains colonisateurs primaires ont aussi évolué en acquérant des stratégies permettant de les outrepasser ou de les vaincre (Franceschi et al. 2005). Le processus de colonisation des arbres par les coléoptères xylophages est spécifique à l‟organisme, mais certaines espèces partagent des stratégies dans leur cycle vital. Afin d‟en faire ressortir les traits avantageux, il est convenable de le diviser en trois phases: la dispersion, la colonisation et la reproduction (Stark 1982).
Généralement, une fois le développement des larves complété, l‟adulte émerge de l‟arbre et entre en phase de dispersion afin de sélectionner un hôte potentiel pour la colonisation. La décision d‟accepter ou non un hôte dépend de signaux olfactifs, visuels, mécaniques et gustatifs, inhibiteurs et stimulants, balancés avec des signaux excitatifs et inhibiteurs internes (Miller et Strickler 1984). Il a été suggéré que certaines espèces ont développé de fortes réponses comportementales lors de la détection de composés volatils émis par l‟espèce hôte indiquant un substrat potentiel pour se reproduire (Byers 1995). Les coléoptères non-agressifs seraient fortement attirés par des monoterpènes, qui sont
8
ironiquement des composés chimiques impliqués dans le système de défense des conifères, et par l‟éthanol produit par le métabolisme anaérobique d‟organismes présents dans les plantes stressées et en décomposition (Byers 1995). Cette attraction primaire, résultant de la détection de produits émis par l‟hôte, démontre que les défenses naturelles des arbres peuvent devenir une faiblesse (Franceschi et al. 2005). Par ailleurs, les espèces agressives possèdent majoritairement des phéromones d‟agrégation qui jouent le rôle d‟attraction secondaire. La plupart des phéromones de scolytes sont synthétisés à partir des monoterpènes (Vanderwel et Oehlschlager 1987) ou par biosynthèse de novo (Ivarsson et al. 1993, Seybold et al. 1995, Seybold et Tittiger 2003), mais certaines espèces non-agressives ont aussi développé ce système qui entre en synergie avec les kairomones issues de l‟arbre. Les défenses naturelles des arbres peuvent alors devenir un avantage pour les saproxyliques primaires qui ont développé cette stratégie durant cette course aux armements.
Une fois l‟hôte choisi, les colonisateurs font face aux défenses naturelles de l‟arbre. Ils doivent forer l‟écorce puis passer à travers les défenses constitutives et ceux qui s‟attaquent aux arbres vivants doivent aussi contrer les défenses induites. Deux issues sont possibles lors de la colonisation d‟arbres vivants: soit l‟arbre se défend efficacement et les coléoptères sont expulsés ou tués, soit l‟arbre est colonisé et meurt (tué par annelage ou par infection) (Berryman et al. 1989). Les scolytes sont les plus grands ravageurs des forêts tempérées de conifères (Franceschi et al. 2005). Trois facteurs plus ou moins complémentaires et impliqués dans la colonisation sont souvent mentionnés comme les plus importants pour expliquer ce phénomène (Franceschi et al. 2005). Premièrement, les comportements d‟agrégation sont utiles pour trouver des hôtes potentiels et des partenaires sexuels mais ils sont aussi décisifs dans le contexte de colonisation. La colonisation massive coordonnée par les phéromones d‟agrégation serait essentielle pour épuiser le système de défense de l‟arbre (Franceschi et al. 2005), car les scolytes ne semblent pas être généralement capables d‟inhiber les défenses induites (Berryman et al. 1989). Deuxièmement, l‟évolution d‟une tolérance envers les mécanismes
de défenses de l‟hôte est importante, par exemple pour les pionniers émettant les phéromones d‟agrégation ou lorsque les populations sont basses et qu‟une attaque massive est impossible. Troisièmement, une des stratégies qui n‟a pas été encore discutée et qui semble être décisive pour la colonisation est l‟association de certaines espèces de coléoptères xylophages, notamment les scolytes, avec des symbiontes
9
fongiques. Cette association commune peut être mutualiste, donc avec de clairs bénéfices pour les deux partenaires comme pour les scolytes «à ambrosia» et leur symbionte. Cependant, l‟association peut aussi être antagoniste (c.-à-d. compétiteurs pour la ressource ou pathogène du coléoptère), commensale ou occasionnelle, ou à effets inconnus (Francke-Grosmann 1967, Beaver et al. 1989, Franceschi 2005). L‟intensité de la relation entre les scolytes et leurs symbiontes varie de facultative à obligatoire, mais elle peut être responsable de spectaculaires épidémies, lorsque favorisée par des conditions climatiques spécifiques, la pathogénicité du symbionte ou le comportement d‟agrégation du scolyte (You et al. 2015). L‟association entre les scolytes et les symbiontes infestant les conifères est complexe. Ainsi, distinguer l‟espèce du symbionte et la manière dont ce dernier est transporté est pertinent dans la détermination de la nature de leur relation (Paine et al. 1997). Généralement, le symbionte est composé de champignons basidiomycètes et d‟ascomycètes transportés sous forme de spores ou de mycélium à l‟intérieur d‟invaginations cuticulaires, les mycanges (Paine et al. 1997). Des espèces pathogènes de champignon de bleuissement, généralement des ascomycètes, sont responsables ou requises pour entraîner la mortalité des arbres attaqués lorsque les espèces de scolytes agressives en transportent (Paine et al. 1997). Une fois à l‟intérieur des tissus sous-corticaux, le champignon transporté progresse dans les trachéides à travers les ponctuations aréolées ou en pénétrant directement les parois cellulaires (Gibbs 1993, Wallin et Raffa 2001). Ensemble, certaines espèces de coléoptères et de champignons interrompent le système vasculaire de l‟arbre rendant le xylème non-conducteur, empêchant donc la circulation d‟eau et de substances nutritives dans l‟aubier, causant ultimement la mort de l‟arbre (Wallin et Raffa 2001).
Enfin, une fois la colonisation effectuée, la phase de reproduction débute. Chez plusieurs Curculionidae, cette phase inclut l‟accouplement, la construction de galeries, l‟oviposition et le développement de la progéniture. Les adultes creusent une galerie principale où la femelle déposera ses œufs. Les galeries ont souvent des formes uniques à l‟espèce à un point tel qu‟elles sont parfois utilisées pour identifier les espèces ayant colonisé les arbres (Stark 1982). Lorsque les œufs éclosent, les larves creusent majoritairement des galeries individuelles qui s‟allongent et croissent en volume tout le long du développement. Lorsque la phase d‟alimentation se termine, la pupaison se produit dans une cellule nymphale. Les autres Curculionidae, les Cerambycidae et les Buprestidae pondent majoritairement leurs œufs dans de petites crevasses ou cavités. Les larves effectuent
10
donc la colonisation dans ce dernier cas et une seule galerie, souvent plus grosse, est observée dans les tissus sous-corticaux et parfois dans l‟aubier. Le développement des larves et des pupes à l‟intérieur des arbres constitue la plus grande partie du cycle vital des coléoptères phloéophages et xylophages. Cela peut durer de quelques semaines à quelques années.
Les comportements de détection d‟hôtes potentiels et d‟agrégation, ainsi que les mécanismes de tolérance aux défenses de l‟arbre sont évolutivement avantageux et expliquent pourquoi les scolytes agressifs sont les agents les plus destructeurs de la forêt boréale tempérée (Franceschi et al. 2005). Cependant, le comportement alimentaire et l‟association avec des symbiontes sont d‟importants facteurs dans l‟évaluation des conséquences économiques reliées à la colonisation. Les phloéophages primaires sont des pionniers de la colonisation et se spécialisent dans la consommation du contenu des cellules du cambium, du phloème primaire et du xylème primaire, donc des tissus sous-corticaux en général situés juste sous l‟écorce (Speigth 1989). Plusieurs espèces de xylophages foreurs primaires possèdent l‟habileté de digérer la cellulose, grâce à des symbiontes ou des (hémi)-cellulases ainsi que des mandibules assez fortes pour forer des tunnels à travers l‟écorce et le bois (Speigth 1989). La mortalité de grandes quantités d‟arbres dans un peuplement cause des pertes importantes de profit reliées à une réduction de rentabilité par rapport à sa productivité potentielle. Des pertes économiques sont aussi entraînées car le bois colonisé est souvent coloré et certaines espèces comme le longicorne noir (Monochamus scutellatus (Say)) (Coleoptera: Cerambycidae) peuvent coloniser des arbres brûlés avec une densité excédant 300 larves par m² de surface de tige creusant chacune des galeries pouvant atteindre 20 cm de long et pénétrant le bois de 7.5 cm en moyenne (Safranyik and Raske 1970, Cerezke 1977). Ces tiges sont toujours déclassées pour des causes d‟esthétisme (Lowell et al. 1992). De plus, l‟exportation de ce bois peut être l‟objet de restrictions pour éviter le transfert potentiel de vecteurs de maladies comme le nématode du pin (Saint-Germain et Greene 2009). Les coléoptères phloéo- et xylophages sont pris au sérieux et peuvent être considérés comme nuisibles par les organismes de réglementation associés aux opérations forestières. Les enjeux économiques reliés à la colonisation des arbres par ces coléoptères sont réels. Ainsi, dans une optique de bonification de l‟aménagement écosystémique et de l‟optimisation de la ressource forestière, l‟étude du processus de colonisation des arbres
11
par les coléoptères xylophages suite à une nouvelle perturbation naturelle, au Québec, est pertinente afin d‟en caractériser les impacts écologiques et économiques.
La colonisation par les coléoptères phloéophages et xylophages
en contexte d’épidémie d’arpenteuse de la pruche
Les défenses induites par l‟arbre dans les tissus sous-corticaux sont influencées non seulement par les nutriments emmagasinés localement mais aussi par la translocation de nouveaux photosynthétats issus des aiguilles (Fransceschi et al. 2005). Les assimilats sont alors utilisés par les structures de défense ainsi que pour synthétiser les composés chimiques et ne sont alors pas utilisés pour supporter la croissance et la reproduction, impliquant donc l‟importance pour l‟arbre de bien équilibrer l‟allocation de ses ressources (Fransceschi et al. 2005). Des facteurs abiotiques variables, tels que le stress hydrique, la pollution de l‟air, la température en plus de l‟attaque par des agents biotiques, comme l‟arpenteuse de la pruche, peut altérer la ressource en assimilats (Fransceschi et al.
2005). Wallin et Raffa (1999) ont montré que la défoliation du pin gris (Pinus banksiana Lamb.), par la tordeuse du pin gris (Choristoneura pinus pinus Freeman) (Lepidoptera: Tortricidae), affecte le contenu et la composition de monoterpènes constitutifs et induits en plus d‟affecter des réponses physiologiques comme la réduction de croissance, la probabilité de mortalité, le taux de confinement fongique (du symbionte de scolytes) et le volume de résine. Ils ont montré que la défoliation entrainait plus de changements dans les tissus du phloème que dans le feuillage directement affecté par l‟insecte ravageur. Ils ont aussi déterminé que la durée du stress (défoliation) était importante et entrait en interaction avec son intensité ainsi qu‟avec le profil de défenses naturelles. Dans un contexte semblable, Iqbal et al. (2011) ont montré que l‟intensité de la défoliation par l‟arpenteuse de la pruche était déterminante dans la diminution des taux de croissance et de mortalité des sapins baumiers et des épinettes noires affectés. De plus, Caron et al. (2013) ont rapporté l‟augmentation de monoterpènes dans les tissus foliaires de sapins baumiers défoliés par la TBE. Il est donc probable que les profils de défense naturelle et de qualité nutritive des sapins baumiers, affectés par l‟épidémie d‟arpenteuse de la pruche de 2012, aient été modifiés. Des espèces de coléoptères xylophages primaires pourraient alors détecter les arbres défoliés et vulnérables, par signal olfactif majoritairement, comme une opportunité à la colonisation.
12
Les travaux de Bowers et al. (1996) ont porté sur la défoliation causée par la TBE et la présence du polygraphe de l‟épinette (Polygraphus rufipennis (Kirby)) (Coleoptera: Curculionidae) dans des peuplements d‟épinette noire (Picea mariana (Mill.) B.S.P.) à Terre-Neuve. Ils ont rapporté l‟augmentation de l‟incidence du coléoptère dans les peuplements sévèrement défoliés, particulièrement dans les arbres morts, mais ils n‟ont pas pu préciser le niveau de défoliation requis pour déclencher la colonisation, car ils ont écorcé les arbres deux ans après la fin de l‟épidémie de TBE. W allin et Raffa (2001) ont étudié l‟impact de la défoliation de la tordeuse du pin gris sur la densité de trous d‟entrée du scolyte à grand corselet (Ips grandicollis Eichoff) (Coleoptera: Curculionidae), ainsi que du longicorne de la Caroline (Monochamus carolinensis (Olivier)) (Coleoptera: Cerambycidae) au Wisconsin. Ils ont montré que la colonisation par ces coléoptères augmentait exponentiellement en relation avec le niveau de défoliation des pins gris . Bien que les résultats soient très intéressants, les auteurs ont utilisé des classes de défoliation comme variable explicative et des décomptes de trous d‟entrée comme variable réponse de la même manière que Bowers et al. (1996). Afin de déterminer un impact plus précis de la défoliation sur la communauté de coléoptères phloéo- et xylophages, et subséquemment, sur le processus de colonisation des arbres post-épidémie d‟arpenteuse de la pruche, il serait pertinent d‟étudier l‟effet d‟un niveau plus précis de défoliation des arbres en temps réel sur les assemblages d‟espèces et les populations de ces coléoptères. La présence, l‟abondance et la succession de ces coléoptères témoigne de leur sensibilité élevée aux changements dans l‟écosystème forestier et pourraient faire office de bioindicateurs comme objectif à atteindre en contexte de simulation de perturbation naturelle. La conservation de telles espèces et donc des rôles qu‟elles jouent suite à l‟épidémie s‟inscrit dans l‟optique de respect et de la préservation des valeurs écologiques véhiculées par l‟aménagement écosystémique. D‟un autre point de vue, la caractérisation et la prédiction d‟une trajectoire successionnelle d‟espèces économiquement importantes en fonction de la défoliation pourrait s‟avérer utile à des fins de détection et de prévention de dommages entraînant des pertes économiques. Les informations issues des résultats de la présente étude serviront à produire des recommandations afin de bonifier l‟aménagement écosystémique. L‟objectif principal est, par conséquent, de caractériser l‟impact de la défoliation par l‟arpenteuse de la pruche sur la communauté de coléoptères phloéo- et xylophages. Plus spécifiquement, les objectifs sont les suivants:
13
Objectif 1: Évaluer la justesse de différentes méthodes d‟estimation de la défoliation annuelle et cumulative afin d‟évaluer l‟impact de l‟arpenteuse de la pruche sur les sapins baumiers et sur les coléoptères phloéo- et xylophages.
Objectif 2: Déterminer si une succession à court terme survient chez les coléoptères phloéo- et xylophages en présence de défoliation et déterminer quelles sont les principales variables environnementales expliquant la ou les possibles trajectoires successionnelles.
Objectif 3: Déterminer un taux de défoliation seuil au-delà duquel survient la colonisation massive des coléoptères phloéo- et xylophages et déterminer si les pièges utilisés pourraient servir d‟outil de détection dans la prédiction des attaques de coléoptères.
15
Chapitre 1
Comparison of several methods for estimating defoliation
from the hemlock looper (Lambdina fiscellaria (Guenée))
(Lepidoptera: Geometridae) on balsam fir
Jean-Michel Béland
1, Christian Hébert
2, Conrad Cloutier
3,
Richard Berthiaume
1and Éric Bauce
11
Université Laval, Faculté de foresterie, de géographie et de géomatique, Pavillon Abitibi- Price, Québec, Québec, G1K 7P4, Canada
2
Natural Resources Canada, Canadian Forest Service, Laurentian Forestry Centre, 1055 du P.E.P.S., P.O. Box 10380, Stn. Sainte-Foy, Québec, Québec, G1V 4C7, Canada 3
Université Laval, Faculté des sciences et de génie, Pavillon Alexandre-Vachon, Québec, Québec, G1V 0A6, Canada
16
Résumé
Une épidémie d‟arpenteuse de la pruche a été observée, pour la première fois, dans la région de Québec en 2012. Cette épidémie a fourni l‟opportunité d‟évaluer la précision d‟une méthode d‟inventaire aérien de la défoliation, récemment développée par le Ministère des Forêts, de la Faune et des Parcs du Québec. De plus, afin de développer une méthode d‟estimation de la défoliation au niveau de l‟arbre, nous avons comparé deux méthodes basées sur des estimations visuelles faites à partir du sol, avec la méthode classique de Fettes (1950). L‟inventaire aérien a fourni des estimés justes de la défoliation après la première année d‟épidémie mais s‟est avéré inexact lors de la deuxième année alors que seuls les peuplements gravement défoliés ont été correctement classés . La défoliation cumulée sur toutes les classes d‟âge de feuillage amène un nouveau défi aux équipes affectées aux programmes d‟évaluation de la défoliation par voie aérienne. La défoliation de la pousse annuelle était fortement corrélée avec l‟estimé de Fettes. Néanmoins, la défoliation estimée dans la couronne supérieure des arbres s‟est avérée le substitut le mieux corrélé avec la défoliation totale estimée par la méthode de Fettes.
Mots-clés: Arpenteuse de la pruche; Défoliation; Inventaire aérien; Méthode d‟estimation; Couronne supérieure
17
Abstract
An outbreak of the hemlock looper was observed in the Québec region for the first time in 2012. This outbreak provided the opportunity to test the efficacy of a newaerial defoliation survey method, developed by the Ministère des Forêts, de la Faune et des Parcs du Québec. In order to estimate defoliation at the tree level, we tested two proxies, which were easy and rapid to use from the ground level, and compared them with the classical Fettes method. First, we observed that aerial survey provided accurate estimates of defoliation severity during the first year of the outbreak but it was far less efficient after a second year of defoliation when only heavily defoliated stands were correctly classified. Cumulative defoliation on all age classes of foliage brings a new challenge to crews affected to aerial survey programs. Defoliation on current year shoots was strongly correlated with Fettes estimates combining all age classes. However, the overall defoliation estimated in the upper crown was more strongly correlated with Fettes estimates than defoliation on current year shoots.
Keywords: Hemlock Looper; Defoliation; Aerial survey; Estimation method; Current-year shoot; Upper crown
18
1 Introduction
The hemlock looper, Lambdina fiscellaria (Guenée) (Lepidoptera: Geometridae), is considered as the second most important insect defoliator of balsam fir (Abies balsamea (L.) Mill.) in eastern North America after the spruce budworm, Choristoneura fumiferana (Clemens) (Lepidoptera: Tortricidae) (Hébert and Brodeur 2013). Except when populations are extremely high, which may induce larval back-feeding on the old foliage, the spruce budworm usually feeds only on current-year shoots and thus kills balsam firs only after 4 to 5 years of severe defoliation (Blais 1958, MacLean 1980). In comparison, hemlock looper larvae chew on fir needles of all age classes without consuming them completely (Hébert and Jobin 2001, Iqbal and MacLean 2010). This wasteful feeding behaviour results in damaged needles eventually drying and falling in late summer and thus, the hemlock looper can kill trees within just one or two years of severe defoliation (Dobesberger 1989, Iqbal and MacLean 2010), which is much faster than for the spruce budworm.
Annual defoliation estimates are useful tools in forest pest management . They are usually obtained through aerial defoliation surveys used for assisting forest managers in their operational decisions (MacLean and McKinnon 1996) but they are also useful for research purposes (MacLean and Lidstone 1982, Gray et al. 2000, Candau and Fleming 2005). These estimates are based upon the characteristic reddish-brown coloration of tree foliage resulting from insect feeding (Iqbal and MacLean 2010). For the spruce budworm, over a 10-year period, MacLean and MacKinnon (1996) concluded that aerial sketch-mapping of defoliation was appropriate for management purposes. Globally, misclassifications resulted mostly from the difficulty in detecting and correctly estimating light defoliation with aerial survey. On the other hand, the heavy defoliation class of aerial survey slightly overestimated defoliation according to ground estimates (MacLean and MacKinnon 1996).
Estimating the annual defoliation by the hemlock looper is more challenging than for the spruce budworm as if affects all age classes of foliage but also because successive years of defoliation include previous years of defoliation (Iqbal and MacLean 2010). Moreover, methods used to produce defoliation estimates vary between jurisdictions and their accuracy also depends upon observers (Dorais and Kettela 1982). Data from Iqbal and MacLean (2010), using a shoot-count defoliation chart, applied in the laboratory on shoots from field-collected branches (e.g. Fettes 1950), showed that aerial surveys, carried out in
19
Newfoundland with helicopters, overestimated defoliation in both moderate and heavy defoliation areas (no data was available for lightly defoliated stands). In Quebec, aerial defoliation surveys are carried out using aircrafts by the Ministère des Forêts, de la Faune et des Parcs (MFFP; named Ministère des Ressources naturelles et de la Faune or MRNF in 2012). The MFFP has developed a spatially explicit mapping tool, which has been used since 2003, but its accuracy for assessing hemlock looper defoliation has not yet been appraised. Moreover, at ground level, it is often impractical to use a shoot-count method, like the Fettes method, due to time and resource limitations. Easier and faster field estimation methods are thus needed for research purposes. For instance, we may hypothesize that, even if the hemlock looper feeds on multiple age classes of foliage, current-year shoot defoliation could be a good proxy of the overall tree defoliation. For a conifer-feeding insect such as the hemlock looper, current-year shoots are the only ones on which defoliation can be categorized with certainty as annual. Also, an estimation of defoliation in the upper crown could be a good proxy of the overall tree defoliation for research purposes that need defoliation estimates at the tree level.
In this paper, our objectives were 1) to verify the accuracy of aerial surveys carried out in Quebec for assessing hemlock looper annual defoliation at the stand level, over the first two years of an outbreak, and 2) to compare two ground-level methods for assessing defoliation at the tree level.
20
2 Methods
2.1 Study area
The study was realized in the Parc national de la Jacques-Cartier (47°06' N, 71°20' W) and in the Forêt Montmorency (47°19' N, 71°09' W), both located approximately 70 km north of Quebec City. Both areas belong to the balsam fir-white birch bioclimatic domain, within the continuous boreal forest sub-zone (Saucier et al. 2011). Balsam fir stands largely dominate the landscape and black spruce (Picea mariana (Mill.) B.S.P.), white spruce (Picea glauca (Moench) Voss.), white birch (Betula papyrifera Marsh.), American mountain-ash (Sorbus americana Marsh.) and trembling aspen (Populus tremuloides Michx.) are the most common companion species in these stands.
2.2 Stand selection and defoliation estimation
Aerial surveys were conducted in August 2012 and 2013 by the MFFP to assess hemlock looper annual defoliation. Aircraft flew at 160 km/h and 240 m altitude along flight lines spaced by 4.5 km (MRNF 2012). Their spatially explicit mapping tool was used to collect and digitize images by delineating defoliated patches with a stylus on a touchscreen computer. To make it easier, a topographic map rolled out as a background on the screen, linked with the aircraft route. Images were directly digitized and georeferenced with a GIS software. Defoliated patches were classified according to three severity classes: light (1−34%), moderate (35−69%) and heavy (70−100%). Light defoliation is defined as a loss of foliage in the upper crown for some trees in a stand, moderate defoliation as a loss of foliage in the upper crown of most trees and on the full crown length of several trees, and heavy defoliation as a loss of foliage along the full crown length of most trees (MRNF 2012).
During spring 2013, 12 balsam fir stands were selected among four defoliation classes (trace, light, moderate and heavy) according to the 2012 MFFP aerial survey. Three 400 m² circular plots, spaced at least by 50 m, were established within each selected stand for estimating defoliation from ground-level. Four co-dominant balsam firs, labeled as «sample trees», were selected in each plot for estimating defoliation during this hemlock looper outbreak. Defoliation was visually estimated on sample trees using the chart developed for the spruce budworm by Fettes (1950). This method is usually applied in the laboratory on