Etude prospective des gènes de prédisposition aux anomalies du tube neural dans une population algér
124
0
0
Texte intégral
Figure

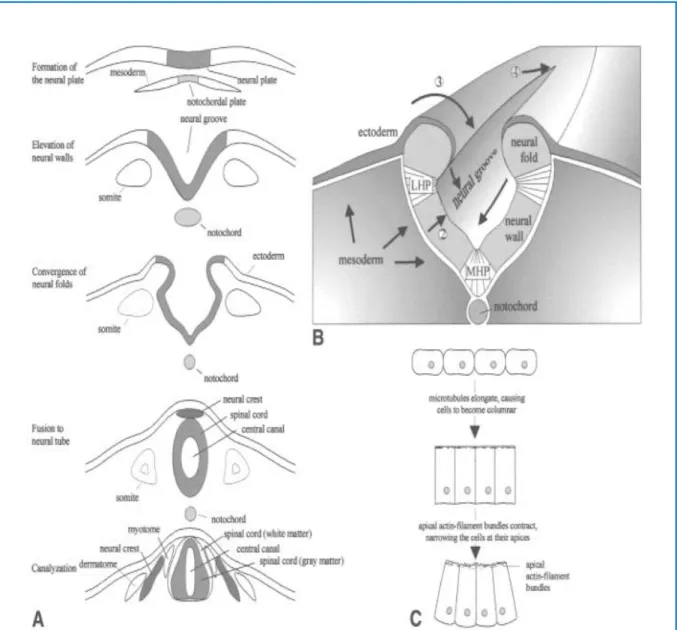
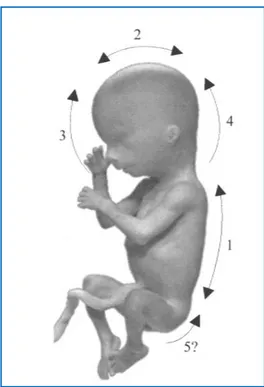
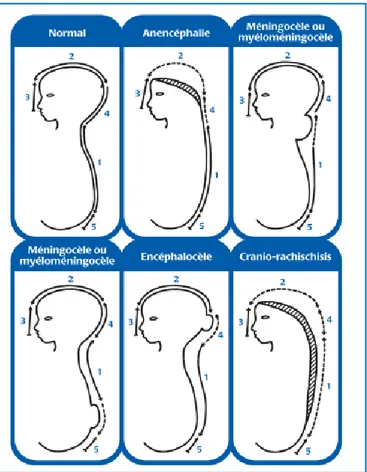
+7

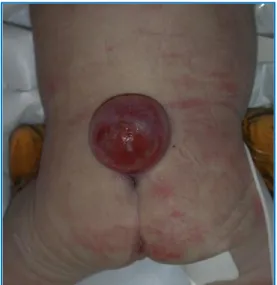




Documents relatifs